
Cool Temperature Enhances Growth, Ferulic Acid and Flavonoid Biosynthesis While Inhibiting Polysaccharide Biosynthesis in Angelica sinensis



Abstract
:1. Introduction
2. Results
2.1. Effect of Temperatures on Growth Characteristics
2.2. Effect of Temperatures on Contents of Ferulic Acid, Flavonoids, Ligustilide, Volatile Oils and Polysaccharides
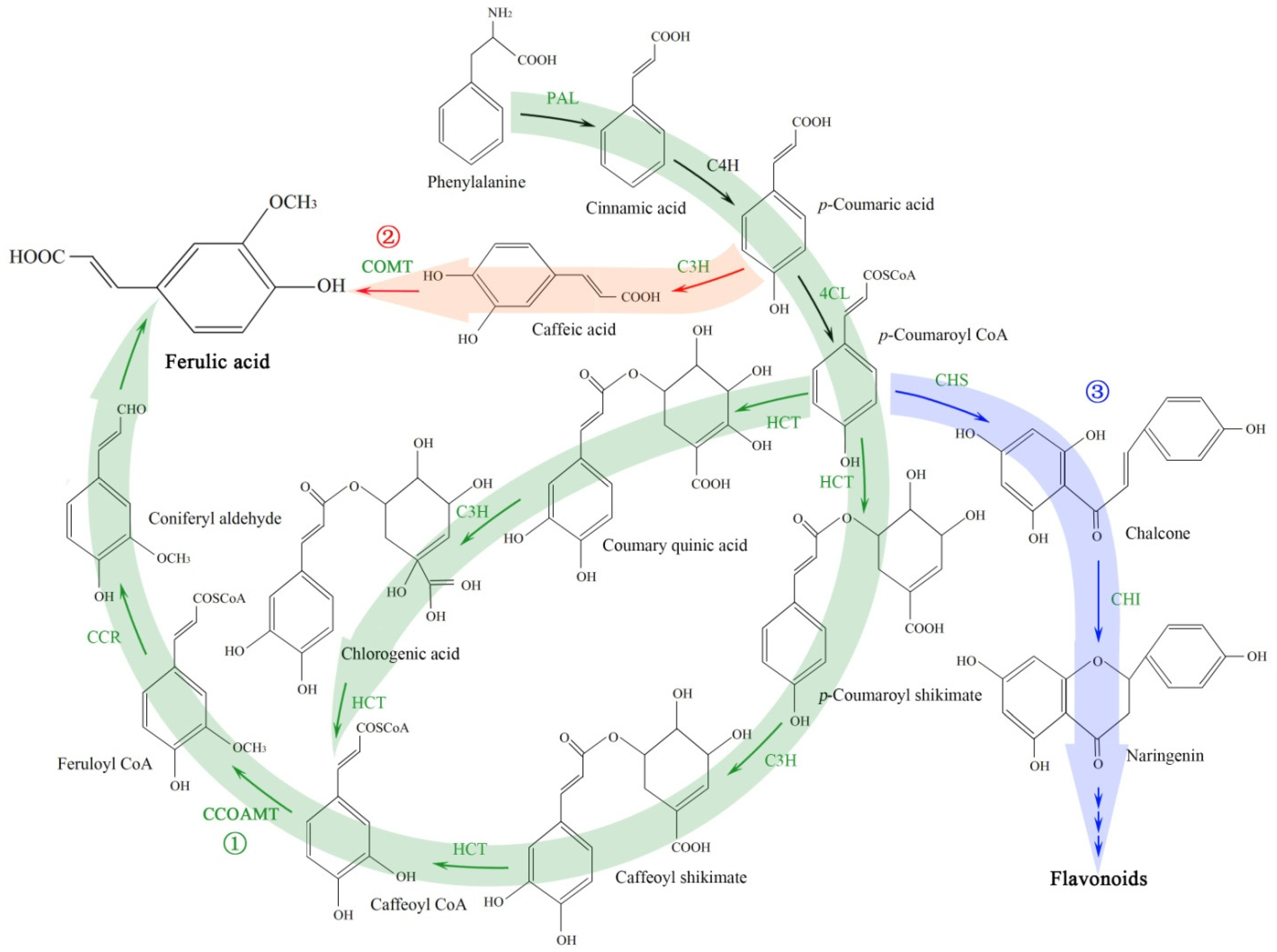
2.3. Effect of Temperatures on Gene Expression Related to Ferulic Acid and Flavonoid Biosynthesis
2.4. Effect of Temperatures on Gene Expression Related to Volatile Oils and Polysaccharide Biosynthesis
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Physiological Measurement
4.3. Metabolites Determination
4.3.1. Extracts Preparation
4.3.2. Determination of Ferulic Acid and Ligustilide Contents
4.3.3. Determination of Flavonoids Content
4.3.4. Determination of Polysaccharides Content
4.3.5. Volatile Oils Determination
4.4. Quantification of mRNA
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
Sample Availability
References
- Upton, R. American Herbal Pharmacopoeia and Therapeutic Compendium: Dang Gui Root-Angelica Sinensis (Oliv.); American Herbal Pharmacopoeia and Therapeutic Compendium: Scotts Valley, CA, USA, 2003; pp. 1–41. [Google Scholar]
- Zhang, H.Y.; Bi, W.G.; Yu, Y.; Liao, W.B. Angelica sinensis (Oliv.) Diels in China: Distribution, cultivation, utilization and variation. Genet. Resour. Crop Evol. 2012, 59, 607–613. [Google Scholar] [CrossRef]
- Chinese Pharmacopoeia Commission. Pharmacopoeia of the People’s Republic of China (2015 Version) Volume I; Chinese Medical Science Press: Beijing, China, 2020; p. 139. [Google Scholar]
- Wei, W.L.; Zeng, R.; Gu, C.M.; Qu, Y.; Huang, L.F. Angelica sinensis in China—A review of botanical profile, ethnopharmacology, phytochemistry and chemical analysis. J. Ethnopharmacol. 2016, 190, 116–141. [Google Scholar] [CrossRef]
- Hook, I.L.I. Danggui to Angelica sinensis root: Are potential benefits to European women lost in translation? A review. J. Ethnopharmacol. 2014, 152, 1–13. [Google Scholar] [CrossRef]
- Xu, X.Q.; Zhang, X.B.; Chen, J.; Zhao, W.L.; Jin, L. Study on ecological suitability of Angelica sinensis in Gansu Province. Chin. Tradit. Herb. Drugs 2020, 51, 3304–3307. [Google Scholar]
- Liang, F.; Li, J.; Zhang, W.; Zhang, R.X. Causes for change in producing areas of geo-authentic herbs. China J. Chin. Mat. Med. 2013, 38, 1649–1651. [Google Scholar]
- Huang, L.Q.; Jin, L. Suitable Technology for Production and Processing of Angelica sinensis; China Pharmaceutical Science and Technology Press: Beijing, China, 2018; pp. 1–14. [Google Scholar]
- Xu, X.Q.; Zhu, T.T.; Shi, T.T.; Chen, J.; Jin, L. Quality suitability regionalization analysis of Angelica sinensis in Gansu, China. PLoS ONE 2020, 15, 0243750. [Google Scholar] [CrossRef] [PubMed]
- Fan, Q.; Zhang, Y.Y.; Chen, H.P.; Lin, X.Y.; Xia, P.F.; Sun, Y.J.; Zhao, L.; Luo, J.; Yang, F.X.; Li, S.H. Influence of ecological factors on the quality of Angelica sinensis based on OPLS model. Chin. Tradit. Pat. Med. 2021, 43, 1226–1230. [Google Scholar]
- Jin, L.; Jin, Y.; Wang, H.Z.; Zhang, X.F.; Zhang, P.S.; Li, Y.D. Effects of altitude on the yield and content of ferulic acid in Angelica sinensis. J. Tradit. Chin. Vet. Med. 2014, 3, 47–49. [Google Scholar]
- Li, M.S.; Hao, Y.R.; Zhang, Z.G. Influence of the high altitude on the yield of Angelica sinensis and principal components of essential oils. Acta Bot. Boreali-Occidentalia Sin. 1985, 5, 155–160. [Google Scholar]
- Li, J.; Gao, S.F.; Li, Y.D.; Jin, L.; Wang, H.Z. Response study of ferulic and ligustilide in the root of Angelica sinensis on altitude. J. Gansu Coll. TCM 2014, 31, 23–26. [Google Scholar]
- Li, M.F.; Liu, X.Z.; Wei, J.H.; Zhang, Z.; Chen, S.J.; Liu, Z.H.; Xing, H. Selection of high altitude planting area of Angelica sinensis based on biomass, bioactive compounds accumulation and antioxidant capacity. Chin. Tradit. Herb. Drugs 2020, 51, 474–481. [Google Scholar]
- Li, M.F.; Kang, T.L.; Jin, L.; Wei, J.H. Research progress on bolting and flowering of Angelica sinensis and regulation pathways. Chin. Tradit. Herb. Drugs 2020, 51, 5894–5899. [Google Scholar]
- Ma, Y.; Jin, L.; Wang, Z.H.; Cui, Z.J.; Zhang, Y.L.; Li, Y.D. Comparative study of cultivated radix Angelica sinensis polysaccharide content from different altitude in Min county of Gansu province. J. Tradit. Chin. Vet. Med. 2013, 2, 7–9. [Google Scholar]
- Wang, H.Z.; Jin, L.; Zhang, E.H. Effect of altitude on the photosynthate accumulation and distribution pattern of Angelica sinensis. J. Chin. Med. Mater. 2012, 35, 1191–1194. [Google Scholar]
- Wang, H.Z.; Jin, L.; Zhang, E.H. Effect of altitude on ferulic acid in Angelica sinensis and analysis in key factors. Chin. Tradit. Herb. Drugs 2013, 44, 219–223. [Google Scholar]
- Lin, H.M.; Wu, Y.A.; Cao, Z.F.; Lv, S.L.; Mao, X.J. Influence of sun shade cultivation on premature bolting in Angelica sinensis and growth environment factors. Chin. J. Exp. Tradit. Med. Form. 2010, 16, 79–83. [Google Scholar]
- Xiang, H.; Li, Y.; Jin, Y. Effects of light intensity on yield and ferulic acid content in Angelica sinensis. J. Tradit. Chin. Vet. Med. 2015, 2, 53–54. [Google Scholar]
- Wang, Z.H.; Wang, Y.Q.; Fan, Q.; Yang, X.; Luo, J.; Li, S.N.; Jia, M.T. Effects of UV-B radiation on the content of phthalides in Angelica sinensis. J. Gansu Coll. TCM 2019, 36, 31–34. [Google Scholar]
- Liu, B.; Rao, B.Y.; Qian, C.; Wu, Z.J. Effect on different irrigation treatments on growth, yield and quality of Angelica sinensis. Mod. Agric. Sci. Tech. 2013, 2, 77–82. [Google Scholar]
- Bohnert, H.J.; Nguyen, H.; Lewis, N.G. Bioengineering and Molecular Biology of Plant. Pathways; Science Press: Beijing, China, 2008; pp. 385–401. [Google Scholar]
- Liu, J.; Li, W.J.; Wang, C.M.; Xie, H.M. Biosynthesis and regulation of ferulic acid in Angelica sinensis. Chin. Tradit. Herb. Drugs 2008, 39, 1909–1912. [Google Scholar]
- Turi, C.E.; Murch, S.J. Targeted and untargeted phytochemistry of Ligusticum canbyi: Indoleamines, phthalides, antioxidant potential, and use of metabolomics as a hypothesis-generating technique for compound discovery. Planta Med. 2013, 79, 1370–1379. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Xu, J.; Li, Y.C.; Dai, Y.T.; Zhang, S.P.; Wang, G.; Liu, Z.G.; Dong, L.L.; Chen, S.L. Integrated chemical and transcriptomic analyses unveils synthetic characteristics of different medicinal root parts of Angelica sinensis. Chin. Herb. Med. 2019, 12, 19–28. [Google Scholar] [CrossRef]
- Liang, F.; Li, J.; Zhang, W.; Zhang, R.X. Brief introduction of geo-authentic herbs. China J. Chin. Mat. Med. 2013, 38, 1455–1457. [Google Scholar]
- Yang, D.L.; Sun, P.; Li, M.F. Chilling temperature stimulates growth, gene over-expression and podophyllotoxin biosynthesis in Podophyllum hexandrum Royle. Plant Physiol. Biol. 2016, 107, 197–203. [Google Scholar] [CrossRef]
- Yu, K.W.; Murthy, H.N.; Hahn, E.J.; Paek, K.Y. Ginsenoside production by hairy root cultures of Panax ginseng: Influence of temperature and light quality. Biol. Eng. J. 2005, 23, 53–56. [Google Scholar] [CrossRef]
- Su, H.Y.; Li, J.; Chen, S.J.; Sun, P.; Xing, H.; Yang, D.L.; Zhang, X.N.; Li, M.F.; Wei, J.H. Physiological and transcriptomic analysis provide insight into low temperature enhancing hypericin biosynthesis in Hypericum perforatum. Molecules 2021, 26, 2294. [Google Scholar] [CrossRef]
- Ramakrishna, A.; Ravishankar, G.A. Influence of abiotic stress signals on secondary metabolites in plants. Plant Signal. Behav. 2011, 6, 1720–1731. [Google Scholar]
- Dixon, R.A.; Paiva, N.L. Stress-induced phenylpropanoid metabolism. Plant Cell 1995, 7, 1085–1097. [Google Scholar] [CrossRef]
- Kienow, L.; Schneider, K.; Bartsch, M.; Stuible, H.P.; Weng, H.; Miersch, O.; Wasternack, C.; Kombrink, E. Jasmonates meet fatty acids: Functional analysis of a new acyl-coenzyme A synthetase family from Arabidopsis thaliana. J. Exp. Bot. 2008, 59, 403–419. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Yao, R.; Zhao, Y.; Xu, S.; Huang, C.; Luo, J.; Kong, L. Cloning, functional characterization and site-directed mutagenesis of 4-coumarate: Coenzyme a ligase (4CL) involved in coumarin biosynthesis in Peucedanum praeruptorum Dunn. Front. Plant Sci. 2017, 8, 4. [Google Scholar] [CrossRef] [Green Version]
- Griesser, M.; Vitzthum, F.; Fink, B.; Bellido, M.L.; Raasch, C.; Munoz-Blanco, J.; Schwab, W. Multi-substrate flavonol O-glucosyltransferases from strawberry (Fragaria × ananassa) achene and receptacle. J. Exp. Bot. 2008, 59, 2611–2625. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.J.; Huhman, D.; Sumner, L.W.; Dixon, R.A. Regiospecific hydroxylation of isoflavones by cytochrome p450 81E enzymes from Medicago truncatula. Plant J. 2003, 36, 471–484. [Google Scholar] [CrossRef] [Green Version]
- Koeduka, T.; Baiga, T.J.; Noel, J.P.; Pichersky, E. Biosynthesis of t-anethole in anise: Characterization of t-anol/isoeugenol synthase and an O-methyltransferase specific for a C7-C8 propenyl side chain. Plant Physiol. 2009, 149, 384–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dudareva, N.; D’Auria, J.C.; Nam, K.H.; Raguso, R.A.; Pichersky, E. Acetyl-CoA: Benzylalcohol acetyltransferase—An enzyme involved in floral scent production in Clarkia breweri. Plant J. 1998, 14, 297–304. [Google Scholar] [CrossRef] [Green Version]
- Miron, D.; Schaffer, A.A. Sucrose phosphate synthase, sucrose synthase, and invertase activities in developing fruit of Lycopersicon esculentum Mill. and the sucrose accumulating Lycopersicon hirsutum Humb. and Bonpl. Plant Physiol. 1991, 95, 623–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visser, R.G.; Stolte, A.; Jacobsen, E. Expression of a chimaeric granule-bound starch synthase-GUS gene in transgenic potato plants. Plant Mol. Biol. 1991, 17, 691–699. [Google Scholar] [CrossRef] [PubMed]
- Date, K.; Satoh, A.; Iida, K.; Ogawa, H. Pancreatic α-Amylase controls glucose assimilation by duodenal retrieval through N-Glycan-specific binding, endocytosis, and degradation. J. Biol. Chem. 2015, 290, 17439–17450. [Google Scholar] [CrossRef] [Green Version]
- Deng, X.Q.; Guan, X.J.; Huang, N.N.; Li, J. Simultaneous determination of eight constituents in Angelica sinensis by HPLC. Chin. Tradit. Pat. Med. 2020, 42, 2075–2079. [Google Scholar]
- Lay, M.M.; Karsani, S.A.; Mohajer, S.; Abd Malek, S.N. Phytochemical constituents, nutritional values, phenolics, flavonols, flavonoids, antioxidant and cytotoxicity studies on Phaleria macrocarpa (Scheff.) Boerl fruits. BMC Complement. Altern. Med. 2014, 14, 152. [Google Scholar] [CrossRef] [Green Version]
- Lv, M.; Su, H.Y.; Li, M.L.; Yang, D.L.; Yao, R.Y.; Li, M.F.; Wei, J.H. Effect of UV-B radiation on growth, flavonoid and podophyllotoxin accumulation, and related gene expression in Sinopodophyllum hexandrum. Plant Biol. 2021, 23, 202–209. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Cao, X.L.; Li, M.L.; Li, J.; Song, Y.X.; Zhang, X.N.; Yang, D.L.; Li, M.F.; Wei, J.H. Co-expression of hydrolase genes improves seed germination of Sinopodophyllum hexandrum. Ind. Crops Prod. 2021, 164, 113414. [Google Scholar] [CrossRef]
- Zheng, W.R.; Tao, Y.H.; Shi, J.; Shao, J.; Xiao, W. Optimize the technique of extracting volatile oil from Angelica sinensis by steam distillation. Mod. Chin. Med. 2013, 15, 879–882. [Google Scholar]
- Li, J.; Li, M.L.; Zhu, T.T.; Zhang, X.N.; Li, M.F.; Wei, J.H. Integrated transcriptomics and metabolites at different growth stages reveals the regulation mechanism of bolting and flowering of Angelica sinensis. Plant Biol. 2021, 23, 574–582. [Google Scholar] [CrossRef]
- Li, M.F.; Li, J.; Wei, J.H.; Paré, P.W. Transcriptional controls for early bolting and flowering in Angelica sinensis. Plants 2021, 10, 1931. [Google Scholar] [CrossRef]
- Willems, E.; Leyns, L.; Vandesompele, J. Standardization of real-time PCR gene expression data from independent biological replicates. Anal. Biol. 2008, 379, 127–129. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Accession No. | Sequences (5′ to 3′) | Amplicon Size (bp) |
|---|---|---|---|
| ACT | [26] | Forward: TGGTATTGTGCTGGATTCTGGT | 109 |
| Reverse: TGAGATCACCACCAGCAAGG | |||
| Ferulic acid Biosynthesis | |||
| PAL1 | XM_017399483.1 | Forward: GGACTTGACAGTAGGGCAG | 146 |
| Reverse: CCCCGTAACTATCCGTTCCTT | |||
| 4CLL4 | XM_017376722.1 | Forward: AAGCAGTGTTTCAGAGGCAG | 105 |
| Reverse: GCTGAGCGCGGTATTGAGTT | |||
| 4CLL5 | XM_017388768.1 | Forward: CGGGACGAGTAAAGGAGTGG | 171 |
| Reverse: AGCGTTGCTACAAACCAAGC | |||
| 4CLL7 | KJ531407.1 | Forward: TGCTCCGTTGGGTAGAGAGT | 164 |
| Reverse: CTCCAGGCACAAGCATTCCT | |||
| 4CLL9 | XM_017397573.1 | Forward: GGTGGGGAAGCTAACAGGTC | 183 |
| Reverse: TCGCCAGTTCTTAACCAGCC | |||
| HCT | XM_017397289.1 | Forward: CCGGTGACATATCTGCGTGT | 171 |
| Reverse: GCGGAATGGCAATGGAAAGG | |||
| C3H | [26] | Forward: CAATCCAAGTTGACGACGAA | 119 |
| Reverse: CGAAGGCGAAACATAGGC | |||
| CCOAMT | AY620245.1 | Forward: TCGGCTACGACAACACCCTA | 157 |
| Reverse: TCGCCAACAGGAAGCATACA | |||
| CCR1 | XM_017403617.1 | Forward: CCATTCATGGATGCGTTGGT | 135 |
| Reverse: CCACACGTCTCACATTGGCT | |||
| COMT1 | XM_010673030.2 | Forward: TGGCGGAAAGGTAGTCGTTG | 130 |
| Reverse: TTCAGTCCTCTCACTTCCGC | |||
| Flavonoid Biosynthesis | |||
| CHS | KP726914.1 | Forward: GCAAAGACGCTGCATCCAAA | 126 |
| Reverse: GGAGCTTGGTGAGCTGGTAG | |||
| CHI | XM_017365109.1 | Forward: GTGTTTCCCCAGCTGCAAAG | 102 |
| Reverse: TTCCGACTTCTGCTTTCCCA | |||
| I3′H | XM_017363227.1 | Forward: GGCCACCTTCACCTCATCAA | 173 |
| Reverse: GGGCGGTCAGCTAAAACAAC | |||
| GT6 | XM_017383880.1 | Forward: TTCGGTGCCCATCACAAGAA | 166 |
| Reverse: AATCCTCCGACAGATGCGTG | |||
| Volatile oils Biosynthesis | |||
| AIMT1 | B8RCD3.1 | Forward: CGCTAGTCTTTTGAGCGAAGC | 119 |
| Reverse: CATGGGCACCTCCTACATCC | |||
| BEAT | O64988.1 | Forward: GATCAAGCCAGCAGTGATGC | 147 |
| Reverse: ACTTCAACACGTGTAGGCCG | |||
| Polysaccharide Biosynthesis | |||
| SUS1 | XM_017363708.1 | Forward: ATGAAGTCCACACAGGAAGCC | 124 |
| Reverse: CGACGACAAGGTGATGAGTG | |||
| Amy2 | V00718.1 | Forward: TCTTCTGAGCCCTGGAGTGT | 117 |
| Reverse: TCCAGGGAAGCCTCATGGAT | |||
| WAXY | AJ006293.1 | Forward: GCACTCATCCTCCATTCAGAG | 167 |
| Reverse: TCCGTTACTGATCCACCAGC | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, H.; Li, M.; Jin, L.; Xie, X.; Li, M.; Wei, J. Cool Temperature Enhances Growth, Ferulic Acid and Flavonoid Biosynthesis While Inhibiting Polysaccharide Biosynthesis in Angelica sinensis. Molecules 2022, 27, 320. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27010320
Dong H, Li M, Jin L, Xie X, Li M, Wei J. Cool Temperature Enhances Growth, Ferulic Acid and Flavonoid Biosynthesis While Inhibiting Polysaccharide Biosynthesis in Angelica sinensis. Molecules. 2022; 27(1):320. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27010320
Chicago/Turabian StyleDong, Han, Meiling Li, Ling Jin, Xiaorong Xie, Mengfei Li, and Jianhe Wei. 2022. "Cool Temperature Enhances Growth, Ferulic Acid and Flavonoid Biosynthesis While Inhibiting Polysaccharide Biosynthesis in Angelica sinensis" Molecules 27, no. 1: 320. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27010320