
Molecular Studies on the Nephroprotective Potential of Celastrus paniculatus against Lead-Acetate-Induced Nephrotoxicity in Experimental Rats: Role of the PI3K/AKT Signaling Pathway

 , ,
, ,
Abstract
:1. Introduction
2. Results
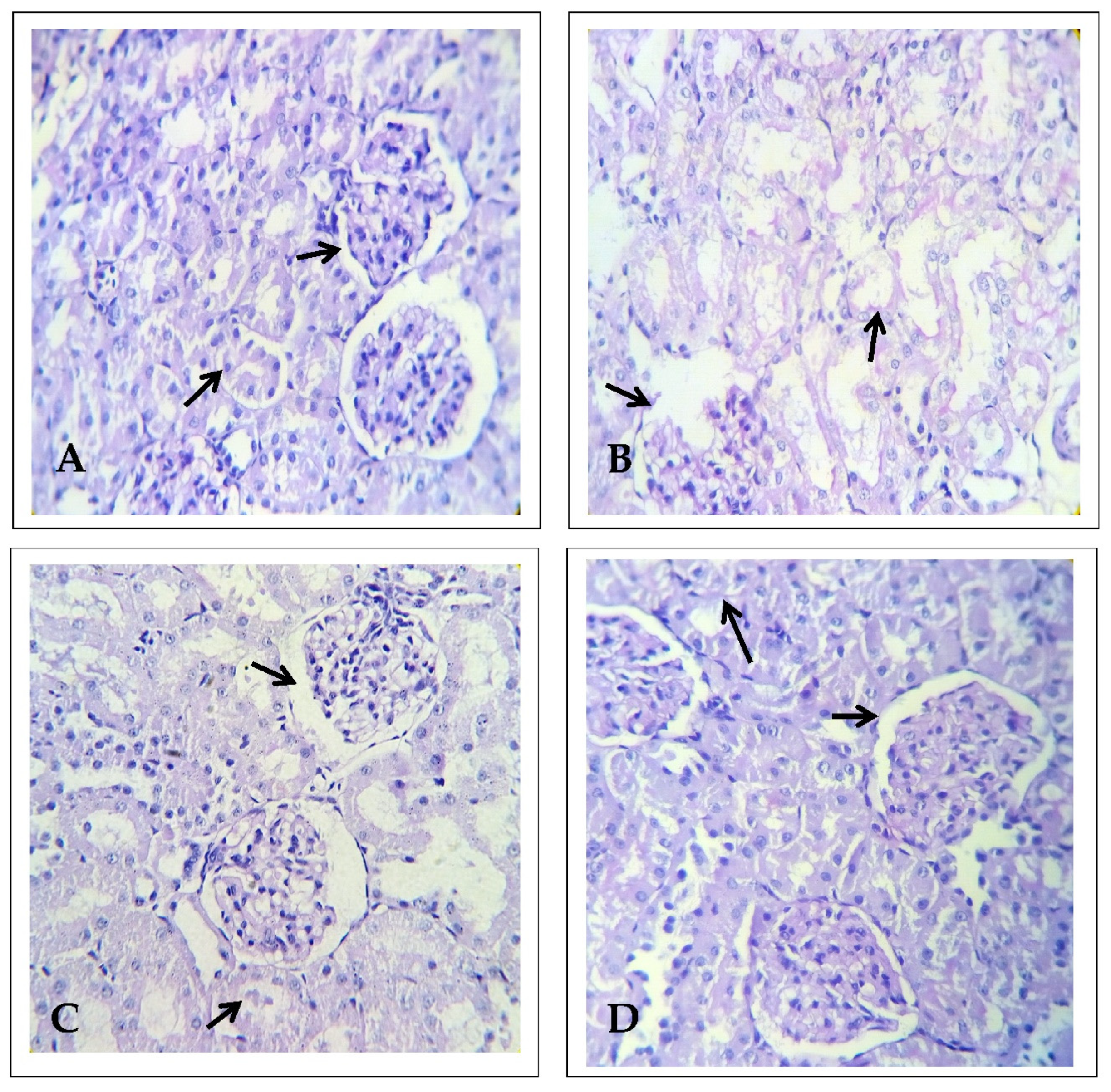
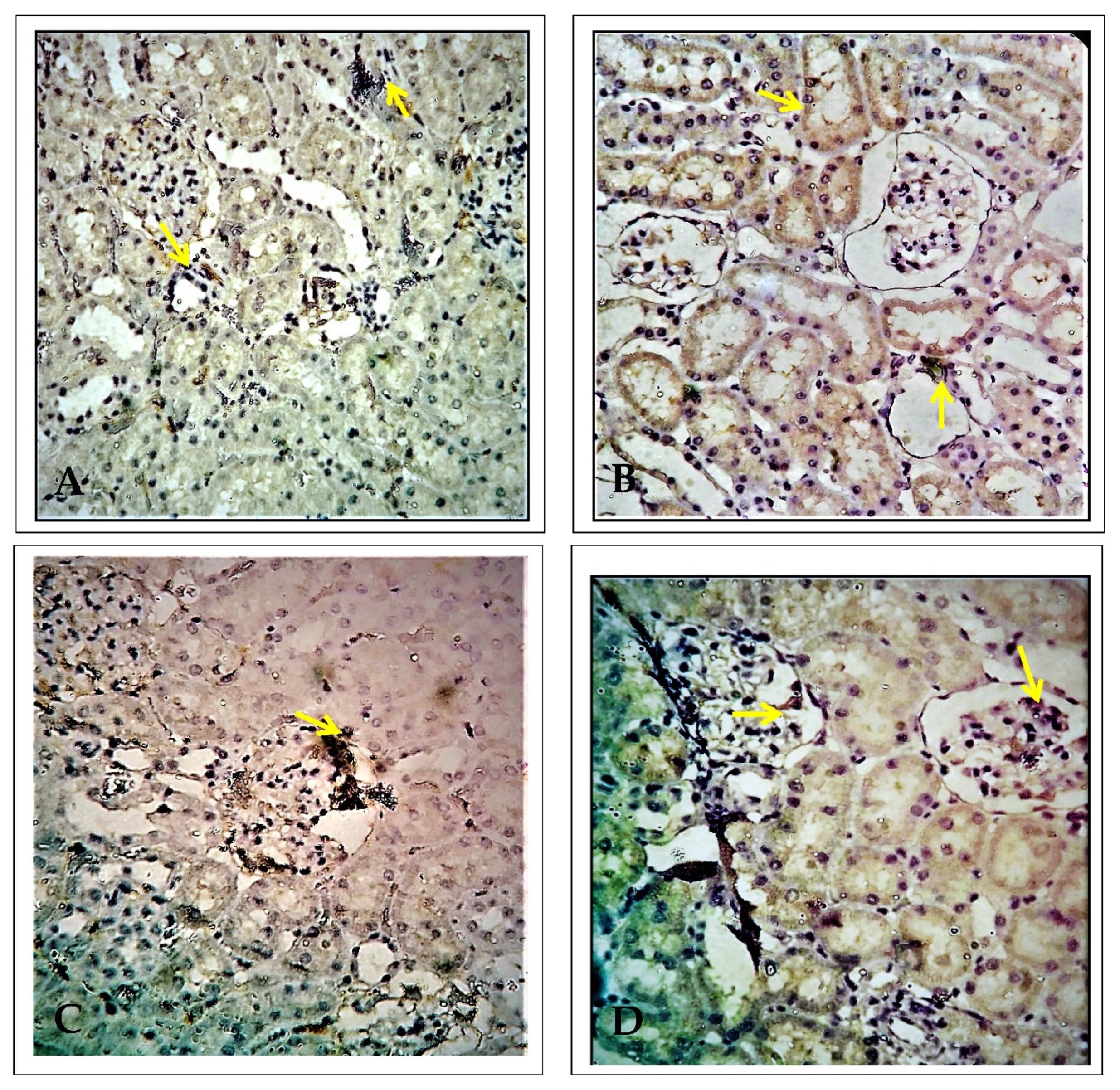
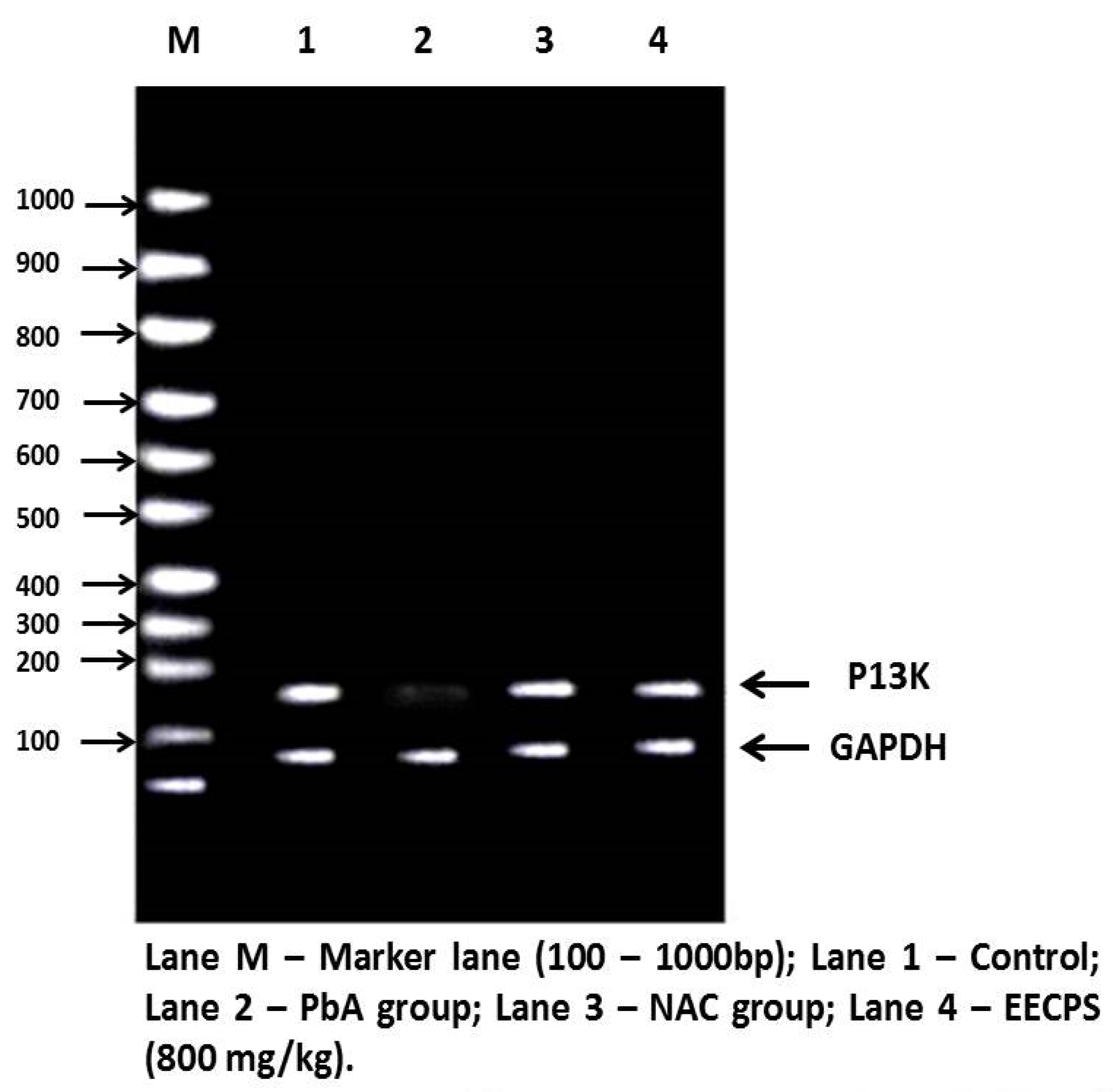
2.1. Effect of EECP on Kidney Histopathology Based on Periodic Acid–Schiff (PAS) Staining
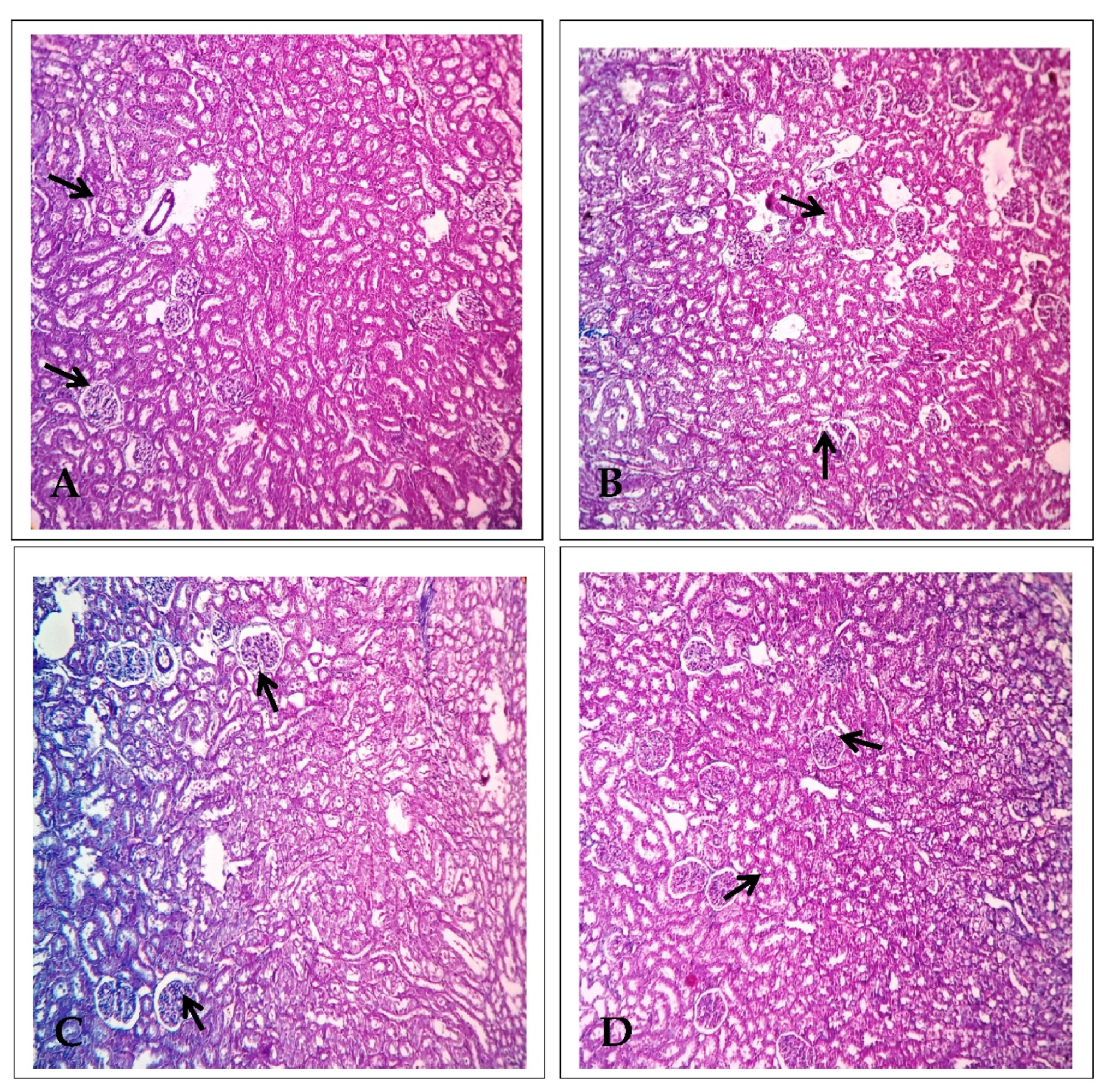
2.2. Effect of EECP on Kidney Histology Based on Using Masson’s Trichrome Staining
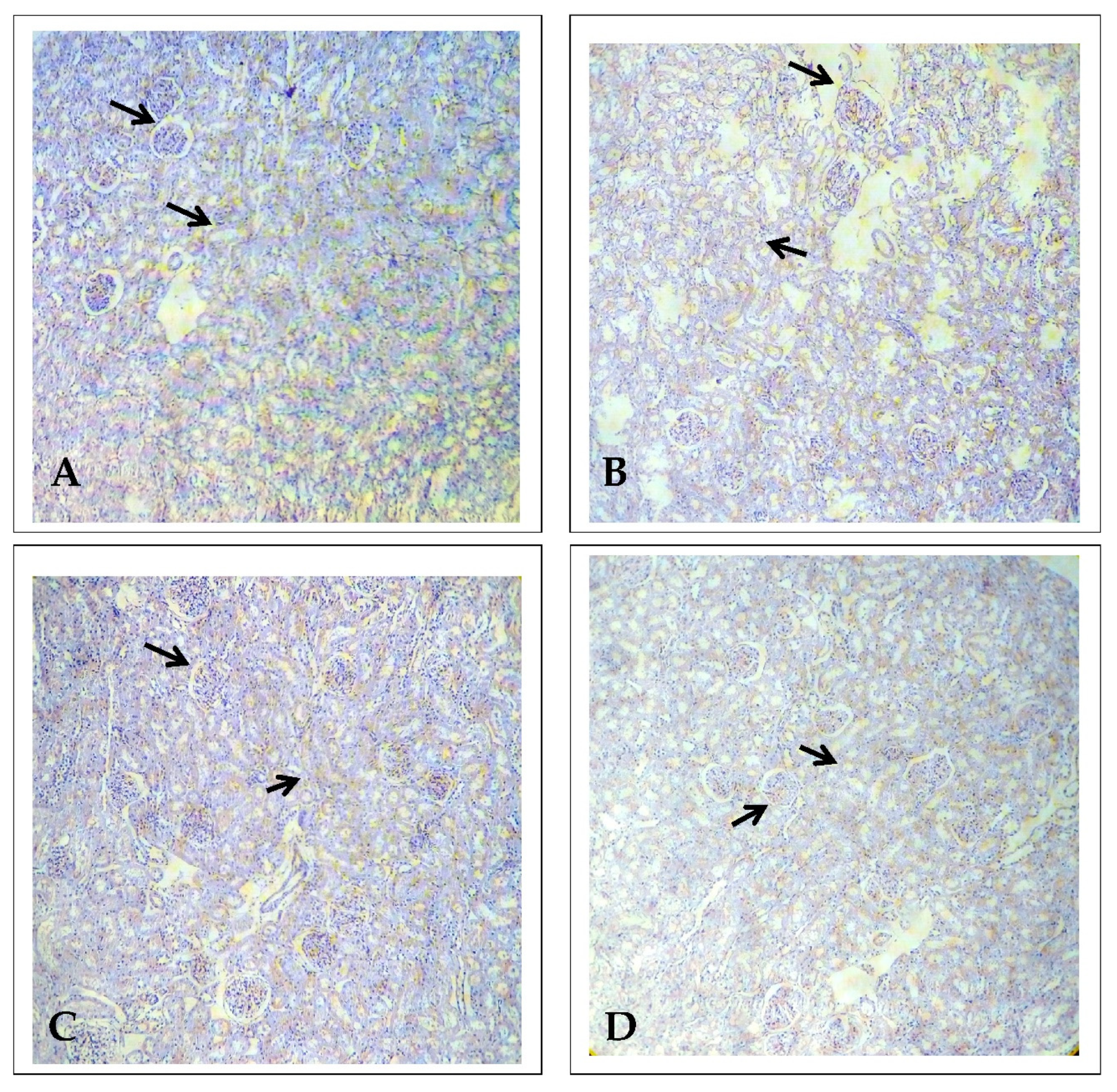
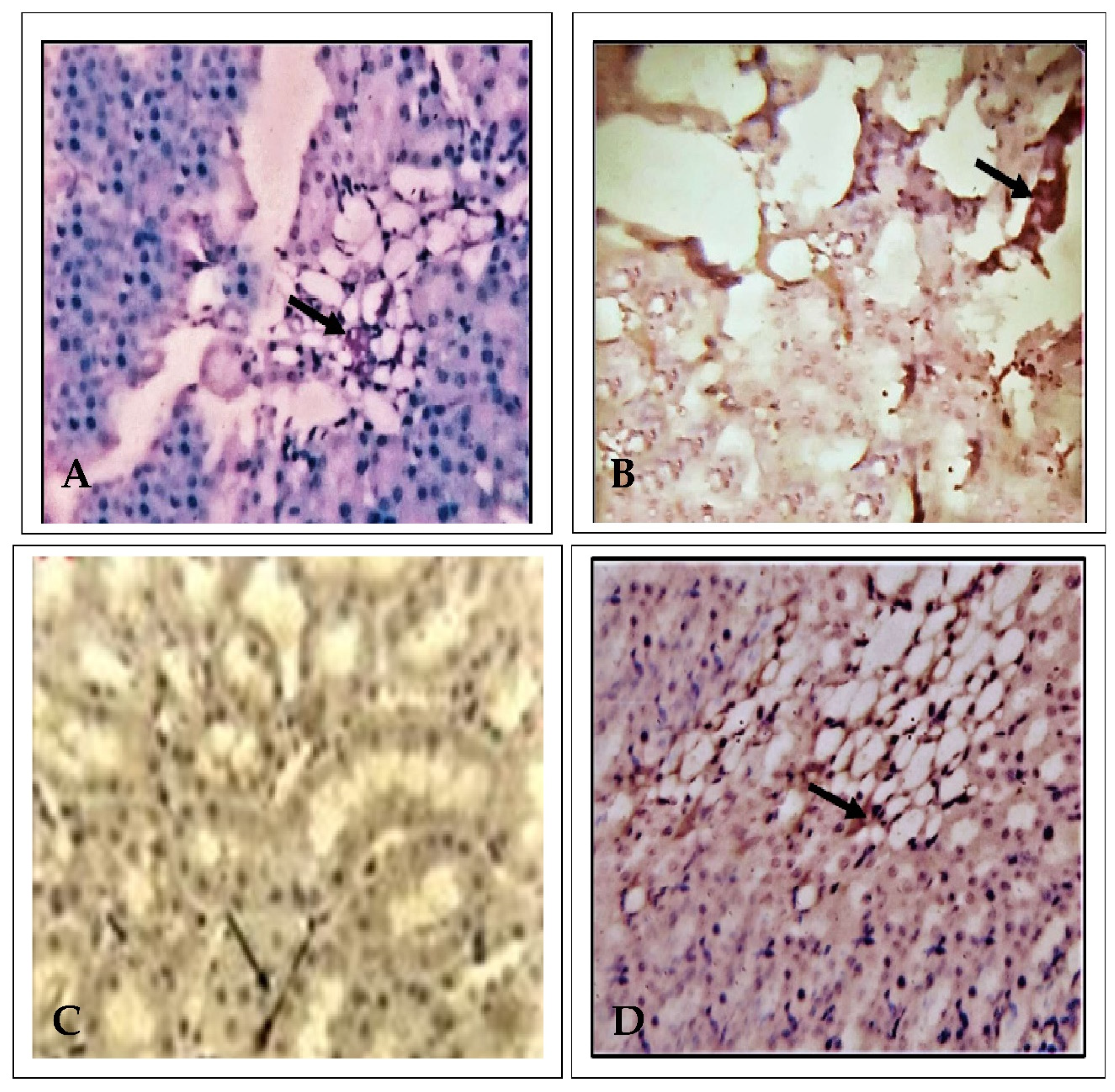
2.3. Effect of EECP on the Expression of Alpha Glutathione S-Transferase (α-GST) Based on Immunohistochemistry
2.4. Effect of EECP on the Expression of Delta-Aminolevulinate Dehydratase (ALAD) Using Immunohistochemistry
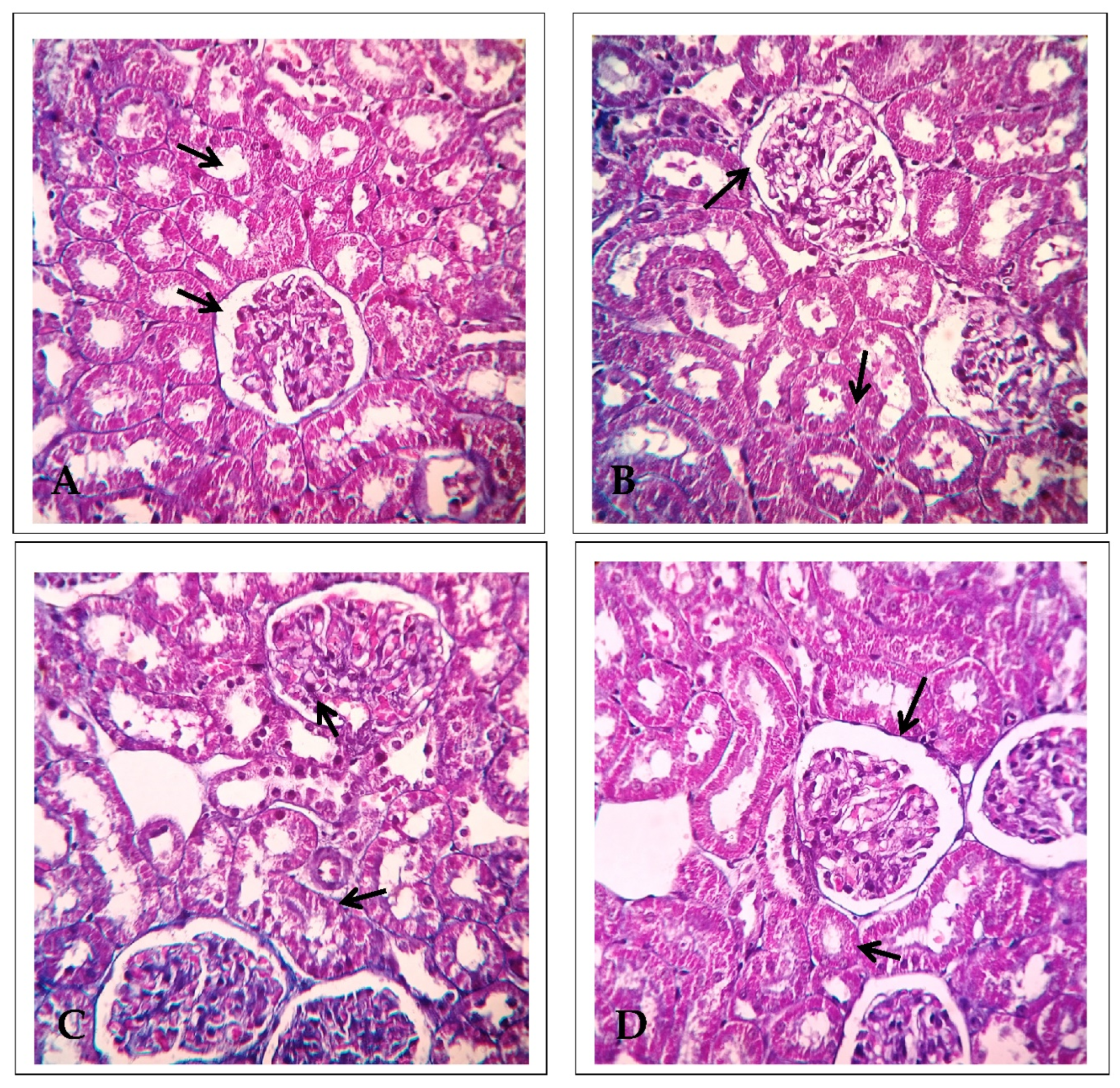
2.5. Effect of EECP against PbA-Induced Nephrotoxicity Based on Scanning Electron Microscopy (SEM)
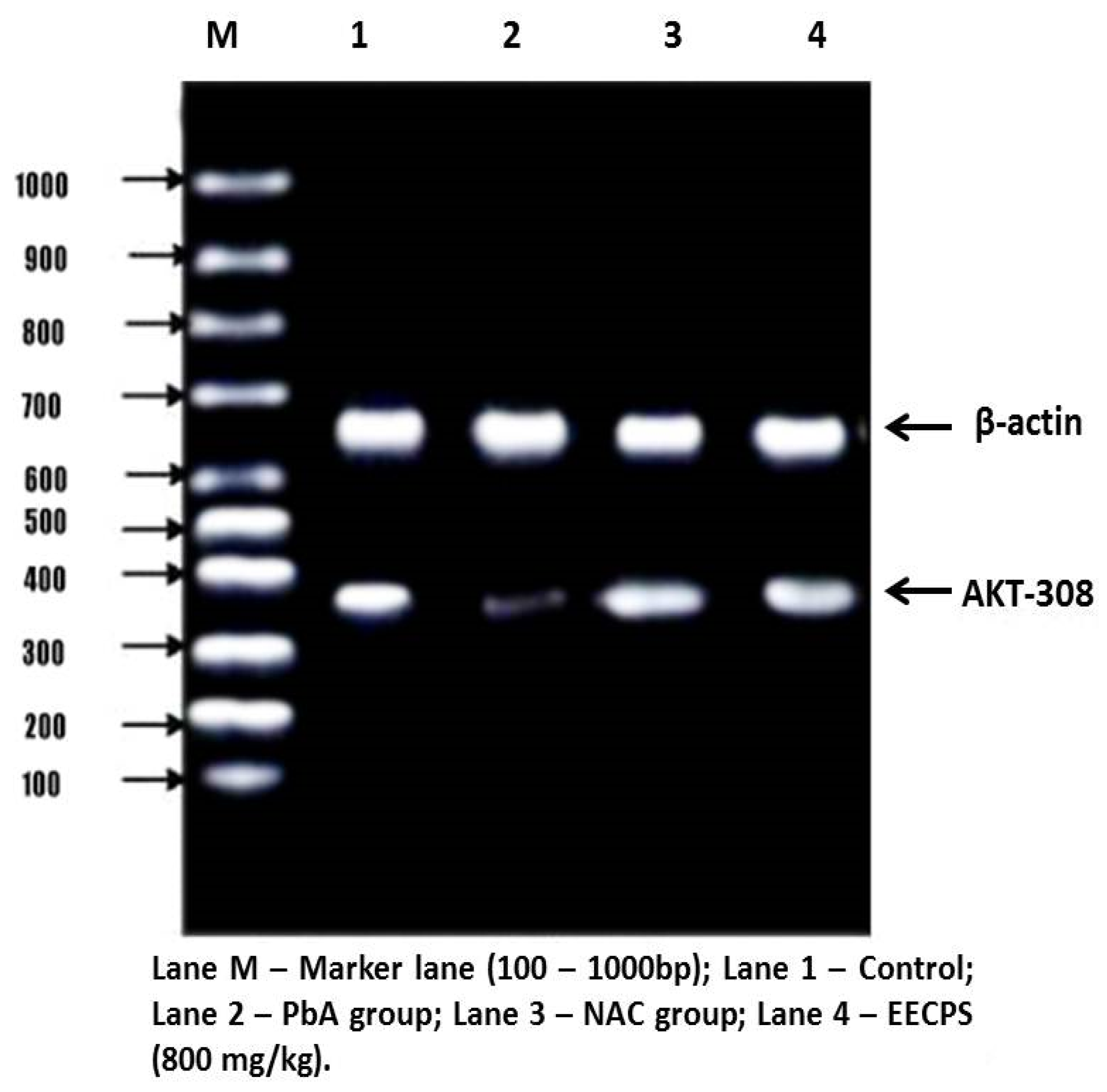
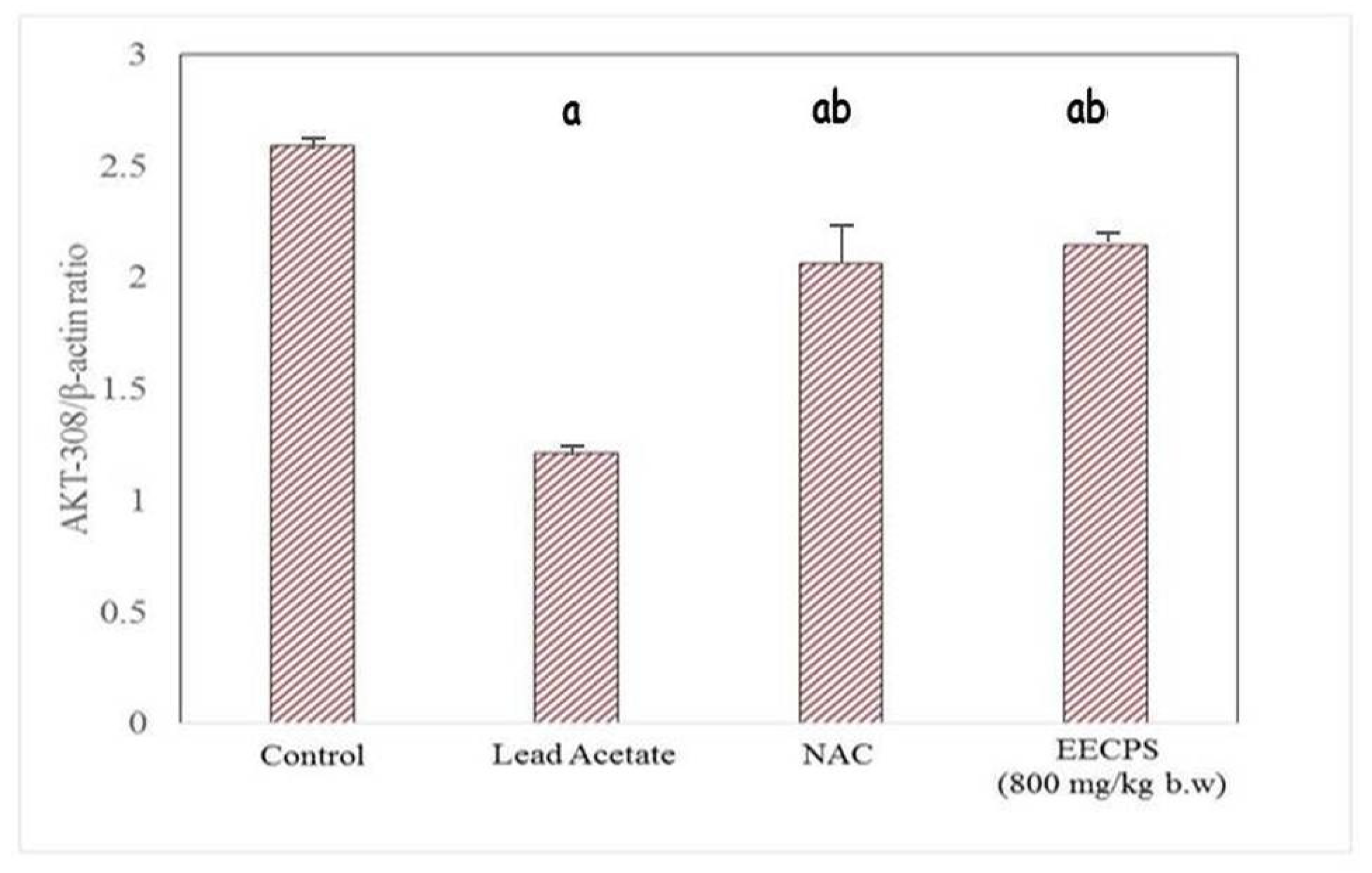
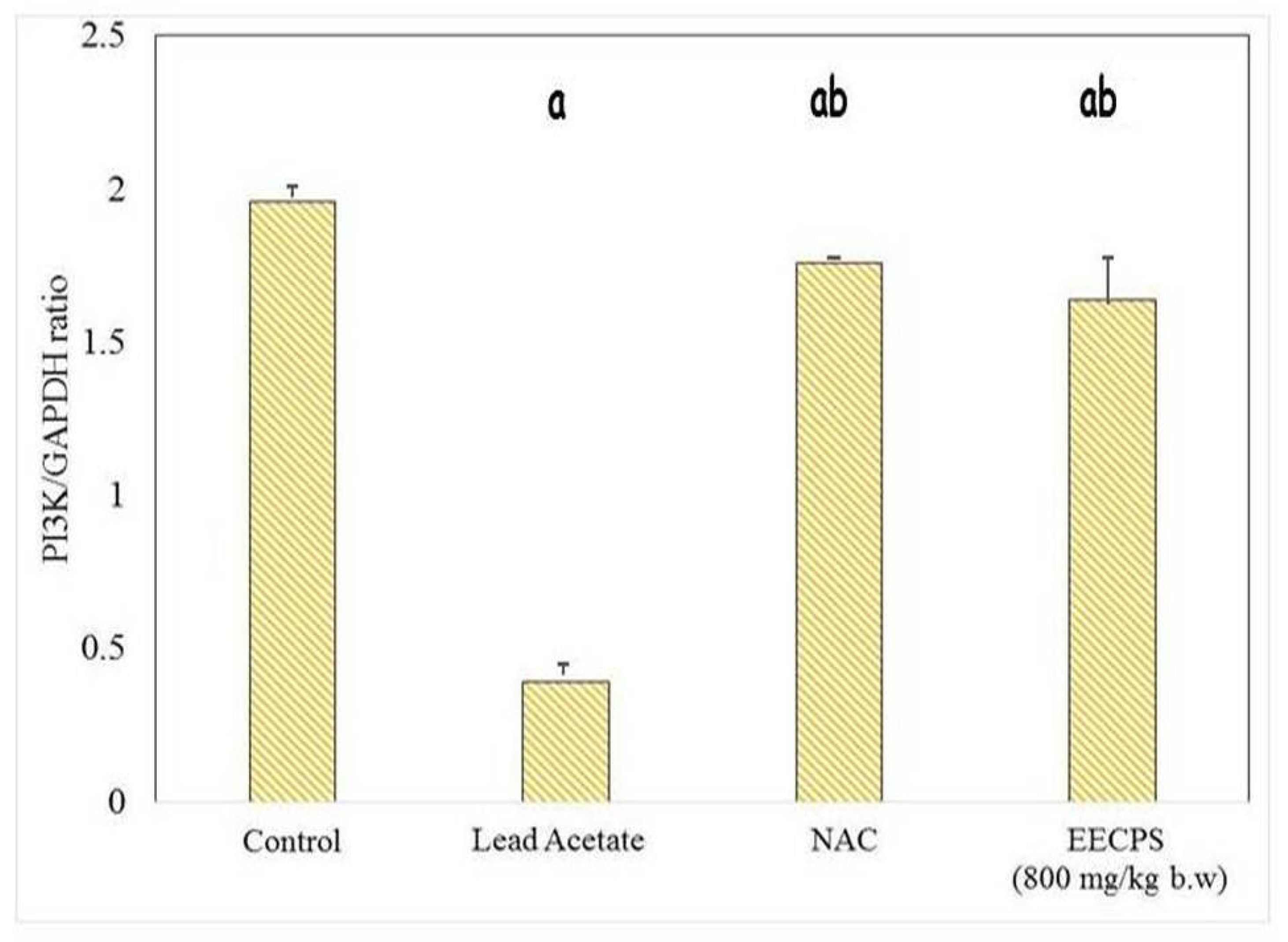
2.6. Effect of EECP on PI3K and AKT mRNA Levels in the Kidney
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Plant Material Preparation and Extraction
4.3. Experimental Animals
4.4. Experimental Design
- Group 1: Saline (2 mL/kg b.w./day, oral)
- Group 2: PbA (30 mg/kg b.w./day, oral)
- Group 3: NAC (200 mg/kg b.w./day, oral) + PbA (30 mg/kg b.w./day, oral)
- Group 4: EECP (800 mg/kg b.w./day, oral) + PbA (30 mg/kg b.w./day, oral)
4.5. Collection of Kdneys
4.6. Tissue Homogenization
4.7. Histopathological Studies
4.8. Expression of α-GST Using Immunohistochemistry
4.9. Expression of ALAD Using Immunohistochemistry
4.10. Electron Microscope Study
4.11. mRNA Expression Analysis by Reverse-Transcription Polymerase Chain Reaction (RT-PCR)
4.11.1. Isolation of Total RNA
4.11.2. cDNA Synthesis and PCR Amplification
4.11.3. Agarose Gel Electrophoresis
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lonappan, L.; Brar, S.K.; Das, R.K.; Verma, M.; Surampalli, R.Y. Diclofenac and its transformation products: Environmental occurrence and toxicity—A review. Environ. Int. 2016, 96, 127–138. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.Y.; Moon, A. Drug-induced nephrotoxicity and its biomarkers. Biomol. Ther. 2012, 20, 268–272. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, E.G.; Virji, M.A.; McClean, M.D.; Weinberg, J.; Woskie, S.; Pepper, L.D. Personal exposure, behavior, and work site conditions as determinants of blood lead among bridge painters. J. Occup. Environ. Hyg. 2010, 7, 80–87. [Google Scholar] [CrossRef]
- Maruthamuthu, M.K.; Ganesh, I.; Ravikumar, S.; Hong, S.H. Evaluation of zraP gene expression characteristics and construction of a lead (Pb) sensing and removal system in a recombinant Escherichia coli. Biotechnol. Lett. 2015, 37, 659–664. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Biological indices of lead exposure and body burden. In IPCS, Inorganic Lead Environmental Health Criteria 118’; WHO: Geneva, Switzerland, 1995; Volume 165, pp. 114–118. [Google Scholar]
- Flora, G.; Gupta, D.; Tiwari, A. Toxicity of lead: A review with recent updates. Interdiscip. Toxicol. 2012, 5, 47–58. [Google Scholar] [CrossRef]
- Negi, K.; Mirza, A. Nephroprotective and Therapeutic Potential of Traditional Medicinal Plants in Renal Diseases. J. Drug Res. Ayurvedic. Sci. 2020, 5, 177–185. [Google Scholar] [CrossRef]
- Godkar, P.B.; Gordon, R.K.; Ravindran, A.; Doctor, B.P. Celastrus paniculatus seed water-soluble extracts protect against glutamate toxicity in neuronal cultures from rat forebrain. J. Ethnopharmacol. 2004, 93, 213–219. [Google Scholar] [CrossRef]
- Lin, J.L.; Huang, P.T. Body lead stores and urate excretion in men with chronic renal disease. J. Rheumatol. 1994, 21, 705–709. [Google Scholar] [PubMed]
- Karmakar, N.; Saxena, R.; Anand, S. Histopathological changes induced in rat tissues by oral intake of lead acetate. Environ. Res. 1986, 41, 23–28. [Google Scholar] [CrossRef]
- Bancroft, J.D.; Stevens, A. Theory and Practice of Histological Techniques, 4th ed.; Churchill Livingstone: New York, NY, USA, 1996. [Google Scholar]
- Kumar, P.P.; Kumaravel, S.; Lalitha, C. Screening of antioxidant activity, total phenolics and GC-MS study of Vitex negundo. Afr. J. Biochem. Res. 2010, 4, 191–195. [Google Scholar]
- Liu, Y. Cellular and molecular mechanisms of renal fibrosis. Nat. Rev. Nephrol. 2011, 7, 684–696. [Google Scholar] [CrossRef]
- Genovese, F.; Manresa, A.A.; Leeming, D.J.; Karsdal, M.A.; Boor, P. The extracellular matrix in the kidney: A source of novel non-invasive biomarkers of kidney fibrosis? Fibrogenes. Tissue Repair. 2014, 7, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cescon, M.; Gattazzo, F.; Chen, P.; Bonaldo, P. Collagen VI at a glance. J. Cell Sci. 2015, 128, 3525–3531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catania, J.M.; Chen, G.; Parrish, A.R. Role of matrix metalloproteinases in renal pathophysiologies. Am. J. Physiol.-Renal Physiol. 2007, 292, 905–911. [Google Scholar] [CrossRef] [PubMed]
- Dieterle, F.; Sistare, F.; Goodsaid, F.; Papaluca, M.; Ozer, J.S.; Webb, C.P. Renal biomarker qualification submission: A dialog between the FDA-EMEA and Predictive Safety Testing Consortium. Nat. Biotechnol. 2010, 28, 455–462. [Google Scholar] [CrossRef]
- Branten, A.J.; Mulder, T.P.; Peters, W.H.; Assmann, K.J.; Wetzels, J.F. Urinary excretion of glutathione S transferases alpha and pi in patients with proteinuria: Reflection of the site of tubular injury. Nephron 2000, 85, 120–126. [Google Scholar] [CrossRef]
- Bieniaś, B.; Zajączkowska, M.; Borzęcka, H.; Sikora, P.; Wieczorkiewicz-Płaza, A.; Wilczyńska, B. Early Markers of Tubulointerstitial Fibrosis in Children With Idiopathic Nephrotic Syndrome: Preliminary Report. Medicine 2015, 94, e1746. [Google Scholar] [CrossRef]
- Lampe, J.W.; Chen, C.; Li, S.; Prunty, J.; Grate, M.T.; Meehan, D.E.; Barale, K.V.; Dightman, D.A.; Feng, Z.; Potter, J.D. Modulation of human glutathione S-transferases by botanically defined vegetable diets. Cancer Epidemiol. Biomark. Prev. 2000, 9, 787–793. [Google Scholar]
- Westhuyzen, J.; Endre, Z.H.; Reece, G.; Reith, D.M.; Saltissi, D.; Morgan, T.J. Measurement of tubular enzymuria facilitates early detection of acute renal impairment in the intensive care unit. Nephrol. Dial. Transplant. 2003, 18, 543–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakai, T.; Morita, Y. delta-Aminolevulinic acid in plasma or whole blood as a sensitive indicator of lead effects, and its relation to the other heme-related parameters. Int. Arch. Occup. Environ. Health 1996, 68, 126–132. [Google Scholar]
- Scinicariello, F.; Murray, H.E.; Moffett, D.B.; Abadin, H.G.; Sexton, M.J.; Fowler, B.A. Lead and delta-aminolevulinic acid dehydratase polymorphism: Where does it lead? A meta-analysis. Environ. Health Perspect. 2007, 115, 35–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asadauskaite, R.; Naginiene, R.; Abdrachmanovas, O. Aminolevulinic acid dehydratase in blood as a biomarker for low-level lead exposure. Cent. Eur. J. Public Health 2007, 11, S11. [Google Scholar]
- La-Llave-Leon, O.; Méndez-Hernandez, E.M.; Castellanos-Juarez, F.X.; Esquivel-Rodríguez, E.; Vázquez-Alaniz, F.; Sandoval-Carrillo, A.; García-Vargas, G.; Duarte-Sustaita, J.; Candelas-Rangel, J.L.; Salas-Pacheco, J.M. Association between Blood Lead Levels and Delta-Aminolevulinic Acid Dehydratase in Pregnant Women. Int. J. Environ. Res. Public Health 2017, 14, 432. [Google Scholar] [CrossRef] [Green Version]
- Chiu, Y.W.; Liu, T.Y.; Chuang, H.Y. The Effects of Lead Exposure on the Activities of [delta]-Aminolevulinic Acid Dehydratase with the Modification of the Relative Genotypes. EDP Sci. 2013, 1, 26005. [Google Scholar]
- Feksa, L.R.; Oliveira, E.; Trombini, T.; Luchese, M.; Bisi, S.; Linden, R.; Berlese, D.B.; Rojas, D.B.; Andrade, R.B.; Schuck, P.F. Pyruvate kinase activity and δ-aminolevulinic acid dehydratase activity as biomarkers of toxicity in workers exposed to lead. Arch. Environ. Contam. Toxicol. 2012, 63, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Jasim, S.M.; AL-Wasiti, E.A.; Subber, Z.J. Lead Exposure Effects on Batteries Manufacturing Factory Workers in Baghdad. Iraqi J. Med. Sci. 2012, 10, 321–327. [Google Scholar]
- Srinivasa Reddy, Y.; Pullakhandam, R.; Radha Krishna, K.; Uday Kumar, P.; Dinesh Kumar, B. Lead and essential trace element levels in school children: A cross-sectional study. Anal. Hum. Biol. 2011, 38, 372–377. [Google Scholar] [CrossRef] [PubMed]
- Jenette, J.C.; Olson, J.L.; Schwartz, M.M.; Silva, F.G. Primer on the Pathologic Diagnosis of Renal Disease. In Hepinstall’s Pathology of the Kidney; Lippincott Williams and Wilkins: Philadelphia, PA, USA, 2007; pp. 98–123. [Google Scholar]
- Pearson, J.M.; McWilliam, L.J.; Coyne, J.D.; Curry, A. Value of electron microscopy in diagnosis of renal disease. J. Clin. Pathol. 1994, 47, 126–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elhefnawy, N. Contribution of Electron Microscopy to the Final Diagnosis of Renal Biopsies in Egyptian Patients. Pathol. Oncol. Res. 2011, 17, 121–125. [Google Scholar] [CrossRef]
- Collan, Y.; Hirsimaki, P.; Aho, H. Value of Electron Microscopy in Kidney Biopsy Diagnosis. Ultrastruct. Pathol. 2005, 29, 461–468. [Google Scholar] [CrossRef] [PubMed]
- Tighe, J.R.; Jones, N.F. The diagnostic value of routine electron microscopy of renal biopsies. Proc. Roy. Soc. Med. 1970, 63, 475–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben-Bassat, M.; Stark, H.; Robson, M.; Rosenfield, J. Value of routine electron microscopy in the differential diagnosis of the nephrotic syndrome. Pathol. Microbiol. 1974, 41, 26–40. [Google Scholar]
- Dische, F.E.; Parsons, V. Experience in the diagnosis of glomerulonephritis using combined light microscopical, ultrastructural and immunofluorescence techniques-ananalysis of 134 cases. Histopathology 1977, 1, 331–362. [Google Scholar] [CrossRef]
- Spargo, B.H. Practical use of electron microscopy for the diagnosis of glomerular disease. Hum. Pathol. 1975, 6, 405–420. [Google Scholar] [CrossRef]
- Siegel, N.J.; Spargo, B.H.; Kashgarian, M.; Hayslett, J.P. An evaluation of routine electron microscopy in the examination of renal biopsies. Nephron 1973, 10, 209–215. [Google Scholar] [CrossRef]
- Cameron, J.S. Indications for renal biopsy, history of the procedure, and relationship of findings to further investigation and treatment. In Diagnostic Renal Pathology; Solez, K., Racusen, L., Olsen, S., Eds.; Transpath Inc.: Canoga Park, CA, USA, 2002; Chapter 2. [Google Scholar]
- Coimbra, T.M. Early events leading to renal injury in obese Zucker (fatty) rats with type II diabetes. Kidney Int. 2000, 57, 167–182. [Google Scholar] [CrossRef] [Green Version]
- Gassler, N. Podocyte injury underlies the progression of focal segmental glomerulosclerosis in the fa/fa Zucker rat. Kidney Int. 2001, 60, 106–116. [Google Scholar] [CrossRef] [Green Version]
- Wolf, G.; Chen, S.; Ziyadeh, F.N. From the periphery of the glomerular capillary wall toward the center of disease: Podocyte injury comes of age in diabetic nephropathy. Diabetes 2005, 54, 1626–1634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mifsud, S.A. Podocyte foot process broadening in experimental diabetic nephropathy: Amelioration with renin-angiotensin blockade. Diabetologia 2001, 44, 878–882. [Google Scholar]
- Kaplan, D.R.; Miller, F.D. Neurotrophin signal transduction in the nervous system. Curr. Opin. Neurobiol. 2000, 10, 381–391. [Google Scholar] [CrossRef]
- Rasul, A.; Ding, C.; Li, X.; Khan, M.; Yi, F.; Ali, M. Dracorhodin perchlorate inhibits PI3K/Akt and NF-KB activation, up-regulates the expression of p53, and enhances apoptosis. Apoptosis 2012, 17, 1104–1119. [Google Scholar] [CrossRef] [PubMed]
- Coffer, P.J.; Jin, J.; Woodgett, J.R. Protein kinase B (c-Akt): A multifunctional mediator of phosphatidylinositol 3 kinase activation. Biochem. J. 1998, 335, 1–3. [Google Scholar] [CrossRef]
- Kulik, G.; Klippel, A.; Weber, M.J. Antiapoptotic signalling by the insulin like growth factor I receptor, phosphatidylinositol 3-kinase, and Akt. Mol. Cell. Biol. 1997, 17, 1595–1606. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.F.; Zhang, H. Phytochemical Constituents, Health Benefits, and Industrial Applications of Grape Seeds: A Mini-Review. Antioxidants 2017, 6, 71. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Ma, Z.F.; Luo, X.; Li, X. Effects of Mulberry Fruit (Morus alba L.) Consumption on Health Outcomes: A Mini-Review. Antioxidants 2018, 7, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khor, B.K.; Chear, N.J.; Azizi, J.; Khaw, K.Y. Chemical Composition, Antioxidant and Cytoprotective Potentials of Carica papaya Leaf Extracts: A Comparison of Supercritical Fluid and Conventional Extraction Methods. Molecules 2021, 26, 1489. [Google Scholar] [CrossRef]
- Yuvaraj, M.F.; Vijayakumar, J.; Gunapriya, R.; Vijayalakshmi, S.; Senthilkumar, S.; Vijayaraghavan, R.; Ethirajan, S. Protective Effect of Carica papaya Leaf Extract against Mercuric Chloride Induced Nephrotoxicity in Wistar Rats. Pharmacogn. Mag. 2020, 16, 379–384. [Google Scholar]
- Balaji, K.; Vijayakumar, J.; Senthilkumar, S.; Vijayaraghavan, R.; Naveenkumar, V. Protective Effect of Celastrus paniculatus Seed Extract against Lead Acetate Induced Nephrotoxicity in Wistar Rats. Indian J. Pharm. Educ. Res. 2021, 55. in press. [Google Scholar]
- Parimala, S.; Shashidhar, G.H.; Sridevi, C.H.; Jyothi, V.; Suthakaran, R. Anti-inflammatory activity of Celastrus paniculatus seeds. Int. J. Pharm. Technol. Res. 2009, 1, 974–4304. [Google Scholar]
- Jho, D.H.; Babcock, T.A.; Tevar, R.; Helton, W.S.; Espat, N.J. Eicosapentaenoic acid supplementation reduces tumor volume and attenuates cachexia in a rat model of progressive non-metastasizing malignancy. J. Parenter. Enter. Nutr. 2002, 26, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Selvamani, V.; Zareei, A.; Elkashif, A.; Muralikannan, M.; Chittiboyina, S.; Delisi, D.; Li, Z.; Cai, L.; Pol, V.G.; Mohamed, N.; et al. Hierarchical Micro/Mesoporous Copper Structure with Enhanced Antimicrobial Property via Laser Surface Texturing. Adv. Mater. Interfaces 2020, 7, 1901890. [Google Scholar] [CrossRef]
- Maruthamuthu, M.K.; Selvamani, V.; Nadarajan, S.P.; Yun, H.; Oh, Y.K.; Eom, G.T.; Hong, S.H. Manganese and cobalt recovery by surface display of metal binding peptide on various loops of OmpC in Escherichia coli. J. Ind. Microbiol. Biotechnol. 2018, 45, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Muralikannan, M.; Lee, W.; Hong, S.H.; Kang, S. Efficacy of antifreeze proteins from Clupea harangues and Anarhichas minor gas hydrate inhibition via cell surface display. Chem. Eng. Sci. 2020, 215, 115470. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Forward Primer | Reverse Primer | Product Size (bp) |
|---|---|---|---|
| AKT | 5′-ATCCCCTCAACAACTTCTCAGT-3′ | R: 5′-CTTCCGTCCACTCTTCTCTTTC-3′ | 447 |
| β-actin | 5′-TGACGGGGTCACCCACACT-3′ | 5′-CTTAGAAGCATTGCGGTGG-3′ | 659 |
| Genes | Forward Primer | Reverse Primer | Product Size |
|---|---|---|---|
| PI3K | 5′-GTTCACCAATCCTGCCTGTG-3′ | 5′-CTGCATCACCTTCATCTGGC-3′ | 248 |
| GAPDH | 5′-TGACGGGGTCACCCACACT-3′ | 5′-CTTAGAAGCATTGCGGTGG-3′ | 192 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balaji, K.; Vijayakumar, J.; Sankaran, P.K.; Senthilkumar, S.; Vijayaraghavan, R.; Selvaraj, J.; Yuvaraj, M.F. Molecular Studies on the Nephroprotective Potential of Celastrus paniculatus against Lead-Acetate-Induced Nephrotoxicity in Experimental Rats: Role of the PI3K/AKT Signaling Pathway. Molecules 2021, 26, 6647. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26216647
Balaji K, Vijayakumar J, Sankaran PK, Senthilkumar S, Vijayaraghavan R, Selvaraj J, Yuvaraj MF. Molecular Studies on the Nephroprotective Potential of Celastrus paniculatus against Lead-Acetate-Induced Nephrotoxicity in Experimental Rats: Role of the PI3K/AKT Signaling Pathway. Molecules. 2021; 26(21):6647. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26216647
Chicago/Turabian StyleBalaji, Karunakaran, Jagadish Vijayakumar, Ponnusamy Kasirajan Sankaran, Sivanesan Senthilkumar, Rajagopalan Vijayaraghavan, Jayaraman Selvaraj, and Maria Francis Yuvaraj. 2021. "Molecular Studies on the Nephroprotective Potential of Celastrus paniculatus against Lead-Acetate-Induced Nephrotoxicity in Experimental Rats: Role of the PI3K/AKT Signaling Pathway" Molecules 26, no. 21: 6647. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26216647