
New Scabimycins A-C Isolated from Streptomyces acidiscabies (Lu19992)

1
Department of Pharmaceutical Biotechnology, Saarland University, 66123 Saarbrücken, Germany
2
Department of Pharmaceutical Biology, Saarland University, 66123 Saarbrücken, Germany
3
AMEG Department, Helmholtz Institute for Pharmaceutical Research Saarland, 66123 Saarbrücken, Germany
*
Author to whom correspondence should be addressed.
Molecules 2021, 26(19), 5922; https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26195922
Submission received: 28 July 2021
/
Revised: 24 September 2021
/
Accepted: 26 September 2021
/
Published: 29 September 2021
(This article belongs to the Special Issue Microbial Natural Products in Drug Discovery Chemistry)
Abstract
:Peptide natural products displaying a wide range of biological activities have become important drug candidates over the years. Microorganisms have been a powerful source of such bioactive peptides, and Streptomyces have yielded many novel natural products thus far. In an effort to uncover such new, meaningful compounds, the metabolome of Streptomyces acidiscabies was analyzed thoroughly. Three new compounds, scabimycins A–C (1–3), were discovered, and their chemical structures were elucidated by NMR spectroscopy. The relative and absolute configurations were determined using ROESY NMR experiments and advanced Marfey’s method.
1. Introduction
The identification of valuable agents is currently more urgent than ever. The rising drug resistance among disease-causing pathogens is only one reason to drive natural product research forward [1]. Agents for treating cancer, bacterial and viral infections or even neurodegenerative diseases require a source of biologically active compounds, which can lead to new treatment options [2,3,4]. In recent years a subgroup called antimicrobial peptides (AMPs) became a new focus of researchers worldwide due to the often observed enhanced activity against resistant pathogens and the potentially lower risk of resistance development [5]. AMPs are small peptides that consist of 2–15 amino acids (short AMPs) up to 150 amino acids and were found throughout most species (mammalians, insects, amphibians, microorganisms) [6,7,8]. Their differences from conventional antibiotics in terms of pharmacodynamics and their unspecific mode of action, which usually results in slower resistance development, make them a potential alternative treatment option. It was indeed shown over the years that several AMPs or combinations of AMPs/antibiotics can effectively treat multi-drug resistant infections and were thus further developed for clinical trials [9,10,11]. Of course, meanwhile, cases of resistance development are also observed with AMPs, although mechanistically different from antibiotics, which gives hope to have them as back-up when other treatments fail [12]. Through thorough studying of pharmacodynamics or dose-response relationships of AMPs, rapid development of AMP resistant strains can be avoided, which should be of major interest when studying new drug candidates. Yu et al. (2018) and Rodríguez-Rojas (2021) et al. were able to confirm that AMPs have a much steeper pharmacodynamic curve, meaning a higher killing rate in a shorter time frame, and, on the other hand, a rapid increase in concentration while administering AMPs avoids the upcoming of non-inherited resistance caused by tolerant or persistent subpopulations [13,14].
Many AMPs have been isolated from bacteria, e.g., the very first described tyrocidins or gramicidins isolated from Bacillus brevis [15] or the well-established nisins [16], and the detection of many more is ongoing. In bacteria, peptides are either ribosomally synthesized and post-translationally modified (RiPPs) or synthesized by large multi-functional enzymes called nonribosomal peptide synthetases (NRPSs) [17,18]. These enzymatic machineries incorporate not only the 20 proteinogenic amino acids, but also many non conventionl amino acids, which causes a huge diversity within peptide structures. Up to 500 different non-proteinogenic amino acid building blocks have been identified so far, including the so-called α,β-dehydroamino acids (dhAAs) [19]. These dhAAs bear a double bond between the α- and the β-carbon of an amino acid and have been often found to be crucial for the activity of a peptide. The physico-chemical properties of dhAAs influence the overall peptide conformation and usually cause the structure to be more rigid, which in turn can have an impact on interaction with targets and thus on the potent activity of certain compounds [20]. Moreover, due to their α,β-double bond, these particular amino acids are more prone to chemical reactions, such as E/Z isomerization, hydrogenation or Michael addition, which is also often the reason why this amino acid contributes significantly to the bioactivity of some compounds [21,22,23].
Herein, we report the isolation and identification of three new peptide compounds that were found in Streptomyces acidiscabies (Lu19992) while screening novel actinomycetes strains. Structural elucidation performed using 1D and 2D NMR spectroscopy revealed that these three compounds are linear peptides consisting of acetyl or lactyl starting units and 4–6 amino acids, including proline, alanine, leucine, allo-isoleucine and the α,β-dehydroamino acid dehydrobutyrine (dhb). The relative configurations of the dhb moieties were determined using ROESY NMR experiments. Furthermore, the absolute configuration of amino acids was assigned by applying the advanced Marfey’s method. Referring to their origin, those compounds were named scabimycins A–C (1–3).
2. Results
Isolation and Structural Elucidation of Scabimycins A–C
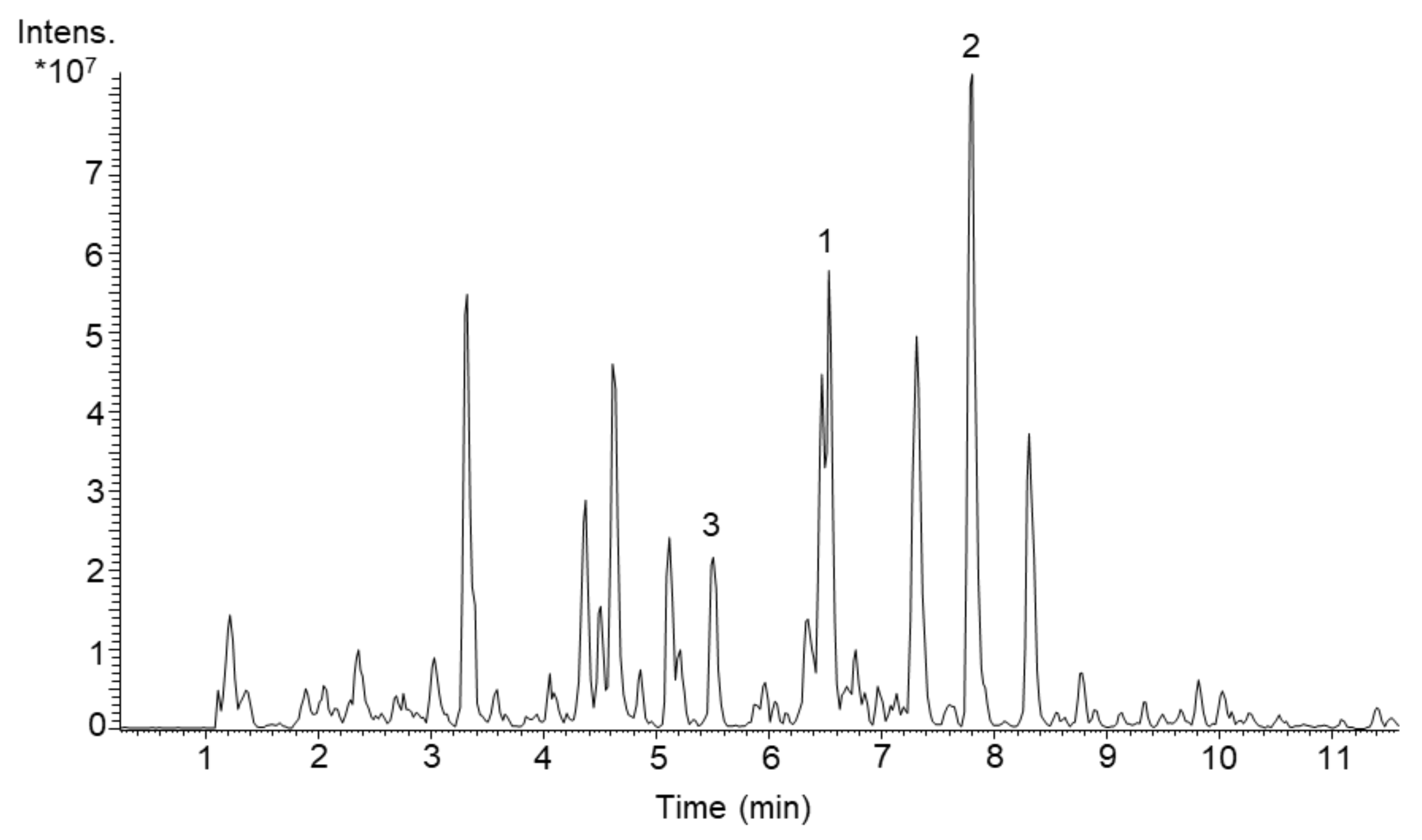
With the primary goal to discover yet unknown compounds in complex bacterial extracts and subsequent structural identification and confirmation of their novelty, we screened several actinomycetes strains from our library. By comparing exact masses and UV spectra of produced metabolites with the natural product database “Dictionary of Natural Products” (DNP) [24], we evaluated the novelty of each produced compound. In the course of this, the strain Streptomyces acidiscabies Lu19992 caught our attention due to the presence of three unknown metabolites (Figure 1). For further characterization of these compounds, we aimed to isolate them and perform structural elucidation. Therefore, S. acidiscabies was cultivated in 10 L DNPM medium (production medium consisting of dextrin, bacto soytone, yeast extract and MOPS (4-morpholinepropanesulfonic acid)). The three new compounds 1–3 were isolated by extracting the culture broth with butanol, and subsequently purified using flash chromatography, size-exclusion chromatography and as last step semi-preparative HPLC.
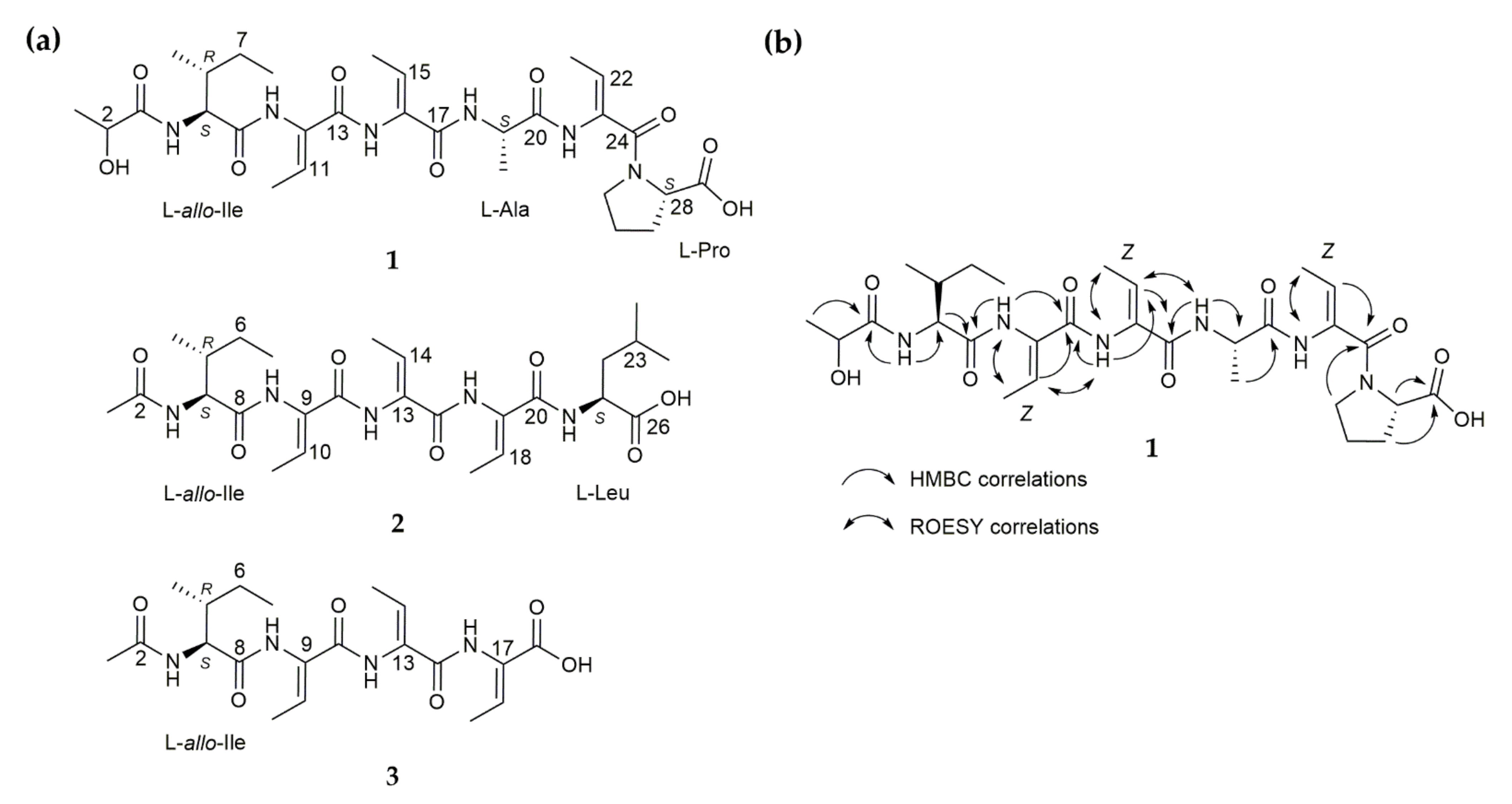
Scabimycin A (1) was isolated as a light-yellow solid with UV absorbance at 219 nm and a molecular ion peak at m/z 621.3229 [M + H]+ determined by high-resolution electrospray ionization mass spectrometry (HRESIMS) (Figure S1). The molecular formula C29H44N6O9, determined from the exact mass, indicates 11 degrees of unsaturation. The 1H NMR spectra acquired in DMSO-d6 revealed the presence of five protons attached to nitrogen (δH 9.75, 9.20, 9.06, 7.66, 7.63), eight methine protons (among them, one secondary alcohol and three as part of double bonds; δH 6.47, 6.17, 5.52, 4.36, 4.33, 4.18, 3.98, 1.83), four methylenes (δH 3.60–3.40, 1.84–1.72, 2.14–1.77, 1.46–1.06) and seven methyl groups (δH 1.70, 2 × 1.64, 1.30, 1.19, 0.88, 0.83). Analysis of HSQC and HMBC spectra (no 13C spectra were acquired) additionally revealed ten quaternary carbons (seven carbonyls and three double bond carbons). The presence of several NH groups and carbonyls and the typical proton/carbon chemical shifts of amino acid α-CH groups (δH/C 4.18/58.5, 4.36/48.1, 4.33/55.9) suggest a peptide-like structure consisting of at least three natural amino acids, proline, alanine and isoleucine (Table S1 and Figures S4–S10). The cross peak of methine proton at δH 6.17 and methyl group at δH 1.70 in COSY spectrum and their HMBC correlations to the quaternary carbons C-10 (δC 130.7) and C-13 (δC 163.5) indicate an α,β-unsaturated amino acid, called dehydrobutyrine (Dhb), of which two additional units were recognized in the NMR spectra. Dehydrobutyrine can be present in the Z or E configuration; however, it is often found as the Z isomer. ROESY correlations indicate the expected Z-configuration for all three dhbs (Figure 2 and Figure S6). Further ROESY cross peaks between 3-NH/H-6 and rather weak signals between the 3-NH/H-5 and 3-NH/H-7 groups of isoleucine (Figure S6) lead to the conclusion that the allo-version of isoleucine is supposedly present. Finally, a lactyl moiety (Lac) could be recognized by its characteristic chemical shifts (δC 174.5, δH 3.98/δC 66.9, δH 1.19/δC 20.6). Careful interpretation of the HMBC correlations led to the peptide sequence Lac-allo-Ile-Dhb-Dhb-Ala-Dhb-Pro-OH (Figure 2).
The absolute configuration of scabimycin A was determined using advanced Marfey’s method [25,26]. The compound was hydrolysed and derivatized with D- and L-FDLA and subsequently subjected to LC-HRMS measurements. After careful analysis of retention times and comparison with standards, allo-isoleucine, alanine and proline were found to have a L-configuration (Table 1, Figure S19). Under the chosen HPLC conditions of Marfey´s method, the L- and D-forms of isoleucine and allo-isoleucine are not clearly distinguishable. Therefore, in addition to the ROESY measurement, we examined the acidic Marfey hydrolysis product by means of 1H NMR and compared the spectrum with those of pure L-isoleucine and L-allo-isoleucine (Figures S21 and S22). Comparison of the relevant α-protons led without any doubt to the detection of L-allo isoleucine.
Scabimycin B (2) was isolated as a pale-yellow solid with UV absorbance at 220 nm and a molecular ion peak with m/z 536.3069 [M + H]+ determined by HRESIMS (Figure S2). The molecular formula C26H41N5O7, determined from the exact mass, indicates 9 degrees of unsaturation. Again, allo-isoleucine could be detected in the 1D and 2D NMR spectra of scabimycin B. However, the Lac moiety was exchanged for an acetyl moiety, which was concluded from the HMBC correlation of H-1 (δH 1.88/δC 22.0) to C-2 (δC 170.8) and the N-HMBC correlation of H-1 to 2-NH (δN 125.0). It was found that the number and type of amino acids in scabimycin B were slightly different from those found in the amino acid sequence of scabimycin A (Table S1 and Figures S11–S14). Alanine was missing; therefore, isoleucine was followed by three dhbs instead of two; this result is supported by the respective HMBC correlations from 16-NH (dhb-3) to C-16 of the second dhb. Proline was replaced by a terminal leucine, which could be identified by its characteristic shifts. The HMBC correlation from 20-NH to carbonyl C-20 indicates a connection to the third dhb (Figure 2). It was assumed that all the double bonds in dhb were in a Z configuration. The absolute configuration was determined using advanced Marfey’s method as described for scabimycin A. Both amino acids, the allo-isoleucine and leucine present in scabimycin B, were found to have an L-configuration (Table 1, Figure S20).
Scabimycin C (3) was isolated as a yellow, oily compound with a molecular ion peak at m/z 423.2227 [M + H]+ determined by HRESIMS (Figure S3); however, a fragment at m/z 322.1755 [M + H]+ predominated. Comparison of the NMR data with the previously identified scabimycin B indicated strong similarities. Only the signals of the C-terminal leucine were missing from the spectrum of scabimycin C; this observation is consistent with the identified mass. Thus, 3 is the leucine-truncated version of 2 (Figure 2). The predominant mass of 322 da in the mass spectrum is identical to a fragment lacking a dhb. It is assumed that the relative and absolute stereochemistry is the same as those of scabimycin A and B.
In conclusion, all three new scabimycin natural products contain the allo-version of isoleucine and in total three units of the non-proteinogenic amino acid dhb. However, in scabimycin B and C, allo-Ile and all three dhbs are connected in series whereas in scabimycin A, the series is interrupted through insertion of alanine between the second and the third dhb. Furthermore, in scabimycin A, proline is inserted as the last amino acid in the sequence, whereas scabimycin B ends with leucine. Scabimycin C does not contain any amino acid other than allo-Ile or dhb. Another peculiarity is that scabimycin A has the different starting unit lactyl instead of the acetyl unit found in scabimycin derivatives B and C. The differences in the amino acid sequence suggest a flexible extender unit choice of employed enzymes during biosynthesis. To explain the differences in the amount and order of amino acids and also the choice of the different starting units, it would be necessary to take a closer look at the underlying biosynthetic machinery of these compounds.
Scabimycin A–C were tested on antimicrobial and antifungal activity. No activity was observed against Gram-positive Bacillus subtilis or Gram-negative Pseudomonas putida and E. coli strains. We also tested the ability of the newly purified compounds to inhibit 3CL protease (SARS-CoV-2). However, none of the scabimycins inhibited the protease activity.
3. Discussion
It is more important than ever to find new natural products to expand the libraries for the development of useful medicines. While screening new actinobacteria strains, a streptomyces strain named Streptomyces acidiscabies Lu19992 attracted our attention due to extensive production of three putatively unknown metabolites. These compounds were isolated and characterized via NMR spectroscopy resulting in three novel short linear peptides that harbor the rather unusual amino acid dhb. This amino acid arises from dehydration of threonine, which occurs either through action of a dehydratase enzyme or via the elimination of phosphorylated residues [27]. Dhbs are often found in ribosomally synthesized and post-translationally modified peptides (RiPPs), e.g., in thiopeptides such as cyclothiazomycin [28] or in depsipeptides such as tumescenamides [29], but also in peptides synthesized by NRPS, e.g., in albopeptin or in the recently discovered malpinins [30,31]. The genome of S. acidiscabies harbors several NRPS and RiPPs biosynthetic gene clusters, which became apparent after genome analysis using the antiSMASH online tool [32]. Detailed analysis of these clusters did not give a clear hint if any of them could be responsible for the production of scabimycin compounds. It would be necessary to perform gene deletion or heterologous expression experiments to identify the correct biosynthetic gene cluster and also to unveil the reasons for the choice of different starting units (lactyl/acetyl) during biosynthesis and the different amino acid sequence of scabimycin A contrary to B and C. Scabimycins were not found to possess antibacterial activity against representative Gram-positive or Gram-negative bacteria tested in our laboratory. To certainly exclude antibacterial activity of these compounds, a wider range of pathogenic bacteria needs to be tested. For example the recently discovered tripeptide albopeptin, which contain also the amino acid dhb, was found to be active against the clinical isolate of vancomycin-resistant Enterococcus faecium K60–39 [30].
Additionally, new compounds should be tested more thoroughly, considering that different conditions while testing antimicrobial activity have a major impact on susceptibility of tested bacteria. As it was nicely summarized in a review by Mercer et al. (2020), if certain AMPs do not perform well under “standard” laboratory test conditions developed for antibiotics, it does not mean that there is not potential activity. It has been proven that AMPs show a lack or attenuation of antimicrobial activity when tested using conditions for conventional antibiotic assays. Various factors can influence antimicrobial activity of AMPs under in vitro testing, e.g., pH, temperature, buffer, nutrient concentrations, charge effects, and growth phase to name a few. Thus, to test the real activity of new drug candidates, standard procedures for antimicrobial susceptibility testing need to be adjusted [33]. For example, it was confirmed that pH regulation within bacterial population can vary significantly due to the presence of persister or viable but non-culturable cells and thus the response to antibiotic treatments [34,35]. Considering new insights into behavior of bacterial subcultures will have an impact on how activity testing can be improved.
The vast application possibilities of short peptides were just recently extensively reviewed by Apostolopoulos et al. (2021) [36]. Referring to this overview, scabimycins could be used alternatively in many different areas. Short peptides are used, for example, in gene delivery systems as alternative targeting moiety that lowers the risk of immunogenicity or as cell penetrating peptides [37,38]. It is also worth noting that (short) peptides have a high impact in the development of vaccines used for the treatment of various infections and cancer [39,40]. Furthermore, peptides are currently also developed to treat neurodegenerative diseases [41]. Considering this wide range of pharmaceutical applications, it becomes clear that peptides will play an important role in the future development of valuable therapeutics. Thus, to ensure that we have a basis to work on, it is important to continue identifying more interesting peptides with various length and types of incorporated amino acids. Bacteria are prolific sources of important peptide molecules, and the continuous screening of interesting new strains can hopefully help to broaden the supply of useful peptides. Herein, we showed that screening of a new streptomyces strain led to the discovery of three new short peptides with an unusual amino acid sequence that can be added to the pool of basic structures for the development of future medicines.
4. Materials and Methods
4.1. Cultivation, Metabolite Extraction and Dereplication
The S. acidiscabies strain was precultivated in a 100 mL flask filled with 10 mL of TSB (tryptic soy broth 30 g/L) at 28 °C and 180 rpm on a rotary shaker for 24 h. For the main culture, 50 mL of DNPM medium (dextrin 40 g/L, bacto soytone 7.5 g/L, yeast extract 5 g/L and MOPS (4-morpholinepropanesulfonic acid) 21 g/L, pH 6.8) was inoculated in a 500 mL flask with 1 mL of preculture and cultivated for 5 days at 28 °C and 180 rpm on a rotary shaker. After cultivation, the biomass and culture liquid were extracted separately with a mixture of acetone/methanol (1:1) and butanol, respectively. The solvent was evaporated until the residue was dry, and then the residue was dissolved in 300 µL MeOH. High-resolution LC-MS data were collected on a Dionex Ultimate 3000 UHPLC system (Thermo Fisher Scientific, Waltham, MA, USA) with stationary phase Acquity UPLC BEH C18, 100 mm × 2.1 mm, 1.7 µm column (Waters Corporation, Milford, MA, USA) using a linear gradient from 5 % [B] (acetonitrile + 0.1 % formic acid) against [A] (ddH2O + 0.1 % formic acid) to 95 % [B] and coupled to a PDA detector operating at 200–600 nm. The coupled LTQ Orbitrap mass spectrometer (Thermo Fisher Scientific, Waltham, MA, USA) was operated at m/z 200–2000. Data were analyzed with Thermo Xcalibur Version 3.0.63 software. Dereplication was performed by comparing the exact masses with the Dictionary of Natural Products database version 10.0 (CRC Press, Boca Raton, FL, USA).
4.2. Isolation, Purification and Structure Elucidation
For isolation and purification of the targeted compounds, S. acidiscabies was cultivated in 10 L of DNPM (100 × 500 mL flasks with 100 mL medium) as described above. After cultivation, the culture liquid was separated from the biomass and extracted twice with butanol. The solvent was removed under reduced pressure, and the residue was dissolved in methanol and centrifuged. The pellet was discarded, and the supernatant was dried, resulting in 16 g of raw material. The crude extract was dissolved in 20 mL methanol and fractionated by flash chromatography (Isolera™ One, Biotage, Uppsala, Sweden) using a Chromabond® Flash RS 330 C18 ec 360 g column (Macherey-Nagel, Düren, Germany). Two runs (10 mL of extract for each) were performed using Milli-Q® (MQ) (Merck Millipore, Burlington, MA, USA) -H2O [A] and methanol [B] as eluents. A linear gradient from 30–80% [B] over 5 column volumes (CV) was applied. Fractions were tested on LC-MS, a Dionex Ultimate 3000 UPLC system using Acquity BEH C18, 50 × 2.1 mm, 1.7 µm dp column (Waters Corporation, Milford, MA, USA) and mobile phase: ddH2O + 0.1% formic acid [A]/acetonitrile + 0.1% formic acid [B], 5–95% [B] over 9 min, at flow rate 0.6 mL/min, coupled to amaZON SL speed mass spectrometer (Bruker, Billerica, MA, USA) with ESI source and mass range m/z 200–2000. Fractions containing the targeted compounds were pooled and further purified with size exclusion chromatography using Sephadex® LH 20 (Sigma Aldrich, Germany) as the stationary phase (50 cm column, filled with a 300 mL volume of Sephadex in methanol) and methanol as the eluent. Fractions were collected every 15 min with a speed of 1–2 drops per second. Every third fraction was analyzed by LC-MS (Bruker amaZon speed, see above). The fractions containing scabimycins were further purified by semipreparative high-performance liquid chromatography (HPLC) using the following equipment: Agilent 1100 and 1260 Series HPLC (Agilent Technologies, Santa Clara, CA, USA) equipped with a Nucleodur C18 HTEC column (250 × 4.6 mm, 5 µm, Macherey-Nagel, Düren, Germany) and a DAD detector operating at 200–600 nm. A linear gradient used solvent [A] MQ-H2O + 0.1% formic acid against solvent [B] acetonitrile + 0.1 % formic acid starting from 35% [B] and increasing to 45% [B] over 20 min with a flow rate of 4.5 mL/min at 45 °C.
NMR spectra were acquired in deuterated dimethyl sulfoxide (DMSO-d6) at 298 K on a Bruker Avance III 700 or 500 MHz spectrometer, both equipped with a 5 mm TXI cryoprobe. The NMR shifts were relative to the position of the residual solvent signal of DMSO-d6 at δ 2.50 1H, or to the solvent itself at δ 39.5 (DMSO-d6) for 13C measurements. NMR data were analysed using Topspin, version 3.5 pl7 (Bruker, Billerica, MA, USA) and Spectrus Processor 2018.2.3 (ACD/Labs, Canada).
Scabimycin A (1). Lac-l-allo-Ile-Dhb-Dhb-l-Ala-Dhb-l-Pro-OH. Yellow oil; 5.0 mg; − 34.4 (c 0.9 mg/mL, MeOH); UV (MeOH) λmax 219 nm; δH (700 MHz, DMSO-d6): 9.75 (s, NH-9), 9.20 (s, NH-20), 9.06 (s, NH-13), 7.66 (d, NH-3), 7.63 (d, NH-17), 6.47 (q, H-15), 6.17 (q, H-11), 5.52 (q, H-22), 4.36 (t, H-18), 4.33 (t, H-4), 4.18 (t, H-28), 3.98 (q, H-2), 3.60–3.40 (m, 2xH-25), 2.14–1.77 (m, 2xH-27), 1.84–1.72 (m, 2xH-26), 1.83 (m, H-5), 1.70 (d, 3xH-12), 2 × 1.64 (d, 3xH-16/3xH-23), 1.46–1.06 (m, 2xH-7), 1.30 (m, 3xH-19), 0.89 (t, 3xH-6), 0.84 (t, H-8); δC (700 MHz, DMSO-d6): 174.5 (C-3), 173.2 (C-29), 2 × 170.9 (C-9, C-20), 165.3 (C-24), 163.6 (C-17), 163.5 (C-13), 131.4 (C-21), 130.7 (C-10), 129.9 (C-14), 129.1 (C-15), 124.8 (C-11), 120.2 (C-22), 66.9 (C-2), 58.5 (C-28), 55.9 (C-4), 48.5 (C-25), 48.1 (C-18), 36.8 (C-5), 28.8 (C-27), 24.8 (C-26), 23.9 (C-7), 20.6 (C-1), 17.0 (C-19), 15.2 (C-6), 2 × 12.4 (C-16, C-12), 11.8 (C-23), 10.9 (C-8); HRESIMS m/z 621.3229 [M + H]+ (calc. for C29H44N6O9 ) (meas. 621.3209 [M + H]+, calc. 621.3247 [M + H]+).
Scabimycin B (2). Ac-l-allo-Ile-Dhb-Dhb-Dhb-l-Leu-OH. Yellow oil; 1.8 mg; − 44.2 (c 2.65 mg/mL, MeOH); UV (MeOH) λmax 220 nm; δH (700 MHz, DMSO-d6): 9.63 (s, NH-8), 9.00 (s, NH-12), 8.67 (s, NH-16), 8.42 (s, NH-2), 7.42 (d, NH-20), 6.49 (q, H-18), 6.43 (q, H-10), 6.32 (q, H-14), 4.18 (m, H-21), 4.06 (m, H-3), 1.88 (s, 3xH-1), 1.77 (m, 3xH-4), 1.69 (d, 3xH-11), 1.68 (d, 3xH-15), 1.62 (m, 3xH-23), 1.61 (d, 3xH-19), 1.60–1.48 (m, 2xH-22); 1.55–1.19 (m, 2xH-6), 0.92 (d, H-5), 0.86 (t, H-7), 0.84 (d, 3xH-24), 0.81 (d, 3xH-25); δC (700 MHz, DMSO-d6): 173.6 (C-26), 171.3 (C-8), 170.8 (C-2), 163.7 (C-12), 163.0 (C-20), 162.6 (C-16), 130.3 (C-13), 2 × 129.8 (C-17/C-9), 129.4 (C-18), 128.3 (C-10), 127.5 (C-14), 58.2 (C-3), 50.5 (C-21), 39.8 (C-22); 34.8 (C-4), 24.5 (C-6), 23.7 (C-23), 22.6 (C-24), 22.0 (C-1), 20.9 (C-25), 15.1 (C-5), 3 × 12.6 (C-11/C-15/C-19), 10.7 (C-7); HRESIMS m/z 536.3069 [M + H]+ (calc. for C26H41N5O7) (meas. 536.3068 [M + H]+, calc. 536.3084 [M + H]+).
Scabimycin C (3). Ac-l-allo-Ile-Dhb-Dhb-Dhb-OH. Yellow oil; 2.0 mg; − 60.7 (c 1.45 mg/mL, MeOH); UV (MeOH) 220 nm; δH (700 MHz, DMSO-d6): 9.61 (s, 8-NH), 8.78 (s, 12-NH), 8.44 (s, 16-NH), 8.23 (d, 2-NH), 6.49 (m, H-14), 6.48 (m, H-18), 6.36 (q, H-10), 4.06 (t, H-3), 1.86 (s, 3xH-1), 1.73 (m, H-4), 1.69 (d, 3xH-11), 2 × 1.63 (d, 3xH-15/3xH-19), 1.52–1.18 (m, 2xH-6), 0.91 (d, H-5), 0.85 (t, 3xH-7); δC (700 MHz, DMSO-d6): 171.7 (C-8), 170.4 (C-2), 163.2 (C-12), 2 × 162.4 (C-20/C-16), 130.7 (C-9), 2 × 130.1 (C-18/C-14), 2 × 129.2 (C-13/C-17), 127.6 (C-10), 58.4 (C-3), 35.5 (C-4), 25.1 (C-6), 22.0 (C-1), 15.4 (C-5), 2 × 13.8 (C-15/C-19), 13.1 (C-11), 11.1 (C-7); HRESIMS m/z 423.2227 [M + H]+ (calc. for C26H41N5O7) (meas. 423.2227 [M + H]+, calc. 423.2243 [M + H]+).
For determination of absolute configuration using advanced Marfey’s method, samples 1 (200 µg) and 2 (250 µg) were treated with 100 µL of a 6 M HCl solution and heated at 110 °C for 45 min. Subsequently, the solvent was removed completely under a nitrogen stream at the same temperature within 20 min. The residues were dissolved in 110 µL H2O, and 50 µL was transferred into two new tubes for each sample. To each tube, 20 µL of 1 M NaHCO3 was added to reach pH 9. For derivatization, the samples were treated with 20 µL of 1% Marfey’s reagent (D-FDLA and L-FDLA, respectively) in acetone. Similarly, 20 µL of 1 M NaHCO3 and 20 µL of a 1% solution of L-FDLA were added to 50 µL of an aqueous solution (50 mmol) of each present amino acid (D/L-proline, D/L-alanine, D/L-leucine, L-isoleucine). All samples were then incubated for 1.5 h at 40 °C and 700 rpm. To stop the reaction, 10 µL of 2 M HCl solution was added. Afterwards, 300 µL acetonitrile was added, and the samples were subjected to LC-HRMS measurements. Samples were run on an Acquity BEH C18, 100 × 2.1 mm, 1.7 µm dp column (Waters Corporation, Milford, MA, USA) using eluent [A] ddH2O + 0.1% formic acid and [B] acetonitrile + 0.1% formic acid. The following gradient was applied: 5–10% [B] (0–2 min), 10–25% [B] (2–15 min), 25–50% [B] (15–22 min), 50–95% [B] (22–24 min), 95 -95% [B] (24–26 min), and 95–5% [B] (26–27 min). The column oven temperature was set to 45 °C, and a mass range from m/z 100–1000 was acquired.
Supplementary Materials
The following are available online: Table S1: NMR spectroscopic data of Scabimycins A-C (1–3) in DMSO-d6, Figure S1: MS spectra of scabimycin A, showing the exact mass m/z 621.3228 [M + H]+, Figure S2: MS spectra of scabimycin B, showing the exact mass m/z 536.3067 [M + H]+, Figure S3: MS spectra of scabimycin C, showing the exact mass without dehydrobutyrine (as explained in the main text, this mass predominates) m/z 322.1753 [M + H]+, Figure S4: 1H NMR spectrum of scabimycin A (DMSO-d6, 700 MHz), Figure S5: 1H-1H-Cosy spectrum of scabimycin A (DMSO-d6, 700 MHz), Figure S6: ROESY spectrum of scabimycin A (DMSO-d6, 700 MHz), Figure S7: HSQC spectrum of scabimycin A (DMSO-d6, 700 MHz), Figure S8: HMBC spectrum of scabimycin A (DMSO-d6, 700 MHz), Figure S9: 15N-HSQC spectrum of scabimycin A (DMSO-d6, 700 MHz), Figure S10: 15N-HMBC spectrum of scabimycin A (DMSO-d6, 700 MHz), Figure S11: 1H NMR spectrum of scabimycin B (DMSO-d6, 700 MHz), Figure S12: 1H-1H-Cosy spectrum of scabimycin B (DMSO-d6, 700 MHz), Figure S13: HSQC spectrum of scabimycin B (DMSO-d6, 700 MHz), Figure S14: HMBC spectrum of scabimycin B (DMSO-d6, 700 MHz), Figure S15: 1H NMR spectrum of scabimycin C (DMSO-d6, 700 MHz), Figure S16: 1H-1H-Cosy spectrum of scabimycin C (DMSO-d6, 700 MHz), Figure S17: HSQC spectrum of scabimycin B (DMSO-d6, 700 MHz), Figure S18: HMBC spectrum of scabimycin C (DMSO-d6, 700 MHz), Figure S19: Section of LC-HRMS chromatogram (retention time 17 to 28 min) of hydrolysed scabimycin A derivatized with D-FDLA and L-FDLA showing single amino acids D/L-alanine (1/2), D/L-proline (3/4) and isoleucine (5) and leucine (1/3). To compare the amino acids present in our probe, pure D/L-amino acids derivatized with L-FDLA (D/L-alanine (1*/2*), D/L-proline (3*/4*) and L-isoleucine (5*)) are shown below. Comparison with the standard amino acids and the later elution of the D-FDLA-derivatized amino acids lead us to the assumption that in all cases, the amino acids possess an L-configuration. It needs to be mentioned that from this test, it is not possible to decide whether L-allo-isoleucine or L-isolelucine is present. The intense peak at RT 19.5 coincides with the mass of the underivatized FDLA, Figure S20: Section of LC-HRMS chromatogram (retention time 17 to 28 min) of hydrolysed scabimycin B derivatized with D-FDLA and L-FDLA showing single amino acids isoleucine (2) and leucine (1/3). To compare the amino acids present in our probe, pure D/L-amino acids derivatized with L-FDLA (D-leucine (1*), L-leucine (3*) and L-isoleucine (2*)) are shown below. Comparison with the standard amino acids and the later elution of the D-FDLA-derivatized amino acids led us to the assumption that in both cases, the amino acids have an L-configuration. It needs to be mentioned that from this test, it is not possible to decide whether L-allo-isoleucine or L-isolelucine is present. The intense peak at RT 19.5 coincides with the mass of the underivatized FLDA, Figure S21: Determination of allo-isoleucine in scabimycin A. Scabimycin A, allo-L-Ile and L-Ile were treated with HCl at 110 °C. 1H-NMR were acquired in DMSO-d6 and compared. The chemical shifts of the α-protons of scabimycin A and allo-L-Ile coincide, while the α-proton of L-Ile is shifted upfield, Figure S22: HSQC spectrum of scabimycin A after treatment with HCl; this spectrum supports the findings for allo-isoleucine. The 13C chemical shift of the α-carbon was found to be approximately 55 ppm, as expected for isoleucine.
Author Contributions
The experiments were designed and evaluated by C.P. and A.L. and all practical work was performed by C.P. NMR experiments were set up, carried out and evaluated by C.P. NMR data was additionally reviewed by J.Z. The manuscript was drafted by C.P. All authors have read and agreed to the published version of the manuscript.
Funding
The project was funded by BASF.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank Helmholtz-Institute for Pharmaceutical Research Saarland, Saarbruecken, Germany (HIPS) for allowing access to their NMR devices. The strain S. acidiscabies Lu19992 was provided by BASF SE Ludwigshafen, Germany. We thank furthermore Birgit Rosenkränzer for performing protease inhibition test (SARS-CoV-2).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Amirkia, V.; Heinrich, M. Natural products and drug discovery: A survey of stakeholders in industry and academia. Front. Pharmacol. 2015, 6, 237. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Tao, G.; Liu, J.; Cai, J.; Huang, Z.; Chen, J.-X. Current Prevention of COVID-19: Natural Products and Herbal Medicine. Front. Pharmacol. 2020, 11, 588508. [Google Scholar] [CrossRef]
- Verma, S.; Twilley, D.; Esmear, T.; Oosthuizen, C.B.; Reid, A.-M.; Nel, M.; Lall, N. Anti-SARS-CoV Natural Products With the Potential to Inhibit SARS-CoV-2 (COVID-19). Front. Pharmacol. 2020, 11, 561334. [Google Scholar] [CrossRef]
- Antonio, A.D.S.; Wiedemann, L.S.M.; Veiga-Junior, V.F. Natural products’ role against COVID-19. RSC Adv. 2020, 10, 23379–23393. [Google Scholar] [CrossRef]
- Lazzaro, B.P.; Zasloff, M.; Rolff, J. Antimicrobial peptides: Application informed by evolution. Science 2020, 368, eaau5480. [Google Scholar] [CrossRef]
- Sharma, K.; Aaghaz, S.; Shenmar, K.; Jain, R. Short Antimicrobial Peptides. Recent Patents Anti-Infect. Drug Discov. 2018, 13, 12–52. [Google Scholar] [CrossRef] [PubMed]
- Huan, Y.; Kong, Q.; Mou, H.; Yi, H. Antimicrobial Peptides: Classification, Design, Application and Research Progress in Multiple Fields. Front. Microbiol. 2020, 11, 582779. [Google Scholar] [CrossRef] [PubMed]
- Clark, S.; Jowitt, T.A.; Harris, L.K.; Knight, C.G.; Dobson, C.B. The lexicon of antimicrobial peptides: A complete set of arginine and tryptophan sequences. Commun. Biol. 2021, 4, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lázár, V.; Martins, A.; Spohn, R.; Daruka, L.; Grézal, G.; Fekete, G.; Számel, M.; Jangir, P.; Kintses, B.; Csörgő, B.; et al. Antibiotic-resistant bacteria show widespread collateral sensitivity to antimicrobial peptides. Nat. Microbiol. 2018, 3, 718–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.-R.; You, D.-G.; Kim, H.K.; Sohn, J.W.; Kim, M.J.; Park, J.K.; Lee, G.Y.; Yoo, Y.D. Romo1-Derived Antimicrobial Peptide Is a New Antimicrobial Agent against Multidrug-Resistant Bacteria in a Murine Model of Sepsis. mBio 2020, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koo, H.B.; Seo, J. Antimicrobial peptides under clinical investigation. Pept. Sci. 2019, 111, 715. [Google Scholar] [CrossRef]
- Rončević, T.; Puizina, J.; Tossi, A. Antimicrobial Peptides as Anti-Infective Agents in Pre-Post-Antibiotic Era? Int. J. Mol. Sci. 2019, 20, 5713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Rojas, A.; Baeder, D.Y.; Johnston, P.; Regoes, R.R.; Rolff, J. Bacteria primed by antimicrobial peptides develop tolerance and persist. PLoS Pathog. 2021, 17, e1009443. [Google Scholar] [CrossRef]
- Yu, G.; Baeder, D.Y.; Regoes, R.R.; Rolff, J. Predicting drug resistance evolution: Insights from antimicrobial peptides and antibiotics. Proc. R. Soc. B Boil. Sci. 2018, 285, 20172687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubos, R.J.; Hobby, G.L.; Dawson, M.H.; Meyer, K.; Chaffee, E. Studies on a bactericidal agent extracted from a soil bacillus. J. Exp. Med. 1939, 70, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Rogers, L.A. The inhibiting effect of Streptococcus lactis on lactobacillus bulgaricus. J. Bacteriol. 1928, 16, 321–325. [Google Scholar] [CrossRef] [Green Version]
- Papagianni, M. Ribosomally synthesized peptides with antimicrobial properties: Biosynthesis, structure, function, and applications. Biotechnol. Adv. 2003, 21, 465–499. [Google Scholar] [CrossRef]
- Finking, R.; Marahiel, M.A. Biosynthesis of Nonribosomal Peptides. Annu. Rev. Microbiol. 2004, 58, 453–488. [Google Scholar] [CrossRef]
- Walsh, C.T.; O’Brien, R.V.; Khosla, C. Nonproteinogenic Amino Acid Building Blocks for Nonribosomal Peptide and Hybrid Polyketide Scaffolds. Angew. Chem. Int. Ed. 2013, 52, 7098–7124. [Google Scholar] [CrossRef] [Green Version]
- Bogart, J.W.; Bowers, A.A. Dehydroamino acids: Chemical multi-tools for late-stage diversification. Org. Biomol. Chem. 2019, 17, 3653–3669. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, J.M.; Chamberlin, A.R. Chemical Synthesis of Natural Product Peptides: Coupling Methods for the Incorporation of Noncoded Amino Acids into Peptides. Chem. Rev. 1997, 97, 2243–2266. [Google Scholar] [CrossRef]
- Jain, R.; Chauhan, V.S. Conformational characteristics of peptides containing α,β-dehydroamino acid residues. Biopolymers 1996, 40, 105–119. [Google Scholar] [CrossRef]
- Siodłak, D. α,β-Dehydroamino acids in naturally occurring peptides. Amino Acids 2015, 47, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, B. Dictionary of Natural Products on DVD: Version 10.0; Chapman & Hall CRC Press: Boca Raton, FL, USA, 2019. [Google Scholar]
- Marfey, P. Determination ofD-amino acids. II. Use of a bifunctional reagent, 1,5-difluoro-2,4-dinitrobenzene. Carlsberg Res. Commun. 1984, 49, 591–596. [Google Scholar] [CrossRef] [Green Version]
- Vijayasarathy, S.; Prasad, P.; Fremlin, L.J.; Ratnayake, R.; Salim, A.A.; Khalil, Z.; Capon, R.J. C3 and 2D C3 Marfey’s Methods for Amino Acid Analysis in Natural Products. J. Nat. Prod. 2016, 79, 421–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, L.H. Dehydroamino acid chemical biology: An example of functional group interconversion on proteins. RSC Chem. Biol. 2020, 1, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yu, Y.; Tang, K.; Liu, W.; He, X.; Huang, X.; Deng, Z. Identification and Analysis of the Biosynthetic Gene Cluster Encoding the Thiopeptide Antibiotic Cyclothiazomycin in Streptomyces hygroscopicus 10–22. Appl. Environ. Microbiol. 2010, 76, 2335–2344. [Google Scholar] [CrossRef] [Green Version]
- Motohashi, K.; Toda, T.; Sue, M.; Furihata, K.; Shizuri, Y.; Matsuo, Y.; Kasai, H.; Shin-Ya, K.; Takagi, M.; Izumikawa, M.; et al. Isolation and structure elucidation of tumescenamides A and B, two peptides produced by Streptomyces tumescens YM23-260. J. Antibiot. 2010, 63, 549–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Fang, Q.; Lu, Z.; Gao, Y.; Trembleau, L.; Ebel, R.; Andersen, J.H.; Philips, C.; Law, S.; Deng, H. Discovery and Biosynthetic Investigation of a New Antibacterial Dehydrated Non-Ribosomal Tripeptide. Angew. Chem. Int. Ed. 2021, 60, 3229–3237. [Google Scholar] [CrossRef] [PubMed]
- Wurlitzer, J.M.; Stanišić, A.; Wasmuth, I.; Jungmann, S.; Fischer, D.; Kries, H.; Gressler, M. Bacterial-Like Nonribosomal Peptide Synthetases Produce Cyclopeptides in the Zygomycetous Fungus Mortierella alpina. Appl. Environ. Microbiol. 2021, 87. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; van Wezel, G.P.; Medema, M.H.; Weber, T. antiSMASH 6.0: Improving cluster detection and comparison capabilities. Nucleic Acids Res. 2021, 49, W29–W35. [Google Scholar] [CrossRef]
- Mercer, D.K.; Torres, M.; Duay, S.S.; Lovie, E.; Simpson, L.; Von Köckritz-Blickwede, M.; De La Fuente-Nunez, C.; O’Neil, D.A.; Angeles-Boza, A.M. Antimicrobial Susceptibility Testing of Antimicrobial Peptides to Better Predict Efficacy. Front. Cell. Infect. Microbiol. 2020, 10, 326. [Google Scholar] [CrossRef] [PubMed]
- Goode, O.; Smith, A.; Zarkan, A.; Cama, J.; Invergo, B.M.; Belgami, D.; Caño-Muñiz, S.; Metz, J.; O’Neill, P.; Jeffries, A.; et al. Persister Escherichia coli Cells Have a Lower Intracellular pH than Susceptible Cells but Maintain Their pH in Response to Antibiotic Treatment. mBio 2021, 12, e0090921. [Google Scholar] [CrossRef]
- Bamford, R.A.; Smith, A.; Metz, J.; Glover, G.; Titball, R.W.; Pagliara, S. Investigating the physiology of viable but non-culturable bacteria by microfluidics and time-lapse microscopy. BMC Biol. 2017, 15, 705. [Google Scholar] [CrossRef]
- Apostolopoulos, V.; Bojarska, J.; Chai, T.-T.; Elnagdy, S.; Kaczmarek, K.; Matsoukas, J.; New, R.; Parang, K.; Lopez, O.P.; Parhiz, H.; et al. A Global Review on Short Peptides: Frontiers and Perspectives. Molecules 2021, 26, 430. [Google Scholar] [CrossRef]
- Vazquez-Lombardi, R.; Phan, T.G.; Zimmermann, C.; Lowe, D.; Jermutus, L.; Christ, D. Challenges and opportunities for non-antibody scaffold drugs. Drug Discov. Today 2015, 20, 1271–1283. [Google Scholar] [CrossRef] [Green Version]
- Guidotti, G.; Brambilla, L.; Rossi, D. Cell-Penetrating Peptides: From Basic Research to Clinics. Trends Pharmacol. Sci. 2017, 38, 406–424. [Google Scholar] [CrossRef] [PubMed]
- Neefjes, J.; Ovaa, H. A peptide’s perspective on antigen presentation to the immune system. Nat. Chem. Biol. 2013, 9, 769–775. [Google Scholar] [CrossRef] [PubMed]
- Thundimadathil, J. Cancer Treatment Using Peptides: Current Therapies and Future Prospects. J. Amino Acids 2012, 2012, 967347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horsley, J.R.; Jovcevski, B.; Wegener, K.L.; Yu, J.; Pukala, T.L.; Abell, A.D. Rationally designed peptide-based inhibitor of Aβ42 fibril formation and toxicity: A potential therapeutic strategy for Alzheimer’s disease. Biochem. J. 2020, 477, 2039–2054. [Google Scholar] [CrossRef]
Figure 1.
LC-MS chromatogram (RT 1–11 min out of overall 20 min analysis) of butanol extract of S. acidiscabies Lu19992 cultivated in DNPM medium. The peaks representing newly identified compounds are marked: (1) Scabimycin A, (2) Scabimycin B, (3) Scabimycin C.
Figure 1.
LC-MS chromatogram (RT 1–11 min out of overall 20 min analysis) of butanol extract of S. acidiscabies Lu19992 cultivated in DNPM medium. The peaks representing newly identified compounds are marked: (1) Scabimycin A, (2) Scabimycin B, (3) Scabimycin C.

Figure 2.
(a) Chemical structures of small peptide NPs scabimycins A (1), B (2) and C (3) isolated from S. acidiscabies (Lu19992); (b) Exemplary for scabimycin A: Key HMBC correlations that were used to assess the peptidic structure and through-space correlations measured by ROESY experiment (Rotational Nuclear Overhauser Effect Spectroscopy) that led to the Z-configuration of all dhbs in scabimycins.
Figure 2.
(a) Chemical structures of small peptide NPs scabimycins A (1), B (2) and C (3) isolated from S. acidiscabies (Lu19992); (b) Exemplary for scabimycin A: Key HMBC correlations that were used to assess the peptidic structure and through-space correlations measured by ROESY experiment (Rotational Nuclear Overhauser Effect Spectroscopy) that led to the Z-configuration of all dhbs in scabimycins.


{kind=link}
{kind=link}
Table 1.
Retention times of amino acids in scabimycins A and B after derivatization with D-FDLA and L-FDLA.
Table 1.
Retention times of amino acids in scabimycins A and B after derivatization with D-FDLA and L-FDLA.
| Scabimycin A | Scabimycin B | ||||
|---|---|---|---|---|---|
| allo-Ile | Ala | Pro | allo-Ile | Leu | |
| D-FDLA | 23.2 min | 20.1 min | 19.8 min | 23.2 min | 23.3 min |
| L-FDLA | 20.7 min | 18.4 min | 18.6 min | 20.7 min | 20.9 min |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Paulus, C.; Zapp, J.; Luzhetskyy, A. New Scabimycins A-C Isolated from Streptomyces acidiscabies (Lu19992). Molecules 2021, 26, 5922. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26195922
AMA Style
Paulus C, Zapp J, Luzhetskyy A. New Scabimycins A-C Isolated from Streptomyces acidiscabies (Lu19992). Molecules. 2021; 26(19):5922. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26195922
Chicago/Turabian StylePaulus, Constanze, Josef Zapp, and Andriy Luzhetskyy. 2021. "New Scabimycins A-C Isolated from Streptomyces acidiscabies (Lu19992)" Molecules 26, no. 19: 5922. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26195922