
Kuwanon T and Sanggenon a Isolated from Morus alba Exert Anti-Inflammatory Effects by Regulating NF-κB and HO-1/Nrf2 Signaling Pathways in BV2 and RAW264.7 Cells

 , and
, and 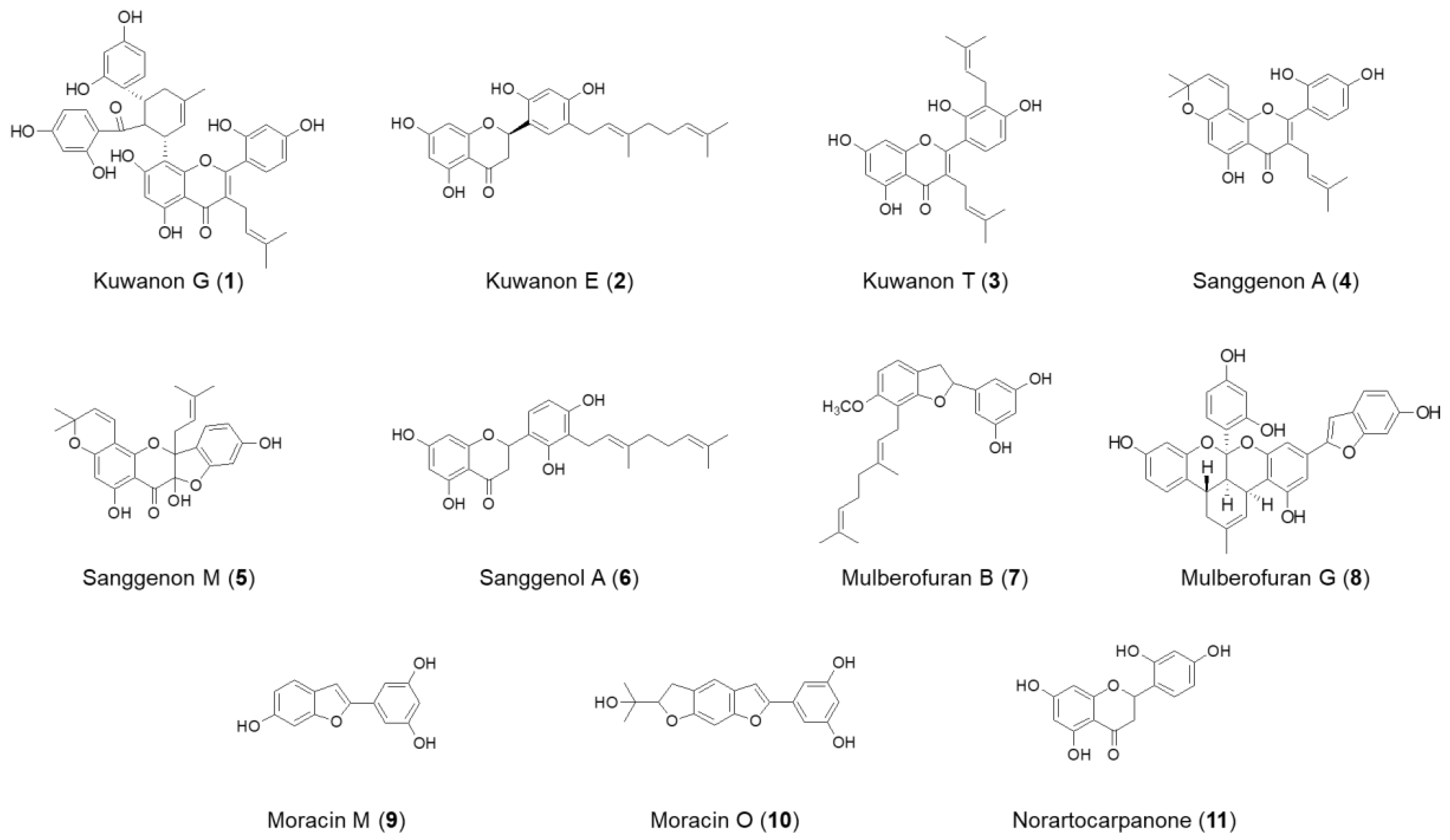{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
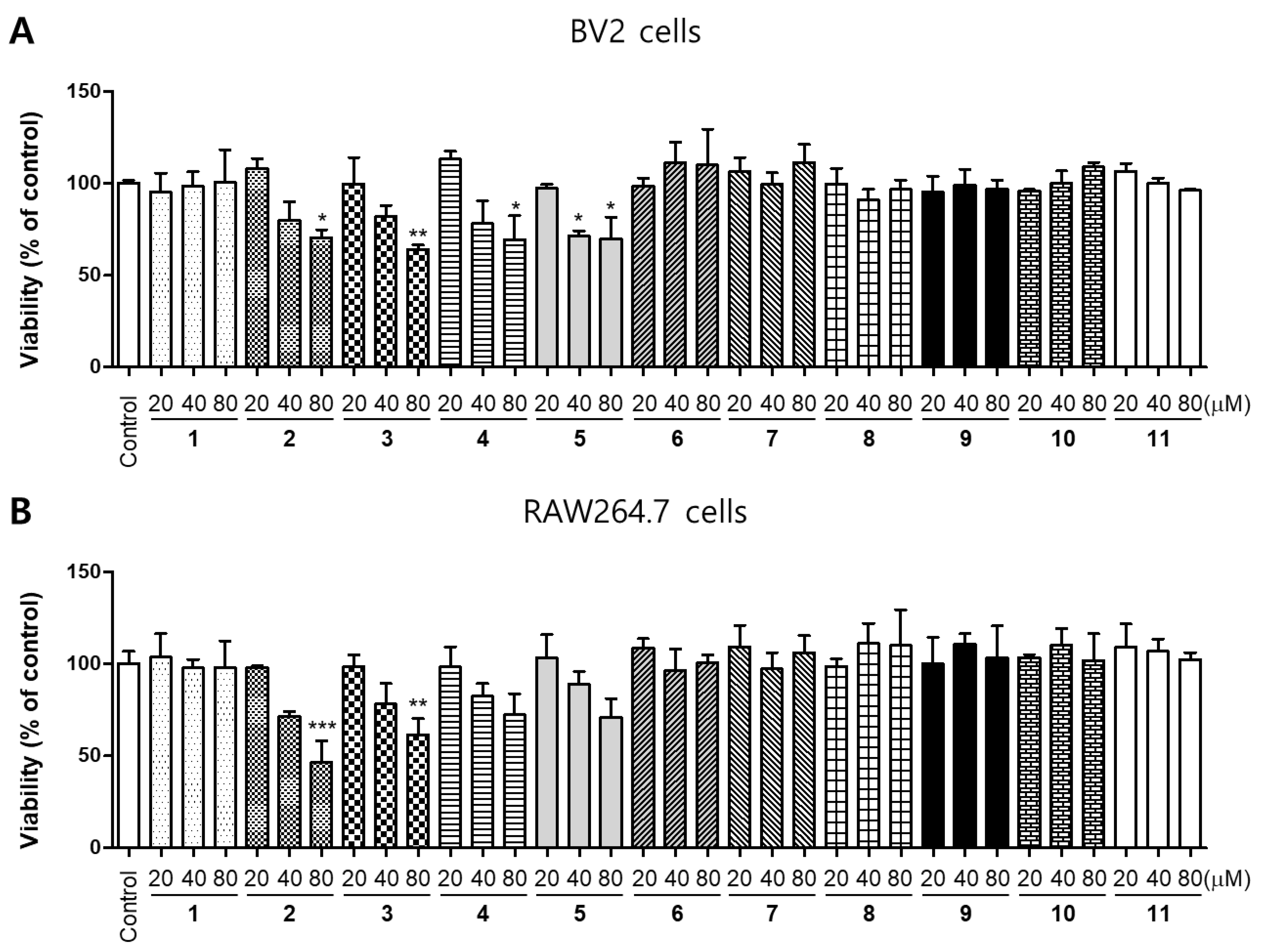
2.1. Effects of 11 Compounds Isolated from M. alba on the Viability of BV2 and RAW264.7 Cells
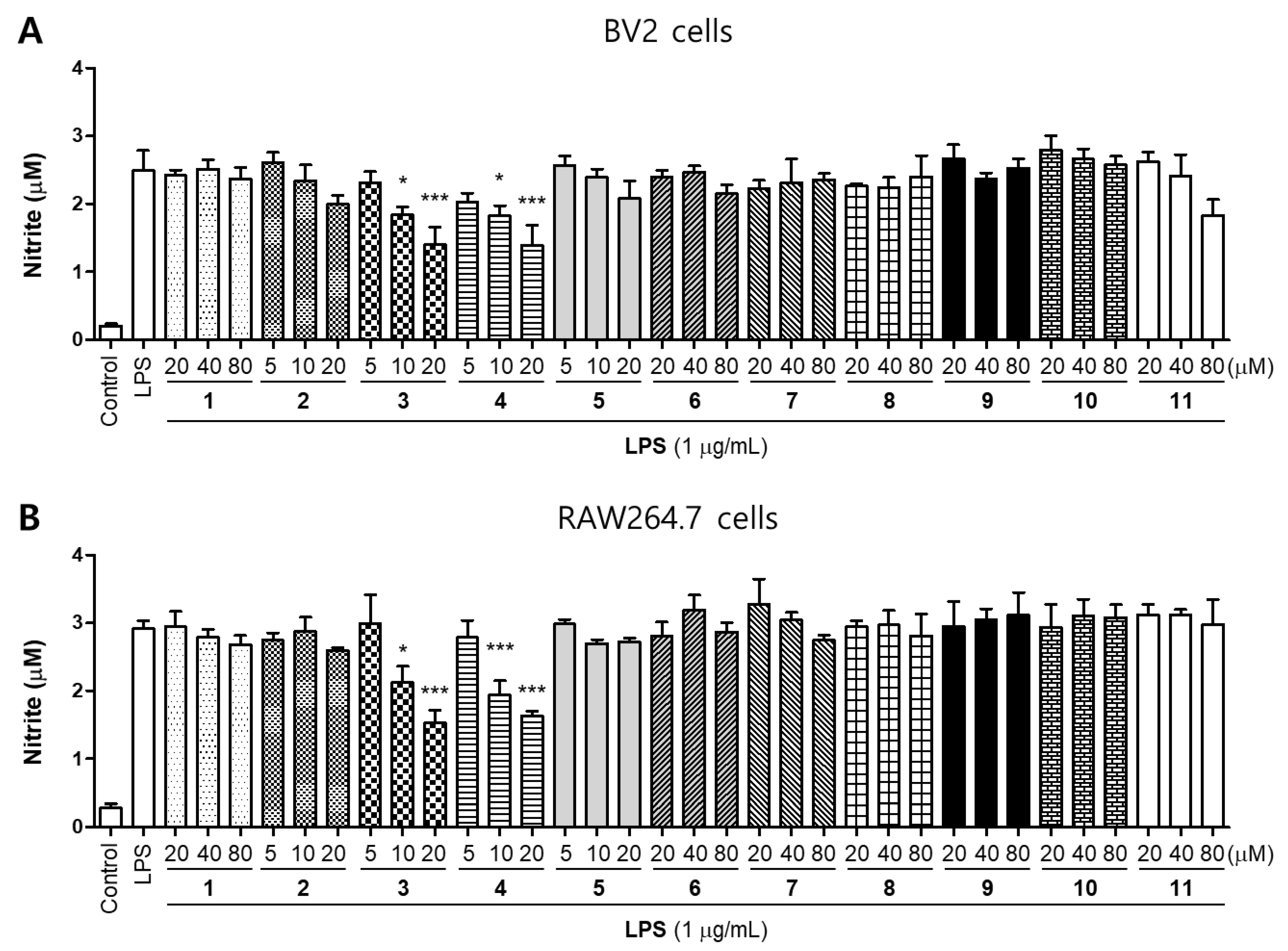
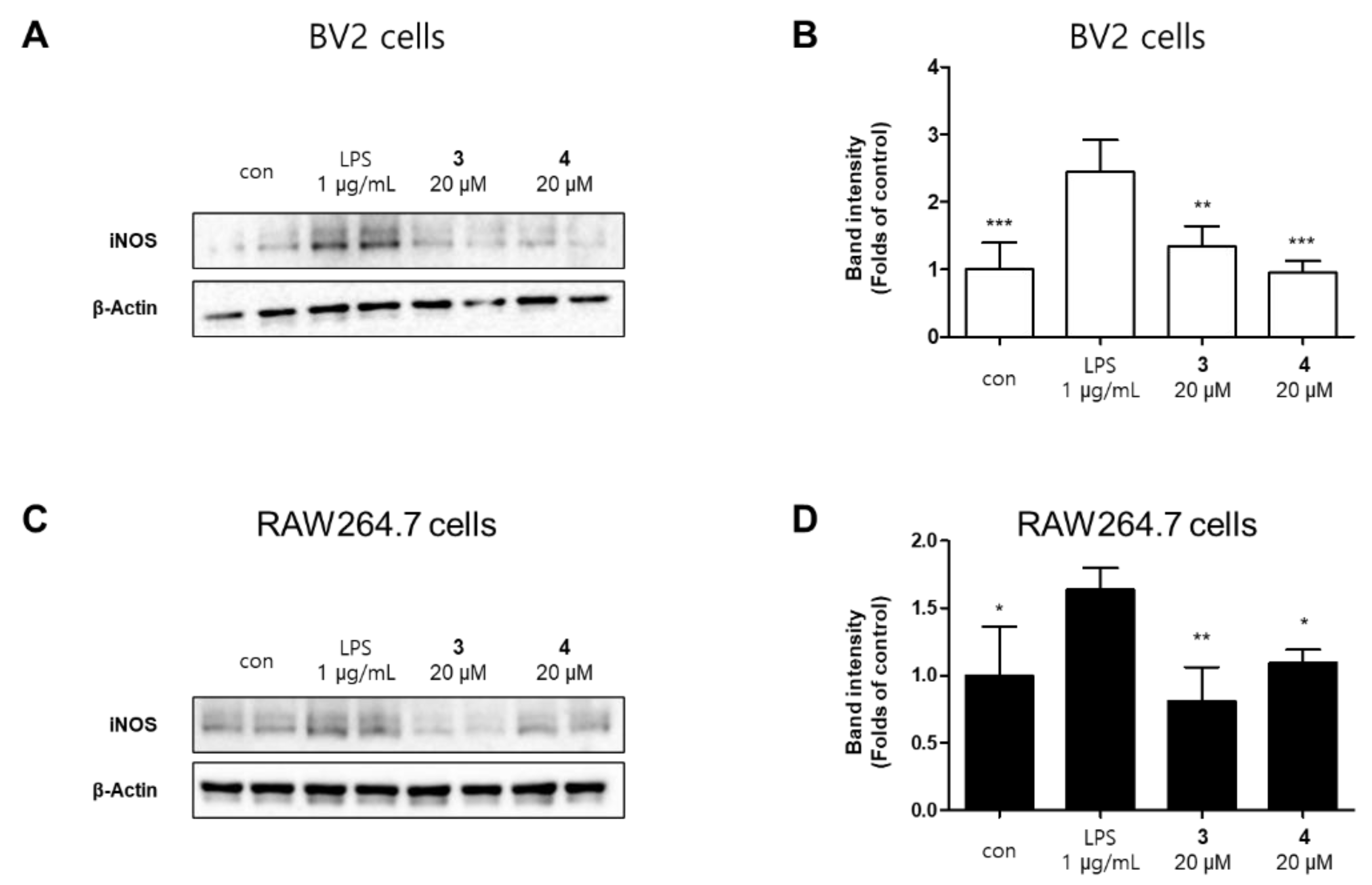
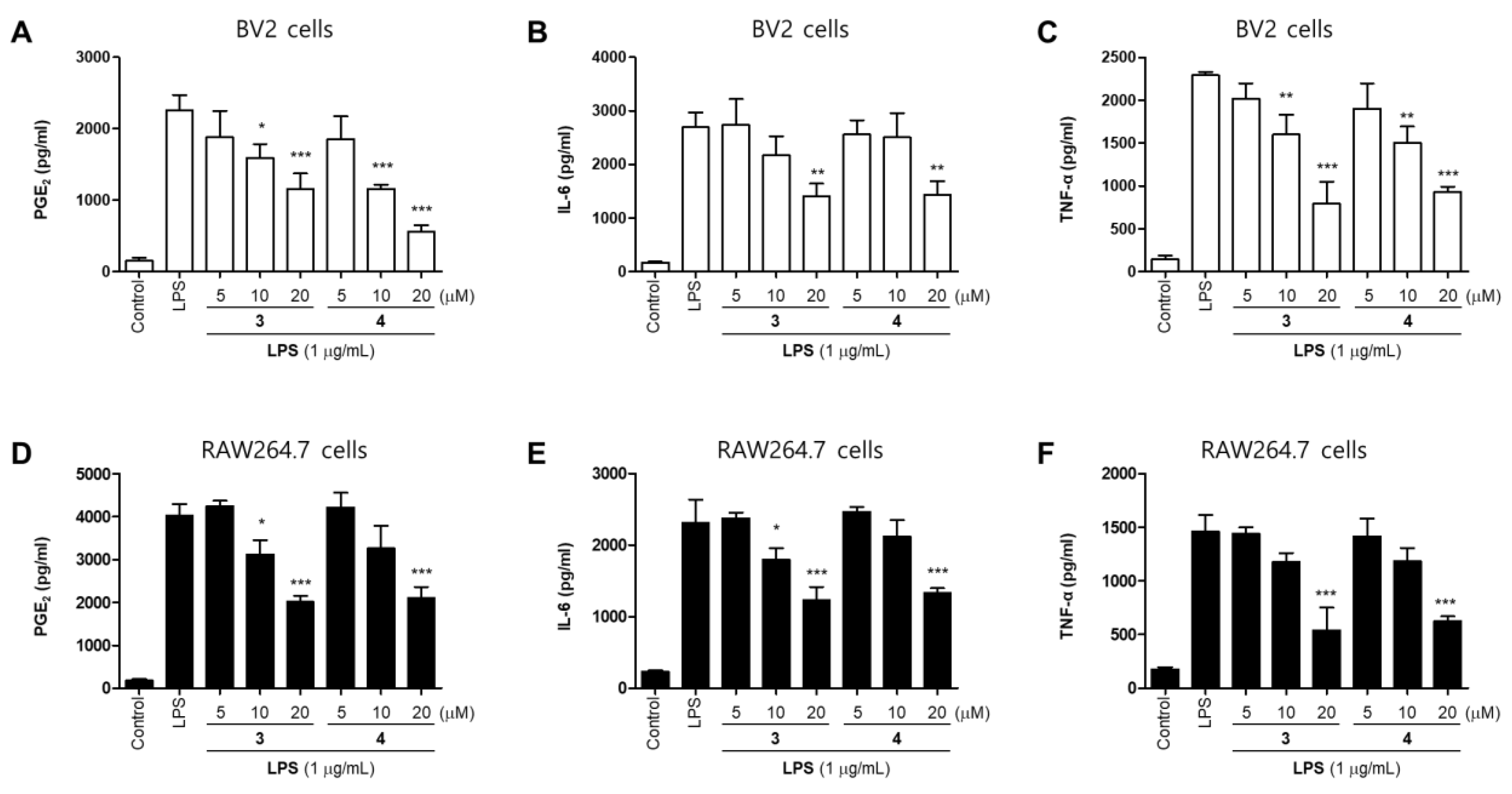
2.2. Effects of 11 Compounds Isolated from M. alba on the Expression of Inflammatory Factors and iNOS Protein in BV2 and RAW264.7 Cells
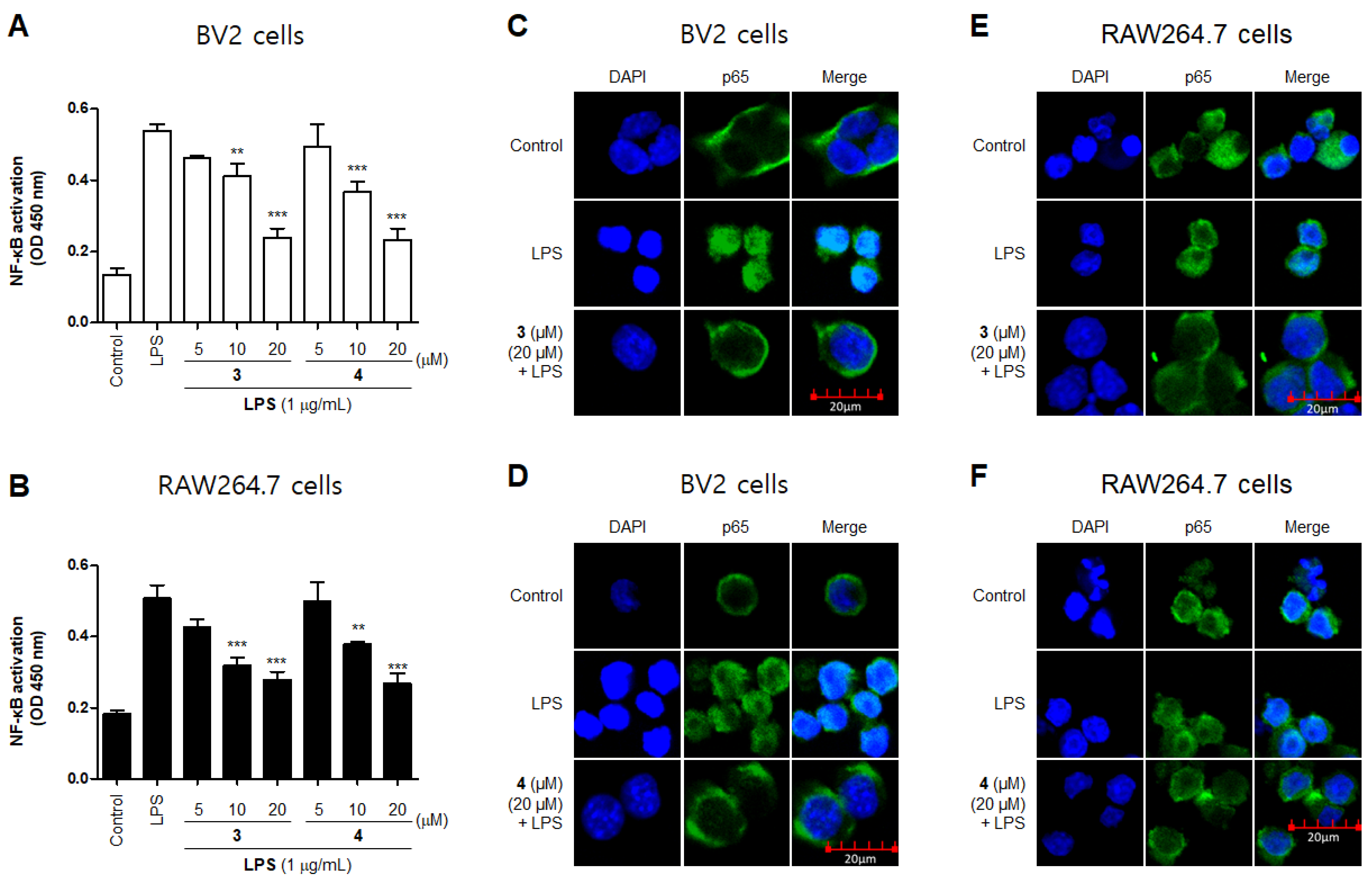
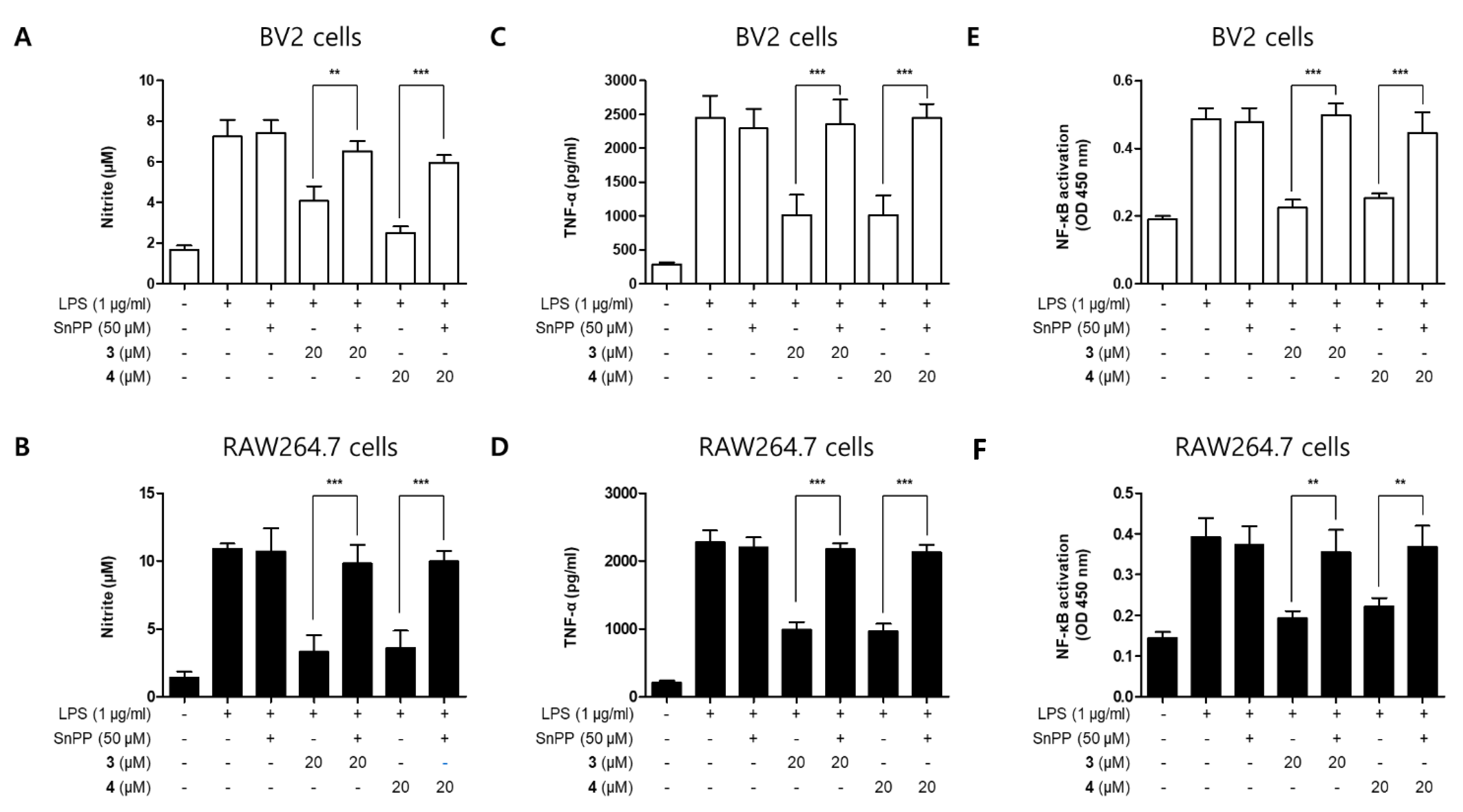
2.3. Effects of Compounds 3 and 4 on NF-κB Translocation in BV2 and RAW264.7 Cells
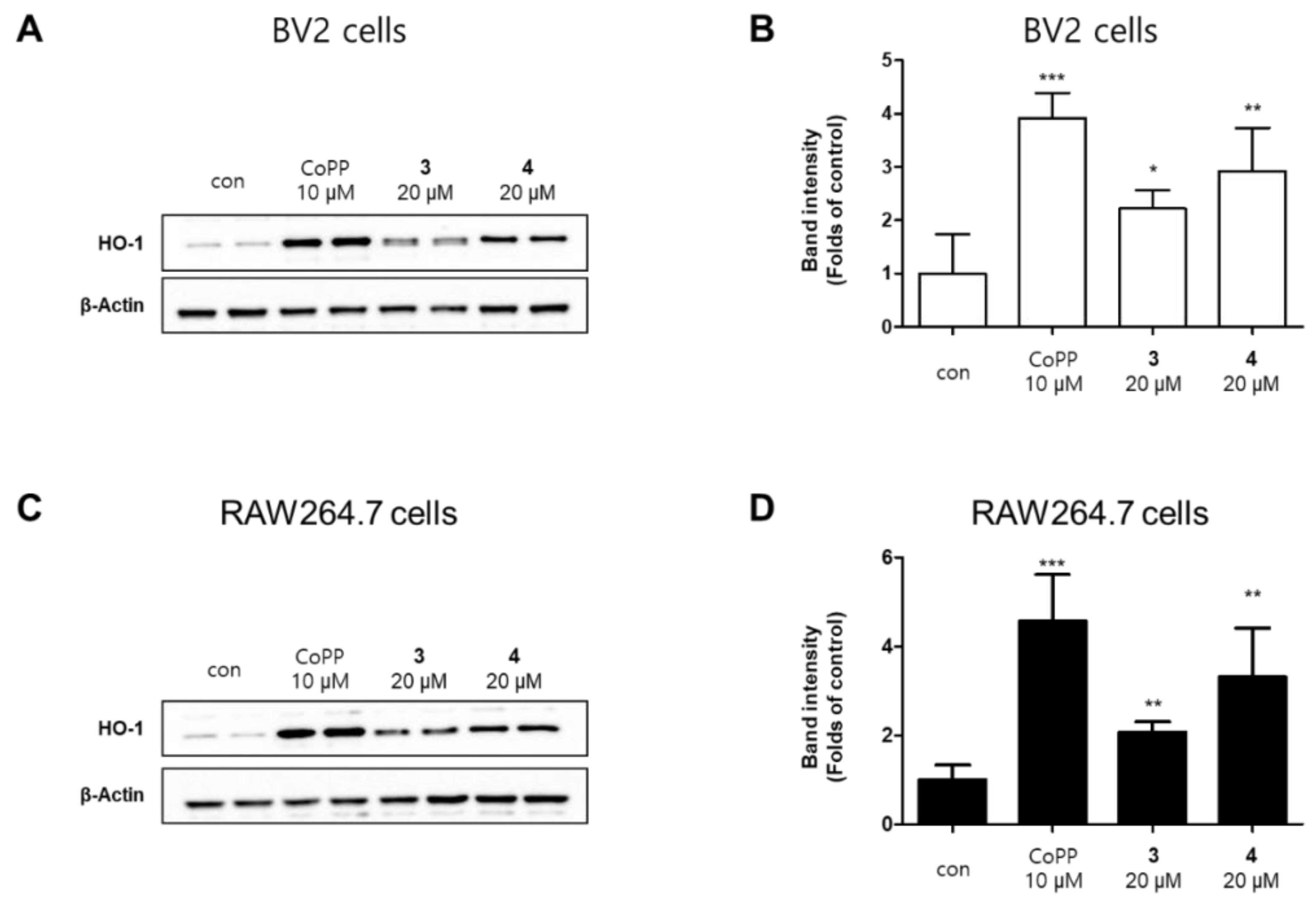
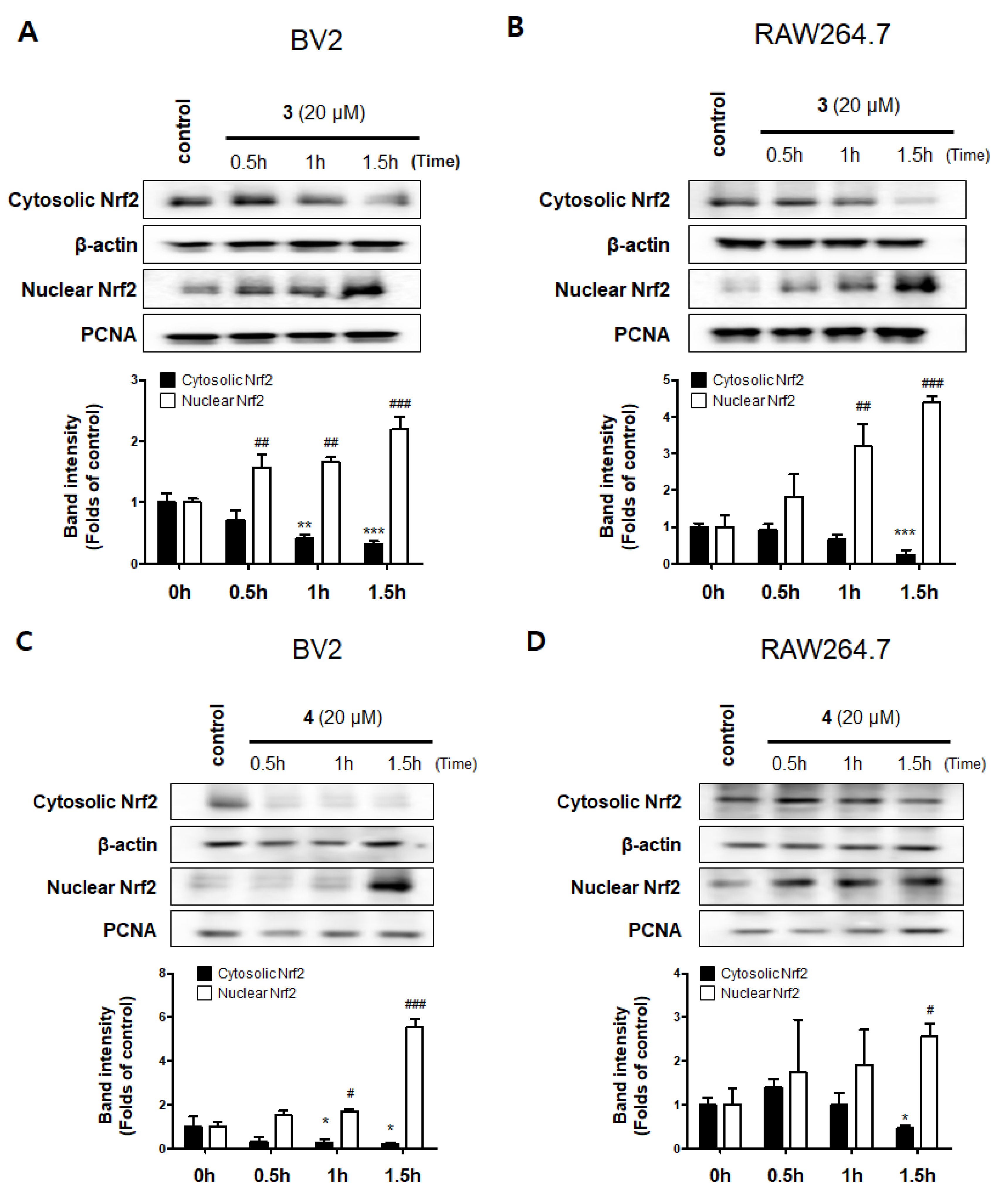
2.4. Effects of Compounds 3 and 4 on the Nrf2/HO-1 Pathway in BV2 and RAW264.7 Cells
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture and Viability Assays
4.3. Measurement of NO Production
4.4. PGE2 Assay
4.5. Measurement of IL-6 and TNF-α Levels
4.6. Western Blot Analysis
4.7. NF-κB Localization and Immunofluorescence Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Bae, K. The Medicinal Plants of Korea; Kyo-Hak Pub. Co.: Seoul, Korea, 2000; p. 73. [Google Scholar]
- Mihara, S.; Hara, M.; Nakamura, M.; Sakurawi, K.; Tokura, K.; Fujimoto, M.; Fukai, T.; Nomura, T. Non-peptide bombesin receptor antagonists, kuwanon G and H, isolated from mulberry. Biochem. Biophys. Res. Commun. 1995, 213, 594–599. [Google Scholar] [CrossRef]
- Ham, A.; Lee, H.J.; Hong, S.S.; Lee, D.; Mar, W. Moracenin D from Mori Cortex radicis protects SH-SY5Y cells against dopamine-induced cell death by regulating nurr1 and α-synuclein expression. Phytother. Res. 2012, 26, 620–624. [Google Scholar] [CrossRef]
- Lee, H.J.; Lyu, D.H.; Koo, U.; Nam, K.W.; Hong, S.S.; Kim, K.O.; Kim, K.H.; Lee, D.; Mar, W. Protection of prenylated flavonoids from Mori Cortex Radicis (Moraceae) against nitric oxide-induced cell death in neuroblastoma SH-SY5Y cells. Arch. Pharm. Res. 2012, 35, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.H.; Zhang, S.D.; Wang, L.T.; Yu, L.; Zhao, X.L.; Ni, H.Y.; Wang, Y.Q.; Wang, J.D.; Shan, C.H.; Fu, Y.J. Anti-inflammatory effects of neochlorogenic acid extract from mulberry leaf (Morus alba L.) against LPS-stimulated inflammatory response through mediating the AMPK/Nrf2 signaling pathway in A549 cells. Molecules 2020, 25, 1385. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Ko, H.J.; Woo, E.R.; Lee, S.K.; Moon, B.S.; Lee, C.W.; Mandava, S.; Samala, M.; Lee, J.; Kim, H.P. Moracin M inhibits airway inflammation by interrupting the JNK/c-Jun and NF-κB pathways in vitro and in vivo. Eur. J. Pharmacol. 2016, 783, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.W.; Kang, S.Y.; Kang, J.S.; Kim, A.R.; Woo, E.R.; Park, Y.K. Effect of Kuwanon G isolated from the root bark of Morus alba on ovalbumin-induced allergic response in a mouse model of asthma. Phytother. Res. 2014, 28, 1713–1719. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.W.; Ko, W.M.; Park, J.H.; Seo, K.H.; Oh, E.J.; Lee, D.Y.; Lee, D.S.; Kim, Y.C.; Lim, D.W.; Han, D.; et al. Isoprenylated flavonoids from the root bark of Morus alba and their hepatoprotective and neuroprotective activities. Arch. Pharm Res. 2015, 38, 2066–2075. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.G.; Matsuzaki, K.; Takamatsu, S.; Kitanaka, S. Inhibitory effects of constituents from Morus alba var. multicaulis on differentiation of 3T3-L1 cells and nitric oxide production in RAW264.7 cells. Molecules 2011, 16, 6010–6022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elizabeth, A.; Adegbuyi, A.; Olusegun, A.; Benneth, B.A.; Anthony, E.; Abayomi, A.; Solomo, U. Morin hydrate attenuates chronic stress-induced memory impairment and degeneration of hippocampal subfields in mice: The role of oxidative, nitrergic and neuroinflammatory pathways. Metab. Brain Dis. 2020, 35, 1145–1156. [Google Scholar] [CrossRef] [PubMed]
- Liang, D.; Li, F.; Fu, Y.; Cao, Y.; Song, X.; Wang, T.; Wang, W.; Guo, M.; Zhou, E.; Li, D.; et al. Thymol inhibits LPS-stimulated inflammatory response via down-regulation of NF-κB and MAPK signaling pathways in mouse mammary epithelial cells. Inflammation 2014, 37, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K. The function of microglia through purinergic receptors: Neuropathic pain and cytokine release. Pharmacol. Ther. 2006, 109, 210–226. [Google Scholar] [CrossRef]
- Tucureanu, M.M.; Rebleanu, D.; Constantinescu, C.A.; Deleanu, M.; Voicu, G.; Butoi, E.; Calin, M.; Manduteanu, I. Lipopolysaccharide-induced inflammation in monocytes/macrophages is blocked by liposomal delivery of G i-protein inhibitor. Int. J. Nanomed. 2017, 13, 63–76. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.B.; Han, A.R.; Park, E.Y.; Kim, J.Y.; Cho, W.; Lee, J.; Seo, E.K.; Lee, K.T. Inhibition of LPS-induced iNOS, COX-2 and cytokines expression by poncirin through the NF-kappaB inactivation in RAW 264.7 macrophage cells. Biol. Pharm. Bull. 2007, 30, 2345–2351. [Google Scholar] [CrossRef] [Green Version]
- Jin, C.Y.; Lee, J.D.; Park, C.; Choi, Y.H.; Kim, G.Y. Curcumin attenuates the release of pro-inflammatory cytokines in lipopolysaccharide-stimulated BV2 microglia. Acta Pharmacol. Sin. 2007, 28, 1645–1651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srinivasan, L.; Harris, M.C.; Kilpatrick, L.E. Cytokines and inflammatory response in the Fetus and Neonate. Chapter 128. In Fetal and Neonatal Phsiology, 5th ed.; Elsevier: Philadelphia, PA, USA, 2017; Volume 2, pp. 1241–1254.e4. [Google Scholar]
- Maines, M.D. Heme oxygenase: Function, multiplicity, regulatory mechanisms, and clinical applications. FASEB J. 1988, 2, 2557–2568, PMID:3290025. [Google Scholar] [CrossRef] [Green Version]
- Vijayan, V.; Sebastian, M.; Eveline, B.V.; Stephan, I. Heme oxygenase-1 as a therapeutic target in inflammatory disorders of the gastrointestinal tract. World J. Gastroenterol. 2010, 16, 3112–3119. [Google Scholar] [CrossRef]
- Chang, Y.S.; Jin, H.G.; Lee, H.; Lee, D.S.; Woo, E.R. Phytochemical constituents of the root bark from Morus alba and their IL-6 inhibitory activity. Nat. Prod. Sci. 2019, 25, 268–274. [Google Scholar] [CrossRef] [Green Version]
- Lim, H.J.; Jin, H.G.; Woo, E.R.; Lee, S.K.; Kim, H.P. The root barks of Morus alba and the flavonoid constituents inhibit airway inflammation. J. Ethnopharmacol. 2013, 149, 169–175. [Google Scholar] [CrossRef]
- Korhonen, R.; Lahti, A.; Kankaanranta, H.; Moilanen, E. Nitric oxide production and signaling in inflammation. Curr. Drug Targets Inflamm. Allergy 2005, 4, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Apple, K.; Honegger, P.; Gebicke-Haerter, P.J. Expression of interleukin-3 and tumor necrosis factor-beta mRNAs in cultured microglia. J. Neuroimmunol. 1995, 60, 83–91. [Google Scholar] [CrossRef]
- Nussler, A.K.; Billiar, T.R. Inflammation, immunoregulation, and inducible nitric oxide synthase. J. Leukoc. Biol. 1993, 54, 171–178. [Google Scholar] [CrossRef]
- Surh, Y.J.; Chun, K.S.; Cha, H.H.; Han, S.S.; Keum, Y.S.; Park, K.K.; Lee, S.S. Molecular mechanisms underlying chemopreventive activities of anti-inflammatory phytochemicals: Down-regulation of COX-2 and iNOS through suppression of NF-κB activation. Mutat. Res. 2001, 480, 243–268. [Google Scholar] [CrossRef]
- Palombella, V.J.; Rando, O.J.; Goldberg, A.L.; Maniatis, T. The ubiquitin-proteasome pathway is required for processing the NF-κappa B1 precursor protein and the activation of NF-κappaB. Cell 1994, 78, 773–785. [Google Scholar] [CrossRef]
- Walsh, D.A. Angoigenesis and arthritis. Rheumatology 1999, 38, 103–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, T.S.; Chau, L.Y. Heme oxygenase-1 mediates the anti-inflammatory effect of interleukin-10 in mice. Nat. Med. 2002, 8, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Maines, M.D. The heme oxygenase system: A regulator of second messenger gases. Annu. Rev. Pharmacol. Toxicol. 1997, 37, 517–554. [Google Scholar] [CrossRef]
- Oh, G.S.; Pae, H.O.; Chung, H.T. Nitric oxide priming protects nitric oxide-mediated apoptosis via heme oxygenase-1 induction. Free Radic. Biol. Med. 2003, 34, 1136–1145. [Google Scholar]
- Ko, W.; Lee, H.; Kim, N.; Jo, H.G.; Woo, E.R.; Lee, K.; Han, Y.S.; Park, S.R.; Ahn, G.; Cheong, S.H.; et al. The anti-oxidative and anti-neuroinflammatory effects of sargassum horneri by heme oxygenase-1 induction in BV2 and HT22 cells. Antioxidants 2021, 10, 859. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.S.; Park, C.M. Alleviated oxidative damage by taraxacum officinale through the induction of Nrf2-MAPK/PI3K mediated HO-1 activation in murine macrophages RAW 264.7 cell line. Biomolecules 2019, 9, 288. [Google Scholar] [CrossRef] [Green Version]
- Blantz, R.C.; Munger, K. Role of nitric oxide in inflammatory conditions. Nephron 2002, 90, 373–378. [Google Scholar] [CrossRef]
- Förstermann, U.; Sessa, W.C. Nitric oxide synthases: Regulation and function. Eur. Heart J. 2012, 33, 829–837. [Google Scholar] [CrossRef] [Green Version]
- Ying, L.; Hofseth, L.J. An emerging role for endothelial nitric oxide synthase in chronic inflammation and cancer. Cancer Res. 2007, 67, 1407–1410. [Google Scholar] [CrossRef] [Green Version]
- Morita, I. Distinct functions of COX-1 and COX-2. Prostaglandins Other Lipid Mediat. 2002, 68–69, 165–175. [Google Scholar] [CrossRef]
- Sales, K.J.; Jabbour, H.N. Cyclooxygenase enzymes and prostaglandins in pathology of the endometrium. Reproduction 2003, 126, 559–567. [Google Scholar] [CrossRef]
- Ricciotti, E.; FitzGerald, G.A. Prostaglandins and inflammation. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 986–1000. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.R.; Kim, H.G.; Ko, W.; Dong, L.; Yoo, D.; Oh, S.M.; Lee, Y.S.; Lee, D.S.; Baek, N.I.; Lee, D.Y. Noble 3,4-seco-triterpenoid glycosides from the fruits of Acanthopanax sessiliflorus and their anti-neuroinflammatory effects. Antioxidants 2021, 10, 1334. [Google Scholar] [CrossRef] [PubMed]
- Sharma, J.N.; Al-Omran, A.; Parvathy, S.S. Role of nitric oxide in inflammatory diseases. Inflammopharmacology 2007, 15, 252–259. [Google Scholar] [CrossRef]
- Chen, L. Current trends in PGE2 targeting for anti-inflammatory therapy. Pharm. Bioprocess. 2016, 4, 48–49. [Google Scholar]
- Ray, A.; Joshi, J. Cytokines and their role in health and disease: A brief overview. MOJ Immunol. 2016, 4, 00121. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.M.; An, J. Cytokines, inflammation, and pain. Int. Anesthesiol Clin. 2007, 45, 27–37. [Google Scholar] [CrossRef] [Green Version]
- Kany, S.; Vollrath, J.T.; Relja, B. Cytokines in inflammatory disease. Int. J. Mol. Sci. 2019, 20, 6008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toshio, T.; Masashi, N.; Tadamitsu, K. IL-6 in inflammation, immunity, and disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef]
- Ye, H.; Wang, Y.; Jenson, A.B.; Yan, J. Identification of inflammatory factor TNFα inhibitor from medicinal herbs. Exp. Mol. Pathol. 2016, 100, 307–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hana, Z.; Jan, H. TNF-α signalling and inflammation: Interactions between old acquaintances. Inflamm. Res. 2013, 62, 641–651. [Google Scholar] [CrossRef]
- Pahl, H.L. Activators and target genes of Rel/NF-kappaB transcription factors. Oncogene 1999, 18, 6853–6866. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Nie, M.; Fan, M.W.; Bian, Z. Anti-inflammatory activity of curcumin in macrophages stimulated by lipopolysaccharide from Porphyromonas gingivalis. Pharmacology 2008, 82, 264–269. [Google Scholar] [CrossRef]
- Lappas, M.; Permezel, M.; Georgiou, H.M.; Rice, G.E. Nuclear factor kappa B regulation of proinflammatory cytokines in human gestational tissues in vitro. Biol. Reprod. 2002, 67, 668–673. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Lim, J.; Kang, B.Y.; Jung, K.; Choi, H.J. Capillarisin augments anti-oxidative and anti-inflammatory responses by activating Nrf2/HO-1 signaling. Neurochem. Int. 2017, 105, 11–20. [Google Scholar] [CrossRef]
- Wu, B.; Wu, Y.; Tang, W. Heme catabolic pathway in inflammation and immune disorders. Front. Pharmacol. 2019, 10, 825. [Google Scholar] [CrossRef] [Green Version]
- Ko, W.; Sohn, J.H.; Jang, J.H.; Ahn, J.S.; Kang, D.G.; Lee, H.S.; Kim, J.S.; Kim, Y.C.; Oh, H. Inhibitory effects of alternaramide on inflammatory mediator expression through TLR4-MyD88-mediated inhibition of NF-κB and MAPK pathway signaling in lipopolysaccharide-stimulated RAW264.7 and BV2 cells. Chem. Biol. Interact. 2016, 244, 16–26. [Google Scholar] [CrossRef]
- Titheradge, M.A. The enzymatic measurement of nitrate and nitrite. Methods Mol. Biol. 1998, 100, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Liu, Z.; Yoon, C.S.; Dong, L.; Ko, W.; Woo, E.R.; Lee, D.S. Anti-neuroinflammatory and anti-inflammatory activities of phenylheptatriyne isolated from the flowers of coreopsis lanceolata L. via NF-κB inhibition and HO-1 expression in BV2 and RAW264.7 cells. Int. J. Mol. Sci. 2021, 22, 7482. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.C.; Yoon, C.S.; Quang, T.H.; Ko, W.; Kim, J.S.; Oh, H.; Kim, Y.C. Prenylated flavonoids from Cudrania tricuspidata suppress lipopolysaccharide-induced neuroinflammatory activities in BV2 microglial cells. Int. J. Mol. Sci. 2016, 17, 255. [Google Scholar] [CrossRef] [PubMed] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ko, W.; Liu, Z.; Kim, K.-W.; Dong, L.; Lee, H.; Kim, N.Y.; Lee, D.-S.; Woo, E.-R. Kuwanon T and Sanggenon a Isolated from Morus alba Exert Anti-Inflammatory Effects by Regulating NF-κB and HO-1/Nrf2 Signaling Pathways in BV2 and RAW264.7 Cells. Molecules 2021, 26, 7642. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26247642
Ko W, Liu Z, Kim K-W, Dong L, Lee H, Kim NY, Lee D-S, Woo E-R. Kuwanon T and Sanggenon a Isolated from Morus alba Exert Anti-Inflammatory Effects by Regulating NF-κB and HO-1/Nrf2 Signaling Pathways in BV2 and RAW264.7 Cells. Molecules. 2021; 26(24):7642. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26247642
Chicago/Turabian StyleKo, Wonmin, Zhiming Liu, Kwan-Woo Kim, Linsha Dong, Hwan Lee, Na Young Kim, Dong-Sung Lee, and Eun-Rhan Woo. 2021. "Kuwanon T and Sanggenon a Isolated from Morus alba Exert Anti-Inflammatory Effects by Regulating NF-κB and HO-1/Nrf2 Signaling Pathways in BV2 and RAW264.7 Cells" Molecules 26, no. 24: 7642. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26247642