
Fucoidan and Alginate from the Brown Algae Colpomenia sinuosa and Their Combination with Vitamin C Trigger Apoptosis in Colon Cancer


Abstract
:1. Introduction
2. Results
2.1. Physicochemical Properties and Monosaccharide Composition of Fucoidan and Alginate
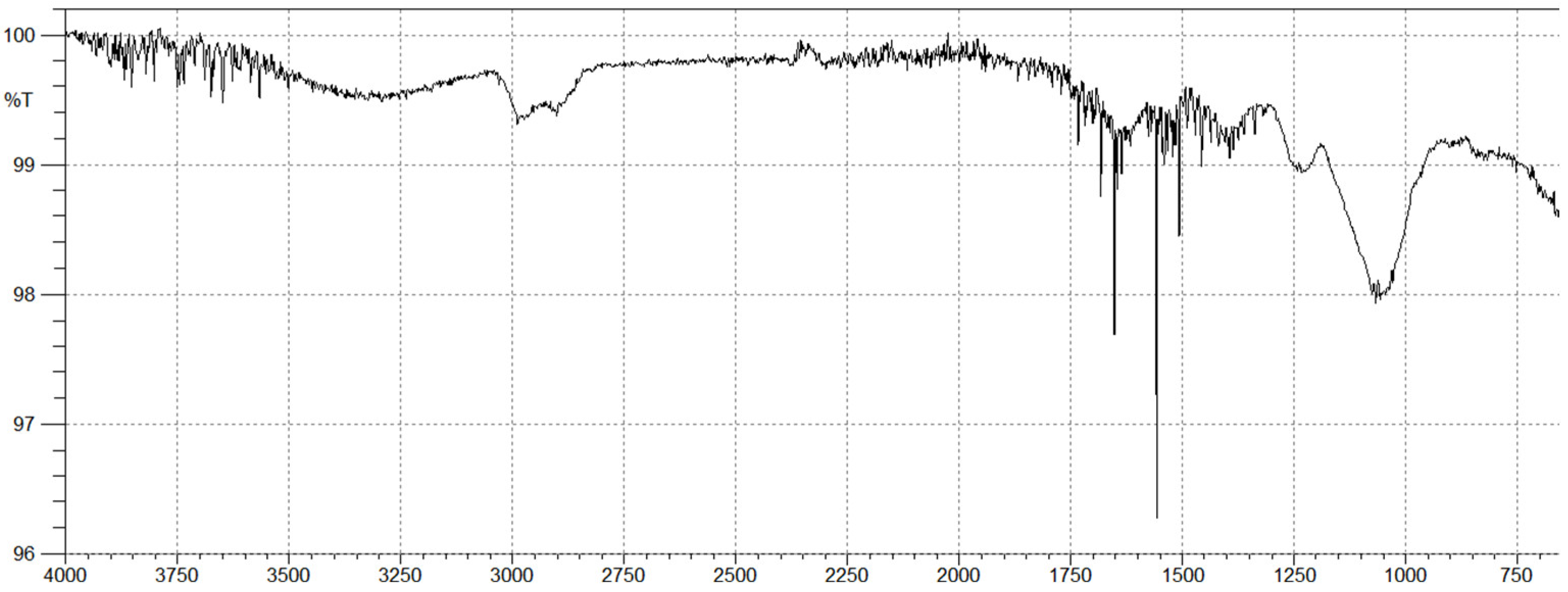
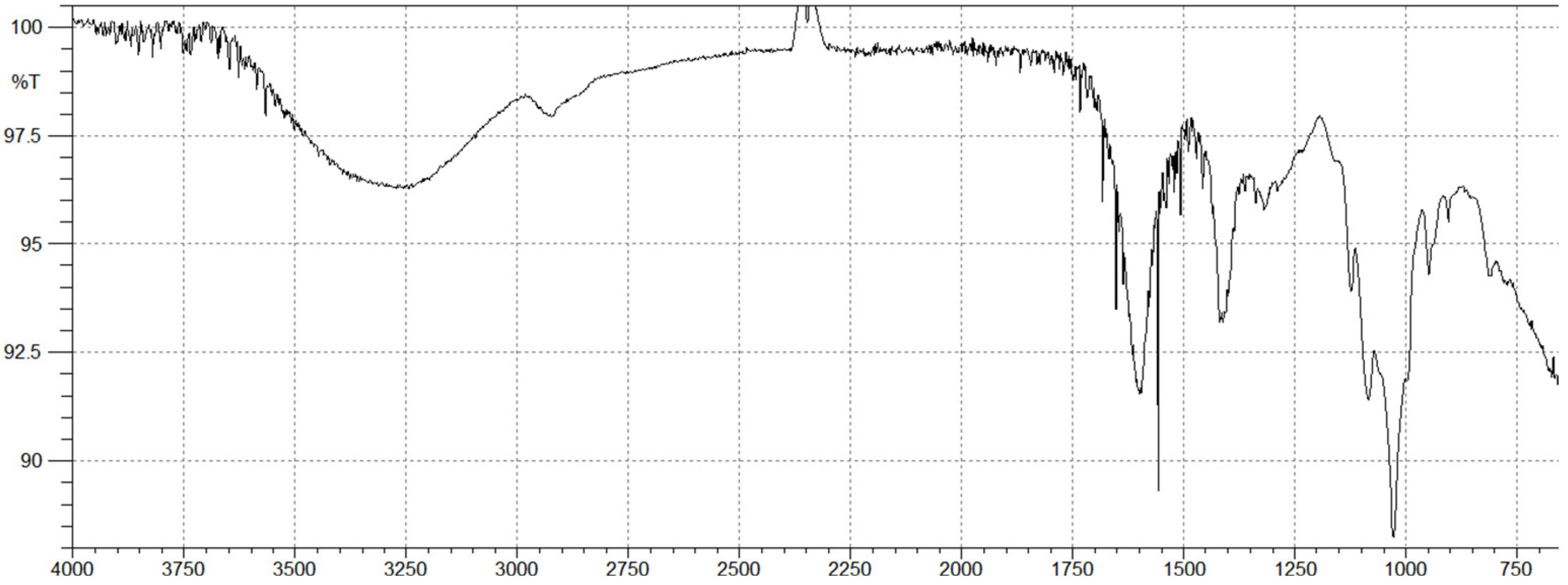
2.2. Brown Algal Polysaccharides Structural Analysis by FTIR
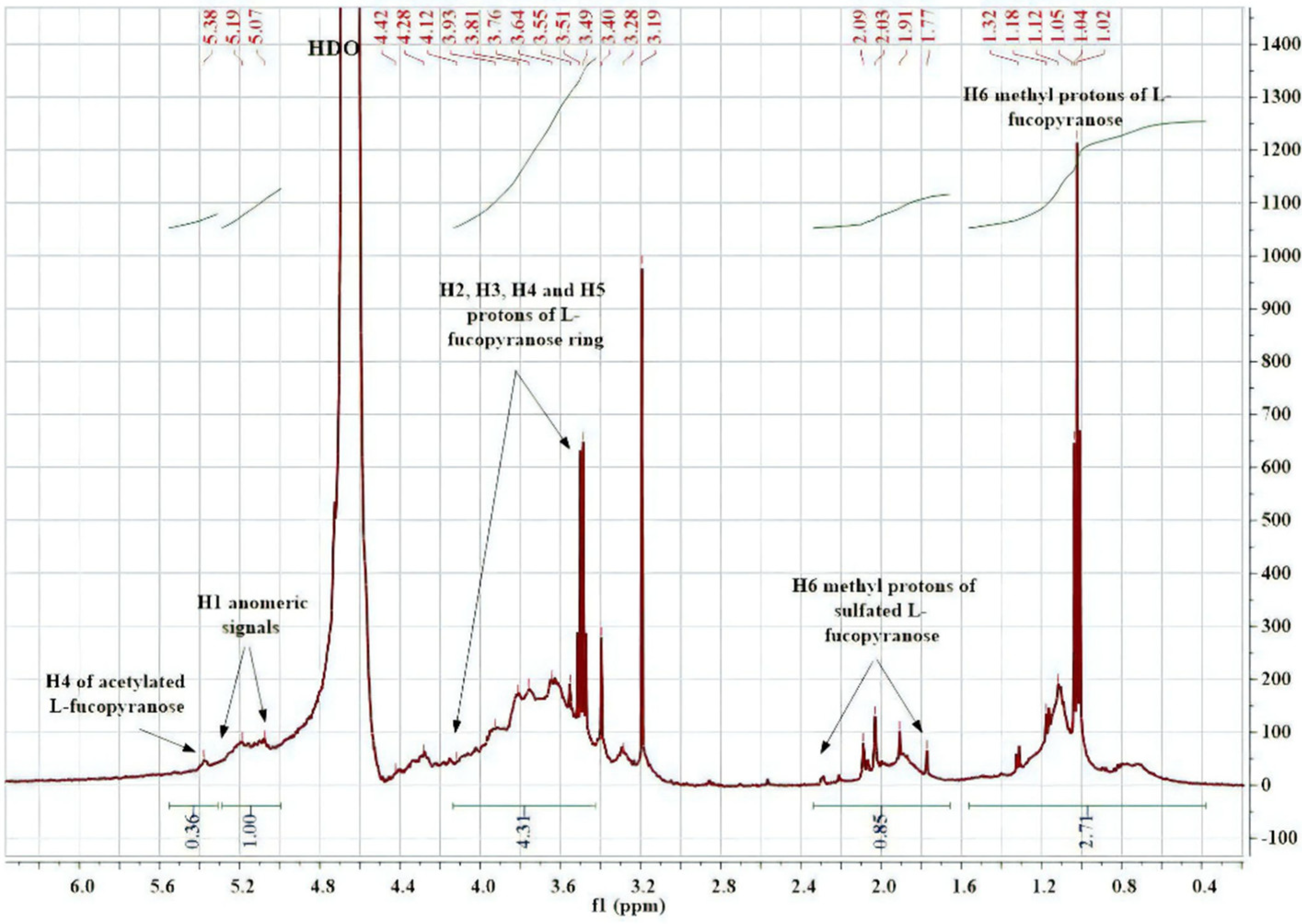
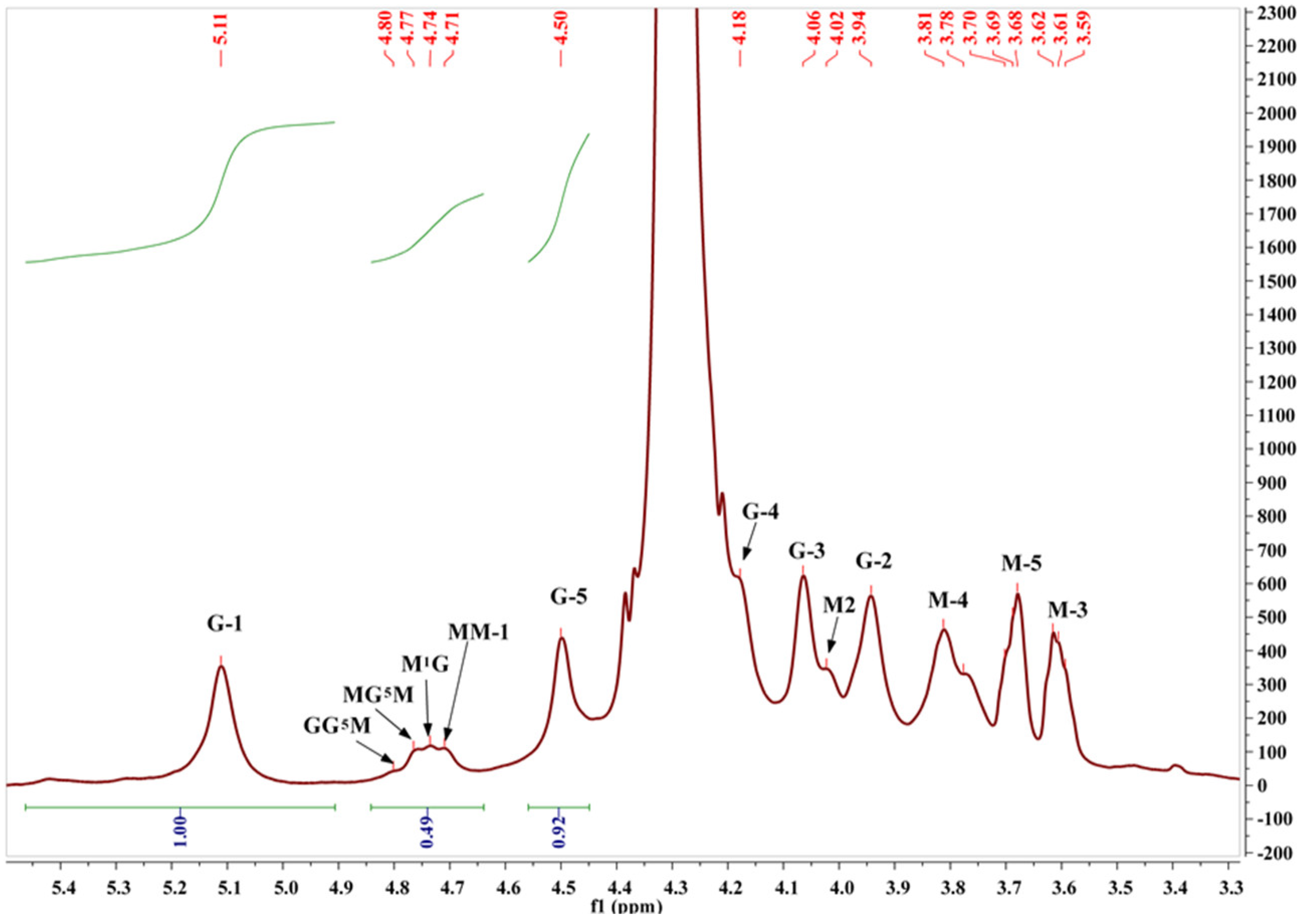
2.3. Structural Characterization of Algal Polysaccharides by NMR Spectroscopy
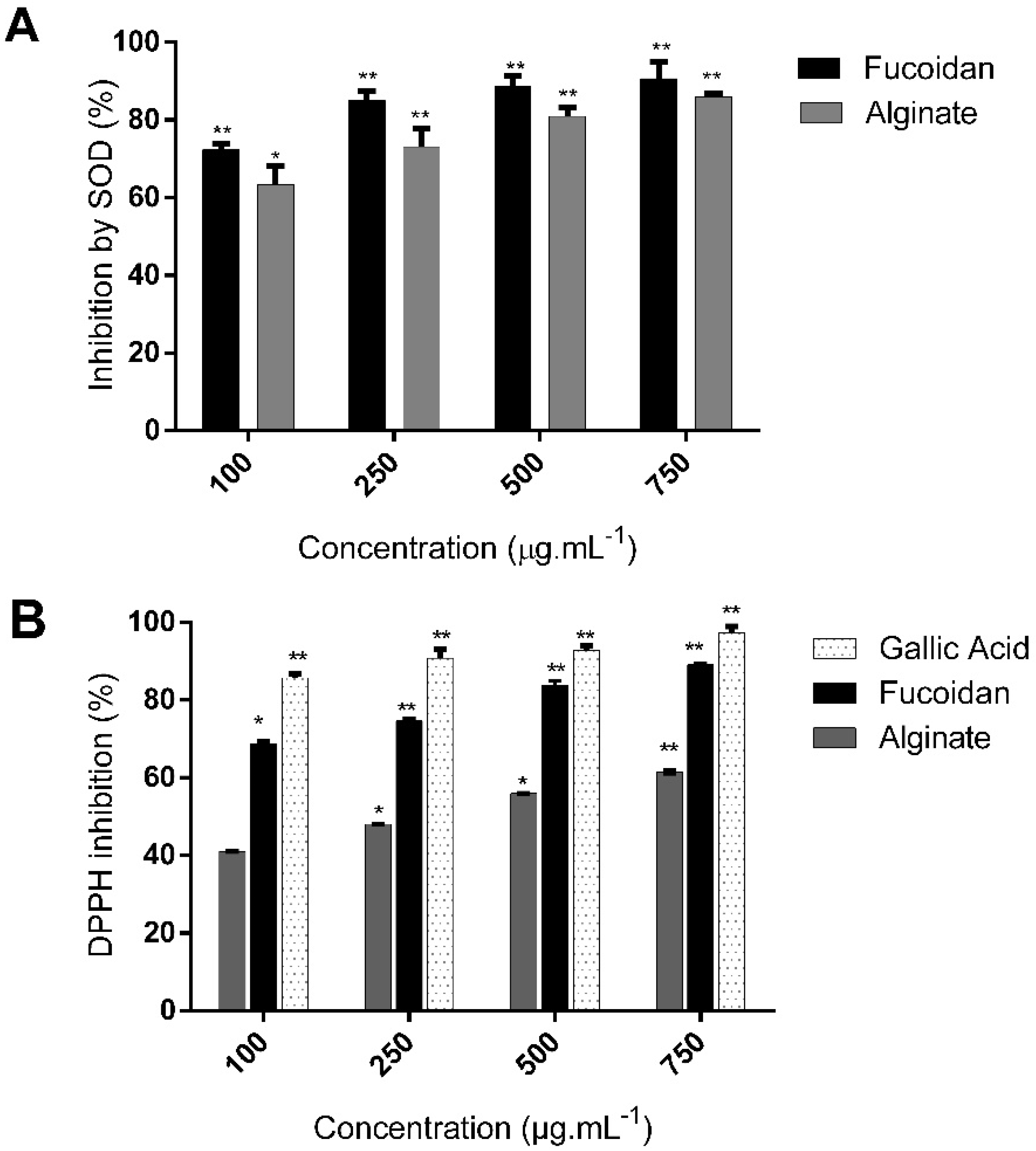
2.4. C. sinuosa Polysaccharides Exhibited Antioxidant Activities
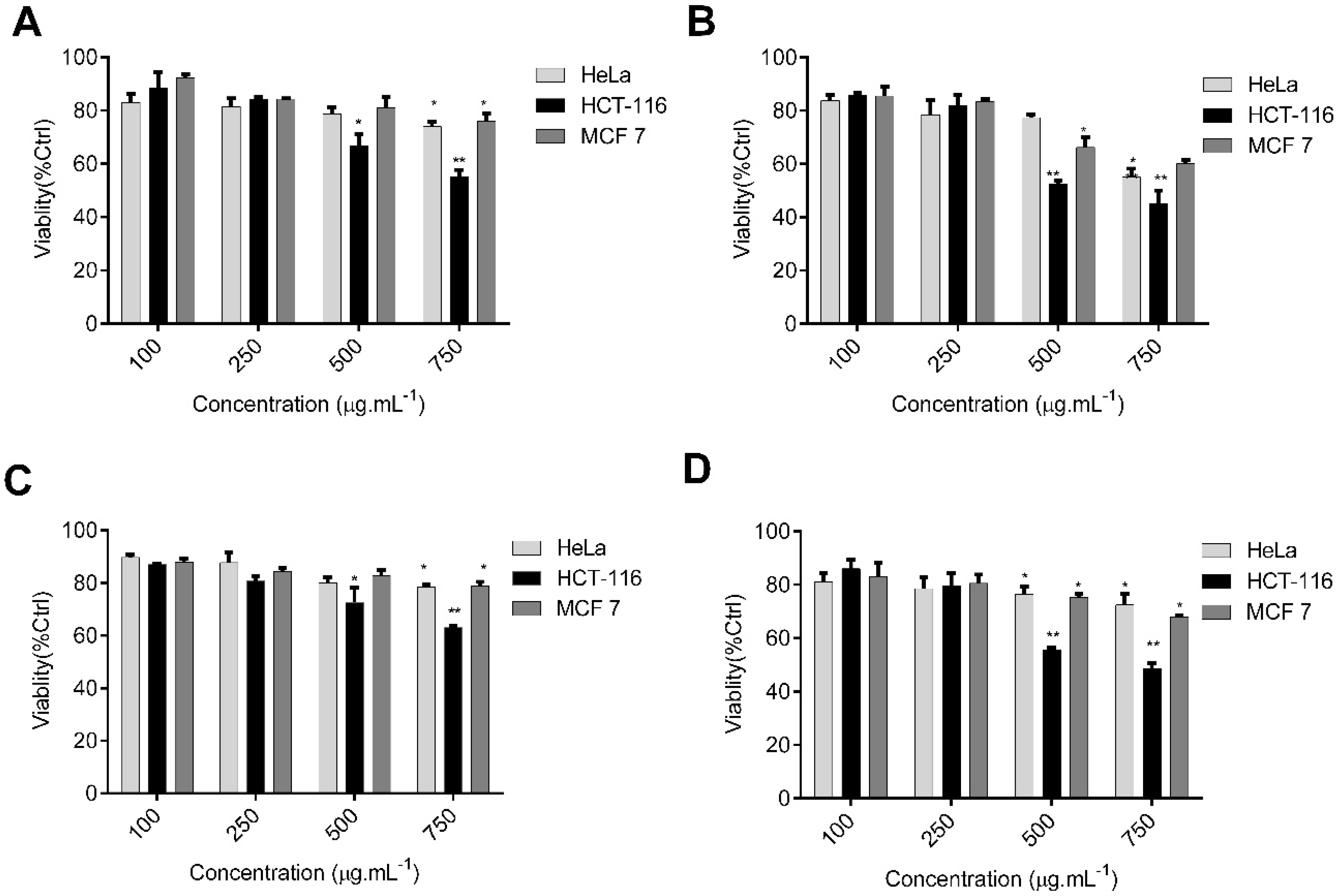
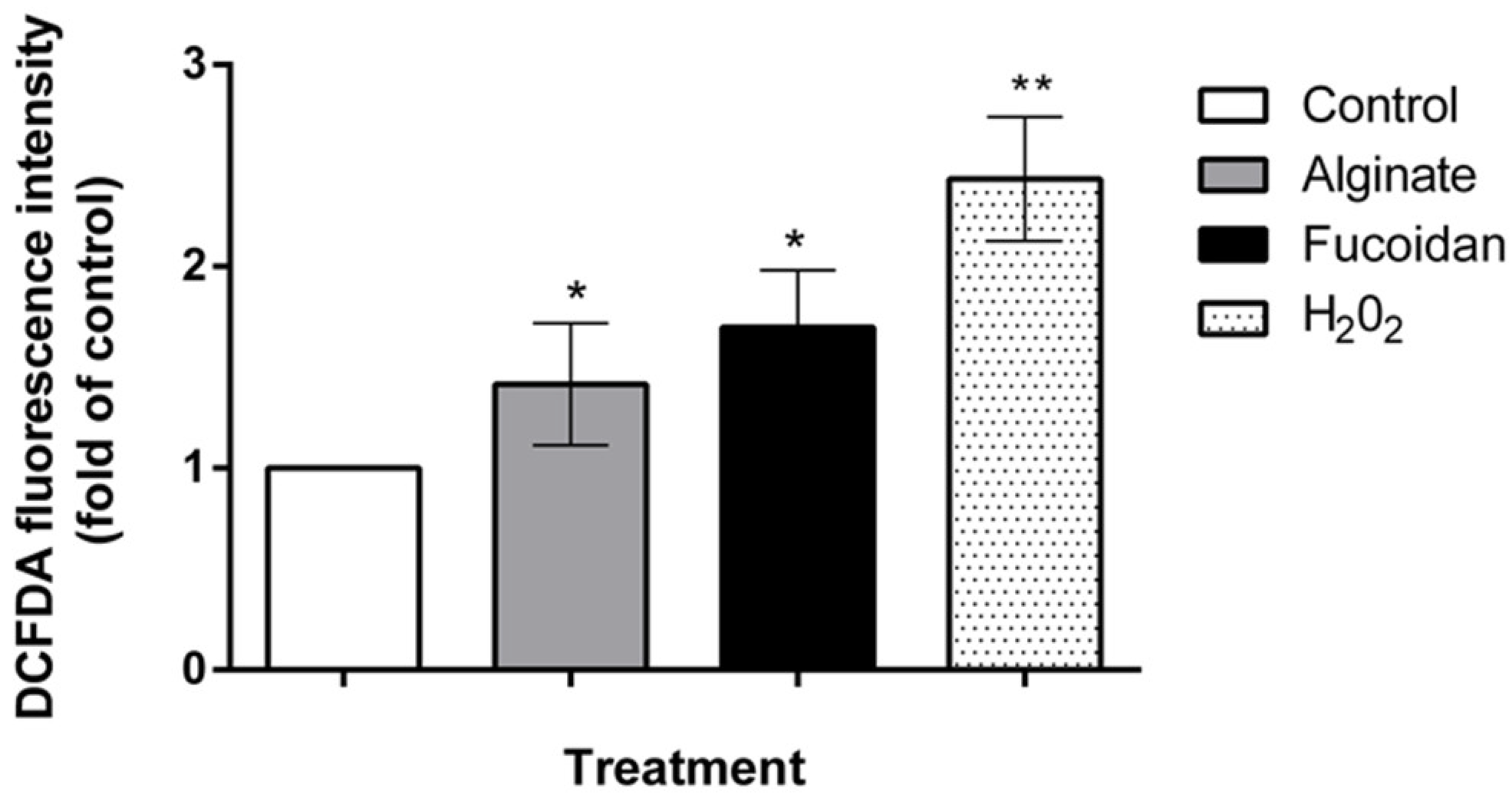
2.5. C. sinuosa Polysaccharides Showed Potent Antitumor and ROS-Inducing Effects against HCT-116 Colon Cancer Cells
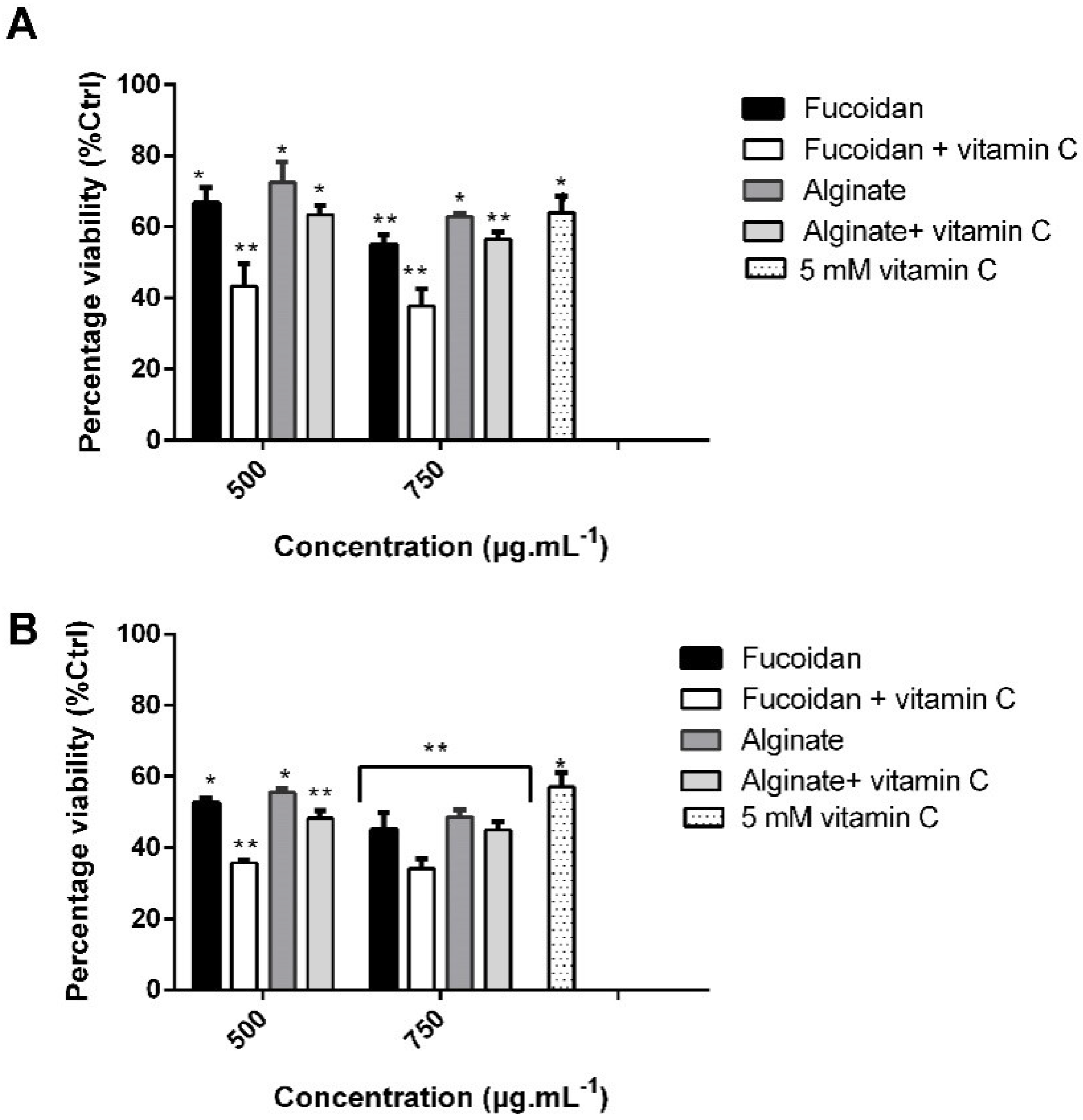
2.6. Combination Treatment of Vitamin C and C. sinuosa Polysaccharides Significantly Increase the Cytotoxic Activity against HCT-116 Cancer Cells
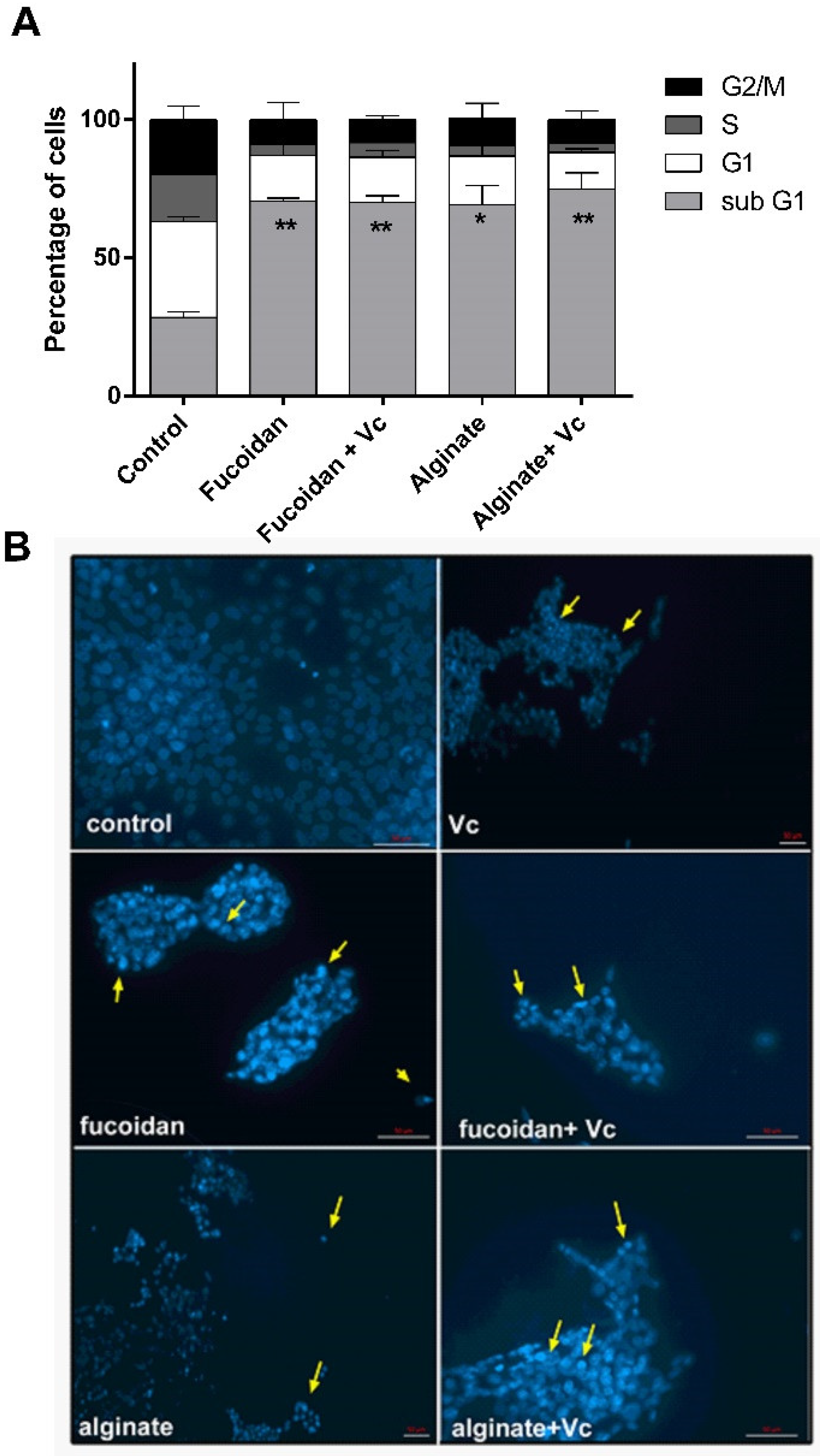
2.7. Combination of Vitamin C and Algal Polysaccharides Trigger Cell Cycle Regulation and Morphological Alterations in HCT-116 Cells
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. Extraction of Polysaccharides from C. sinuosa
4.2.1. Fucoidan Isolation
4.2.2. Alginate Isolation
4.2.3. Fucoidan and Alginate Purification
4.3. Physicochemical Properties
4.4. Monosaccharide Composition
4.5. Fourier Transform Infrared Spectroscopy (FTIR) Analysis
4.6. Nuclear Magnetic Resonance (NMR) Spectroscopy
4.7. Preparation of Polysaccharides
4.8. Antioxidant Assays
4.8.1. DPPH Free Radical Scavenging Assay
4.8.2. Superoxide Dismutase (SOD) Inhibition Assay
4.9. Cell Lines and Culture
4.10. Cell Viability Assay
4.11. Quantitative Determination of ROS
4.12. Cell Cycle Analysis
4.13. DAPI Staining
4.14. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Khalifa, S.A.M.; Elias, N.; Farag, M.A.; Chen, L.; Saeed, A.; Hegazy, M.-E.F.; Moustafa, M.S.; Abd El-Wahed, A.; Al-Mousawi, S.M.; Musharraf, S.G.; et al. Marine Natural Products: A Source of Novel Anticancer Drugs. Mar. Drugs 2019, 17, 491. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, C.; Neves, N.M.; Reis, R.L.; Martins, A.; Silva, T.H. A Review on Fucoidan Antitumor Strategies: From a Biological Active Agent to a Structural Component of Fucoidan-Based Systems. Carbohydr. Polym. 2020, 239, 116131. [Google Scholar] [CrossRef] [PubMed]
- Dobrinčić, A.; Balbino, S.; Zorić, Z.; Pedisić, S.; Bursać Kovačević, D.; Elez Garofulić, I.; Dragović-Uzelac, V. Advanced Technologies for the Extraction of Marine Brown Algal Polysaccharides. Mar. Drugs 2020, 18, E168. [Google Scholar] [CrossRef] [Green Version]
- Hsu, H.-Y.; Hwang, P.-A. Clinical Applications of Fucoidan in Translational Medicine for Adjuvant Cancer Therapy. Clin. Trans. Med. 2019, 8, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbosa, A.I.; Coutinho, A.J.; Costa Lima, S.A.; Reis, S. Marine Polysaccharides in Pharmaceutical Applications: Fucoidan and Chitosan as Key Players in the Drug Delivery Match Field. Mar. Drugs 2019, 17, 654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isnansetyo, A.; Lutfia, F.; Nursid, M.; Nursid, M.; Susidarti, R. Cytotoxicity of Fucoidan from Three Tropical Brown Algae against Breast and Colon Cancer Cell Lines. Pharmacogn. J. 2017, 9, 14–20. [Google Scholar] [CrossRef] [Green Version]
- Ikeguchi, M.; Yamamoto, M.; Aral, Y.; Maeta, Y.; Ashida, K.; Katano, K.; Miki, Y.; Kimura, T. Fucoidan Reduces the Toxicities of Chemotherapy for Patients with Unresectable Advanced or Recurrent Colorectal Cancer. Oncol. Lett. 2011, 2, 319–322. [Google Scholar] [CrossRef] [PubMed]
- Burney, M.; Mathew, L.; Gaikwad, A.; Nugent, E.K.; Gonzalez, A.O.; Smith, J.A. Evaluation Fucoidan Extracts From Undaria Pinnatifida and Fucus Vesiculosus in Combination With Anticancer Drugs in Human Cancer Orthotopic Mouse Models. Integr. Cancer Ther. 2018, 17, 755–761. [Google Scholar] [CrossRef] [Green Version]
- Oh, B.; Kim, J.; Lu, W.; Rosenthal, D. Anticancer Effect of Fucoidan in Combination with Tyrosine Kinase Inhibitor Lapatinib. Evid.-Based Complement. Altern. Med. 2014, 2014, 865375. [Google Scholar] [CrossRef]
- Fernando, I.P.S.; Kim, D.; Nah, J.-W.; Jeon, Y.-J. Advances in Functionalizing Fucoidans and Alginates (Bio)Polymers by Structural Modifications: A Review. Chem. Eng. 2019, 355, 33–48. [Google Scholar] [CrossRef]
- Gomaa, M.; Hifney, A.; Fawzy, M.; Abdel-Gawad, K. Use of Seaweed and Filamentous Fungus Derived Polysaccharides in the Development of Alginate-Chitosan Edible Films Containing Fucoidan: Study of Moisture Sorption, Polyphenol Release and Antioxidant Properties. Food Hydrocoll. 2018, 82, 239–247. [Google Scholar] [CrossRef]
- Rhein-Knudsen, N.; Ale, M.; Ajalloueian, F.; Meyer, A. Characterization of Alginates from Ghanaian Brown Seaweeds: Sargassum spp. and Padina spp. Food Hydrocoll. 2017, 71, 236–244. [Google Scholar] [CrossRef]
- Das, R.K.; Kasoju, N.; Bora, U. Encapsulation of Curcumin in Alginate-Chitosan-Pluronic Composite Nanoparticles for Delivery to Cancer Cells. Nanomed. Nanotechnol. Biol. Med. 2010, 6, 153–160. [Google Scholar] [CrossRef]
- Ivanovska, J.; Zehnder, T.; Lennert, P.; Sarker, B.; Boccaccini, A.R.; Hartmann, A.; Schneider-Stock, R.; Detsch, R. Biofabrication of 3D Alginate-Based Hydrogel for Cancer Research: Comparison of Cell Spreading, Viability, and Adhesion Characteristics of Colorectal HCT116 Tumor Cells. Tissue Eng. Part C Methods 2016, 22, 708–715. [Google Scholar] [CrossRef] [PubMed]
- Sorasitthiyanukarn, F.N.; Muangnoi, C.; Bhuket, P.R.N.; Rojsitthisak, P.; Rojsitthisak, P. Pranee Chitosan/Alginate Nanoparticles as a Promising Approach for Oral Delivery of Curcumin Diglutaric Acid for Cancer Treatment. Mater. Sci. Eng. C 2018, 93, 178–190. [Google Scholar] [CrossRef]
- Szekalska, M.; Puciłowska, A.; Szymańska, E.; Ciosek, P.; Winnicka, K. Alginate: Current Use and Future Perspectives in Pharmaceutical and Biomedical Applications. Int. J. Polym. Sci. 2016, 2016, 7697031. [Google Scholar] [CrossRef] [Green Version]
- Olukman, M.; Oya, S.; Solak, E.K. Release of Anticancer Drug 5-Fluorouracil from Different Ionically Crosslinked Alginate Beads. J. Biomater. Nanobiotechnol. 2012, 3, 469. [Google Scholar] [CrossRef] [Green Version]
- Rahaiee, S.; Hashemi, M.; Shojaosadati, S.A.; Moini, S.; Razavi, S.H. Nanoparticles Based on Crocin Loaded Chitosan-Alginate Biopolymers: Antioxidant Activities, Bioavailability and Anticancer Properties. Int. J. Biol. Macromol. 2017, 99, 401–408. [Google Scholar] [CrossRef]
- Rezk, A.I.; Obiweluozor, F.O.; Choukrani, G.; Park, C.H.; Kim, C.S. Drug Release and Kinetic Models of Anticancer Drug (BTZ) from a PH-Responsive Alginate Polydopamine Hydrogel: Towards Cancer Chemotherapy. Int. J. Biol. Macromol. 2019, 141, 388–400. [Google Scholar] [CrossRef]
- Sánchez, B.G.; Bort, A.; Mateos-Gómez, P.A.; Rodríguez-Henche, N.; Díaz-Laviada, I. Combination of the Natural Product Capsaicin and Docetaxel Synergistically Kills Human Prostate Cancer Cells through the Metabolic Regulator AMP-Activated Kinase. Cancer Cell Int. 2019, 19, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, M.K.; Baguley, B.C.; Wall, C.; Jameson, M.B.; Findlay, M.P. Review of High-Dose Intravenous Vitamin C as an Anticancer Agent. Asia Pac. J. Clin. Oncol. 2014, 10, 22–37. [Google Scholar] [CrossRef] [PubMed]
- De Francesco, E.M.; Bonuccelli, G.; Maggiolini, M.; Sotgia, F.; Lisanti, M.P. Vitamin C and Doxycycline: A Synthetic Lethal Combination Therapy Targeting Metabolic Flexibility in Cancer Stem Cells (CSCs). Oncotarget 2017, 8, 67269–67286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mostafavi-Pour, Z.; Ramezani, F.; Keshavarzi, F.; Samadi, N. The Role of Quercetin and Vitamin C in Nrf2-dependent Oxidative Stress Production in Breast Cancer Cells. Oncol. Lett. 2017, 13, 1965–1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Men’shova, R.V.; Ermakova, S.P.; Rachidi, S.M.; Al-Hajje, A.H.; Zvyagintseva, T.N.; Kanaan, H.M. Seasonal Variations of the Composition, Structural Features, and Antitumor Properties of Polysaccharides from Padina Pavonica (Lebanon) as a Function of Composition. Chem. Nat. Compd. 2012, 47, 870–875. [Google Scholar] [CrossRef]
- Tannoury, M.; Khayrallah, M.; Saab, A.M.; Diab-Assaf, M.; Rizk, F.; Hamade, A.; Najjar, F. Evaluation of Cytotoxic Activity of Chorda Filum from the Lebanese Coast against MCF-7 Cancer Cell Line. Eur. J. Biomed. 2018, 5, 56–62. [Google Scholar]
- Tannoury, M.; Saab, A.M.; Elia, J.M.; Makhlouf, H.; Diab-Assaf, M. In Vitro Cytotoxic Activity of Laurencia Papillosa, Marine Red Algae from the Lebanese Coast. J. Appl. Pharm. Sci. 2017, 7, 175–179. [Google Scholar]
- Tannoury, M.; Elia, J.M.; Saab, A.M.; Makhlouf, H.; Daouchabo, R.; Diab-Assaf, M. Evaluation of Cytotoxic Activity of Sargassum Vulgare from the Lebanese Coast Against Jurkat Cancer Cell Line. J. Appl. Pharm. Sci. 2016, 6, 108–112. [Google Scholar] [CrossRef] [Green Version]
- Al Monla, R.; Dessouki, Z.; Gali-Muhtasib, H.; Mawalwi, H. Chemical Analysis and Biological Potentials of Extracts from Colpomenia Sinuosa. Pharmacogn. Res. 2020, 12, 272–277. [Google Scholar]
- Hernández-Garibay, E.; Zertuche-González, J.A.; Pacheco-Ruiz, I. Sulfated Polysaccharides (Fucoidan) from the Brown Seaweed Silvetia Compressa (J. Agardh) E. Serrão, TO Cho, SM Boo & Brawley. J. Appl. Phycol. 2019, 31, 3841–3847. [Google Scholar] [CrossRef]
- Grasdalen, H. High-Field, 1HNMR Spectroscopy of Alginate: Sequential Structure and Linkage Conformations. Carbohydr. Res. 1983, 118, 255–260. [Google Scholar] [CrossRef]
- Davis, T.A.; Llanes, F.; Volesky, B.; Mucci, A. Metal Selectivity of Sargassum Spp. and Their Alginates in Relation to Their α-l-Guluronic Acid Content and Conformation. Environ. Sci. Technol. 2003, 37, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.A.; Llanes, F.; Volesky, B.; Diaz-Pulido, G.; McCook, L.; Mucci, A. 1H-NMR Study of Na Alginates Extracted from Sargassum Spp. in Relation to Metal Biosorption. Appl. Biochem. Biotechnol. 2003, 110, 75–90. [Google Scholar] [CrossRef]
- Diniz do Nascimento, L.; Moraes, A.A.B.; Costa, K.S.D.; Pereira Galúcio, J.M.; Taube, P.S.; Costa, C.M.L.; Neves Cruz, J.; de Aguiar Andrade, E.H.; Faria, L.J.G.D. Bioactive Natural Compounds and Antioxidant Activity of Essential Oils from Spice Plants: New Findings and Potential Applications. Biomolecules 2020, 10, 988. [Google Scholar] [CrossRef]
- Kedare, S.B.; Singh, R.P. Genesis and Development of DPPH Method of Antioxidant Assay. J. Food Sci. Technol. 2011, 48, 412–422. [Google Scholar] [CrossRef] [Green Version]
- Yamanaka, Y.; Yoshida-Yamamoto, S.; Doi, H. Microtubule Formation and Activities of Antioxidative Enzymes in PC12 Cells Exposed to Phosphatidylcholine Hydroperoxides. Int. J. Mol. Sci. 2012, 13, 15510–15522. [Google Scholar] [CrossRef] [Green Version]
- Perillo, B.; Donato, M.D.; Pezone, A.; Zazzo, E.D.; Giovannelli, P.; Galasso, G.; Castoria, G.; Migliaccio, A. ROS in Cancer Therapy: The Bright Side of the Moon. Exp. Mol. Med. 2020, 52, 192–203. [Google Scholar] [CrossRef] [PubMed]
- Wen, S.; Wang, Z.-H.; Zhang, C.-X.; Yang, Y.; Fan, Q.-L. Caspase-3 Promotes Diabetic Kidney Disease Through Gasdermin E-Mediated Progression to Secondary Necrosis During Apoptosis. Diabetes Metab. Syndr. Obes. Targets Ther. 2020, 13, 313–323. [Google Scholar] [CrossRef] [Green Version]
- Sanjeewa, K.K.A.; Kang, N.; Ahn, G.; Jee, Y.; Kim, Y.-T.; Jeon, Y.-J. Bioactive Potentials of Sulfated Polysaccharides Isolated from Brown Seaweed Sargassum Spp in Related to Human Health Applications: A Review. Food Hydrocoll. 2018, 81, 200–208. [Google Scholar] [CrossRef]
- Al Monla, R.; Dassouki, Z.; Kouzayha, A.; Salma, Y.; Gali-Muhtasib, H.; Mawlawi, H. The Cytotoxic and Apoptotic Effects of the Brown Algae Colpomenia Sinuosa Are Mediated by the Generation of Reactive Oxygen Species. Molecules 2020, 25, 1993. [Google Scholar] [CrossRef]
- Arumugam, P.; Arunkumar, K.; Sivakumar, L.; Murugan, M.; Murugan, K. Anticancer Effect of Fucoidan on Cell Proliferation, Cell Cycle Progression, Genetic Damage and Apoptotic Cell Death in HepG2 Cancer Cells. Toxicol. Rep. 2019, 6, 556–563. [Google Scholar] [CrossRef]
- Mathlouthi, M.; Koenig, J.L. Vibrational Spectra of Carbohydrates. Adv. Carbohydr. Chem. Biochem. 1986, 44, 7–89. [Google Scholar] [CrossRef]
- Leal, D.; Matsuhiro, B.; Rossi, M.; Caruso, F. FT-IR Spectra of Alginic Acid Block Fractions in Three Species of Brown Seaweeds. Carbohydr. Res. 2008, 343, 308–316. [Google Scholar] [CrossRef]
- Sinha, S.; Astani, A.; Ghosh, T.; Schnitzler, P.; Ray, B. Polysaccharides from Sargassum Tenerrimum: Structural Features, Chemical Modification and Anti-Viral Activity. Phytochemistry 2010, 71, 235–242. [Google Scholar] [CrossRef]
- Zhang, Z.; Lin, T.; Li, S.; Chen, X.; Que, X.; Sheng, L.; Hu, Y.; Peng, J.; Ma, H.; Li, J.; et al. Polyacrylamide/Copper-Alginate Double Network Hydrogel Electrolyte with Excellent Mechanical Properties and Strain-Sensitivity. Macromol. Biosci. 2021, 578, 2100361. [Google Scholar] [CrossRef]
- Hilbig, J.; Hartlieb, K.; Gibis, M.; Herrmann, K.; Weiss, J. Rheological and Mechanical Properties of Alginate Gels and Films Containing Different Chelators. Food Hydrocoll. 2020, 101, 105487. [Google Scholar] [CrossRef]
- Sari-Chmayssem, N.; Taha, S.; Mawlawi, H.; Guégan, J.-P.; Jeftić, J.; Benvegnu, T. Extracted and Depolymerized Alginates from Brown Algae Sargassum Vulgare of Lebanese Origin: Chemical, Rheological, and Antioxidant Properties. J. Appl. Phycol. 2016, 28, 1915–1929. [Google Scholar] [CrossRef]
- Haddad, M.; Zein, S.; Hazeime, G.; Krivoruchko, E.; Makhour, Y.; Kassem, Z.; Kanaan, H. Structural Characteristics, Antitumor and Antioxidant Properties of Polysaccharides Isolated from the Brown Algae Stypopodium schimperi Growing on the Lebanese Coast. ARJMD 2017, 17, 36–43. [Google Scholar]
- Magalhaes, K.D.; Costa, L.S.; Fidelis, G.P.; Oliveira, R.M.; Nobre, L.T.D.B.; Dantas-Santos, N.; Camara, R.B.G.; Albuquerque, I.R.L.; Cordeiro, S.L.; Sabry, D.A.; et al. Anticoagulant, Antioxidant and Antitumor Activities of Heterofucans from the Seaweed Dictyopteris delicatula. Int. J. Mol. Sci. 2011, 12, 3352–3365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Usov, A.; Bilan, M.I. Fucoidans-Sulfated Polysaccharides of Brown Algae. Russ. Chem. Rev. 2009, 78, 785–799. [Google Scholar] [CrossRef]
- Yu, P.; Sun, H. Purification of a Fucoidan from Kelp Polysaccharide and Its Inhibitory Kinetics for Tyrosinase. Carbohydr. Polym. 2014, 99, 278–283. [Google Scholar] [CrossRef] [PubMed]
- Thangapandi, M.; Ajithkumar, T.; Shanmugaasokan, L.; Suja, G.; Thirumalairaj, V. In Vitro Anticancer Activity of Fucoidan from Turbinaria Conoides against A549 Cell Lines. Int. J. Biol. Macromol. 2014, 72, 919–923. [Google Scholar] [CrossRef]
- Sinurat, E.; Saefudin, E.; Peranginangin, R.; Pws, S.H. Immunostimulatory Activity of Brown Seaweed-Derived Fucoidans at Different Molecular Weights and Purity Levels towards White Spot Syndrome Virus (WSSV) in Shrimp Litopenaeus vannamei. J. Appl. Pharm. Sci. 2016, 6, 82–91. [Google Scholar] [CrossRef] [Green Version]
- Alwarsamy, M.; Gooneratne, R.; Ravichandran, R. Effect of Fucoidan from Turbinaria Conoides on Human Lung Adenocarcinoma Epithelial (A549) Cells. Carbohydr. Polym. 2016, 152, 207–213. [Google Scholar] [CrossRef]
- Ptak, S.H.; Fretté, X.C. Algae Polysaccharides as Nutraceuticals: Isolation, Characterization and Bioactivities of Fucoidans. In Algae for Food; CRC Press: Boca Raton, FL, USA, 2021; ISBN 978-1-00-316594-1. [Google Scholar]
- Szabó, L.; Gerber-Lemaire, S.; Wandrey, C. Strategies to Functionalize the Anionic Biopolymer Na-Alginate without Restricting Its Polyelectrolyte Properties. Polymers 2020, 12, 919. [Google Scholar] [CrossRef] [Green Version]
- Shao, L.; Sun, Y.; Liang, J.; Li, M.; Li, X. Decolorization Affects the Structural Characteristics and Antioxidant Activity of Polysaccharides from Thesium Chinense Turcz: Comparison of Activated Carbon and Hydrogen Peroxide Decolorization. Int. J. Biol. Macromol. 2020, 155, 1084–1091. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-Y.; Wang, S.-H.; Huang, C.-Y.; Dong, C.-D.; Huang, C.-Y.; Chang, C.-C.; Chang, J.-S. Effect of Molecular Mass and Sulfate Content of Fucoidan from Sargassum Siliquosum on Antioxidant, Anti-Lipogenesis, and Anti-Inflammatory Activity. J. Biosci. Bioeng. 2021, 132, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Lutfia, F.N.L.; Isnansetyo, A.; Susidarti, R.A.; Nursid, M. Chemical Composition Diversity of Fucoidans Isolated from Three Tropical Brown Seaweeds (Phaeophyceae) Species. Biodiversitas J. Biol. Divers. 2020, 21, 3170–3177. [Google Scholar] [CrossRef]
- Palanisamy, S.; Vinosha, M.; Marudhupandi, T.; Rajasekar, P.; Prabhu, N.M. In Vitro Antioxidant and Antibacterial Activity of Sulfated Polysaccharides Isolated from Spatoglossum Asperum. Carbohydr Polym. 2017, 170, 296–304. [Google Scholar] [CrossRef]
- Liu, J.; Wu, S.-Y.; Chen, L.; Li, Q.-J.; Shen, Y.-Z.; Jin, L.; Zhang, X.; Chen, P.-C.; Wu, M.-J.; Choi, J.; et al. Different Extraction Methods Bring About Distinct Physicochemical Properties and Antioxidant Activities of Sargassum Fusiforme Fucoidans. Int. J. Biol. Macromol. 2010, 155, 1385–1392. [Google Scholar] [CrossRef]
- Kurnialahi, I.D.; Husni, A.; Sinurat, E.; Isnansetyo, A. Antioxidant Activity of Tropical Seaweed Sargassum Muticum Fucoidan. Aquac. Aquar. Conserv. Legis. 2020, 13, 230–240. [Google Scholar]
- Wang, S.-H.; Huang, C.-Y.; Chen, C.-Y.; Chang, C.-C.; Huang, C.-Y.; Dong, C.-D.; Chang, J.-S. Structure and Biological Activity Analysis of Fucoidan Isolated from Sargassum siliquosum. ACS Omega 2020, 5, 32447–32455. [Google Scholar] [CrossRef]
- Kim, E.J.; Park, S.Y.; Lee, J.-Y.; Park, J.H.Y. Fucoidan Present in Brown Algae Induces Apoptosis of Human Colon Cancer Cells. BMC Gastroenterol. 2010, 10, 96. [Google Scholar] [CrossRef] [Green Version]
- Sokolova, R.V.; Ermakova, S.P.; Awada, S.M.; Zvyagintseva, T.N.; Kanaan, H.M. Composition, Structural Characteristics, and Antitumor Properties of Polysaccharides from the Brown Algae Dictyopteris Polypodioides and Sargassum sp. Chem. Nat. Compd. 2011, 47, 329–334. [Google Scholar] [CrossRef]
- Yang, L.; Wang, P.; Wang, H.; Li, Q.; Teng, H.; Liu, Z.; Yang, W.; Hou, L.; Zou, X. Fucoidan Derived from Undaria Pinnatifida Induces Apoptosis in Human Hepatocellular Carcinoma SMMC-7721 Cells via the ROS-Mediated Mitochondrial Pathway. Mar. Drugs 2013, 11, 1961–1976. [Google Scholar] [CrossRef] [Green Version]
- Wei, C.; Xiao, Q.; Kuang, X.; Zhang, T.; Yang, Z.; Wang, L. Fucoidan Inhibits Proliferation of the SKM-1 Acute Myeloid Leukaemia Cell Line via the Activation of Apoptotic Pathways and Production of Reactive Oxygen Species. Mol. Med. Rep. 2015, 12, 6649–6655. [Google Scholar] [CrossRef] [Green Version]
- Han, M.H.; Lee, D.-S.; Jeong, J.-W.; Hong, S.-H.; Choi, I.-W.; Cha, H.-J.; Kim, S.; Kim, H.-S.; Park, C.; Kim, G.-Y.; et al. Fucoidan Induces ROS-Dependent Apoptosis in 5637 Human Bladder Cancer Cells by Downregulating Telomerase Activity via Inactivation of the PI3K/Akt Signaling Pathway. Drug Dev. Res. 2017, 78, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Narayani, S.S.; Saravanan, S.; Ravindran, J.; Ramasamy, M.S.; Chitra, J. In Vitro Anticancer Activity of Fucoidan Extracted from Sargassum Cinereum against Caco-2 Cells. Int. J. Biol. Macromol. 2019, 138, 618–628. [Google Scholar] [CrossRef]
- Zhou, L.; Yi, Y.; Yuan, Q.; Zhang, J.; Li, Y.; Wang, P.; Xu, M.; Xie, S. VAOS, a Novel Vanadyl Complexes of Alginate Saccharides, Inducing Apoptosis via Activation of AKT-Dependent ROS Production in NSCLC. Free Radic. Biol. Med. 2018, 129, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, O.M.; El-Deeb, N.M.; Abbas, H.; Elmasry, S.M.; El-Aassar, M.R. Alginate Based Tamoxifen/Metal Dual Core-Folate Decorated Shell: Nanocomposite Targeted Therapy for Breast Cancer via ROS-Driven NF-ΚB Pathway Modulation. Int. J. Biol. Macromol. 2020, 146, 119–131. [Google Scholar] [CrossRef]
- Laurienzo, P. Marine Polysaccharides in Pharmaceutical Applications: An Overview. Mar. Drugs 2010, 8, 2435–2465. [Google Scholar] [CrossRef] [Green Version]
- Saitoh, Y.; Nagai, Y.; Miwa, N. Fucoidan-Vitamin C Complex Suppresses Tumor Invasion through the Basement Membrane, with Scarce Injuries to Normal or Tumor Cells, via Decreases in Oxidative Stress and Matrix Metalloproteinases. Int. J. Oncol. 2009, 35, 1183–1189. [Google Scholar] [CrossRef] [Green Version]
- Park, H.Y.; Kim, G.-Y.; Moon, S.-K.; Kim, W.J.; Yoo, Y.H.; Choi, Y.H. Fucoidan Inhibits the Proliferation of Human Urinary Bladder Cancer T24 Cells by Blocking Cell Cycle Progression and Inducing Apoptosis. Molecules 2014, 19, 5981–5998. [Google Scholar] [CrossRef] [Green Version]
- Singh, T.R.; Ezhilarasan, D. Ethanolic Extract of Lagerstroemia Speciosa (L.) Pers., Induces Apoptosis and Cell Cycle Arrest in HepG2 Cells. Nutr. Cancer 2020, 72, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Atashrazm, F.; Lowenthal, R.M.; Woods, G.M.; Holloway, A.F.; Dickinson, J.L. Fucoidan and Cancer: A Multifunctional Molecule with Anti-Tumor Potential. Mar. Drugs 2015, 13, 2327–2346. [Google Scholar] [CrossRef] [Green Version]
- Park, H.Y.; Park, S.-H.; Jeong, J.-W.; Yoon, D.; Han, M.H.; Lee, D.-S.; Choi, G.; Yim, M.-J.; Lee, J.M.; Kim, D.-H.; et al. Induction of P53-Independent Apoptosis and G1 Cell Cycle Arrest by Fucoidan in HCT116 Human Colorectal Carcinoma Cells. Mar. Drugs 2017, 15, 154. [Google Scholar] [CrossRef] [Green Version]
- Dahmoune, F.; Nayak, B.; Moussi, K.; Remini, H.; Madani, K. Optimization of Microwave-Assisted Extraction of Polyphenols from Myrtus Communis L. Leaves. Food Chem. 2015, 166, 585–595. [Google Scholar] [CrossRef]
- Dodgson, K.S.; Price, R.G. A Note on the Determination of the Ester Sulphate Content of Sulphated Polysaccharides. Biochem. J. 1962, 84, 106–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saboural, P.; Chaubet, F.; Rouzet, F.; Al-Shoukr, F.; Azzouna, R.B.; Bouchemal, N.; Picton, L.; Louedec, L.; Maire, M.; Rolland, L.; et al. Purification of a Low Molecular Weight Fucoidan for SPECT Molecular Imaging of Myocardial Infarction. Mar. Drugs 2014, 12, 4851–4867. [Google Scholar] [CrossRef] [Green Version]
- Sułkowska-Ziaja, K.; Muszyńska, B.; Ekiert, H. Chemical Composition and Cytotoxic Activity of the Polysaccharide Fractions in Sarcodon Imbricatus (Basidiomycota). Acta Mycol. 2013, 47, 49–56. [Google Scholar] [CrossRef]
- Gómez-Ordóñez, E.; Rupérez, P. FTIR-ATR Spectroscopy as a Tool for Polysaccharide Identification in Edible Brown and Red Seaweeds. Food Hydrocoll. 2011, 25, 1514–1520. [Google Scholar] [CrossRef]
- Guo, X.; Ye, X.; Sun, Y.; Wu, D.; Wu, N.; Hu, Y.; Chen, S. Ultrasound Effects on the Degradation Kinetics, Structure, and Antioxidant Activity of Sea Cucumber Fucoidan. J. Agric. Food Chem. 2014, 62, 1088–1095. [Google Scholar] [CrossRef]
- Wang, L.; Xiao, R.; Mo, J. Quantitative Detection Method of Semiquinone Free Radicals on Particulate Matters Using Electron Spin Resonance Spectroscopy. Sustain. Cities Soc. 2019, 49, 101614. [Google Scholar] [CrossRef]
- Yen, G.-C.; Chen, H.-Y. Antioxidant Activity of Various Tea Extracts in Relation to Their Antimutagenicity. J. Agric. Food Chem. 1995, 43, 27–32. [Google Scholar] [CrossRef]
- Elansary, H.O.; Yessoufou, K.; Abdel-Hamid, A.M.E.; El-Esawi, M.A.; Ali, H.M.; Elshikh, M.S. Seaweed Extracts Enhance Salam Turfgrass Performance during Prolonged Irrigation Intervals and Saline Shock. Front. Plant Sci. 2017, 8, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luis, C.; Castaño-Guerrero, Y.; Soares, R.; Sales, G.; Fernandes, R. Avoiding the Interference of Doxorubicin with MTT Measurements on the MCF-7 Breast Cancer Cell Line. Methods Protoc. 2019, 2, 29. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Yield (%) | Protein (%) | Sulfate (%) | Phenol (mg GAE/gDW) | D-Glucuronic Acid (%) |
|---|---|---|---|---|---|
| Fucoidan | 11.6 ± 0.3 | 1.88 ± 1.1 | 18.8 ± 2.1 | 0.045 ± 0.01 | 3.8 ± 0.5 |
| Alginate | 13.6 ± 0.4 | 2.63 ± 1.4 | 5.53 ± 2.6 | 0.043 ± 0.01 | 50 ± 0.7 |
| Sample | Monosaccharide Composition (%) | |||||
|---|---|---|---|---|---|---|
| Glucose | Xylose | mannose | Arabinose | Galactose | fucose | |
| Fucoidan | 5.45 ± 2.97 | 2.62 ± 2.12 | 3.58 ± 1.34 | 3.4 ± 1.39 | 5.94 ± 3.88 | 67.4 ± 12.1 |
| Alginate | 2.67 ± 3.59 | 2.14 ± 2.48 | 5.11 ± 2.64 | 4.2 ± 3.67 | 4.98 ± 4.66 | 0.2 ± 0.34 |
| Numeric Values | FG | FM | M/G Ratio | FGG | FGM = FMG * | FMM |
|---|---|---|---|---|---|---|
| Alginate | 0.71 | 0.29 | 0.41 | 0.65 | 0.06 | 0.23 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al Monla, R.; Dassouki, Z.; Sari-Chmayssem, N.; Mawlawi, H.; Gali-Muhtasib, H. Fucoidan and Alginate from the Brown Algae Colpomenia sinuosa and Their Combination with Vitamin C Trigger Apoptosis in Colon Cancer. Molecules 2022, 27, 358. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27020358
Al Monla R, Dassouki Z, Sari-Chmayssem N, Mawlawi H, Gali-Muhtasib H. Fucoidan and Alginate from the Brown Algae Colpomenia sinuosa and Their Combination with Vitamin C Trigger Apoptosis in Colon Cancer. Molecules. 2022; 27(2):358. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27020358
Chicago/Turabian StyleAl Monla, Reem, Zeina Dassouki, Nouha Sari-Chmayssem, Hiba Mawlawi, and Hala Gali-Muhtasib. 2022. "Fucoidan and Alginate from the Brown Algae Colpomenia sinuosa and Their Combination with Vitamin C Trigger Apoptosis in Colon Cancer" Molecules 27, no. 2: 358. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27020358