
Microwave-Assisted Desulfation of the Hemolytic Saponins Extracted from Holothuria scabra Viscera

 , , and
, , and
Abstract
:1. Introduction
2. Results and Discussion
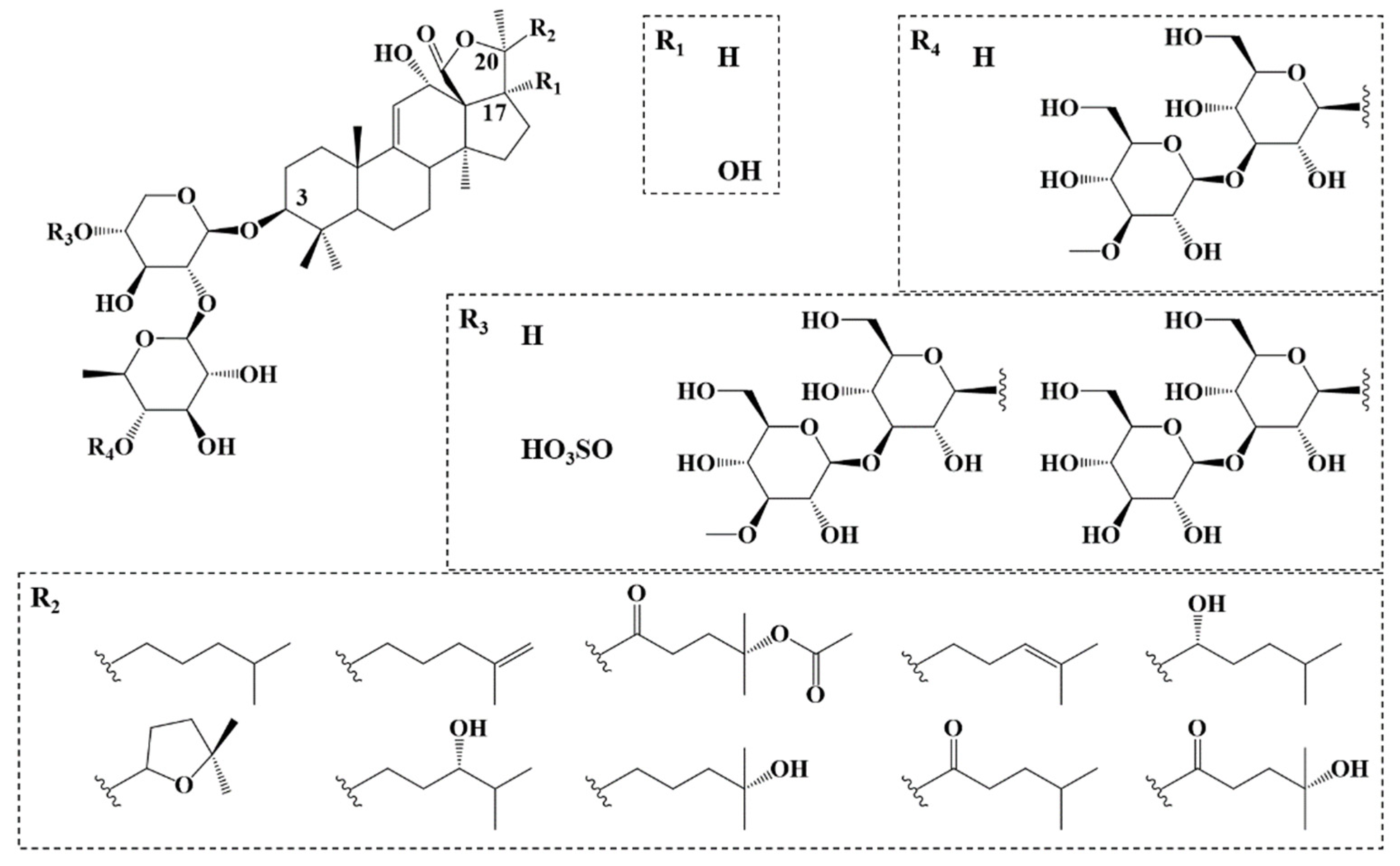
2.1. Saponin Identification and Quantification in the Natural Extract
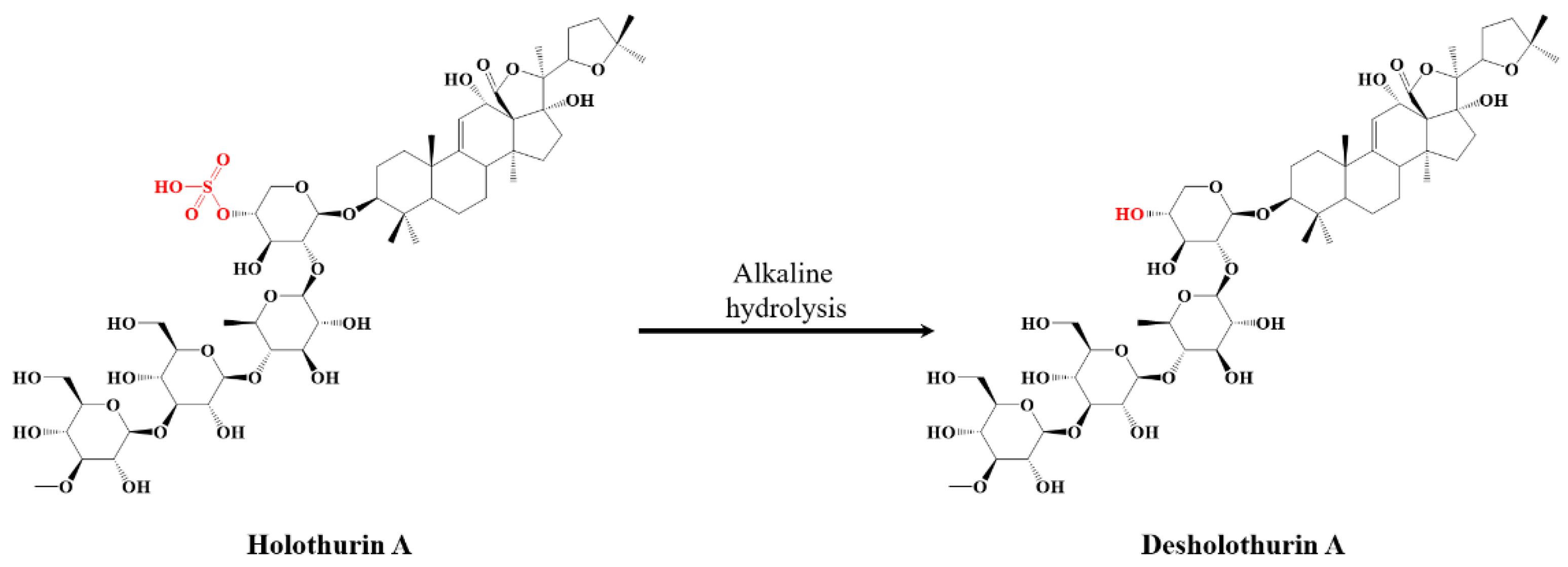
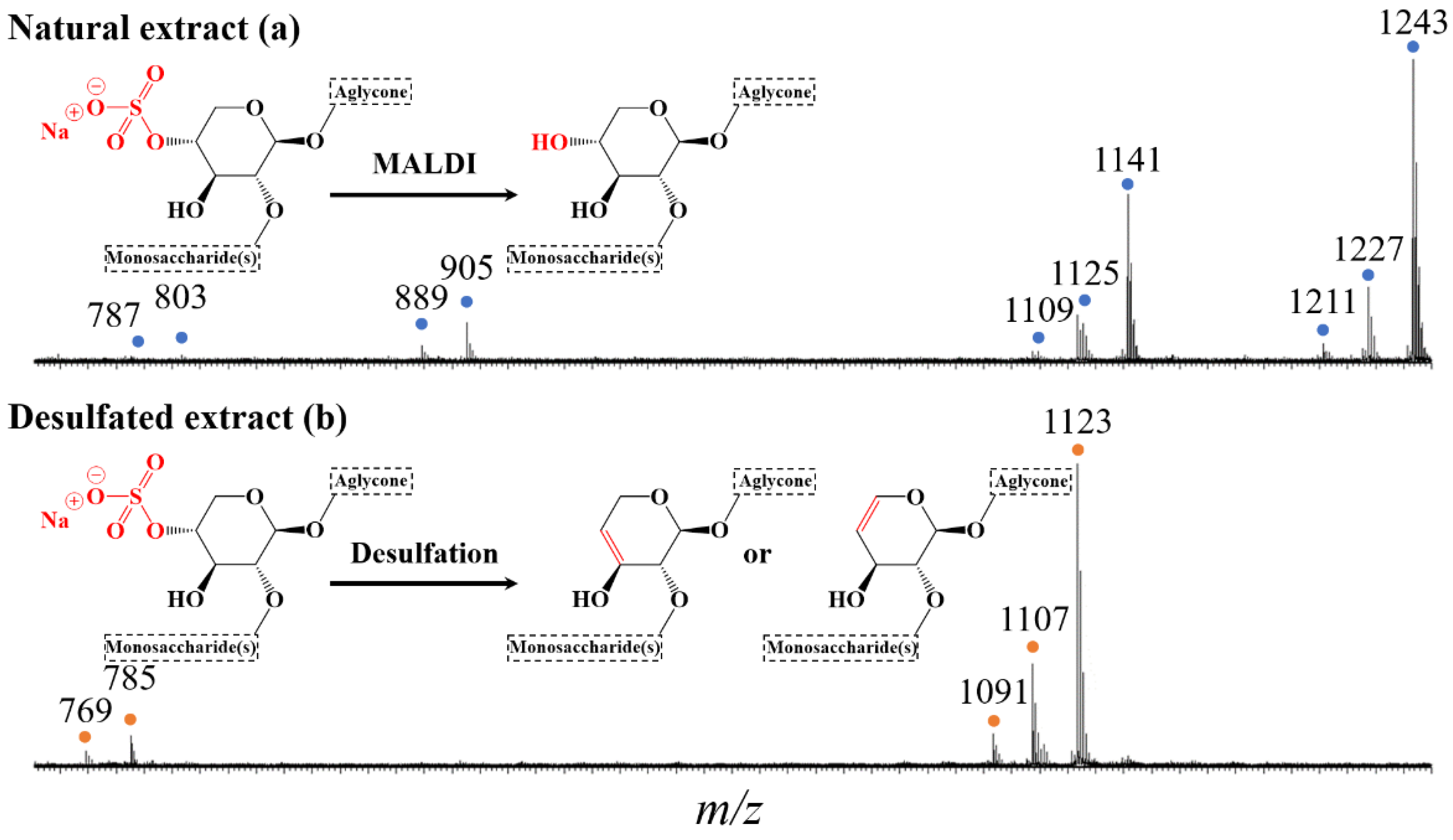
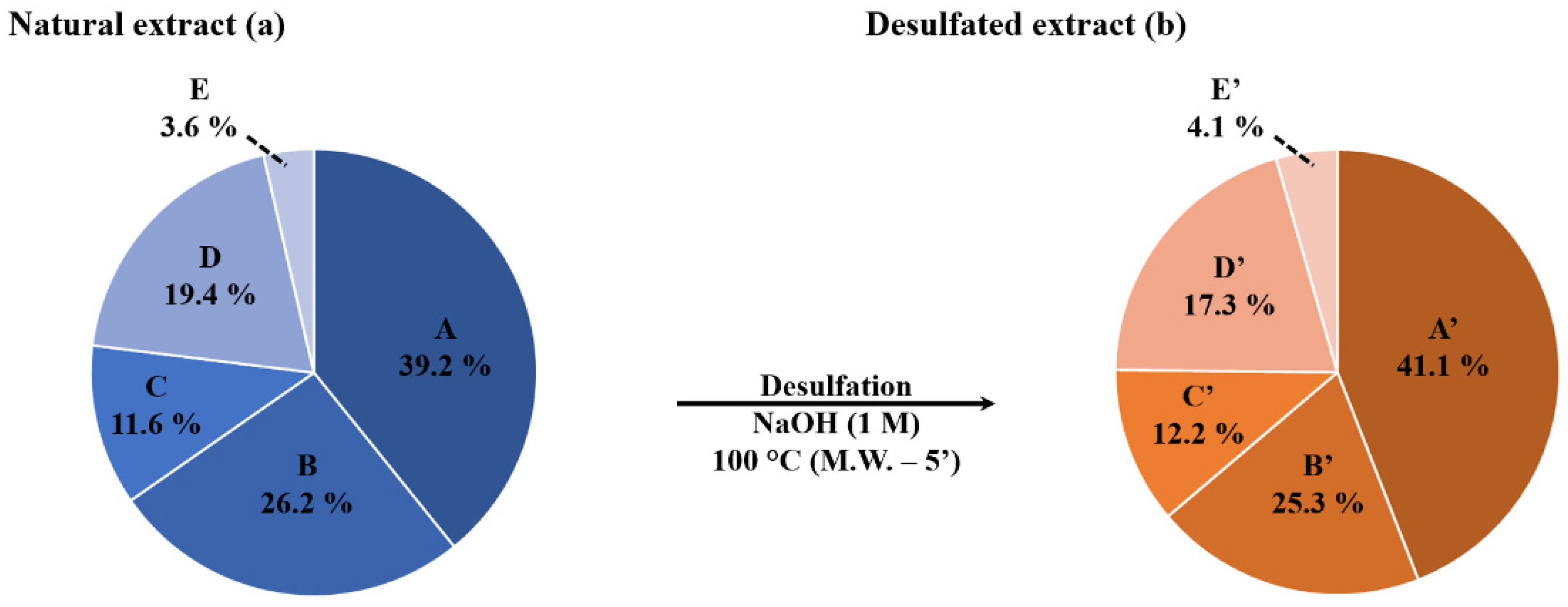
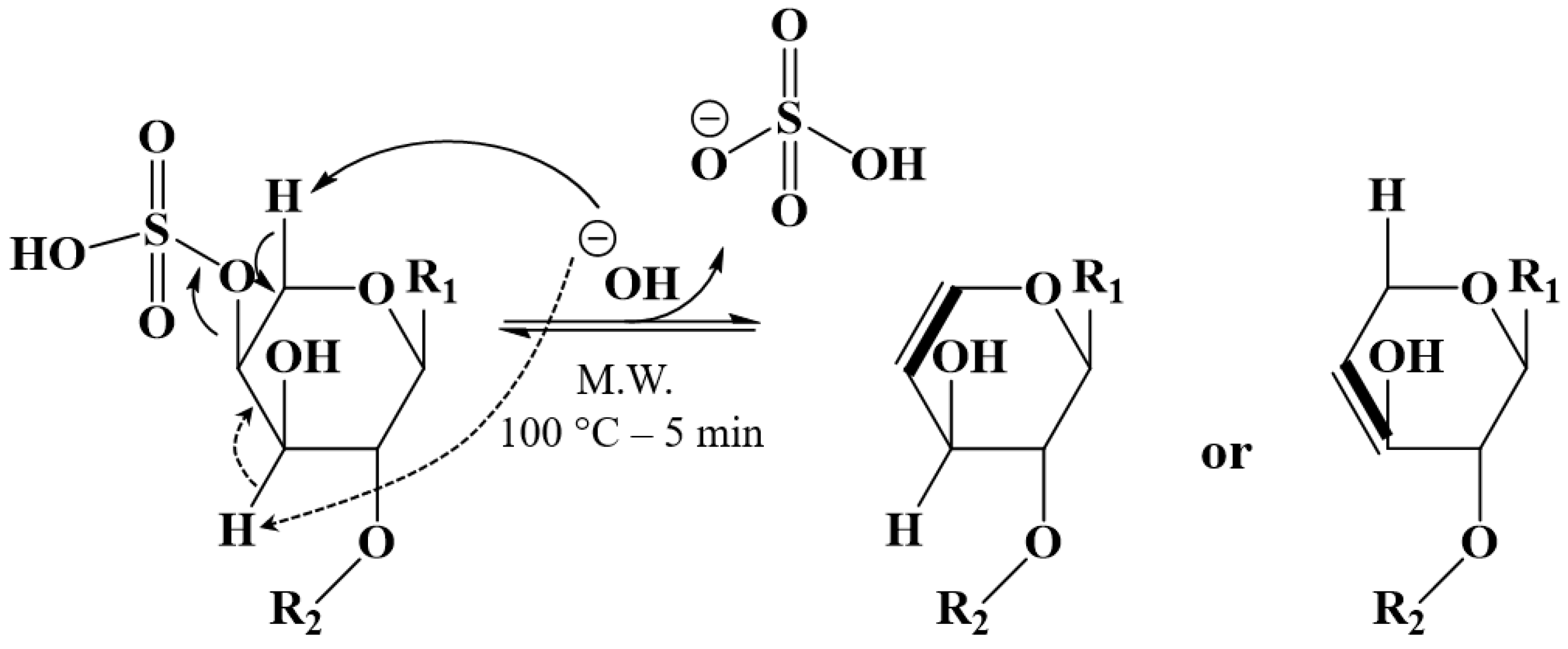
2.2. Selective Microwave-Assisted Desulfation of Saponins
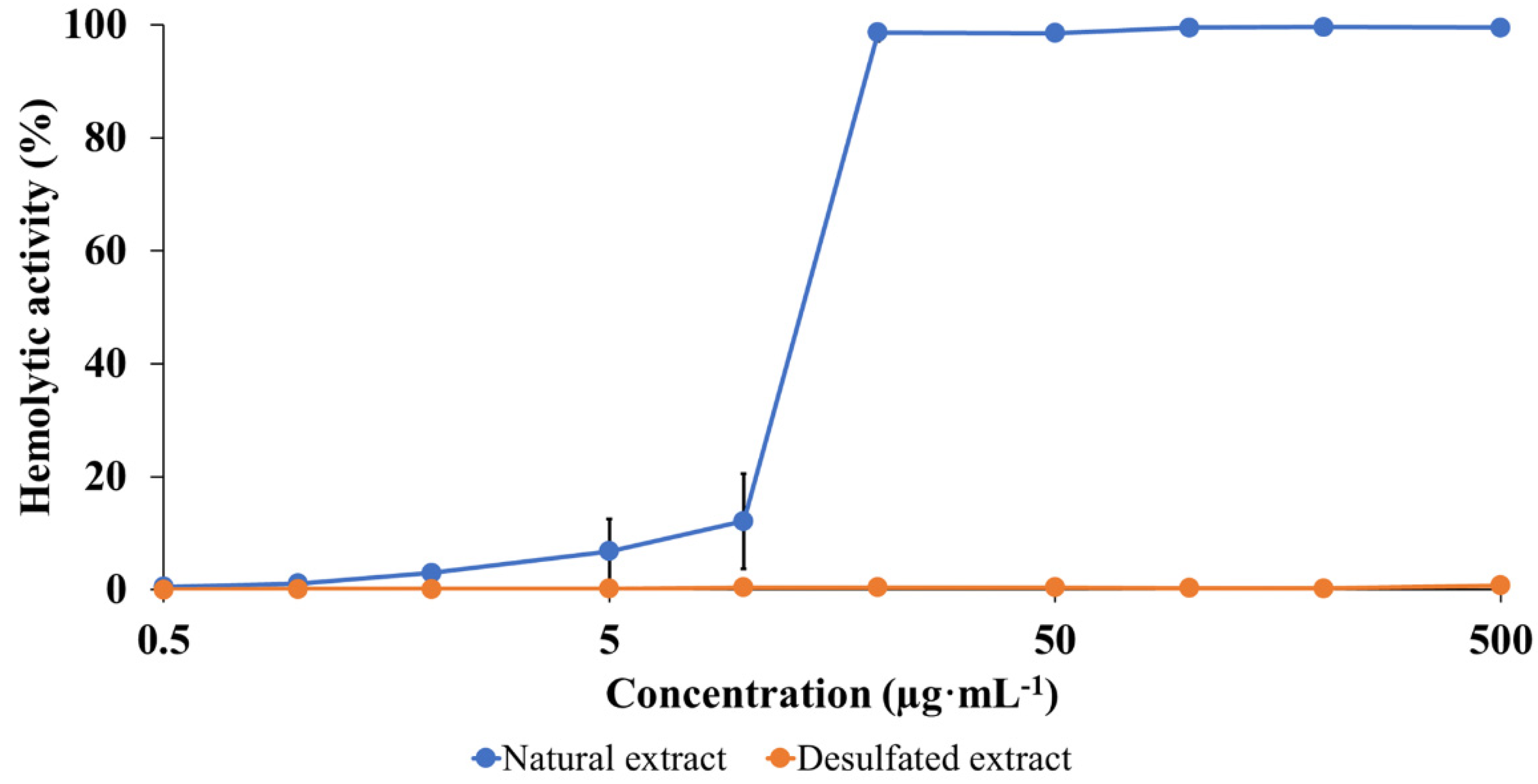
2.3. Hemolytic Activity (HA) Modulation
3. Materials and Methods
3.1. Chemicals
3.2. Extraction & Purification
3.3. Microwave-Assisted Alkaline Desulfation
3.4. Mass Spectrometry
3.5. Hemolytic Activity Assay
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Dutt, R.; Sharma, A.K.; Keservani, R.K.; Garg, V. Promising Drug Molecules of Natural Origin; CRC Press: Coca Raton, FL, USA, 2020; ISBN 9781771888868. [Google Scholar]
- Kornprobst, J.-M. Substances Naturelles D’origine Marine: Chimiodiversité, Pharmacodiversité, Biotechnologies; Lavoisier: Cachan, France, 2005; ISBN 2-7430-0721-4. [Google Scholar]
- Seca, A.M.L.; Pinto, D.C.G.A. Biological Potential and Medical Use of Secondary Metabolites. Medicines 2019, 6, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kabera, J.N.; Semana, E.; Mussa, A.R.; He, X. Plant Secondary Metabolites: Biosynthesis, Classification, Function and Pharmacological Properties. J. Pharm. Pharmacol. 2014, 2, 377–392. [Google Scholar]
- Caulier, G.; Flammang, P.; Gerbaux, P.; Eeckhaut, I. When a repellent becomes an attractant: Harmful saponins are kairomones attracting the symbiotic Harlequin crab. Sci. Rep. 2013, 3, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyazaki, S.; Ichiba, T.; Reimer, J.D.; Tanaka, J. Chemoattraction of the pearlfish Encheliophis vermicularis to the sea cucumber Holothuria leucospilota. Chemoecology 2014, 24, 121–126. [Google Scholar] [CrossRef]
- Moghimipour, E.; Handali, S. Saponin: Properties, methods of evaluation and applications. Annu. Res. Rev. Biol. 2015, 5, 207–220. [Google Scholar] [CrossRef] [Green Version]
- da Silveira Agostini-Costa, T.; Vieira, R.; Bizzo, H.; Silveira, D. Secondary Metabolites. In Chromatography and Its Applications; IntechOpen: London, UK, 2012; pp. 131–164. ISBN 00319422. [Google Scholar]
- Kassem, A.S.; Ahmed, A.M.; Tariq, M.R. Study of saponins in methanol extract of the leaves of Acacia etbaica subspecies etbaica. Res. J. Pharm. Biol. Chem. Sci. 2014, 5, 803–810. [Google Scholar]
- Demeyer, M.; De Winter, J.; Caulier, G.; Eeckhaut, I.; Flammang, P.; Gerbaux, P. Molecular diversity and body distribution of saponins in the sea star Asterias rubens by mass spectrometry. Comp. Biochem. Physiol. Part. B Biochem. Mol. Biol. 2014, 168, 1–11. [Google Scholar] [CrossRef]
- Bahrami, Y.; Zhang, W.; M. M. Franco, C. Distribution of Saponins in the Sea Cucumber Holothuria lessoni; the Body Wall Versus the Viscera, and Their Biological Activities. Mar. Drugs 2018, 16, 423. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Laval, S.; Yu, B. Chemical Synthesis of Saponins, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2014; Volume 71, ISBN 9780128001288. [Google Scholar]
- De Simone, F.; Dini, A.; Finamore, E.; Minale, L.; Pizza, C.; Riccio, R.; Zollo, F. Starfish saponins. Part 5. Structure of sepositoside A, a novel steroidal cyclic glycoside from the starfish Echinaster sepositus. J. Chem. Soc. Perkin Trans. 1981, 1, 1855–1862. [Google Scholar] [CrossRef]
- Tantry, M.A.; Khan, I.A. Saponins from Glycine max Merrill (soybean). Fitoterapia 2013, 87, 49–56. [Google Scholar] [CrossRef]
- Yoshikawa, M.; Harada, E.; Murakami, T.; Matsuda, H.; Wariishi, N.; Yamahara, J.; Murakami, N.; Kitagawa, I. Escins-Ia, Ib, IIa, IIb, and IIIa, Bioactive Triterpene Oligoglycosides From the Seeds of Aesculus Hippocastanum, L.: Their Inhibitory Effects on Ethanol Absorption and Hypoglycemic Activity on Glucose Tolerance Test. Chem. Pharm. Bull. 1994, 42, 1357–1359. [Google Scholar] [CrossRef] [Green Version]
- Jæger, D.; Ndi, C.P.; Crocoll, C.; Simpson, B.S.; Khakimov, B.; Guzman-Genuino, R.M.; Hayball, J.D.; Xing, X.; Bulone, V.; Weinstein, P.; et al. Isolation and Structural Characterization of Echinocystic Acid Triterpenoid Saponins from the Australian Medicinal and Food Plant Acacia ligulata. J. Nat. Prod. 2017, 80, 2692–2698. [Google Scholar] [CrossRef]
- Honey-Escandón, M.; Arreguín-Espinosa, R.; Solís-Marín, F.A.; Samyn, Y. Biological and taxonomic perspective of triterpenoid glycosides of sea cucumbers of the family Holothuriidae (Echinodermata, Holothuroidea). Comp. Biochem. Physiol. Part.B Biochem. Mol. Biol. 2015, 180, 16–39. [Google Scholar] [CrossRef]
- Caulier, G.; Mezali, K.; Soualili, D.L.; Decroo, C.; Demeyer, M.; Eeckhaut, I.; Gerbaux, P.; Flammang, P. Chemical characterization of saponins contained in the body wall and the Cuvierian tubules of the sea cucumber Holothuria (Platyperona) sanctori (Delle Chiaje, 1823). Biochem. Syst. Ecol. 2016, 68, 119–127. [Google Scholar] [CrossRef]
- Mert-Türk, F. Saponins versus plant fungal pathogens. J. Cell Mol. Biol. 2006, 5, 13–17. [Google Scholar]
- Mohammadizadeh, F.; Ehsanpor, M.; Afkhami, M.; Mokhlesi, A.; Khazaali, A.; Montazeri, S. Evaluation of antibacterial, antifungal and cytotoxic effects of Holothuria scabra from the North Coast of the Persian Gulf. J. Mycol. Med. 2013, 23, 225–229. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-J.; Sung, J.-H.; Lee, S.-J.; Moon, C.-K.; Lee, B.-H. Antitumor activity of a novel ginseng saponin metabolite in human pulmonary adenocarcinoma cells resistant to cisplatin. Cancer Lett. 1999, 144, 39–43. [Google Scholar] [CrossRef]
- Kalinin, V.I.; Prokofieva, N.G.; Likhatskaya, G.N.; Schentsova, E.B.; Agafonova, I.G.; Avilov, S.A.; Drozdova, O.A. Hemolytic activities of triterpene glycosides from the holothurian order dendrochirotida: Some trends in the evolution of this group of toxins. Toxicon 1996, 34, 475–483. [Google Scholar] [CrossRef]
- Lorent, J.H.; Quetin-Leclercq, J.; Mingeot-Leclercq, M.P. The amphiphilic nature of saponins and their effects on artificial and biological membranes and potential consequences for red blood and cancer cells. Org. Biomol. Chem. 2014, 12, 8803–8822. [Google Scholar] [CrossRef] [PubMed]
- Keukens, E.A.J.; De Vrije, T.; Van Den Boom, C.; De Waard, P.; Plasman, H.H.; Thiel, F.; Chupin, V.; Jongen, W.M.F.; De Kruijff, B. Molecular basis of glycoalkaloid induced membrane disruption. Biochim. Biophys. Acta 1995, 1240, 216–228. [Google Scholar] [CrossRef] [Green Version]
- Lorent, J.; Lins, L.; Domenech, Ò.; Quetin-Leclercq, J.; Brasseur, R.; Mingeot-Leclercq, M.-P. Domain Formation and Permeabilization Induced by the Saponin α-Hederin and Its Aglycone Hederagenin in a Cholesterol-Containing Bilayer. Langmuir 2014, 30, 4556–4569. [Google Scholar] [CrossRef] [PubMed]
- Lorent, J.; Le Duff, C.S.; Quetin-Leclercq, J.; Mingeot-Leclercq, M.-P. Induction of Highly Curved Structures in Relation to Membrane Permeabilization and Budding by the Triterpenoid Saponins, α- and δ-Hederin. J. Biol. Chem. 2013, 288, 14000–14017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Böttger, S.; Melzig, M.F. The influence of saponins on cell membrane cholesterol. Bioorg. Med. Chem. 2013, 21, 7118–7124. [Google Scholar] [CrossRef] [PubMed]
- Qu, C.; Yang, L.; Yu, S.; Wang, S.; Bai, Y.; Zhang, H. Investigation of the interactions between ginsenosides and amino acids by mass spectrometry and theoretical chemistry. Spectrochim. Acta Part. A Mol. Biomol. Spectrosc. 2009, 74, 478–483. [Google Scholar] [CrossRef]
- Zelepuga, E.A.; Silchenko, A.S.; Avilov, S.A.; Kalinin, V.I. Structure-activity relationships of holothuroid’s triterpene glycosides and some in silico insights obtained by molecular dynamics study on the mechanisms of their membranolytic action. Mar. Drugs 2021, 19, 604. [Google Scholar] [CrossRef]
- Colson, E.; Savarino, P.; J.S. Claereboudt, E.; Cabrera-Barjas, G.; Deleu, M.; Lins, L.; Eeckhaut, I.; Flammang, P.; Gerbaux, P. Enhancing the Membranolytic Activity of Chenopodium quinoa Saponins by Fast Microwave Hydrolysis. Molecules 2020, 25, 1731. [Google Scholar] [CrossRef]
- Feng, J.; Chen, Y.; Liu, X.; Liu, S. Efficient improvement of surface activity of tea saponin through Gemini-like modification by straightforward esterification. Food Chem. 2015, 171, 272–279. [Google Scholar] [CrossRef]
- Monti, D.; Candido, A.; Cruz Silva, M.M.; Křen, V.; Riva, S.; Danieli, B. Biocatalyzed generation of molecular diversity: Selective modification of the saponin asiaticoside. Adv. Synth. Catal. 2005, 347, 1168–1174. [Google Scholar] [CrossRef] [Green Version]
- Ramos-Morales, E.; de la Fuente, G.; Duval, S.; Wehrli, C.; Bouillon, M.; Lahmann, M.; Preskett, D.; Braganca, R.; Newbold, C.J. Antiprotozoal effect of saponins in the rumen can be enhanced by chemical modifications in their structure. Front. Microbiol. 2017, 8, 399. [Google Scholar] [CrossRef]
- Hamel, J.-F.; Mercier, A.; Conand, C.; Purcell, S.; Toral-Granda, T.-G.; Gamboa, R. Holothuria scabra, golden sandfish. IUCN Red List Threat. Species 2013 2013, e.T180257A1606648. [Google Scholar] [CrossRef]
- Bordbar, S.; Anwar, F.; Saari, N. High-Value Components and Bioactives from Sea Cucumbers for Functional Foods—A Review. Mar. Drugs 2011, 9, 1761–1805. [Google Scholar] [CrossRef] [Green Version]
- Lavitra, T.; Rasolofonirina, R.; Jangoux, M.; Eeckhaut, I. Problems related to the farming of Holothuria scabra (Jaeger, 1833). SPC Beche-de-mer Inf. Bull. 2009, 29, 20–30. [Google Scholar]
- Caulier, G.; Flammang, P.; Rakotorisoa, P.; Gerbaux, P.; Demeyer, M.; Eeckhaut, I. Preservation of the bioactive saponins of Holothuria scabra through the processing of trepang. Cah. Biol. Mar. 2013, 54, 685–690. [Google Scholar]
- Van Thanh, N.; Dang, N.H.; Van Kiem, P.; Cuong, N.X.; Huong, H.T.; Van Minh, C. A New Triterpene Glycoside from the Sea Cucumber Holothuria Scabra Collected in Vietnam. ASEAN J. Sci. Technol. Dev. 2017, 23, 253. [Google Scholar] [CrossRef] [Green Version]
- Dang, N.H.; Van Thanh, N.; Van Kiem, P.; Huong, L.M.; Van Minh, C. Two new triterpene glycosides from the vietnamese sea cucumber Holothuria scabra. Arch. Pharm. Res. 2007, 30, 1387–1391. [Google Scholar] [CrossRef]
- Han, H.; Yi, Y.; Xu, Q.; La, M.; Zhang, H. Two New Cytotoxic Triterpene Glycosides from the Sea Cucumber Holothuria scabra. Planta Med. 2009, 75, 1608–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puspitasari, Y.E.; De Bruyne, T.; Foubert, K.; Aulanni’am, A.; Pieters, L.; Hermans, N.; Tuenter, E. Holothuria triterpene glycosides: A comprehensive guide for their structure elucidation and critical appraisal of reported compounds. Phytochem. Rev. 2021, 0123456789. [Google Scholar] [CrossRef]
- Hamel, J.-F.; Conand, C.; Pawson, D.; Mercier, A. The sea cucumber Holothuria scabra (Holothuroidea: Echinodermata): Its biology and exploitation as Beche-de-mer. In Advances in Marine Biology; Elsevier Inc.: Amsterdam, The Netherlands, 2001; Volume 41 , pp. 129–223. ISBN 0120261413. [Google Scholar]
- Maier, M.S. Biological Activities of Sulfated Glycosides from Echinoderms. In Studies in Natural Products Chemistry; Elsevier B.V.: Amsterdam, The Netherlands, 2008; Volume 35, pp. 311–354. ISBN 9780444531810. [Google Scholar]
- Yu, S.; Ye, X.; Huang, H.; Peng, R.; Su, Z.; Lian, X.-Y.; Zhang, Z. Bioactive Sulfated Saponins from Sea Cucumber Holothuria moebii. Planta Med. 2015, 81, 152–159. [Google Scholar] [CrossRef] [Green Version]
- Bedini, E.; Laezza, A.; Parrilli, M.; Iadonisi, A. A review of chemical methods for the selective sulfation and desulfation of polysaccharides. Carbohydr. Polym. 2017, 174, 1224–1239. [Google Scholar] [CrossRef] [PubMed]
- Richel, A.; Paquot, M. Conversion of Carbohydrates Under Microwave Heating. In Carbohydrates: Comprehensive Studies on Glycobiology and Glycotechnology; IntechOpen: London, UK, 2012. [Google Scholar]
- Witkowska, H.E.; Bialy, Z.; Jurzysta, M.; Waller, G.R. Analysis of Saponin Mixtures from Alfalfa (Medicago Sativa L.) Roots using Mass Spectrometry with MALDI Techniques. Nat. Prod. Commun. 2008, 3, 1395–1410. [Google Scholar] [CrossRef] [Green Version]
- Ji, S.; Wang, Q.; Qiao, X.; Guo, H.; Yang, Y.; Bo, T.; Xiang, C.; Guo, D.; Ye, M. New triterpene saponins from the roots of Glycyrrhiza yunnanensis and their rapid screening by LC/MS/MS. J. Pharm. Biomed. Anal. 2014, 90, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Gevrenova, R.; Bardarov, V.; Bardarov, K.; Voutquenne-Nazabadioko, L.; Henry, M. Selective Profiling of Saponins from Gypsophila trichotoma Wend. by HILIC Separation and HRMS Detection. Phytochem. Anal. 2018, 29, 250–274. [Google Scholar] [CrossRef] [PubMed]
- Ackloo, S.Z.; Smith, R.W.; Terlouw, J.K.; McCarry, B.E. Characterization of ginseng saponins using electrospray mass spectrometry and collision-induced dissociation experiments of metal-attachment ions. Analyst 2000, 125, 591–597. [Google Scholar] [CrossRef] [PubMed]
- Savarino, P.; Demeyer, M.; Decroo, C.; Colson, E.; Gerbaux, P. Mass spectrometry analysis of saponins. Mass Spectrom. Rev. 2021. [Google Scholar] [CrossRef] [PubMed]
- Van Dyck, S.; Gerbaux, P.; Flammang, P. Elucidation of molecular diversity and body distribution of saponins in the sea cucumber Holothuria forskali (Echinodermata) by mass spectrometry. Comp. Biochem. Physiol. Part.B 2009, 152, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Herrera, T.; Navarro del Hierro, J.; Fornari, T.; Reglero, G.; Martin, D. Acid hydrolysis of saponin-rich extracts of quinoa, lentil, fenugreek and soybean to yield sapogenin-rich extracts and other bioactive compounds. J. Sci. Food Agric. 2019, 99, 3157–3167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dyck, S.; Flammang, P.; Meriaux, C.; Bonnel, D.; Salzet, M.; Fournier, I.; Wisztorski, M. Localization of Secondary Metabolites in Marine Invertebrates: Contribution of MALDI MSI for the Study of Saponins in Cuvierian Tubules of H. forskali. PLoS ONE 2010, 5, e13923. [Google Scholar] [CrossRef]
- Vo, N.N.Q.; Fukushima, E.O.; Muranaka, T. Structure and hemolytic activity relationships of triterpenoid saponins and sapogenins. J. Nat. Med. 2017, 71, 50–58. [Google Scholar] [CrossRef]
- Voutquenne, L.; Lavaud, C.; Massiot, G.; Men-Olivier, L. Le Structure-Activity Relationships of Haemolytic Saponins. Pharm. Biol. 2002, 40, 253–262. [Google Scholar] [CrossRef]
- Takechi, M.; Yasuo, T. Structure-activity relationships of the saponin α-hederin. Phytochemistry 1990, 29, 451–452. [Google Scholar] [CrossRef]
- Mackie, A.M.; Grant, P.T.; Lasker, R. Avoidance reactions of a mollusc Buccinum undatum to saponin-like surface-active substances in extracts of the starfish Asterias rubens and Marthasterias glacialis. Comp. Biochem. Physiol 1968, 26, 415–428. [Google Scholar] [CrossRef]
- Domanski, D.; Zegrocka-Stendel, O.; Perzanowska, A.; Dutkiewicz, M.; Kowalewska, M.; Grabowska, I.; Maciejko, D.; Fogtman, A.; Dadlez, M.; Koziak, K. Molecular Mechanism for Cellular Response to β-Escin and Its Therapeutic Implications. PLoS ONE 2016, 11, e0164365. [Google Scholar] [CrossRef] [PubMed]
- Van Dyck, S.; Gerbaux, P.; Flammang, P. Qualitative and Quantitative Saponin Contents in Five Sea Cucumbers from the Indian Ocean. Mar. Drugs 2010, 8, 173–189. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Saponin | Composition (M) | m/z (Δ ppm) [M−H+2Na]+ | %-Weight in Extract (%) | Mass Fraction in Viscera (mg·g−1) | Retention Time (min) | Composition Molar Proportion (%) | Isomer Molar Proportion (%) |
|---|---|---|---|---|---|---|---|
| A | C54H86O27S | 1243.4795 (1.3) | 37.45 ± 0.37 | 0.102 | 5.97 | 39.18 ± 1.24 | 0.57 ± 0.11 |
| 6.24 | 0.75 ± 0.11 | ||||||
| 6.48 | 0.11 ± 0.02 | ||||||
| 6.76 | 0.20 ± 0.02 | ||||||
| 7.20 | 0.48 ± 0.07 | ||||||
| 7.81 | 0.21 ± 0.04 | ||||||
| 8.01 | 97.54 ± 0.38 | ||||||
| 8.74 | 0.14 ± 0.02 | ||||||
| B | C54H86O26S | 1227.4845 (1.5) | 24.65 ± 1.04 | 0.067 | 6.09 | 26.14 ± 0.56 | 0.67 ± 0.11 |
| 6.34 | 0.78 ± 0.14 | ||||||
| 8.47 | 26.87 ± 0.41 | ||||||
| 11.01 | 2.77 ± 1.40 | ||||||
| 11.16 | 68.91 ± 0.88 | ||||||
| C | C54H86O25S | 1211.4896 (1.2) | 10.82 ± 0.84 | 0.029 | 11.14 | 11.64 ± 1.22 | 3.80 ± 0.11 |
| 11.26 | 96.20 ± 0.11 | ||||||
| D | C41H64O17S | 905.3582 (1.2) | 13.41 ± 0.20 | 0.036 | 7.19 | 19.41 ± 1.04 | 0.92 ± 0.04 |
| 7.56 | 0.97 ± 0.08 | ||||||
| 9.25 | 0.32 ± 0.03 | ||||||
| 9.37 | 0.37 ± 0.01 | ||||||
| 11.02 | 97.42 ± 0.15 | ||||||
| E | C41H64O16S | 889.3632 (3.1) | 2.46 ± 0.67 | 0.007 | 7.29 | 3.63 ± 0.32 | 1.96 ± 0.59 |
| 7.37 | 1.66 ± 0.45 | ||||||
| 7.74 | 4.26 ± 0.81 | ||||||
| 11.36 | 53.64 ± 2.89 | ||||||
| 12.04 | 0.85 ± 0.16 | ||||||
| 12.16 | 37.63 ± 2.81 |
| Saponin | Composition | m/z (Δ ppm) [M+Na]+ | Retention Time (min) | Composition Molar Proportion (%) | Isomer Molar Proportion (%) |
|---|---|---|---|---|---|
| A’ | C54H84O23 | 1123.5301 (0.7) | 7.60 7.90 8.76 8.89 10.26 11.16 11.50 11.68 | 41.08 ± 0.26 | 0.93 ± 0.06 |
| 0.91 ± 0.04 | |||||
| 0.59 ± 0.09 | |||||
| 0.66 ± 0.09 | |||||
| 1.88 ± 0.06 | |||||
| 6.47 ± 0.22 | |||||
| 87.66 ± 0.08 | |||||
| 0.90 ± 0.16 | |||||
| B’ | C54H84O22 | 1107.5376 (1.9) | 8.39 8.61 9.13 11.67 12.04 | 25.34 ± 0.53 | 2.70 ± 0.12 |
| 2.42 ± 0.11 | |||||
| 7.71 ± 0.16 | |||||
| 59.82 ± 0.83 | |||||
| 27.35 ± 0.80 | |||||
| C’ | C54H84O23 | 1091.5403 (1.8) | 12.11 12.24 | 12.18 ± 0.32 | 96.37 ± 0.15 |
| 3.63 ± 0.15 | |||||
| D’ | C41H62O13 | 785.4038 (2.8) | 8.67 9.30 11.65 11.87 12.01 | 17.34 ± 0.51 | 0.43 ± 0.03 |
| 0.79 ± 0.05 | |||||
| 4.64 ± 0.08 | |||||
| 93.56 ± 0.05 | |||||
| 0.58 ± 0.01 | |||||
| E’ | C41H63O12 | 769.4139 (0.5) | 10.18 10.75 10.97 11.62 12.07 12.43 | 4.06 ± 0.26 | 2.06 ± 0.82 |
| 2.12 ± 0.43 | |||||
| 36.44 ± 0.69 | |||||
| 3.36 ± 0.22 | |||||
| 51.98 ± 0.18 | |||||
| 4.04 ± 0.43 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Savarino, P.; Colson, E.; Caulier, G.; Eeckhaut, I.; Flammang, P.; Gerbaux, P. Microwave-Assisted Desulfation of the Hemolytic Saponins Extracted from Holothuria scabra Viscera. Molecules 2022, 27, 537. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27020537
Savarino P, Colson E, Caulier G, Eeckhaut I, Flammang P, Gerbaux P. Microwave-Assisted Desulfation of the Hemolytic Saponins Extracted from Holothuria scabra Viscera. Molecules. 2022; 27(2):537. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27020537
Chicago/Turabian StyleSavarino, Philippe, Emmanuel Colson, Guillaume Caulier, Igor Eeckhaut, Patrick Flammang, and Pascal Gerbaux. 2022. "Microwave-Assisted Desulfation of the Hemolytic Saponins Extracted from Holothuria scabra Viscera" Molecules 27, no. 2: 537. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27020537