
Metabolomic Profiling of Fresh Goji (Lycium barbarum L.) Berries from Two Cultivars Grown in Central Italy: A Multi-Methodological Approach

 , , , , , , , , , and
, , , , , , , , , and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Goji Berries Metabolite Profile
2.1.1. 1H NMR Assignment
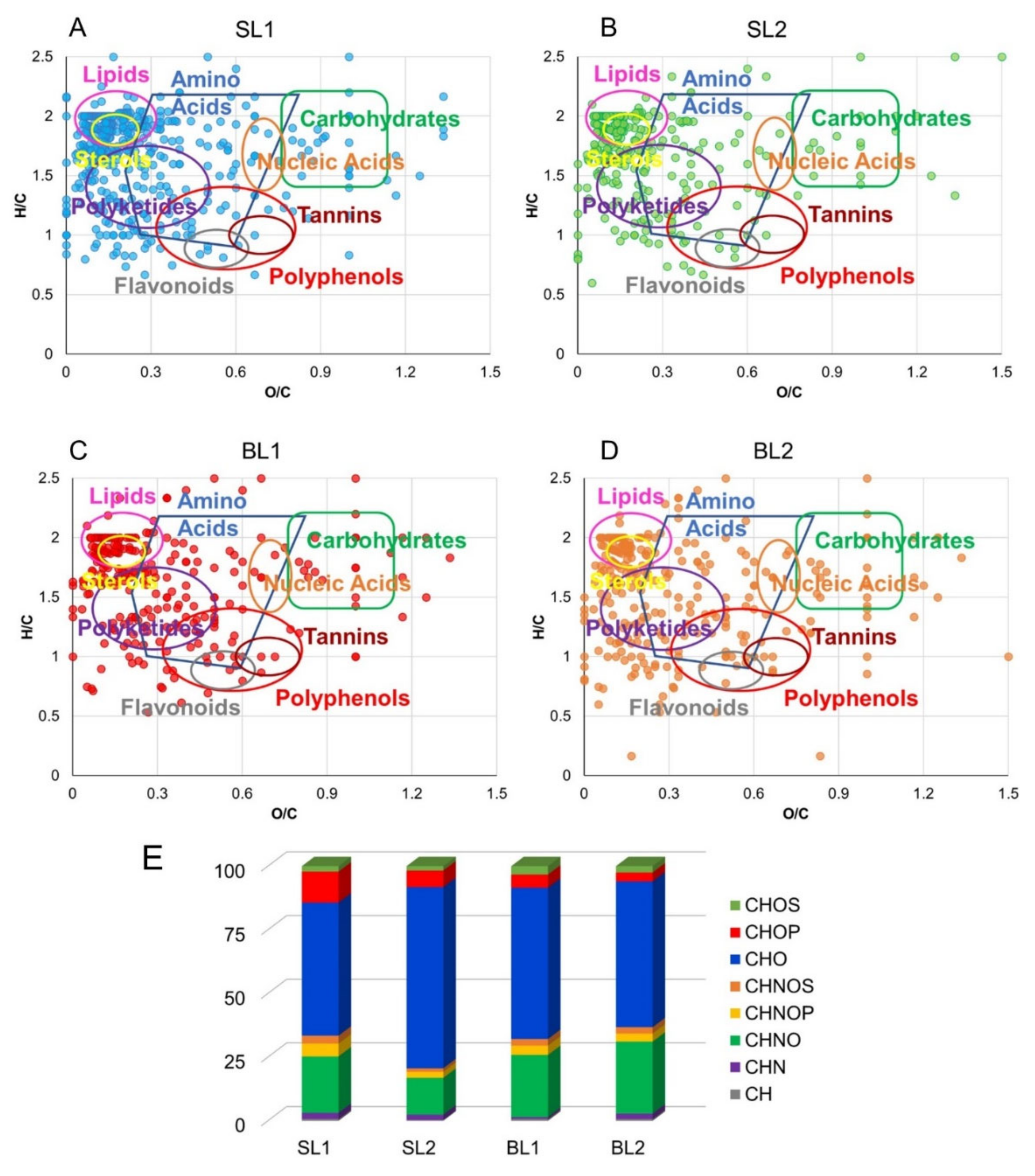
2.1.2. ESI FT-ICR Assignment
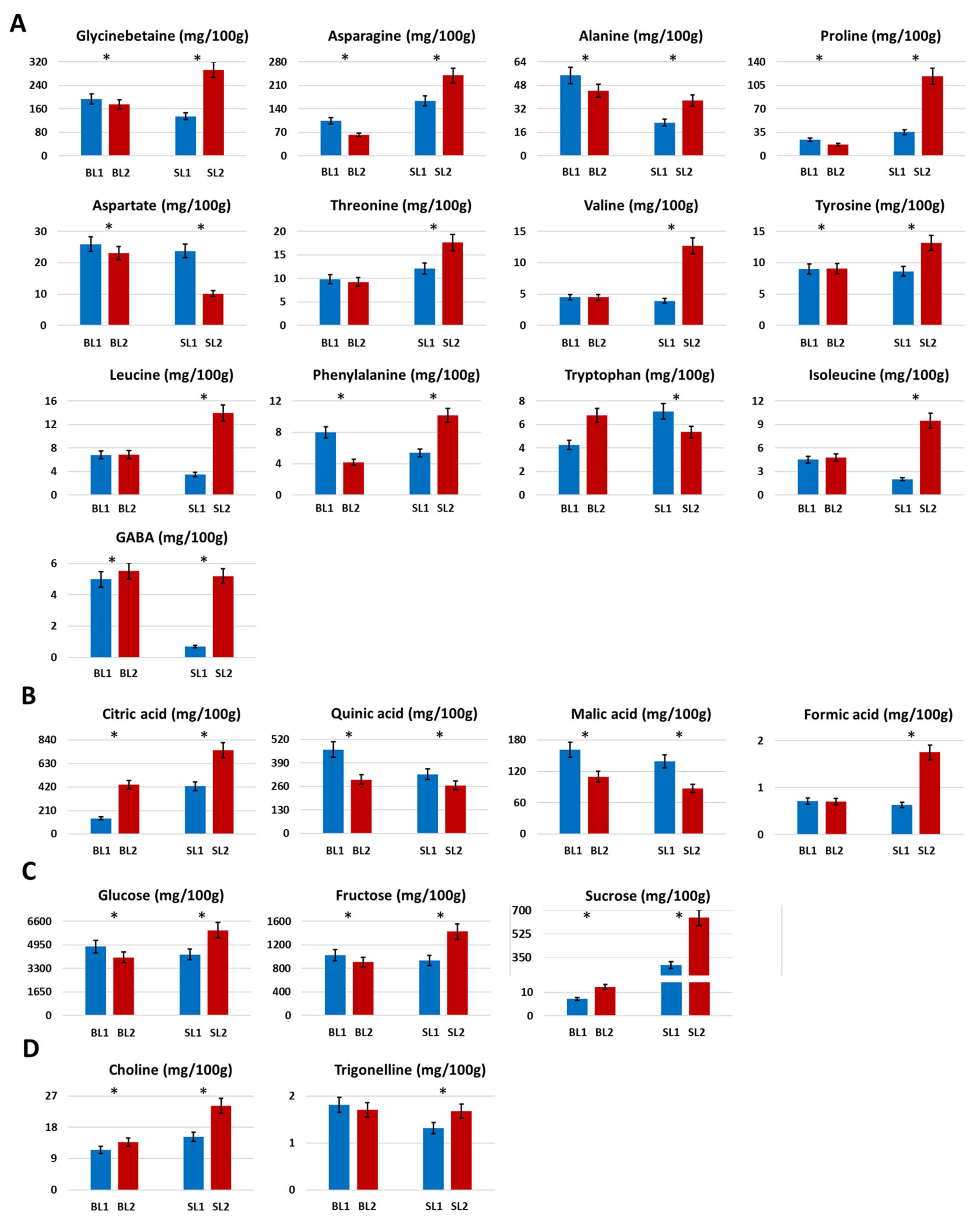
2.1.3. Amino Acids and Derivatives
2.1.4. Organic Acids
2.1.5. Sugars
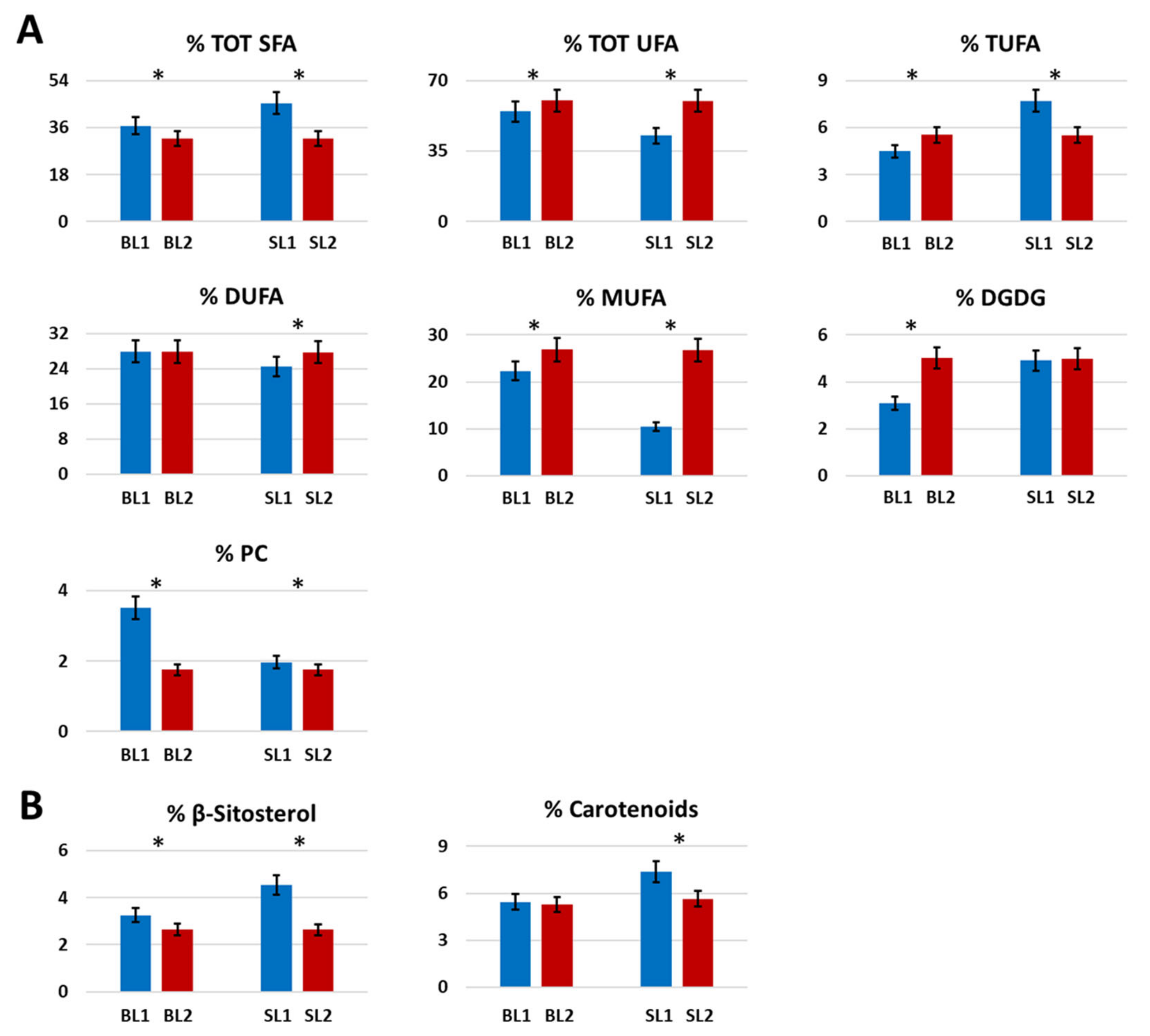
2.1.6. Fatty Acids and Polar Lipids
2.1.7. Miscellaneous
2.2. Pigments Analysis
2.3. Biological Assays on Berries Extracts and Metabolomic Characterization of Leaves
2.3.1. Effects of BL1 Goji Berries Extracts on MG63 Cell Viability
2.3.2. Effects of Goji Berries Extracts on Inflammatory Mediators in MG63 Cells
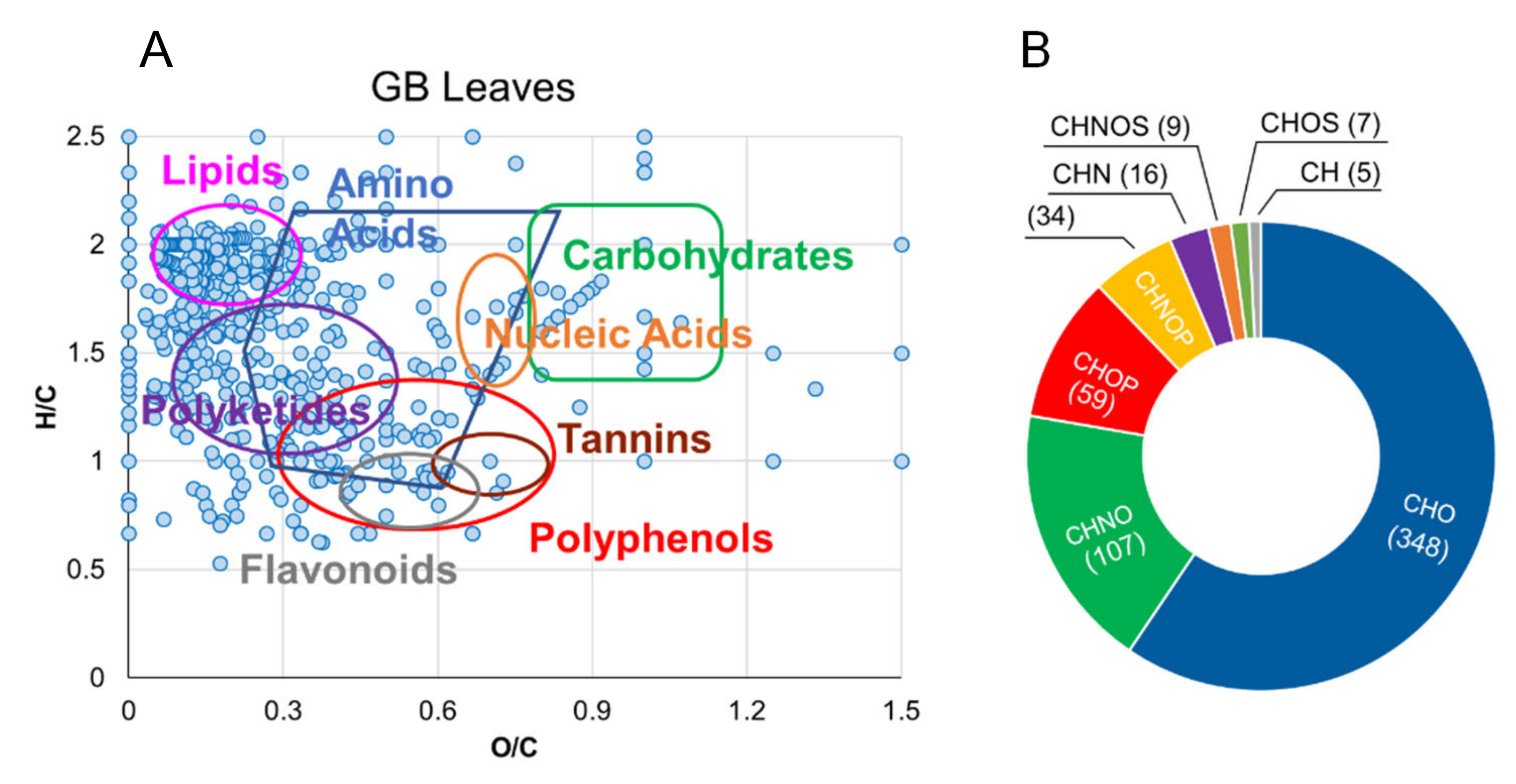
2.3.3. Metabolomic Characterization of Goji Leaves
3. Materials and Methods
3.1. Chemicals
3.2. Plant Material
3.3. Extraction Procedure
3.4. NMR Analysis
3.5. Mass-Spectrometry-Based Metabolomics
3.6. Metabolomics Visualization Tools
3.7. Spectrophotometric Pigments Analysis
3.8. Biological Assays
3.8.1. Cell Viability
3.8.2. Quantitative Real-Time PCR
3.8.3. ELISA Assay
3.8.4. Immunofluorescence Measures
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Yao, R.; Heinrich, M.; Weckerle, C.S. The genus Lycium as food and medicine: A botanical, ethnobotanical and historical review. J. Ethnopharmacol. 2018, 212, 50–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Z.F.; Zhang, H.; Teh, S.S.; Wang, C.W.; Zhang, Y.; Hayford, F.; Wang, L.; Ma, T.; Dong, Z.; Zhang, Y.; et al. Goji berries as a potential natural antioxidant medicine: An insight into their molecular mechanisms of action. Oxid. Med. Cell. Longev. 2019, 2019, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, D.; Zhao, Y.; Yang, G.; Huang, L. Systematic review of chemical constituents in the genus lycium (solanaceae). Molecules 2017, 22, 911. [Google Scholar] [CrossRef] [Green Version]
- Kulczyński, B.; Gramza-Michałowska, A. Goji Berry (Lycium barbarum): Composition and Health Effects—A Review. Polish J. Food Nutr. Sci. 2016, 66, 67–75. [Google Scholar] [CrossRef]
- Wang, C.C.; Chang, S.C.; Inbaraj, B.S.; Chen, B.H. Isolation of carotenoids, flavonoids and polysaccharides from Lycium barbarum L. and evaluation of antioxidant activity. Food Chem. 2010, 120, 184–192. [Google Scholar] [CrossRef]
- Inbaraj, B.S.; Lu, H.; Kao, T.H.; Chen, B.H. Simultaneous determination of phenolic acids and flavonoids in Lycium barbarum Linnaeus by HPLC-DAD-ESI-MS. J. Pharm. Biomed. Anal. 2010, 51, 549–556. [Google Scholar] [CrossRef]
- Masci, A.; Carradori, S.; Casadei, M.A.; Paolicelli, P.; Petralito, S.; Ragno, R.; Cesa, S. Lycium barbarum polysaccharides: Extraction, purification, structural characterisation and evidence about hypoglycaemic and hypolipidaemic effects. A review. Food Chem. 2018, 254, 377–389. [Google Scholar] [CrossRef]
- Zou, S.; Zhang, X.; Yao, W.; Niu, Y.; Gao, X. Structure characterization and hypoglycemic activity of a polysaccharide isolated from the fruit of Lycium barbarum L. Carbohydr. Polym. 2010, 80, 1161–1167. [Google Scholar] [CrossRef]
- Skenderidis, P.; Lampakis, D.; Giavasis, I.; Leontopoulos, S.; Petrotos, K.; Hadjichristodoulou, C.; Tsakalof, A. Chemical properties, fatty-acid composition, and antioxidant activity of Goji berry (Lycium barbarum l. and Lycium Chinense mill.) fruits. Antioxidants 2019, 8, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mocan, A.; Cairone, F.; Locatelli, M.; Cacciagrano, F.; Carradori, S.; Vodnar, D.C.; Crișan, G.; Simonetti, G.; Cesa, S. Polyphenols from lycium barbarum (Goji) fruit european cultivars at different maturation steps: Extraction, hplc-dad analyses, and biological evaluation. Antioxidants 2019, 8, 562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertoldi, D.; Cossignani, L.; Blasi, F.; Perini, M.; Barbero, A.; Pianezze, S.; Montesano, D. Characterisation and geographical traceability of Italian goji berries. Food Chem. 2019, 275, 585–593. [Google Scholar] [CrossRef]
- Dhekney, S.A.; Baldwin, M.R. Prospects for goji berry (Lycium barbarum L.) production in North America. In Medicinal Plants: From Farm to Pharmacy; SpringerLink: Berlin, Germany, 2019; pp. 129–140. ISBN 9783030312695. [Google Scholar]
- Bondia-Pons, I.; Savolainen, O.; Törrönen, R.; Martinez, J.A.; Poutanen, K.; Hanhineva, K. Metabolic profiling of Goji berry extracts for discrimination of geographical origin by non-targeted liquid chromatography coupled to quadrupole time-of-flight mass spectrometry. Food Res. Int. 2014, 63, 132–138. [Google Scholar] [CrossRef]
- Mocan, A.; Vlase, L.; Vodnar, D.C.; Bischin, C.; Hanganu, D.; Gheldiu, A.M.; Oprean, R.; Silaghi-Dumitrescu, R.; Crişan, G. Polyphenolic content, antioxidant and antimicrobial activities of Lycium barbarum L. And Lycium chinense Mill. Leaves. Molecules 2014, 19, 10056–10073. [Google Scholar] [CrossRef]
- Vitanza, L.; Maccelli, A.; Marazzato, M.; Scazzocchio, F.; Comanducci, A.; Fornarini, S.; Crestoni, M.E.; Filippi, A.; Fraschetti, C.; Rinaldi, F.; et al. Satureja montana L. essential oil and its antimicrobial activity alone or in combination with gentamicin. Microb. Pathog. 2019, 126, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Sobolev, A.P.; Mannina, L.; Capitani, D.; Sanzò, G.; Ingallina, C.; Botta, B.; Fornarini, S.; Crestoni, M.E.; Chiavarino, B.; Carradori, S.; et al. A multi-methodological approach in the study of Italian PDO “Cornetto di Pontecorvo” red sweet pepper. Food Chem. 2018, 255, 120–131. [Google Scholar] [CrossRef]
- Ingallina, C.; Maccelli, A.; Spano, M.; Matteo, G.D.; Sotto, A.D.; Giusti, A.M.; Vinci, G.; Giacomo, S.D.; Rapa, M.; Ciano, S.; et al. Chemico-biological characterization of torpedino di fondi® tomato fruits: A comparison with san marzano cultivar at two ripeness stages. Antioxidants 2020, 9, 1027. [Google Scholar] [CrossRef] [PubMed]
- Ingallina, C.; Spano, M.; Sobolev, A.P.; Esposito, C.; Santarcangelo, C.; Baldi, A.; Daglia, M.; Mannina, L. Characterization of Local Products for Their Industrial Use: The Case of Italian Potato Cultivars Analyzed by Untargeted and Targeted Methodologies. Foods 2020, 9, 1216. [Google Scholar] [CrossRef]
- Liu, H.; Fan, Y.; Wang, W.; Liu, N.; Zhang, H.; Zhu, Z.; Liu, A. Polysaccharides from Lycium barbarum leaves: Isolation, characterization and splenocyte proliferation activity. Int. J. Biol. Macromol. 2012, 51, 417–422. [Google Scholar] [CrossRef]
- Mocan, A.; Zengin, G.; Simirgiotis, M.; Schafberg, M.; Mollica, A.; Vodnar, D.C.; Crişan, G.; Rohn, S. Functional constituents of wild and cultivated Goji (L. barbarum L.) leaves: Phytochemical characterization, biological profile, and computational studies. J. Enzyme Inhib. Med. Chem. 2017, 32, 153–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingallina, C.; Sobolev, A.P.; Circi, S.; Spano, M.; Fraschetti, C.; Filippi, A.; Di Sotto, A.; Di Giacomo, S.; Mazzoccanti, G.; Gasparrini, F.; et al. Cannabis sativa L. inflorescences from monoecious cultivars grown in central Italy: An untargeted chemical characterization from early flowering to ripening. Molecules 2020, 25, 1908. [Google Scholar] [CrossRef] [PubMed]
- Di Matteo, G.; Spano, M.; Esposito, C.; Santarcangelo, C.; Baldi, A.; Daglia, M.; Mannina, L.; Ingallina, C.; Sobolev, A.P. Nmr characterization of ten apple cultivars from the piedmont region. Foods 2021, 10, 289. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kramer, R.W.; Hatcher, P.G. Graphical Method for Analysis of Ultrahigh-Resolution Broadband Mass Spectra of Natural Organic Matter, the Van Krevelen Diagram. Anal. Chem. 2003, 75, 5336–5344. [Google Scholar] [CrossRef]
- Song, Y.; Xu, B. Diffusion profiles of health beneficial components from goji berry (Lyceum barbarum) marinated in alcohol and their antioxidant capacities as affected by alcohol concentration and steeping time. Foods 2013, 2, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Fan, T.W.M. Metabolite profiling by one- and two-dimensional NMR analysis of complex mixtures. Prog. Nucl. Magn. Reson. Spectrosc. 1996, 28, 161–219. [Google Scholar] [CrossRef]
- Donno, D.; Beccaro, G.L.; Mellano, M.G.; Cerutti, A.K.; Bounous, G. Goji berry fruit (Lycium spp.): Antioxidant compound fingerprint and bioactivity evaluation. J. Funct. Foods 2015, 18, 1070–1085. [Google Scholar] [CrossRef]
- Mikulic-Petkovsek, M.; Schmitzer, V.; Slatnar, A.; Stampar, F.; Veberic, R. Composition of Sugars, Organic Acids, and Total Phenolics in 25 Wild or Cultivated Berry Species. J. Food Sci. 2012, 77. [Google Scholar] [CrossRef] [PubMed]
- Rekhter, D.; Lüdke, D.; Ding, Y.; Feussner, K.; Zienkiewicz, K.; Lipka, V.; Wiermer, M.; Zhang, Y.; Feussner, I. Isochorismate-derived biosynthesis of the plant stress hormone salicylic acid. Science 2019, 365, 498–502. [Google Scholar] [CrossRef]
- Kosińska-Cagnazzo, A.; Weber, B.; Chablais, R.; Vouillamoz, J.F.; Molnár, B.; Crovadore, J.; Lefort, F.; Andlauer, W. Bioactive compound profile and antioxidant activity of fruits from six goji cultivars cultivated in Switzerland. J. Berry Res. 2017, 7, 43–59. [Google Scholar] [CrossRef]
- Pedro, A.C.; Sánchez-Mata, M.C.; Pérez-Rodríguez, M.L.; Cámara, M.; López-Colón, J.L.; Bach, F.; Bellettini, M.; Haminiuk, C.W.I. Qualitative and nutritional comparison of goji berry fruits produced in organic and conventional systems. Sci. Hortic. 2019, 257. [Google Scholar] [CrossRef]
- Ilić, T.; Dodevska, M.; Marčetić, M.; Božić, D.; Kodranov, I.; Vidović, B. Chemical Characterization, Antioxidant and Antimicrobial Properties of Goji Berries Cultivated in Serbia. Foods 2020, 9, 1614. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.Z.; Deng, G.; Liang, Q.; Chen, D.F.; Guo, R.; Lai, R.C. Antioxidant activity of quercetin and its glucosides from propolis: A theoretical study. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Maková, B.; Mik, V.; Lišková, B.; Gonzalez, G.; Vítek, D.; Medvedíková, M.; Monfort, B.; Ručilová, V.; Kadlecová, A.; Khirsariya, P.; et al. Cytoprotective activities of kinetin purine isosteres. Bioorganic Med. Chem. 2021, 33. [Google Scholar] [CrossRef] [PubMed]
- Cao, D.; Pi, J.; Shan, Y.; Tang, Y.; Zhou, P. Anti-inflammatory effect of Resolvin D1 on LPS-treated MG-63 cells. Exp. Ther. Med. 2019, 16, 4283–4288. [Google Scholar] [CrossRef] [Green Version]
- Leonidou, A.; Lepetsos, P.; Mintzas, M.; Kenanidis, E.; Macheras, G.; Tzetis, M.; Potoupnis, M.; Tsiridis, E. Inducible nitric oxide synthase as a target for osteoarthritis treatment. Expert Opin. Ther. Targets 2018, 22, 299–318. [Google Scholar] [CrossRef] [PubMed]
- Tu, M.; Yang, M.; Yu, N.; Zhen, G.; Wan, M.; Liu, W.; Ji, B.; Ma, H.; Guo, Q.; Tong, P.; et al. Inhibition of cyclooxygenase-2 activity in subchondral bone modifies a subtype of osteoarthritis. Bone Res. 2019, 7. [Google Scholar] [CrossRef]
- Salvemini, D.; Kim, S.F.; Mollace, V. Reciprocal regulation of the nitric oxide and cyclooxygenase pathway in pathophysiology: Relevance and clinical implications. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 304, 473–487. [Google Scholar] [CrossRef] [Green Version]
- Solymosi, K.; Mysliwa-Kurdziel, B. Chlorophylls and their Derivatives Used in Food Industry and Medicine. Mini-Rev. Med. Chem. 2016, 17. [Google Scholar] [CrossRef] [Green Version]
- Gülçin, I. Antioxidant properties of resveratrol: A structure-activity insight. Innov. Food Sci. Emerg. Technol. 2010, 11, 210–218. [Google Scholar] [CrossRef]
- Taofiq, O.; González-Paramás, A.M.; Barreiro, M.F.; Ferreira, I.C.F.R.; McPhee, D.J. Hydroxycinnamic acids and their derivatives: Cosmeceutical significance, challenges and future perspectives, a review. Molecules 2017, 22, 281. [Google Scholar] [CrossRef] [PubMed]
- BLIGH, E.G.; DYER, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guijas, C.; Montenegro-Burke, J.R.; Domingo-Almenara, X.; Palermo, A.; Warth, B.; Hermann, G.; Koellensperger, G.; Huan, T.; Uritboonthai, W.; Aisporna, A.E.; et al. METLIN: A Technology Platform for Identifying Knowns and Unknowns. Anal. Chem. 2018, 90, 3156–3164. [Google Scholar] [CrossRef] [Green Version]
- Marshall, A.G.; Chen, T. 40 years of Fourier transform ion cyclotron resonance mass spectrometry. Int. J. Mass Spectrom. 2015, 377, 410–420. [Google Scholar] [CrossRef]
- Wägele, B.; Witting, M.; Schmitt-Kopplin, P.; Suhre, K. Masstrix reloaded: Combined analysis and visualization of tran-scriptome and metabolome data. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spano, M.; Di Matteo, G.; Ingallina, C.; Botta, B.; Quaglio, D.; Ghirga, F.; Balducci, S.; Cammarone, S.; Campiglia, E.; Giusti, A.M.; et al. A multimethodological characterization of cannabis sativa l. Inflorescences from seven dioecious cultivars grown in Italy: The effect of different harvesting stages. Molecules 2021, 26, 2912. [Google Scholar] [CrossRef] [PubMed]
- Wellburn, A.R. The Spectral Determination of Chlorophylls a and b, as well as Total Carotenoids, Using Various Solvents with Spectrophotometers of Different Resolution. J. Plant. Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite | Group | 1H (ppm) | Multiplicity [J(Hz)] | 13C (ppm) |
|---|---|---|---|---|
| Sugars | ||||
| α-D-Fructofuranose b,l | C-2 | 105.4 | ||
| CH-3 | 4.12 * | 83.0 | ||
| CH-4 | 4.01 | |||
| CH-5 | 4.07 | 82.5 | ||
| β-D-Fructofuranose b,l | C-2 | 102.9 | ||
| CH-3 | 4.12 * | 76.5 | ||
| CH-4 | 4.12 * | 75.4 | ||
| CH-5 | 3.83 | |||
| β-D-Fructopyranose b,l | C-2 | 99.3 | ||
| CH-3 | 3.80 | 68.6 | ||
| CH-4 | 4.00 | 77.0 | ||
| CH2-6,6′ | 3.72; 4.03 | 64.4 | ||
| α-Glucose b,l | CH-1 | 5.25 * | d [3.8] | 93.2 |
| CH-2 | 3.55 | 72.0 | ||
| CH-3 | 3.72 | 73.3 | ||
| CH-4 | 3.42 | 70.7 | ||
| CH-5 | 3.84 | 72.5 | ||
| CH2-6,6′ | 3.89; 3.78 | 61.8 | ||
| β-Glucose b,l | CH-1 | 4.66 * | d [8.0] | 96.9 |
| CH-2 | 3.26 | 75.1 | ||
| CH-3 | 3.51 | 76.9 | ||
| CH-4 | 3.42 | 70.7 | ||
| CH-5 | 3.49 | 76.8 | ||
| CH2-6,6′ | 3.90; 3.75 | 61.8 | ||
| Sucrose b,l | CH-1 (Glucose) | 5.42 * | d [3.8] | 93.1 |
| CH-2 | 3.57 | 71.8 | ||
| CH-3 | 3.77 | 73.6 | ||
| CH-4 | 3.48 | 70.2 | ||
| CH-5 | 3.85 | 73.5 | ||
| C-2 (Fructose) | 104.8 | |||
| CH-3′ | 4.23 | d [8.8] | 77.4 | |
| CH-4′ | 4.07 | 75.1 | ||
| CH-5′ | 3.90 | 82.4 | ||
| CH2-6′ | 3.83 | 61.2 | ||
| Organic acids | ||||
| Acetic acid b,l | α-CH3 | 1.93 | s | 24.4 |
| COOH | 182.6 | |||
| Citric acid b,l | α,γ-CH | 2.55 * | d [15.4] | 46.6 |
| α’,γ’-CH | 2.68 | d [15.4] | 46.6 | |
| β-C | 76.5 | |||
| 1,5-COOH | 180.1 | |||
| 6-COOH | 183.0 | |||
| Formic acid b,l | HCOOH | 8.47 * | s | |
| Fumaric Acid b,l | α,β-CH=CH | 6.53 | s | |
| Malic acid b,l | α-CH | 4.30 * | dd [9.9; 3.2] | 71.3 |
| β-CH | 2.70 | dd [15.4; 3.2] | 43.5 | |
| β’-CH | 2.39 | dd [15.4; 9.9] | 43.5 | |
| Quinic acid b,l | CH2-2,2′ | 1.88 *; 2.07 | dd [13.5; 10.8]; m | 41.7 |
| CH-3 | 4.16 | |||
| Amino acids | ||||
| Alanine b,l | α-CH | 3.80 | 51.7 | |
| β-CH3 | 1.49 * | d [7.3] | 17.1 | |
| COOH | 177.1 | |||
| Arginine b,l | α-CH | 3.79 | 55.3 | |
| β-CH2 | 1.91 | m | 28.6 | |
| γ-CH | 1.67 | m | 25.1 | |
| γ’-CH | 1.74 | m | 25.1 | |
| δ-CH3 | 3.24 | 41.6 | ||
| Asparagine b,l | α-CH | 4.02 | 52.4 | |
| β,β’-CH2 | 2.89 *; 2.96 | dd [7.3; 16.9] | 35.5 | |
| γ-CONH2 | 175.4 | |||
| Aspartate b,l | α-CH | 3.94 | 53.1 | |
| β,β’-CH2 | 2.70; 2.81 * | dd [3.9; 17.4] | 37.5 | |
| γ-COO- | 179.9 | |||
| GABA b,l | α-CH2 | 2.30 * | t [7.4] | 35.4 |
| β-CH2 | 1.91 | 25.0 | ||
| γ-CH2 | 3.02 | 41.0 | ||
| Glycinebetaine b,l | N(CH3)3+ | 3.27 * | s | 54.4 |
| α-CH2 | 3.90 | 67.5 | ||
| Glutamine b,l | α-CH | 3.79 | 55.4 | |
| β,β’-CH2 | 2.14 | m | 27.5 | |
| γ-CH | 2.46 | m | 31.9 | |
| Isoleucine b,l | β-CH | 1.99 | 37.1 | |
| γ-CH3 | 1.27 | 25.8 | ||
| γ-CH3 | 1.01 * | d [7.1] | 15.7 | |
| δ-CH3 | 0.95 | 12.0 | ||
| Leucine b,l | β-CH2 | 1.74 | 40.8 | |
| γ-CH | 1.71 | |||
| δ-CH3 | 0.97 * | d [6.2] | 23.0 | |
| δ’-CH3 | 0.96 | d [6.2] | 22.0 | |
| Phenylalanine b,l | CH-2,6 | 7.34 | m | 130.3 |
| CH-4 | 7.38 | m | 128.6 | |
| CH-3,5 | 7.43 * | m | 130.2 | |
| Proline b | α-CH | 4.15 | 62.3 | |
| β,β’- CH2 | 2.07, 2.35 | 30.3 | ||
| γ-CH2 | 2.01 * | m | 24.8 | |
| δ,δ’- CH2 | 3.35, 3.42 | 47.2 | ||
| Threonine b,l | α-CH | 3.61 | 61.6 | |
| β-CH | 4.27 | 67.2 | ||
| γ-CH3 | 1.34 * | d [6.6] | 20.4 | |
| Tyrosine b,l | CH-3,5 | 7.20 | d [8.5] | 131.8 |
| CH-2,6 | 6.90 * | d [8.5] | 116.7 | |
| Tryptophan b | CH-4 | 7.27 | m | |
| CH-5 | 7.19 | m | ||
| CH-6 | 7.73 | d [8.0] | 119.5 | |
| CH-7 | 7.54 * | d [8.0] | 113.0 | |
| Valine b,l | α-CH | 3.62 | 61.4 | |
| β-CH | 2.28 | 30.3 | ||
| γ-CH3 | 1.00 | d [7.1] | 17.7 | |
| γ’-CH3 | 1.05 * | d [7.1] | 18.9 | |
| Miscellaneous metabolites | ||||
| Choline b,l | N(CH3)3+ | 3.21 * | s | 54.8 |
| α-CH2 | 3.81 | 68.5 | ||
| Trigonelline b,l | CH-1 | 9.13 * | s | |
| CH-3,5 | 8.84 | |||
| CH-4 | 8.09 |
| Metabolite | Group | 1H (ppm) | Multiplicity [J (Hz)] | 13C (ppm) |
|---|---|---|---|---|
| Oleic fatty chain b,l | COO | 174.5 | ||
| (C18:1 Δ9) | CH2-2 | 2.30 * | 34.6 | |
| CH2-3 | 1.59 | m | 25.4 | |
| CH2-4-7 | 1.30 | m | 29.5 | |
| CH2-8 | 2.03 | m | 27.6 | |
| CH=CH-9,10 | 5.30 * | m | 128.4 | |
| CH2-11 | 2.03 | m | 27.6 | |
| CH2-12-15 | 1.33–1.30 | m | 29.5–29.9 | |
| CH2-16 | 1.29 | m | 29.7 | |
| CH2-17 | 1.24 | m | 32.2 | |
| CH3-18 | 0.84 | t | 14.1 | |
| Linoleic fatty chain b,l | COO | 174.5 | ||
| (C18:2 Δ9,12) | CH2-2 | 2.30 * | 34.6 | |
| CH2-3 | 1.59 | m | 25.4 | |
| CH2-4-7 | 1.32–1.28 | m | 29.5 | |
| CH2-8 | 2.03 | m | 27.6 | |
| CH= 9 | 5.33 * | m | 130.5 | |
| CH= 10 | 5.30 * | m | 128.4 | |
| CH2-11 | 2.75 * | t [6.8] | 25.9 | |
| CH= 12 | 5.30 * | m | 128.4 | |
| CH= 13 | 5.33 * | m | 130.5 | |
| CH2-14 | 2.03 | m | 27.6 | |
| CH2-15 | 1.29 | m | 29.7 | |
| CH2-16 | 1.29 | m | 29.7 | |
| CH2-17 | 1.24 | m | 32.2 | |
| CH3-18 | 0.86 | t | 14.1 | |
| Linolenic fatty chain b,l | COO | 174.5 | ||
| (C18:3 Δ9,12,15) | CH2-2 | 2.30 * | 34.6 | |
| CH2-3 | 1.57 | m | 25.4 | |
| CH2-4-7 | 1.30 | m | 29.5 | |
| CH2-8 | 2.04 | m | 27.5 | |
| CH=CH 9,10 | 5.33 * | m | 128.4 | |
| CH2 11 | 2.79 * | t [6.2] | 25.9 | |
| CH=CH 12,13 | 5.33 * | m | 128.4 | |
| CH2-14 | 2.79 * | t [6.2] | 25.9 | |
| CH= 15 | 5.28 * | m | 128.6 | |
| CH= 16 | 5.35 * | m | 132.2 | |
| CH2-17 | 2.04 | m | 20.6 | |
| CH3-18 | 0.95 | t [7.5] | 14.3 | |
| Saturated fatty acids b,l | COO | 174.5 | ||
| CH2-2 | 2.28 * | 34.8 | ||
| CH2-3 | 1.57 | m | 25.4 | |
| CH2 | 1.28–1.22 | m | 29.9–32.0 | |
| CH2 n-1 | 1.25 | 23.0 | ||
| CH3 n | 0.84 | t | 14.1 | |
| β-Sitosterol b,l | C-13 | 42.4 | ||
| CH-17 | 1.16 | 56.4 | ||
| CH3-18 | 0.66 * | s | 12.0 | |
| Squalene b,l | CH3-a | 1.56 | ||
| CH3-b | 1.65 | |||
| CH-c | 5.08 | m | 124.7 | |
| CH2-d | 2.04 | |||
| CH2-e | 1.96 | |||
| 1,2-Diacyl-sn-glycero-3-Phosphatidylcholine b,l | CH2-sn1 | 4.30, 4.14 | 62.5 | |
| CH-sn2 | 5.22 | 70.7 | ||
| CH2-sn3 | 3.94, 3.70 | 68.2 | ||
| N(CH3)3+ | 3.19 * | s | 54.4 | |
| Digalactosyldiacylglycerol b,l | CH2-sn1 | 4.30, 4.14 | 62.5 | |
| CH-sn2 | 5.06 | 70.7 | ||
| CH2-sn3 | 3.66 | 68.2 | ||
| CH’’-1 | 4.87 * | d [3.8] | 99.7 | |
| CH’’-2 | 3.78 | 68.5 | ||
| CH’’-3,5 | 3.70 | 70.6 | ||
| CH’’-4 | 3.92 | 70.1 | ||
| CH’-1 | 4.20 | 104.3 | ||
| CH’-2 | 3.50 | 71.3 | ||
| CH’-3 | 3.47 | 73.5 | ||
| CH’-4 | 3.87 | 66.6 | ||
| Carotenoids b | CH-7 | 6.09 | 125.6 | |
| CH-8 | 6.09 | 139.2 | ||
| CH-10 | 6.14 | 131.7 | ||
| CH-11 | 6.63 | 125.2 | ||
| CH-12 | 6.35 * | 138.0 | ||
| CH-14 | 6.24 | 133.0 | ||
| CH-15 | 6.62 | 130.4 |
| Extract | Hydroalcoholic a | Organic b | ||||
|---|---|---|---|---|---|---|
| Ion Mode | Identified MF | Ion Mode | Identified MF | |||
| BL1 | ESI(+) | 96 | 191 | ESI(+) | 55 | 109 |
| ESI(−) | 114 | ESI(−) | 65 | |||
| BL2 | ESI(+) | 118 | 219 | ESI(+) | 71 | 104 |
| ESI(−) | 121 | ESI(−) | 38 | |||
| SL1 | ESI(+) | 165 | 273 | ESI(+) | 114 | 169 |
| ESI(−) | 114 | ESI(−) | 63 | |||
| SL2 | ESI(+) | 93 | 138 | ESI(+) | 101 | 170 |
| ESI(−) | 57 | ESI(−) | 84 | |||
| Chlorophyll a | Chlorophyll b | Tot Carotenoids | ||
|---|---|---|---|---|
| BL1 | Pulp | 0.0112 ± 0.0011 | 0.0214 ± 0.0012 | 1.4210 ± 0.0452 |
| Peel | 0.0043 ± 0.0008 | 0.0083 ± 0.0002 | 0.5300 ± 0.0340 | |
| BL2 | Pulp | 0.0195 ± 0.0015 | 0.0320 ± 0.0018 | 1.1980 ± 0.0420 |
| Peel | 0.0139 ± 0.0018 | 0.0281 ± 0.0014 | 0.3274 ± 0.0285 | |
| SL1 | Pulp | 0.0300 ± 0.0080 | 0.0589 ± 0.0035 | 2.4130 ± 0.0490 |
| Peel | 0.0157 ± 0.0023 | 0.0302 ± 0.0070 | 0.9822 ± 0.0130 | |
| SL2 | Pulp | 0.0184 ± 0.0026 | 0.0370 ± 0.0052 | 1.2810 ± 0.0772 |
| Peel | 0.0046 ± 0.0003 | 0.0090 ± 0.0021 | 0.4652 ± 0.0084 |
| Metabolite | mg/100 g | Metabolite | mg/100 g | Metabolite | mg/100 g |
|---|---|---|---|---|---|
| Valine | 3.98 ± 0.40 (4.5) | Citric acid | 52.67 ± 4.82 (139.9) | Glucose | 356.66 ± 4.20 (4829.9) |
| Isoleucine | 2.57 ± 0.31 (4.5) | Aspartate | 27.38 ± 1.84 (39.4) | Sucrose | 430.13 ± 4.32 (7.3) |
| Leucine | 6.33 ± 0.56 (6.8) | Asparagine | 5.57 ± 0.36 (103.9) | Tyrosine | 4.49 ± 0.54 (9.0) |
| Threonine | 5.30 ± 0.47 (9.8) | Choline | 10.44 ± 1.40 (11.4) | Phenylalanine | 3.81 ± 0.57 (8.0) |
| Alanine | 3.84 ± 0.43 (54.7) | Glycinbetaine | 110.09 ± 8.23 (194) | Formic acid | 1.57 ± 0.09 (0.7) |
| Quinic acid | 82.32 ± 2.31 (60.4) | Fructose | 67.42 ± 5.42 (1025.5) | Trigonelline | 7.60 ± 0.82 (1.8) |
| GABA | 24.87 ± 3.78 (5.0) | Malic acid | 306.02 ± 25.48 (161.7) |
| Metabolite | Molar % |
|---|---|
| TOT SFA | 39.57 ± 3.21 |
| TOT UFA | 57.67 ± 4.83 |
| TUFA | 48.60 ± 4.65 |
| DUFA | 9.07 ± 1.03 |
| MUFA | / |
| DGDG | 17.70 ± 1.70 |
| PC | 5.02 ± 0.65 |
| β-Sitosterol | 6.80 ± 0.70 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spano, M.; Maccelli, A.; Di Matteo, G.; Ingallina, C.; Biava, M.; Crestoni, M.E.; Bardaud, J.-X.; Giusti, A.M.; Mariano, A.; Scotto D’Abusco, A.; et al. Metabolomic Profiling of Fresh Goji (Lycium barbarum L.) Berries from Two Cultivars Grown in Central Italy: A Multi-Methodological Approach. Molecules 2021, 26, 5412. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26175412
Spano M, Maccelli A, Di Matteo G, Ingallina C, Biava M, Crestoni ME, Bardaud J-X, Giusti AM, Mariano A, Scotto D’Abusco A, et al. Metabolomic Profiling of Fresh Goji (Lycium barbarum L.) Berries from Two Cultivars Grown in Central Italy: A Multi-Methodological Approach. Molecules. 2021; 26(17):5412. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26175412
Chicago/Turabian StyleSpano, Mattia, Alessandro Maccelli, Giacomo Di Matteo, Cinzia Ingallina, Mariangela Biava, Maria Elisa Crestoni, Jean-Xavier Bardaud, Anna Maria Giusti, Alessia Mariano, Anna Scotto D’Abusco, and et al. 2021. "Metabolomic Profiling of Fresh Goji (Lycium barbarum L.) Berries from Two Cultivars Grown in Central Italy: A Multi-Methodological Approach" Molecules 26, no. 17: 5412. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26175412