
Hydrothermal Processing and In Vitro Simulated Human Digestion Affects the Bioaccessibility and Bioactivity of Phenolic Compounds in African Pumpkin (Momordica balsamina) Leaves

 , , and
, , and
Abstract
:1. Introduction
2. Results and Discussion
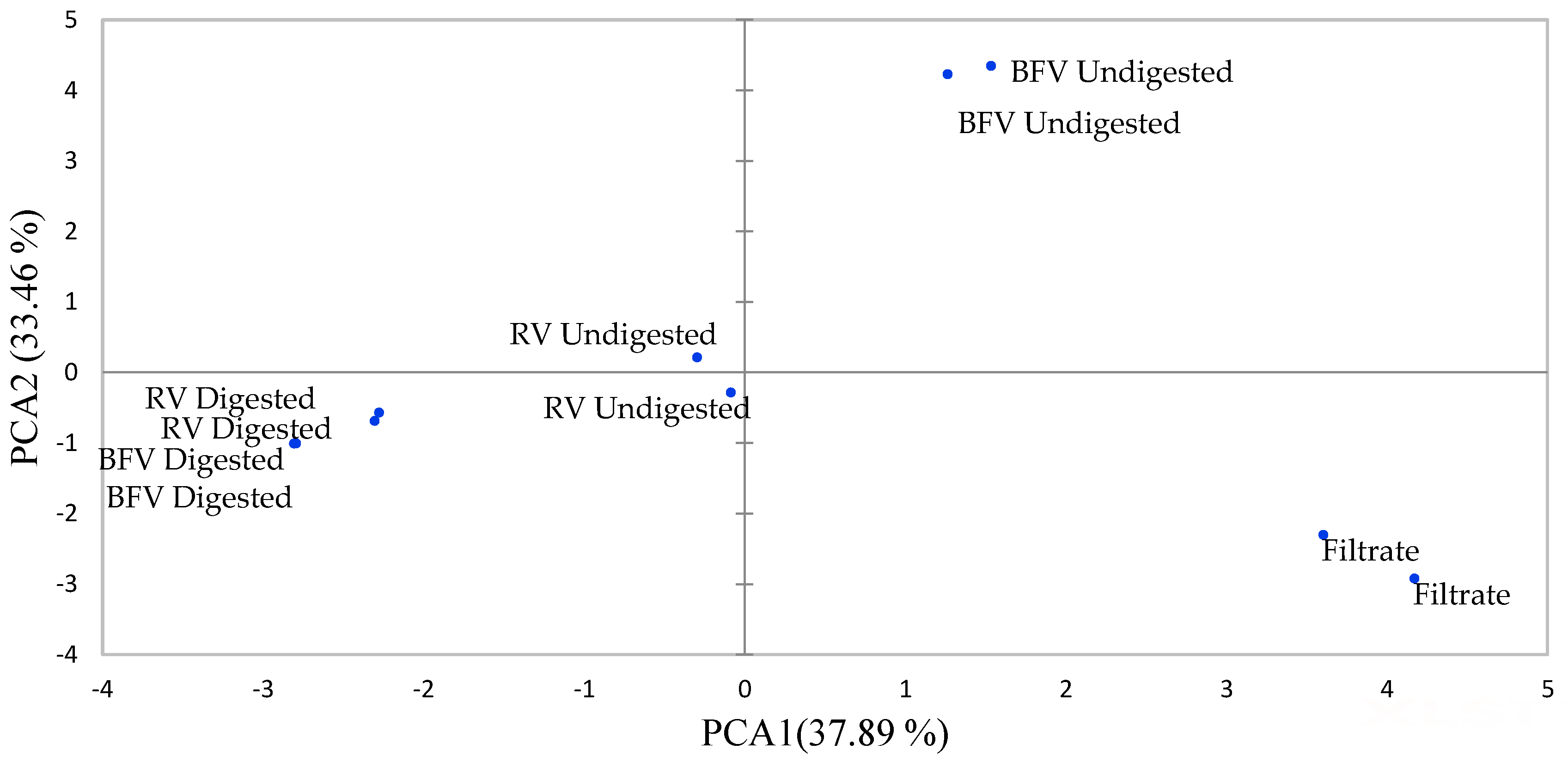
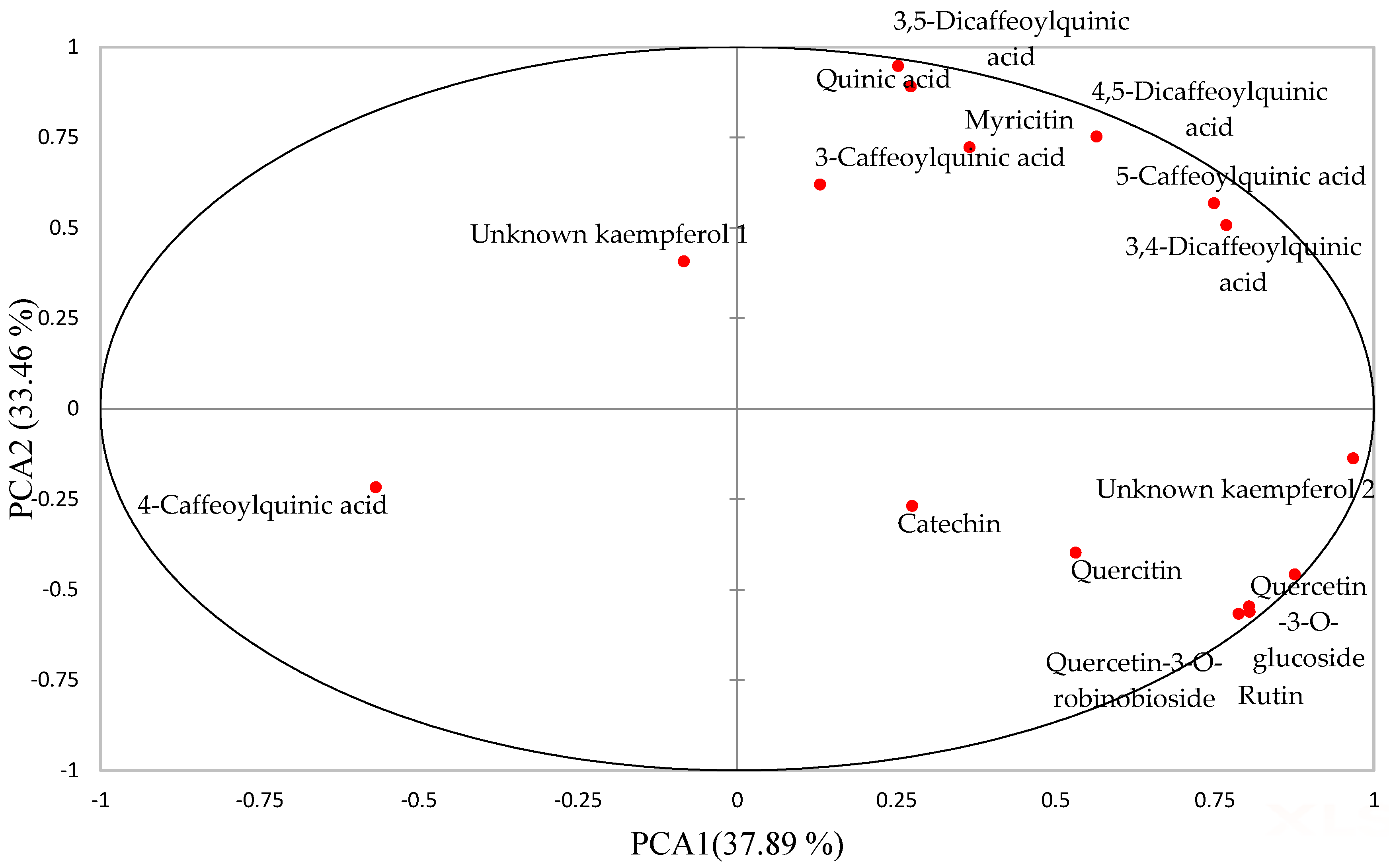
2.1. Effect of Boiling and In Vitro Digestion on Compounds Identified in Raw and Boiled AFRICAN Pumpkin by UPLC-QTOF-MS
2.2. Effect of Boiling on TPC and TFC of African Pumpkin and Spinach
2.3. Effect of In Vitro Digestion on TPC and TFC of African Pumpkin and Spinach
2.4. Effect of Boiling on Radical Scavenging Activity/Redox Potential of African Pumpkin and Spinach
2.5. Effect of In Vitro Digestion on Radical Scavenging Activity/Redox Potential African Pumpkin and Spinach
2.6. Cellular Antioxidant Activity
2.7. Macromolecule Protective Ability
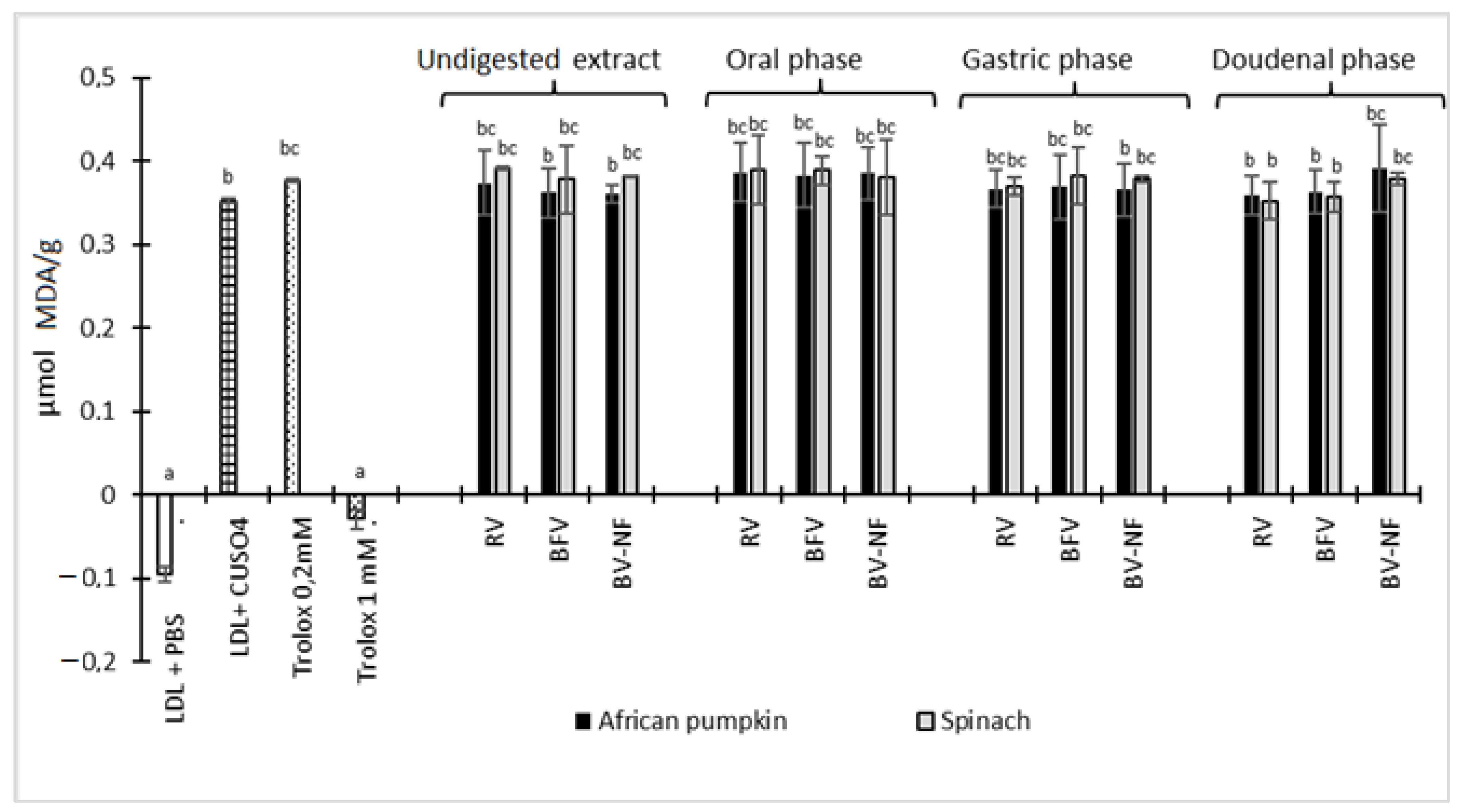
2.7.1. Copper-Mediated Human LDL Oxidation
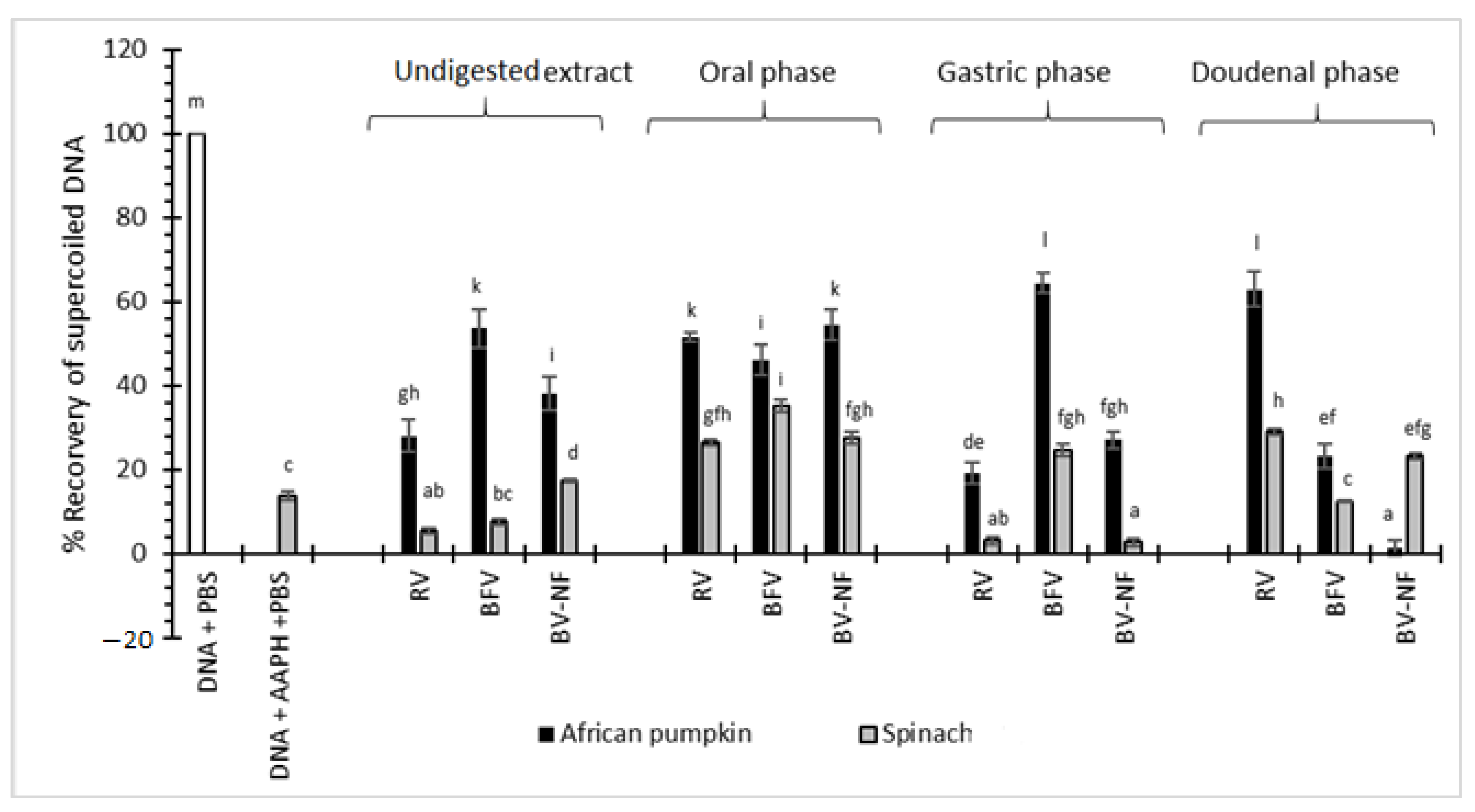
2.7.2. Inhibition of AAPH-Induced Oxidative DNA Damage
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Sample Preparation
3.3. Methanolic Extraction for Undigested Samples
3.4. Human Simulated Gastrointestinal Digestion
3.5. Determination of Phenolic Composition
3.5.1. Analytical Conditions for UPLC-DAD-QTOF-MS
3.5.2. Determination of Total Phenolic Content
3.5.3. Determination of Total Flavonoid Content
3.5.4. Estimation of Bioaccessibility
3.6. Determination of Radical Scavenging Activity/Redox Potential
3.6.1. Trolox Equivalence Antioxidant Capacity (TEAC) or the ABTS Assay
3.6.2. Ferric Reducing Antioxidant Power (FRAP) Assay
3.6.3. Oxygen Radical Antioxidant Capacity (ORAC) Assay
3.7. Cellular Antioxidant Activity
2′,7′-Dichlorodihydrofluorescein Diacetate (DCFH-DA) Assay
3.8. Macromolecule Protective Ability
3.8.1. Copper-Mediated LDL Oxidation
3.8.2. Inhibition of Oxidative DNA Damage
3.9. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Uchegbu, R.I.; Bako, S.S.; Olehi, L.C.N.; Achinihu, I.O. GC/Ms Analysis and identification of phytochemicals present in the fruits of Mormodica balsamina Linn. IOSR J. Appl. Chem. 2015, 8, 39–42. [Google Scholar]
- Mshelia, H.S.; Karumi, Y.; Dibal, N.I. Therapeutic effect of Momordica balsamina leaf extract on ethanol-induced gastric ulcer in Wistar rats. Ann. Res. Hosp. 2017, 1, 1–5. [Google Scholar] [CrossRef]
- Siboto, A.; Sibiya, N.; Khathi, A.; Ngubane, P. The Effects of Momordica balsamina methanolic extract on kidney function in STZ-Induced diabetic rats: Effects on selected metabolic markers. J. Diabetes Res. 2018, 2018, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boshielo, I.T. In Vitro Evaluation of Anticancer Effect of Momordica balsamina Linn. Leaf Extract in Human Breast MCF-7 Cancer Cells. Master’s Thesis, University of Limpopo, Polokwane, South Africa, 2017. [Google Scholar]
- Dufour, C.; Loonis, M.; Delosière, M.; Buffière, C.; Hafnaoui, N.; Santé-Lhoutellier, V.; Rémond, D. The matrix of fruit & vegetables modulates the gastrointestinal bioaccessibility of polyphenols and their impact on dietary protein digestibility. Food Chem. 2018, 240, 314–322. [Google Scholar] [PubMed]
- Gunathilake, K.; Ranaweera, K.; Rupasinghe, H. Effect of different cooking methods on polyphenols, carotenoids and antioxidant activities of selected edible leaves. Antioxidants 2018, 7, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madala, N.E.; Tugizimana, F.; Steenkamp, P.A. Development and optimization of an UPLC-QTOF-MS/MS method based on an in-source collision induced dissociation approach for comprehensive discrimination of chlorogenic acids isomers from Momordica plant species. J. Anal. Methods Chem. 2014, 2014, 1–7. [Google Scholar] [CrossRef]
- Madala, N.E.; Piater, L.; Dubery, I.; Steenkamp, P. Distribution patterns of flavonoids from three Momordica species by ultra-high performance liquid chromatography quadrupole time of flight mass spectrometry: A metabolomic profiling approach. Rev. Bras. Farmacogn. 2016, 26, 507–513. [Google Scholar] [CrossRef] [Green Version]
- Souda, S.; George, S.; Mannathoko, N.; Goercke, I.; Chabaesele, K. Antioxidant and antibacterial activity of methanol extract of Momordica balsamina. IRA-Int. J. Appl. Sci. 2018, 10, 7. [Google Scholar] [CrossRef] [Green Version]
- Akula, U.S.; Odhav, B. In vitro 5-Lipoxygenase inhibition of polyphenolic antioxidants from undomesticated plants of South Africa. J. Med. Plants Res. 2008, 2, 207–212. [Google Scholar]
- De Santiago, E.; Pereira-Caro, G.; Moreno-Rojas, J.M.; Cid, C.; De Peña, M.P. Digestibility of (poly)phenols and antioxidant activity in raw and cooked cactus cladodes (Opuntia ficus-indica). J. Agric. Food Chem. 2018, 66, 5832–5844. [Google Scholar] [CrossRef]
- Alminger, M.; Aura, A.M.; Bohn, T.; Dufour, C.; El, S.N.; Gomes, A.; Karakaya, S.; Martínez-Cuesta, M.C.; Mcdougall, G.J.; Requena, T.; et al. In vitro models for studying secondary plant metabolite digestion and bioaccessibility. Compr. Rev. Food Sci. Food Saf. 2014, 13, 413–436. [Google Scholar] [CrossRef] [Green Version]
- Parada, J.; Aguilera, J.M. Food microstructure affects the bioavailability of several nutrients. J. Food Sci. 2007, 72, R21–R32. [Google Scholar] [CrossRef]
- Chen, Y.; Fu, Z.; Tu, Z.-C.; Wang, H.; Zhang, L.; Xie, X.; Liu, G. Influence of in vitro gastrointestinal digestion on the bioavailability and antioxidant activity of polyphenols from Ipomoea batatas leaves. Int. J. Food Sci. Technol. 2017, 52, 1131–1137. [Google Scholar] [CrossRef]
- Alegría, A.; Garcia-llatas, G.; Cilla, A. In vitro intestinal tissue models: General introduction. In The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Verhoeckx, K., Cotter, P., López-Expósito, I., Kleiveland, C., Lea, T., Mackie, A., Requena, T., Swiateck, D., Wichers, H., Eds.; Springer: Cham, Switzerland; New York, NY, USA, 2015; pp. 239–244. ISBN 9783319161044. [Google Scholar]
- Nagarani, G.; Abirami, A.; Siddhuraju, P. Food prospects and nutraceutical attributes of Momordica species: A potential tropical bioresources—A review. Food Sci. Hum. Wellness 2014, 3, 117–126. [Google Scholar] [CrossRef] [Green Version]
- Takenaka, M.; Nanayama, K.; Isobe, S.; Murata, M. Changes in caffeic acid derivatives in sweet potato (Ipomoea batatas L.) during cooking and processing. Biosci. Biotechnol. Biochem. 2006, 70, 172–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Zhang, B.; Xia, Y.; Li, H.; Shi, X.; Wang, J.; Deng, Z. Bioaccessibility and transformation pathways of phenolic compounds in processed mulberry (Morus alba L.) leaves after in vitro gastrointestinal digestion and faecal fermentation. J. Funct. Foods 2019, 60, 103–406. [Google Scholar] [CrossRef]
- Spínola, V.; Llorent-Martínez, E.J.; Castilho, P.C. Antioxidant polyphenols of Madeira sorrel (Rumex maderensis): How do they survive to in vitro simulated gastrointestinal digestion? Food Chem. 2018, 259, 105–112. [Google Scholar] [CrossRef]
- Pinto, J.; Spínola, V.; Llorent-Martínez, E.J.; Córdova, M.L.F.; Molina-García, L.; Castilho, P.C. Polyphenolic profile and antioxidant activities of Madeiran elderberry (Sambucus lanceolata) as affected by simulated in vitro digestion. Food Res. Int. 2017, 100, 404–410. [Google Scholar] [CrossRef]
- Murador, D.; Braga, A.R.; Da Cunha, D.; De Rosso, V. Alterations in phenolic compound levels and antioxidant activity in response to cooking technique effects: A meta-analytic investigation. Crit. Rev. Food Sci. Nutr. 2018, 58, 169–177. [Google Scholar] [CrossRef]
- Amin, I.; Norazaidah, Y.; Hainida, K.I.E. Antioxidant activity and phenolic content of raw and blanched Amaranthus species. Food Chem. 2006, 94, 47–52. [Google Scholar] [CrossRef]
- Celep, E.; Akyüz, S.; İnan, Y.; Yesilada, E. Assessment of potential bioavailability of major phenolic compounds in Lavandula stoechas L. ssp. stoechas. Ind. Crops Prod. 2018, 118, 111–117. [Google Scholar] [CrossRef]
- Dolinsky, M.; Agostinho, C.; Ribeiro, D.; Rocha, G.D.S.; Barroso, S.G.; Ferreira, D.; Polinati, R.; Ciarelli, G.; Fialho, E. Effect of different cooking methods on the polyphenol concentration and antioxidant capacity of selected vegetables. J. Culin. Sci. Technol. 2016, 14, 1–12. [Google Scholar] [CrossRef]
- Subudhi, B.B.; Bhoi, A. Antioxidative effects of Brassica juncea and Moringa oliefera prepared by different processing methods. J. Food Sci. Technol. 2014, 51, 790–794. [Google Scholar] [CrossRef] [Green Version]
- Scrob, T.; Hosu, A.; Cimpoiu, C. The influence of in vitro gastrointestinal digestion of Brassica oleracea florets on the antioxidant activity and chlorophyll, carotenoid and phenolic content. Antioxidants 2019, 8, 212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonçalves, S.; Moreira, E.; Andrade, P.B.; Valentão, P.; Romano, A. Effect of in vitro gastrointestinal digestion on the total phenolic contents and antioxidant activity of wild Mediterranean edible plant extracts. Eur. Food Res. Technol. 2018, 245, 753–762. [Google Scholar] [CrossRef]
- Seraglio, S.K.T.; Gonzaga, L.V.; Schulz, M.; Vitali, L.; Micke, G.A.; Costa, A.C.O.; Fett, R.; Borges, G.D.S.C. Effects of gastrointestinal digestion models in vitro on phenolic compounds and antioxidant activity of juçara (Euterpe edulis). Int. J. Food Sci. Technol. 2018, 53, 1824–1831. [Google Scholar] [CrossRef]
- Pinacho, R.; Yolanda, R.; Astiasarán, I.; Ansorena, D.; Isabel, M.; Cavero, R.Y.; Astiasarán, I.; Ansorena, D.; Calvo, M.I. Phenolic compounds of blackthorn (Prunus spinosa L.) and influence of in vitro digestion on their antioxidant capacity. J. Funct. Foods 2015, 19, 49–62. [Google Scholar] [CrossRef]
- Palafox-Carlos, H.; Ayala-Zavala, J.F.; González-Aguilar, G.A. The role of dietary fiber in the bioaccessibility and bioavailability of fruit and vegetable antioxidants. J. Food Sci. 2011, 76, 6–15. [Google Scholar] [CrossRef] [Green Version]
- Acosta-estrada, B.A.; Gutiérrez-uribe, J.A.; Serna-saldívar, S.O. Bound phenolics in foods, a review. Food Chem. 2014, 152, 46–55. [Google Scholar] [CrossRef]
- Lucas-González, R.; Viuda-Martos, M.; Pérez-Alvarez, J.A.; Fernández-López, J. In vitro digestion models suitable for foods: Opportunities for new fields of application and challenges. Food Res. Int. 2018, 107, 423–436. [Google Scholar] [CrossRef]
- Reddy, P.E.; Manohar, S.M.; Reddy, S.V.; Bitla, A.R.; Vishnubhotla, S.; Rao, S.; Narasimha, P.V.L. Ferric reducing ability of plasma and lipid peroxidation in hemodialysis patients: Intradialytic changes. Int. J. Nephrol. Urol. 2010, 2, 414–421. [Google Scholar]
- Tomović, V.; Jokanović, M.; Šojić, B.; Škaljac, S.; Ivić, M. Plants as natural antioxidants for meat products. In IOP Conference Series: Earth and Environmental Science, Proceedings of the 59th International Meat Industry Conference MEATCON2017, Zlatibor, Serbia, 1–4 October 2017; IOP Publishing: Bristol, UK, 2017; pp. 1–9. [Google Scholar]
- Ali, M.; Mahsa, M.; Mehrzad, J.B. Effect of boiling cooking on antioxidant activities and phenolic content of selected Iranian vegetables. Res. J. Pharm. Biol. Chem. Sci. 2015, 6, 636–641. [Google Scholar]
- Kapusta-Duch, J.; Kusznierewicz, B.; Leszczyńska, T.; Borczak, B. Effect of culinary treatment on changes in the contents of selected nutrients and non-nutrients in curly Kale (Brassica oleracea Var. acephala). J. Food Process. Preserv. 2016, 40, 1280–1288. [Google Scholar] [CrossRef]
- Armesto, J.; Gómez-Limia, L.; Carballo, J.; Martínez, S. Effects of different cooking methods on the antioxidant capacity and flavonoid, organic acid and mineral contents of Galega Kale (Brassica oleracea var. acephala cv. Galega). Int. J. Food Sci. Nutr. 2019, 70, 136–149. [Google Scholar] [CrossRef] [PubMed]
- Ratseewo, J.; Tangkhawanit, E.; Meeso, N.; Kaewseejan, N.; Siriamornpun, S. Changes in antioxidant properties and volatile compounds of kaffir lime leaf as affected by cooking processes. Int. Food Res. J. 2016, 23, 188–196. [Google Scholar]
- López-García, G.; Baeza-Jiménez, R.; Garcia-Galindo, H.S.; Dublán-García, O.; Lopez-Martinez, L.X. Cooking treatments effect on bioactive compounds and antioxidant activity of quintonil (Amaranthus hybridus) harvested in spring and fall seasons. CYTA-J. Food 2018, 16, 707–714. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Hardin, R.; Ogutu, S.; Verghese, M.; Boateng, J. Preliminary analysis of in vitro digestion and bioactivity assessment of basil and ginger in human liver cancer cell line. J. Biol. Sci. 2016, 16, 202–214. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, R.; Pal, D.; Roy, S. Characterization of Leucas aspera and evaluation of antioxidant activities before and after being subjected to digestion enzymes. Int. J. Veg. Sci. 2020, 26, 302–320. [Google Scholar] [CrossRef]
- Somchaidee, P.; Tedsree, K. Green synthesis of high dispersion and narrow size distribution of zero-valent iron nanoparticles using guava leaf (Psidium guajava L) extract. Adv. Nat. Sci. Nanosci. Nanotechnol. 2018, 9, 035006. [Google Scholar] [CrossRef]
- Halliwell, B. Dietary polyphenols: Good, bad, or indifferent for your health? Cardiovasc. Res. 2007, 73, 341–347. [Google Scholar] [CrossRef]
- Rychlik, J.; Olejnik, A.; Olkowicz, M.; Kowalska, K.; Juzwa, W.; Myszka, K.; Dembczyński, R.; Moyer, M.P.; Grajek, W. Antioxidant capacity of broccoli sprouts subjected to gastrointestinal digestion. J. Sci. Food Agric. 2015, 95, 1892–1902. [Google Scholar] [CrossRef]
- Pérez-Burillo, S.; Rufián-Henares, J.Á.; Pastoriza, S. Effect of home cooking on the antioxidant capacity of vegetables: Relationship with Maillard reaction indicators. Food Res. Int. 2019, 121, 514–523. [Google Scholar] [CrossRef]
- Trpkovic, A.; Resanovic, I.; Stanimirovic, J.; Radak, D.; Mousa, S.A.; Cenic-Milosevic, D.; Jevremovic, D.; Isenovic, E.R. Oxidized low-density lipoprotein as a biomarker of cardiovascular diseases. Crit. Rev. Clin. Lab. Sci. 2015, 52, 70–85. [Google Scholar] [CrossRef] [PubMed]
- Moyo, S.M.; Serem, J.C.; Bester, M.J.; Mavumengwana, V.; Kayitesi, E. Influence of boiling and subsequent phases of digestion on the phenolic content, bioaccessibility, and bioactivity of Bidens pilosa (Blackjack) leafy vegetable. Food Chem. 2020, 311, 126023. [Google Scholar] [CrossRef]
- Moyo, S.M.; Serem, J.C.; Bester, M.J.; Mavumengwana, V.; Kayitesi, E. The impact of boiling and in vitro human digestion of Solanum nigrum complex (Black nightshade) on phenolic compounds bioactivity and bioaccessibility. Food Res. Int. 2020, 137, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Taira, J.; Taira, K.; Ohmine, W.; Nagata, J. Mineral determination and anti-LDL oxidation activity of sweet potato (Ipomoea batatas L.) leaves. J. Food Compos. Anal. 2013, 29, 117–125. [Google Scholar] [CrossRef]
- Kiokias, S.; Proestos, C.; Oreopoulou, V. Effect of natural food antioxidants against LDL and DNA oxidative changes. Antioxidants 2018, 7, 133. [Google Scholar] [CrossRef] [Green Version]
- Unlu, S.; Saglar, E. Evaluation of cytogenetic and genotoxic effects of oxalic acid by the alkaline comet assay and QRT PCR in human buccal epithelial cells. Anal. Quant. Cytopathol. Histopathol. 2015, 37, 347–352. [Google Scholar]
- Lin, K.H.; Yang, Y.Y.; Yang, C.M.; Huang, M.Y.; Lo, H.F.; Liu, K.C.; Lin, H.S.; Chao, P.Y. Antioxidant activity of herbaceous plant extracts protect against hydrogen peroxide-induced DNA damage in human lymphocytes. Food Nutr. Sci. 2013, 6, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moyo, S.M.; Kayitesi, E.; Mavumengwana, V.; Madala, N.E. Effects of cooking and drying on the total phenolic, total flavonoid content, antioxidant and antibacterial activity of Cleome gynandra (Spider plant). In Proceedings of the International Conference on Advances in Science Engineering, Technology & Natural Resources, Parys, South Africa, 24–25 November 2016; pp. 136–141. [Google Scholar]
- Serem, J.C.; Bester, M.J. Physicochemical properties, antioxidant activity and cellular protective effects of honeys from southern Africa. Food Chem. 2012, 133, 1544–1550. [Google Scholar] [CrossRef]
- Amaral, S.; Mira, L.; Nogueira, J.M.; Silva, A.P.; Helena, F.M. Plant extracts with anti-inflammatory properties—A new approach for characterization of their bioactive compounds and establishment of structure–antioxidant activity relationships. Bioorg. Med. Chem. 2009, 17, 1876–1883. [Google Scholar] [CrossRef] [PubMed]
- Serem, J.C. Identification and Characterization of Bioactivity of Simulated Gastrointestinal Digested Indigenous Southern African Honey Samples. Ph.D. Thesis, University of Pretoria, Pretoria, South Africa, 2018. [Google Scholar]
- Antoni, A.; Muñoz, M.F.; Sandro, A. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-Hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 1–31. [Google Scholar]
- Kayitesi, E. Micronisation of Cowpeas: The Effects on Sensory Quality, Phenolic Compounds and Bioactive Properties. Ph.D. Thesis, University of Pretoria, Pretoria, South Africa, 2013. [Google Scholar]
- Akoglu, H. User’s guide to correlation coefficients. Turk. J. Emerg. Med. 2018, 18, 91–93. [Google Scholar] [CrossRef] [PubMed]
- Moyo, S.M. Influence of In Vitro Digestion on the Bioaccessibility and Bioactivity of Phenolic Compounds from Raw and Boiled African Indigenous Green Leafy Vegetables. Ph.D. Thesis, University of Johannesburg, Johannesburg, South Africa, 2020. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rt | Precursor Ion [M − H]− | Formula [M − H]− | Diagnostic Fragments | Uvmax | Compound Name | Effect of Boiling * | Effect of Digestion * | ||
|---|---|---|---|---|---|---|---|---|---|
| RV | BFV | RV | BFV | ||||||
| 1.71 | 191.06 | C7H12O6 | 127, 111, 93, 85 | 210 | Quinic acid | 0.13 ± 0.01 | 3.38 ± 0.15 | 0.61 ± 0.14 | 0.46 ± 0.04 |
| 9.86 | 353.09 | C16H17O9 | 191, 179, 173, 135 | 326 | 3-Caffeoylquinic acid | 16.40 ± 0.61 | 11.97 ± 0.39 | ND | 0.83 ± 0.03 |
| 11.36 | 289.07 | C15H13O6 | 245, 205, 203, 123, 109 | 278 | Catechin | 79.21 ± 8.62 | 1.15 ± 0.19 | 6.07 ± 0.24 | 1.13 ± 0.44 |
| 11.42 | 353.09 | C16H17O9 | 191, 179,173,135 | 326 | 4-Caffeoylquinic acid | ND | ND | ND | 0.77 ± 0.07 |
| 12.41 | 353.09 | C16H17O9 | 191, 179, 173, 135 | 326 | 5-Caffeoylquinic acid | 36.64 ± 0.29 | 52.56 ± 3.63 | ND | ND |
| 16.57 | 317.24 | C15H9O8 | 315 | 255, 375 | Myricitin | ND | 22.38 ± 1.55 | 2.64 ± 0.68 | 7.87 ± 0.39 |
| 16.87 | 609.15 | C27H29O16 | 300, 301, 271, 255 | 257, 353 | Quercetin-3-O-robinobioside | 1.87 ± 0.16 | 2.50 ± 0.09 | 0.76 ± 0.03 | 0.71 ± 0.01 |
| 17.14 | 609.15 | C27H29O16 | 300, 301, 271, 255 | 353 | Rutin | 6.84 ± 0.12 | 2.85 ± 0.30 | 1.26 ± 0.03 | 0.68 ± 0.05 |
| 17.64 | 463.09 | C21H19O12 | 300, 301, 271, 255 | 348 | Quercetin-3-O-glucoside | 111.70 ± 1.66 | 122.08 ± 0.40 | ND | 0.81 ± 0.01 |
| 18.9 | 491 | C23H23O12 | 285, 151, 135 | 307 | Unknown-kaempferol 1 | ND | 3.68 ± 0.57 | 5.05 ± 0.04 | ND |
| 19.01 | 515.12 | C25H23O12 | 353, 179, 173 | 325 | 3,4-Dicaffeoylquinic acid | ND | 47.71 ± 6.86 | ND | 0.77 ± 0.07 |
| 19.69 | 515.12 | C25H23O12 | 353, 179, 173 | 325 | 3,5-Dicaffeoylquinic acid | 24.68 ± 1.08 | 62.27 ± 2.85 | 1.28 ± 0.04 | 1.61 ± 0.02 |
| 20.84 | 515.12 | C25H23O12 | 353, 179, 173 | 325 | 4,5-Dicaffeoylquinic acid | 68.93 ± 13.1 | 110.64 ± 12.2 | 1.94 ± 0.09 | ND |
| 22.35 | 491 | C23H23O12 | 285, 151, 135 | 307 | Unknown-kaempferol 2 | 3.72 ± 0.31 | 7.99 ± 0.25 | 1.13 ± 0.12 | 1.17 ± 0.64 |
| 23.86 | 301.03 | C15H9O7 | 151, 179 | 256 | Quercetin | 15.94 ± 0.13 | 2.19 ± 0.21 | 3.60 ± 0.05 | 0.77 ± 0.08 |
| Samples | TPC mg GAE/g dw | TFC mg QE/g dw | TPC% BA | TFC% BA |
|---|---|---|---|---|
| African pumpkin (African pumpkin) | ||||
| Undigested | ||||
| RV | 64.53 ± 9.1 bc | 69.00 ± 1.3 i | - | - |
| BFV | 60.41 ± 15 bc | 44.34 ± 1.2 gh | - | - |
| BV-NF | 52.27 ± 2.0 b | 63.90 ± 5.6 i | - | - |
| Oral digestion | ||||
| RV | 37.13 ± 0.9 ab | 4.33 ± 0.3 a | 57.54 | 6.27 |
| BFV | 43.38 ± 1.9 b | 18.44 ± 7.2 b | 71.81 | 41.58 |
| BV-NF | 53.83 ± 13 b | 25.31 ± 3.7 bcd | 102.98 | 39.61 |
| Gastric digestion | ||||
| RV | 37.42 ± 4.3 ab | 30.22 ± 0.2 cde | 57.99 | 43.94 |
| BFV | 40.98 ± 5.5 b | 28.43 ± 2.4 cd | 67.83 | 64.12 |
| BV-NF | 42.58 ± 1.5 b | 64.69 ± 16 i | 81.46 | 101.24 |
| Duodenal digestion | ||||
| RV | 44.82 ± 2.5 b | 50.07 ± 3.7 h | 69.46 | 72.57 |
| BFV | 44.55 ± 3.4 b | 39.20 ± 6.5 efg | 73.74 | 88.40 |
| BV-NF | 4.03 ± 0.7 a | 3.50 ± 3.6 a | 7.71 | 5.48 |
| Digestion effect on | ||||
| RV | Unchanged | Reduced | ||
| BFV | Unchanged | Unchanged | ||
| BV-NF | Reduced | Reduced | ||
| Spinach | ||||
| Undigested | ||||
| RV | 349.38 ± 7.8 h | 133.09 ± 2.6 k | - | - |
| BFV | 279.10 ± 1.7 fg | 93.41 ± 0.89 j | - | - |
| BV-NF | 289.78 ± 4.4 g | 93.82 ± 2.7 j | - | - |
| Oral digestion | ||||
| RV | 248.58 ± 4.8 fg | 69.14 ± 3.2 i | 71.14 | 51.94 |
| BFV | 115.18 ± 5.2 d | 23.76 ± 1.4 bcd | 41.27 | 25.44 |
| BV-NF | 128.15 ± 3.9 d | 42.11 ± 2.1 fgh | 44.22 | 44.88 |
| Gastric digestion | ||||
| RV | 260.03 ± 2.4 fg | 48.83 ± 4.7 gh | 74.43 | 36.69 |
| BFV | 201.68 ± 2.9 e | 33.76 ± 0.18 def | 72.26 | 36.14 |
| BV-NF | 401.21 ± 2.9 h | 33.17 ± 4.2 k | 138.45 | 35.35 |
| Duodenal digestion | ||||
| RV | 124.25 ± 8.9 d | 29.06 ± 3.13 cd | 35.56 | 21.83 |
| BFV | 93.88 ± 6.0 cd | 16.44 ± 2.1 b | 33.64 | 17.60 |
| BV-NF | 69.01 ± 2.7 bc | 17.37 ± 1.3 b | 23.81 | 18.51 |
| Digestion effect on | ||||
| RV | Reduced | Reduced | - | - |
| BFV | Reduced | Reduced | - | - |
| BV-NF | Reduced | Reduced | - | - |
| Samples | TEAC µM TE/g dw | ORAC µM TE/g dw | FRAP µM TE/g dw |
|---|---|---|---|
| African pumpkin | |||
| Undigested extract | |||
| RV | 216.50 ± 37 bcd | 531.29 ± 5.7 cd | 50.14 ± 6.0 b |
| BFV | 211.50 ± 21 bcd | 406.66 ± 12 bc | 48.36 ± 11 b |
| BV-NF | 262.11 ± 17 bcde | 340.46 ± 4.0 bc | 148.78 ± 16 ef |
| Oral digestion | |||
| RV | 221.08 ± 6.2 bcd | 450.56 ± 1.1 bc | 88.62 ± 1.0 cd |
| BFV | 290.85 ± 1.8 cde | 429.60 ± 10 bc | 45.68 ± 5.6 b |
| BV-NF | 298.35 ± 7.1 de | 193.87 ± 0.3 a | 60.18 ± 0.7 bc |
| Gastric digestion | |||
| RV | 154.84 ± 37 b | 497.99 ± 6.4 bc | 52.34 ± 15 b |
| BFV | 167.76 ± 23 b | 483.05 ± 7.6 bc | 47.83 ± 3.9 b |
| BV-NF | 184.42 ± 17 bc | 186.57 ± 1.3 a | 40.91 ± 2.5 b |
| Duodenal digestion | |||
| RV | 179.00 ± 23 b | 283.53 ± 6.0 ab | 47.43 ± 7.8 b |
| BFV | 193.79 ± 9 bcd | 326.53 ± 2.1 bc | 116.93 ± 1.2 de |
| BV-NF | −203.21 ± 20 a | 191.30 ± 0.1 a | −36.92 ± 4.8 a |
| Digestion effect on | |||
| RV | Unchanged | Reduced | Unchanged |
| BFV | Unchanged | Unchanged | Increased |
| BV-NF | Reduced | Reduced | Reduced |
| Spinach | |||
| Undigested extract | |||
| RV | 1090.99 ± 1.4 i | 756.20 ± 1.5 d | 334.16 ± 4.5 k |
| BFV | 1021.96 ± 2.4 i | 1223.21 ± 9.7 ef | 263.46 ± 0.96 j |
| BV-NF | 1084.84 ± 1.5 i | 1181.94 ± 2.5 ef | 351.83 ± 2.9 k |
| Oral digestion | |||
| RV | 712.53 ± 9.8 g | 637.67 ± 3.7 f | 172.38 ± 2.8 fg |
| BFV | 495.93 ± 9.3 f | 1363.33 ± 1.7 f | 190.73 ± 1.7 gh |
| BV-NF | 370.37 ± 3.3 e | 540.65 ± 4.3 cd | 184.62 ± 0.96 g |
| Gastric digestion | |||
| RV | 857.14 ± 7.0 h | 1681.07 ± 5.2 g | 228.12 ± 6.7 i |
| BFV | 748.83 ± 5.8 gh | 1179.40 ± 7.2 ef | 139.07 ± 5.8 e |
| BV-NF | 1090.99 ± 9.3 i | 546.13 ± 4.6 cd | 249.86 ± 2.7 ij |
| Duodenal digestion | |||
| RV | 534.01 ± 6.6 f | 1000.71 ± 1.2 e | 226.76 ± 2.9 i |
| BFV | 523.30 ± 5.9 f | 1004.29 ± 7.8 e | 218.60 ± 2.9 hi |
| BV-NF | 172.21 ± 1.5 b | 563.79 ± 2.2 cd | 122.76 ± 1.5 e |
| Digestion effect on | |||
| RV | Reduced | Increased | Reduced |
| BFV | Reduced | Unchanged | Unchanged |
| BV-NF | Reduced | Reduced | Reduced |
| Samples | % Oxidative Damage DCFH-DAL929 | % Oxidative Damage DCFH-DACaco-2 |
|---|---|---|
| African pumpkin | ||
| Undigested extract | Control (AAPH + L929 cells + PBS) 100% | Control (AAPH + Caco-2 cells + PBS) 100% |
| RV | −16.54 ± 12 a | −8.88 ± 8.9 ab |
| BFV | −11.84 ± 3.8 ab | −6.50 ± 1.1 ab |
| BV-NF | −15.50 ± 6.1 a | −10.91 ± 1.3 a |
| Oral digestion | ||
| RV | 78.39 ± 5.6 d | 515.25 ± 6.0 h |
| BFV | 8.77 ± 1.5 b | 29.96 ± 4.5 f |
| BV-NF | −5.17 ± 5.1 ab | −0.96 ± 1.3 abcd |
| Gastric digestion | ||
| RV | 55.65 ± 3.6 c | −13.83 ± 2.0 a |
| BFV | −13.45 ± 5.5 ab | 6.40 ± 0.1 bcde |
| BV-NF | −9.22 ± 0.1 ab | −5.65 ± 0.6 ab |
| Duodenal digestion | ||
| RV | 64.22 ± 0.7 cd | −0.90 ± 1.7 abcd |
| BFV | 114.93 ± 0.8 e | 68.76 ± 2.3 g |
| BV-NF | −2.93 ± 6.2 ab | −4.46 ± 1.6 abcd |
| Digestion effect on | ||
| RV | Increased | Unchanged |
| BFV | Increased | Increased |
| BV-NF | Unchanged | Unchanged |
| Spinach | ||
| Undigested extract | ||
| RV | −21.52 ± 4.7 a | −5.39 ± 2.7 abc |
| BFV | −20.33 ± 4.1 a | −13.39 ± 6.8 a |
| BV-NF | −20.57 ± 4.8 a | −10.71 ± 1.0 a |
| Oral digestion | ||
| RV | 102.05 ± 1.0 e | 16.49 ± 0.8 ef |
| BFV | −6.86 ± 3.7 ab | 10.19 ± 1.1 cde |
| BV-NF | −7.22 ± 4.1 ab | 6.64 ± 0.3 bcde |
| Gastric digestion | ||
| RV | −0.006 ± 7.5 ab | −0.83 ± 0.2 abcd |
| BFV | −20.59 ± 5.1 a | −0.05 ± 0.04 abcd |
| BV-NF | −10.21 ± 6.1 ab | 10.74 ± 1.9 de |
| Duodenal digestion | ||
| RV | −3.19 ± 0.02 ab | −10.90 ± 5.1 a |
| BFV | −3.09 ± 0.8 ab | −5.26 ± 2.7 bcde |
| BV-NF | −5.61 ± 1.4 ab | −7.16 ± 1.1 ab |
| Digestion effect on | ||
| RV | Unchanged | Unchanged |
| BFV | Unchanged | Increased |
| BV-NF | Unchanged | Unchanged |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moyo, S.M.; Serem, J.C.; Bester, M.J.; Mavumengwana, V.; Kayitesi, E. Hydrothermal Processing and In Vitro Simulated Human Digestion Affects the Bioaccessibility and Bioactivity of Phenolic Compounds in African Pumpkin (Momordica balsamina) Leaves. Molecules 2021, 26, 5201. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26175201
Moyo SM, Serem JC, Bester MJ, Mavumengwana V, Kayitesi E. Hydrothermal Processing and In Vitro Simulated Human Digestion Affects the Bioaccessibility and Bioactivity of Phenolic Compounds in African Pumpkin (Momordica balsamina) Leaves. Molecules. 2021; 26(17):5201. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26175201
Chicago/Turabian StyleMoyo, Siphosanele Mafa, June C. Serem, Megan J. Bester, Vuyo Mavumengwana, and Eugenie Kayitesi. 2021. "Hydrothermal Processing and In Vitro Simulated Human Digestion Affects the Bioaccessibility and Bioactivity of Phenolic Compounds in African Pumpkin (Momordica balsamina) Leaves" Molecules 26, no. 17: 5201. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26175201