
Production of Bioactive Compounds with Broad Spectrum Bactericidal Action, Bio-Film Inhibition and Antilarval Potential by the Secondary Metabolites of the Endophytic Fungus Cochliobolus sp. APS1 Isolated from the Indian Medicinal Herb Andrographis paniculata



Abstract
:1. Introduction
2. Results
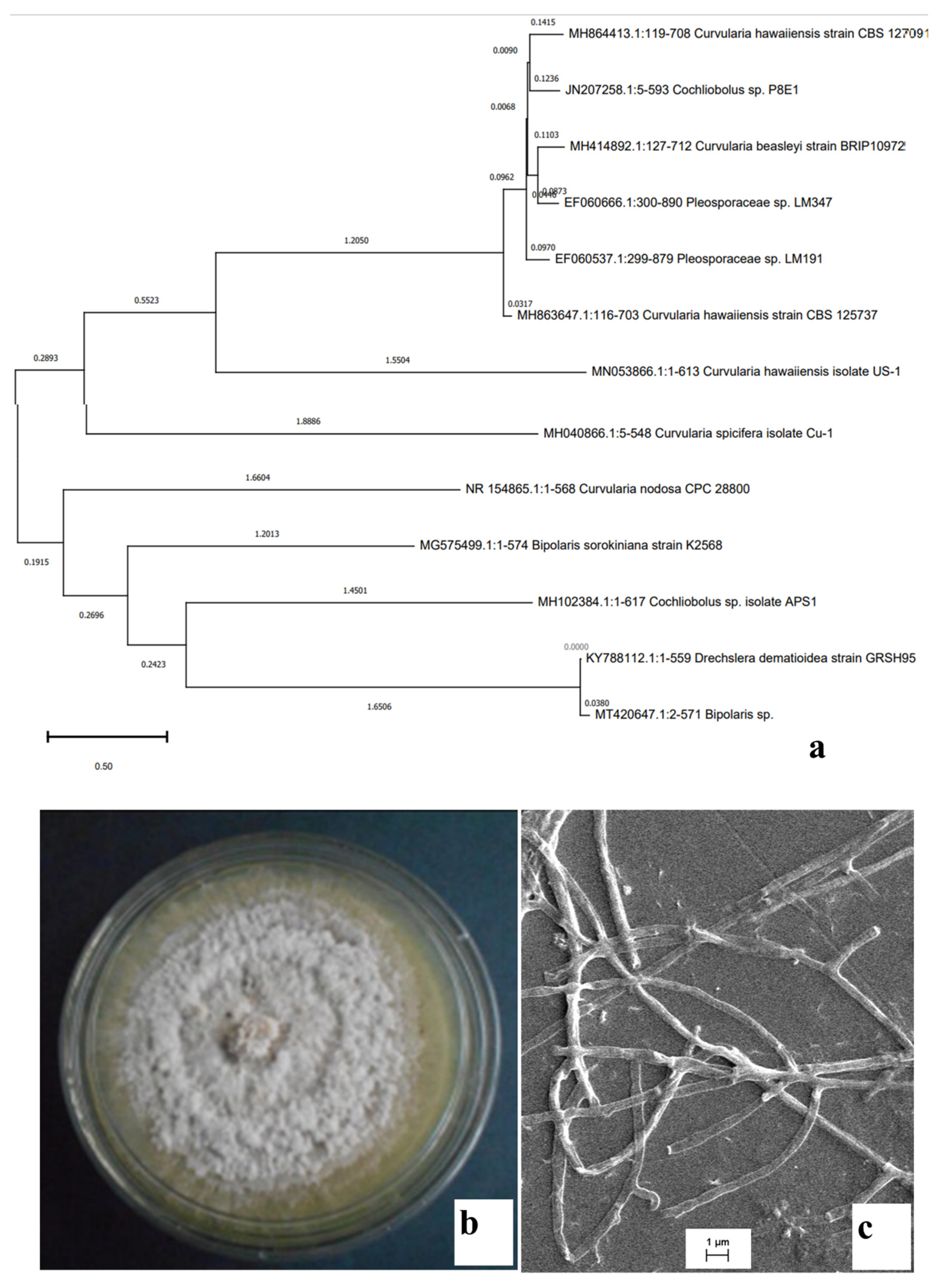
2.1. Identification of the Isolates
2.2. Antibacterial Activity of the Endophytic Fungal Isolates
2.3. Thermostable and Non-Proteinaceous Nature of the Antibacterial Components
2.4. Selection of the Appropriate Extraction Agent
2.5. MIC and MBC Determination
2.6. Effect on Bacterial Growth Kinetics
2.7. Leakage of Intracellular Materials
2.8. EA Fraction of APS1 Hampers the Central Carbohydrate Metabolism of the Pathogens
2.9. Synergistic Action of EA Fraction of APS1 and Antibiotic Ciprofloxacin
2.10. OVAT Optimization
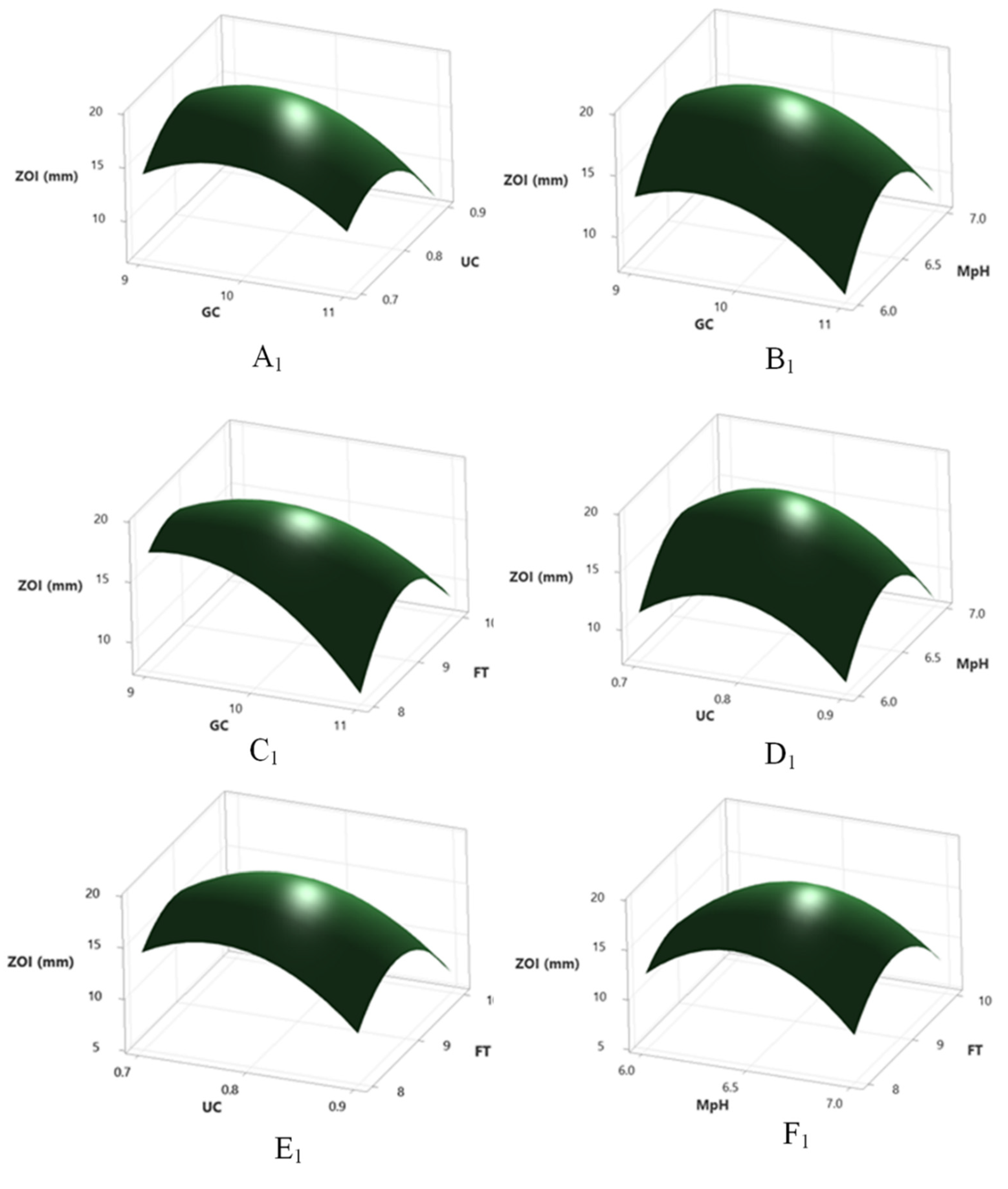
2.11. Optimization by RSM
2.12. Bio-Fim Inhibitory Activity of Cochliobolus sp. APS1
2.13. Larvicidal Potency
2.14. Identification of the Active Components
3. Discussion
4. Materials and Methods
4.1. Isolation and Identification of Endophytic Fungi
4.2. Study of Antibacterial Activity of the Cell Free Supernatant of Endophytic Fungal Isolate (Cochliobolus sp. APS1)
4.2.1. Antibacterial Action of Endophyte Cochliobolus sp. APS1
4.2.2. Determination of the Nature of Antibacterial Principles
4.2.3. Extraction of Antibacterial Components Using Solvent-Ethyl Acetate
4.2.4. Antibacterial Activity of EA Fraction of Endophytic APS1
4.2.5. Detection of MIC and MBC Values of Endophytic Metabolites
4.2.6. Time-Killing Kinetics
4.2.7. Detection of Release of Intracellular Material of MRSA upon Treatment by Cochliobolus sp. APS1 EA Extract
4.2.8. Effect of APS1 EA Fraction on Bacterial key Enzymes
4.3. Determination of Synergistic Activity of EA Fraction of APS1 and Antibiotic Ciprofloxacin by Checkerboard Method
4.4. Study of Inhibition of Bio-Film Formation
4.5. Detection of Antilarval Potency of Endophytic Isolates
4.5.1. Collection of Mosquito Larvae and Evaluation of Larvicidal Potency
4.5.2. Dose–Response Bioassay and Inhibition of Acetyl Choline Esterase Enzyme
4.6. Optimization for Antibacterial Production
4.7. Identification of the Active Components
4.7.1. Fourier Transformed Infrared Spectroscopy (FTIR)
4.7.2. GC-MS (Gas Chromatography and Mass Spectrometry) of the Active Compounds
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gómez-Lama Cabanás, C.; Schilirò, E.; Valverde-Corredor, A.; Mercado-Blanco, J. The biocontrol endophytic bacterium Pseudomonas fluorescens PICF7 induces systemic defense responses in aerial tissues upon colonization of olive roots. Front. Microbiol. 2014, 5, 427. [Google Scholar] [PubMed]
- Saikkonen, K.; Faeth, S.H.; Helander, M.; Sullivan, T.J. Fungal endophytes: A continuum of interactions with host plants. Annu. Rev. Ecol. Syst. 1998, 29, 319–343. [Google Scholar] [CrossRef]
- Redman, R.S.; Sheehan, K.B.; Stout, R.G.; Rodriguez, R.J.; Henson, J.M. Thermotolerance generated by plant/fungal symbiosis. Science 2002, 298, 1581. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, D.; Strobel, G.; Geary, B.; Sears, J.; Ezra, D.; Liarzi, O. Muscodor albus strain GBA, an endophytic fungus of Ginkgo biloba from United States of America produces volatile antimicrobials. Mycology 2010, 1, 179–186. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, D.; Strobel, G.; Booth, B.; Sears, J.; Spakowicz, D.; Busse, S. An endophytic Myrothecium inundatum producing volatile organic compounds. Mycosphere 2010, 1, 241–247. [Google Scholar]
- Banerjee, D.; Pandey, A.; Jana, M.; Strobel, G. Muscodor albus MOW12 an Endophyte of Piper nigrum L. (Piperaceae) Collected from North East India Produces Volatile Antimicrobials. Ind. J. Microbiol. 2014, 54, 27–32. [Google Scholar] [CrossRef] [Green Version]
- Katoch, M.; Phull, S.; Vaid, S.; Singh, S. Diversity, Phylogeny, anticancer and antimicrobial potential of fungal endophytes associated with Monarda citriodora L. BMC Microbiol. 2017, 17, 44. [Google Scholar] [CrossRef] [Green Version]
- Roy, S.; Parvin, R.; Ghosh, S.; Bhattacharya, S.; Maity, S.; Banerjee, D. Occurrence of a novel tannase (tan B LP) in endophytic Streptomyces sp. AL1L from the leaf of Ailanthus excelsa Roxb. 3 Biotech 2018, 8, 33. [Google Scholar] [CrossRef]
- Tomsheck, A.R.; Strobel, G.A.; Booth, E.; Geary, B.; Spakowicz, D.; Knighton, B.; Floerchinger, C.; Sears, J.; Liarzi, O.; Ezra, D. Hypoxylon sp., an endophyte of Persea indica, producing 1, 8-cineole and other bioactive volatiles with fuel potential. Microb. Ecol. 2010, 60, 903–914. [Google Scholar] [CrossRef]
- Clardy, J.; Walsh, C. Lessons from natural molecules. Nature 2004, 432, 829–837. [Google Scholar] [CrossRef]
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial biofilms: A common cause of persistent infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, P.S. Mechanisms of antibiotic resistance in bacterial biofilms. Int. J. Med. Microbiol. 2002, 292, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Brady, O.J.; Gething, P.W.; Bhatt, S.; Messina, J.P.; Brownstein, J.S.; Hoen, A.G.; Moyes, C.L.; Farlow, A.W.; Scott, T.W.; Hay, S.I. Refining the global spatial limits of dengue virus transmission by evidence-based consensus. PLoS Negl. Trop. Dis. 2012, 6, e1760. [Google Scholar] [CrossRef] [PubMed]
- Naqqash, M.N.; Gökçe, A.; Bakhsh, A.; Salim, M. Insecticide resistance and its molecular basis in urban insect pests. Parasitol. Res. 2016, 115, 1363–1373. [Google Scholar] [CrossRef]
- Kirschbaum, J.B. Potential implication of genetic engineering and other biotechnologies to insect control. Annu. Rev. Entomol. 1985, 30, 51–70. [Google Scholar] [CrossRef]
- Ragavendran, C.; Manigandan, V.; Kamaraj, C.; Balasubramani, G.; Prakash, J.S.; Perumal, P.; Natarajan, D. Larvicidal, histopathological, antibacterial activity of indigenous fungus Penicillium sp. against Aedes aegypti L and Culex quinquefasciatus (Say)(Diptera: Culicidae) and its acetylcholinesterase inhibition and toxicity assessment of zebrafish (Danio rerio). Front. Microbiol. 2019, 10, 427. [Google Scholar]
- Hossain, M.D.; Urbi, Z.; Sule, A.; Rahman, K.M. Andrographis paniculata (Burm. f.) Wall. ex Nees: A review of ethnobotany, phytochemistry, and pharmacology. Sci. World J. 2014, 2014, 274905. [Google Scholar] [CrossRef] [Green Version]
- Ham, J.E.; Wells, J.R. Surface chemistry of dihydromyrcenol (2, 6-dimethyl-7-octen-2-ol) with ozone on silanized glass, glass, and vinyl flooring tiles. Atmos. Environ. 2009, 43, 4023–4032. [Google Scholar] [CrossRef]
- Bicker, U. Biochemical and pharmacological properties of new 2-substituted aziridines. Contribution to experimental tumor research. Fortschr. Med. 1978, 96, 661–664. [Google Scholar]
- Fisher, N.L.; Burgess, L.W.; Toussoun, T.A.; Nelson, P.E. Carnation leaves as a substrate and for preserving cultures of Fusarium species. Phytopathology 1982, 72, 151–153. [Google Scholar] [CrossRef]
- Ismail, F.M.; Levitsky, D.O.; Dembitsky, V.M. Aziridine alkaloids as potential therapeutic agents. Eur. J. Med. Chem. 2009, 44, 3373–3387. [Google Scholar] [CrossRef] [PubMed]
- Dembitsky, V.M.; Terent’ev, A.O.; Levitsky, D.O. Aziridine Alkaloids: Origin, Chemistry and Activity. In Natural Products; Ramawat, K.G., Ed.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 977–1006. [Google Scholar]
- Dermer, O.C.; Ham, G.E. Ethylenimine and Other Aziridines; Chemistry and Application; Academic Press: New York, NY, USA, 1969. [Google Scholar]
- Stapley, E.O.; Hendlin, D.; Jackson, M.; Miller, A.K.; Hernandez, S.; Martinez, M. Azirinomycin. I. Microbial production and biological characteristics. J. Antibiot. 1971, 24, 42–47. [Google Scholar] [CrossRef]
- Argoudelis, A.D.; Reusser, F.; Whaley, H.A. Antibiotics produced by Streptomyces ficellus II. Feldamycin and nojirimycin. J. Antibiot. 1976, 29, 1007–1014. [Google Scholar] [CrossRef] [PubMed]
- Mahapatra, S.; Banerjee, D. Fungal exopolysaccharide: Production, composition and applications. Microbiol. Insight 2013, 6, MBI-S10957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; He, G. Influence of nutritional conditions on exopolysaccharide production by submerged cultivation of the medicinal fungus Shiraia bambusicola. World J. Microbiol. Biotechnol. 2008, 24, 2903–2907. [Google Scholar] [CrossRef]
- Xu, C.P.; Kim, S.W.; Hwang, H.J.; Choi, J.W.; Yun, J.W. Optimization of submerged culture conditions for mycelial growth and exo-biopolymer production by Paecilomyces tenuipes C240. Proc. Biochem. 2003, 38, 1025–1030. [Google Scholar] [CrossRef]
- Wonglumsom, W.; Vishnubhatla, A.; Fung, D.Y.C. Effect of volume of liquid enrichment medium containing Oxyrase@ on growth of Campylobacter jejuni. J. Rapid Methods Autom. Microbiol. 2000, 8, 111–139. [Google Scholar] [CrossRef]
- Huang, C.; Feng, Y.; Patel, G.; Xu, X.-Q.; Qian, J.; Liu, Q.; Kai, G.-Y. Production, immobilization and characterization of beta-glucosidase for application in cellulose degradation from a novel Aspergillus versicolor. Int. J. Biol. Macromol. 2021, 177, 437–446. [Google Scholar] [CrossRef]
- Patel, G.; Khobragade, T.P.; Avaghade, S.R.; Patil, M.D.; Nile, S.H.; Kai, G.; Banerjee, U.C. Optimization of media and culture conditions for the production of tacrolimus by Streptomyces tsukubaensis in shake flask and fermenter level. Biocatal. Agric. Biotechnol. 2020, 29, 101803. [Google Scholar] [CrossRef]
- Miller, T.W.; Tristram, E.W.; Wolf, F.J. Azirinomycin. II Isolation and chemical characterization as 3-methyl-2 (2H) azirinecarboxylic acid. J. Antibiot. 1971, 24, 48–50. [Google Scholar] [CrossRef] [Green Version]
- Tsuchida, T.; Iinuma, H.; Kinoshita, N.; Ikeda, T.; Sawa, T.; Hamada, M.; Takeuchi, T. Azicemicins A and B, a new antimicrobial agent produced by Amycolatopsis I. Taxonomy, fermentation, isolation, characterization and biological activities. J. Antibiot. 1995, 48, 217–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gracia, M. Torwards the Total Synthesis of Ficellomycin. Ph.D. Thesis, University of Warwick, Coventry, UK, 2007. [Google Scholar]
- Argoudelis, A.D.; Reusser, F.; Whaley, H.A.; Baczynskyj, L.; Mizsak, S.A.; Wnuk, R.J. Antibiotics produced by Streptomyces ficellus. I. Ficellomycin. J. Antibiot. 1976, 29, 1001–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; He, H.; Ma, R.; Ji, Z.; Wei, Q.; Dai, H.; Zhang, L.; Song, F. Madurastatin B3, a rare aziridine derivative from actinomycete Nocardiopsis sp. LS150010 with potent anti-tuberculosis activity. J. Ind. Microbiol. Biotechnol. 2017, 44, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.J.; Nam, S.-J.; Paul, L.; Beatty, D.; Kauffman, C.; Jensen, P.R.; Fenical, W. Previously uncultured marine bacteria linked to novel alkaloid production. Chem. Biol. 2015, 22, 1270–1279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabares, P.; Degel, B.; Schaschke, N.; Hentschel, U.; Schirmeister, T. Identification of the protease inhibitor miraziridine A in the Red sea sponge Theonella swinhoei. Pharmacogn. Res. 2012, 4, 63. [Google Scholar]
- Bhattacharya, S.; Ghosh, A.K.; Sen, D. A new alkaloid isolated from Abies webbiana leaf. Pharmacogn. Res. 2012, 2, 186. [Google Scholar] [CrossRef] [Green Version]
- Ruan, Q.; Patel, G.; Wang, J.; Luo, E.; Zhou, W.; Sieniawska, E.; Hao, X.; Kai, G. Current advances of endophytes as a platform for production of anti-cancer drug camptothecin. Food Chem. Toxicol. 2021, 151, 112113. [Google Scholar] [CrossRef]
- Mandal, S.; Maity, S.; Banerjee, D. Antioxidative Compounds from the Secreted Metabolome of Strain ‘Mucor irregularis Isolate Dro2′—An Endophyte of the Carnivorous Plant Drosera burmannii. Appl. Biochem. Microbiol. 2021, 57, S88–S97. [Google Scholar] [CrossRef]
- Santra, H.K.; Banerjee, D. Production, optimization, characterization and drought stress resistance by β-glucan rich heteropolysaccharide from an endophytic fungi Colletotrichum alatae LCS1 isolated from clubmoss (Lycopodium clavatum). Front. Fungal Biol. 2022, 2, 65. [Google Scholar] [CrossRef]
- Lagha, R.; Ben Abdallah, F.; Al-Sarhan, B.O.; Al-Sodany, Y. Antibacterial and biofilm inhibitory activity of medicinal plant essential oils against Escherichia coli isolated from UTI patients. Molecules 2019, 24, 1161. [Google Scholar] [CrossRef] [Green Version]
- Sandasi, M.; Leonard, C.M.; Viljoen, A.M. The effect of five common essential oil co mponents on Listeria monocytogenes biofilms. Food Control 2008, 19, 1070–1075. [Google Scholar] [CrossRef]
- Santra, H.K.; Banerjee, D. Natural products as fungicide and their role in crop protection. In Natural Bioactive Products in Sustainable Agriculture; Springer: Singapore, 2020; pp. 131–219. [Google Scholar]
- Santra, H.K.; Banerjee, D. Fungal Endophytes: A Source for Biological Control Agents. In Agriculturally Important Fungi for Sustainable Agriculture; Springer: Cham, Switzerland, 2020; pp. 181–216. [Google Scholar]
- Santra, H.K.; Banerjee, D. Microbial Exopolysaccharides: Structure and Therapeutic Properties. In Microbial Polymers; Springer: Singapore, 2021; pp. 375–420. [Google Scholar]
- Cannon, P.F.; Simmons, C.M. Diversity and host preference of leaf endophytic fungi in the Iwokrama Forest Reserve, Guyana. Mycology 2002, 94, 210–220. [Google Scholar] [CrossRef]
- Maity, S.; Mandal, S.; Santra, H.K.; Maity, S.K.; Banerjee, D. Broad-spectrum antimicrobial activity of Streptomyces griseus BLS4, a newly isolated endophyte of Blumea lacera. Res. J. Biotechnol. 2020, 15, 3. [Google Scholar]
- Mahapatra, S.; Banerjee, D. Structural elucidation and bioactivity of a novel exopolysaccharide from endophytic Fusarium solani SD5. Carbohydr. Polym. 2012, 90, 683–689. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Garayzabal, J.F.; Delgado, C.; Blanco, M.; Vazquez-Boland, J.A.; Briones, V.; Suárez, G.; Dominguez, L. Role of potassium tellurite and brain heart infusion in expression of the haemolytic phenotype of Listeria spp. on agar plates. Appl. Environ. Microbiol. 1992, 58, 434–438. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, S.; Ghosh, R.; Mandal, N.C. Production of bioactive compounds with bactericidal and antioxidant potential by endophytic fungus Alternaria alternata AE1 isolated from Azadirachta indica A. Juss. PLoS ONE 2019, 14, e0214744. [Google Scholar] [CrossRef] [Green Version]
- Zajmi, A.; Mohd Hashim, N.; Noordin, M.I.; Khalifa, S.A.; Ramli, F.; MohdAli, H.; El-Seedi, H.R. Ultrastructural study on the antibacterial activity of artonin e versus streptomycin against Staphylococcus aureus strains. PLoS ONE 2015, 10, e0128157. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Burton, K. A study of the conditions and mechanism of the diphenylamine reaction for the colorimetric estimation of deoxyribonucleic acid. Biochem. J. 1956, 62, 315. [Google Scholar] [CrossRef] [Green Version]
- Mandal, N.C.; Chakrabarty, P.K. Succinate-mediated catabolite repression of enzymes of glucose metabolism in root-nodule bacteria. Curr. Microbiol. 1993, 26, 247–251. [Google Scholar] [CrossRef]
- Orhan, G.; Bayram, A.; Zer, Y.; Balci, I. Synergy tests by E test and checkerboard methods of antimicrobial combinations against Brucella melitensis. J. Clin. Microbiol. 2005, 43, 140–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prinsloo, A.; Van Straten, A.M.S.; Weldhagen, G.F. Antibiotic synergy profiles of multidrug-resistant Pseudomonas aeruginosa in a nosocomial environment. S. Afr. J. Epidemiol. Infect. 2008, 23, 7–9. [Google Scholar]
- Jadhav, S.; Shah, R.; Bhave, M.; Palombo, E.A. Inhibitory activity of yarrow essential oil on Listeria planktonic cells and biofilms. Food Control 2013, 29, 125–130. [Google Scholar] [CrossRef]
- Rueda, L.M. Pictorial keys for the identification of mosquitoes (Diptera: Culicidae) associated with dengue virus transmission. Zootaxa 2004, 589, 1–60. [Google Scholar] [CrossRef]
- Tyagi, B.K.; Munirathinam, A.; Krishnamoorthy, R.; Venkatesh, A. A Field-Based Handbook of Identification Keys to Mosquitoes of Public Health Importance in India; Centre for Research in Medical Entomology: Madurai, India, 2012; pp. 25–27. [Google Scholar]
- Chandrasekar, G.; Arner, A.; Kitambi, S.S.; Dahlman-Wright, K.; Lendahl, M.A. Developmental toxicity of the environmental pollutant 4- nonylphenol in zebrafish. Neurotoxicol. Teratol. 2011, 33, 752–764. [Google Scholar] [CrossRef] [PubMed]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Finney, D.J. Probit Analysis; Cambridge University Press: London, UK, 1971; pp. 68–72. [Google Scholar]
- Mahapatra, S.; Banerjee, D. Production and structural elucidation of exopolysaccharide from endophytic Pestalotiopsis sp. BC55. Int. J. Biol. Macromol. 2016, 82, 182–191. [Google Scholar] [CrossRef]
- Mahapatra, S.; Banerjee, D. Optimization of a bioactive exopolysaccharide production from endophytic Fusarium solani SD5. Carbohydr. Polym. 2013, 97, 627–634. [Google Scholar] [CrossRef]
- Vivek, R.; Thangam, R.; Muthuchelian, K.; Gunasekaran, P.; Kaveri, K.; Kannan, S. Green biosynthesis of silver nanoparticles from Annona squamosa leaf extract and its in vitro cytotoxic effect on MCF-7 cells. Process Biochem. 2012, 47, 2405–2410. [Google Scholar] [CrossRef]
- R Core Team. A Language and Environment for Statistical Computing, Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2013. [Google Scholar]
- Venables, W.; Ripley, B. Modern Applied Statistics, 4th ed.; Springer: New York, NY, USA, 2002. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sl. No. | Human Bacterial Pathogens | Cochliobolus sp. APS1 | CPFX * | ||
|---|---|---|---|---|---|
| MIC Values (µg/mL) | MBC Values (µg/mL) | MIC Values (µg/mL) | MBC Values (µg/mL) | ||
| 1 | B. cereus (ATCC 14579) | 15.62 | 31.25 | 0.7 | 0.7 |
| 2 | B. subtilis (ATCC 11774) | 15.62 | 31.25 | 0.8 | 0.8 |
| 3 | S.aureus (ATCC 25923) | 31.25 | 62.5 | 0.8 | 0.8 |
| 4 | MRSA (ATCC 33591) | 62.5 | 125 | 1 | 1 |
| 5 | VRSA | 62.5 | 125 | 1 | 1 |
| 6 | P. mirabilis (ATCC 12453) | 125 | 250 | 1.2 | 1.2 |
| 7 | P. aeruginosa (ATCC 9027) | 62.5 | 125 | 1.2 | 1.2 |
| 8 | E. coli (MTCC 4296) | 125 | 250 | 1.4 | 1.4 |
| 9 | S. flexneri (ATCC 12022) | 62.5 | 125 | 1.5 | 1.5 |
| Ciprofloxacin (µg/mL) | 0.7 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 0.6 | 0.007 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| 0.5 | 0.098 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| 0.4 | 0.161 | 0.132 | 0.097 | 0 | 0 | 0 | 0 | 0 | |
| 0.3 | 0.209 | 0.168 | 0.101 | 0.051 | 0 | 0 | 0 | 0 | |
| 0.2 | 0.486 | 0.279 | 0.137 | 0.095 | 0.042 | 0 | 0 | 0 | |
| 0.1 | 0.792 | 0.478 | 0.371 | 0.279 | 0.097 | 0.037 | 0 | 0 | |
| 0 | 0.993 | 0.89 | 0.718 | 0.571 | 0.294 | 0.168 | 0.097 | 0 | |
| 0 | 0.99 | 1.985 | 3.97 | 7.81 | 15.62 | 31.25 | 62.5 | ||
| EA extract of APS1 (µg/mL) | |||||||||
| Parameters | Effectors | Percentage Added (g%) | Biomass (g/L) | Antibacterial Activity (ZOI in mm) |
|---|---|---|---|---|
| Incubation time (in day) | 3rd day | - | 4.665 ± 0.076 a | 4.33 ± 0.58 a |
| 5th day | - | 5.565 ± 0.134 b | 6 ± 0 b | |
| 7th day | - | 6.442 ± 0.064 c | 9.67 ± 0.58 c | |
| 9th day | - | 6.015 ± 0.039 c | 7.33 ± 0.58 d | |
| 11th day | - | 5.232 ± 0.076 b | 7 ± 1 d | |
| Incubation temperature (°C) | 22 | - | 7.069 ± 0.045 a | 9 ± 0 a |
| 24 | - | 7.855 ± 0.068 a | 10.33 ± 0.58 b | |
| 26 | - | 8.962 ± 0.049 b | 13.33 ± 0.58 c | |
| 28 | - | 6.062 ± 0.065 c | 10.67 ± 0.58 b | |
| 30 | - | 4.175 ± 0.034 d | 8 ± 0 d | |
| Initial medium pH | 4 | - | 3.385 ± 0.024 a | 3.33 ± 0.58 a |
| 5 | - | 5.005 ± 0.029 b | 4.33 ± 0.58 b | |
| 6 | - | 7.255 ± 0.068 c | 7 ± 1 c | |
| 6.5 | - | 8.995 ± 0.034 d | 9 ± 0 d | |
| 7 | - | 8.132 ± 0.068 d | 6.33 ± 0.58 c | |
| 7.5 | - | 7.995 ± 0.034 e | 5.33 ± 0.58 e | |
| 8 | - | 7.111 ± 0.03 c | 5 ± 0 e | |
| Additional carbon source | Starch | 1 | 8.976 ± 0.02 a | 9.67 ± 0.58 a |
| Fructose | 1 | 8.266 ± 0.036 b | 8.67 ± 0.58 b | |
| Glucose | 1 | 9.162 ± 0.073 a | 11 ± 1 c | |
| Maltose | 1 | 8.183 ± 0.037 b | 9 ± 1 b | |
| Additional nitrogen sources | Tryptone | 0.3 | 9.172 ± 0.023 a | 10.33 ± 0.58 a |
| Urea | 0.3 | 9.528 ± 0.029 a | 13 ± 1 b | |
| NH4NO3 | 0.3 | 9.165 ± 0.024 a | 9 ± 0.58 c | |
| Glucose concentration | Glucose | 2 | 9.522 ± 0.023 a | 11.33 ± 0.58 a |
| 4 | 9.842 ± 0.024 a | 12 ± 0 a | ||
| 6 | 9.965 ± 0.024 a | 12.33 ± 0.58 b | ||
| 8 | 10.213 ± 0.021 b | 13.33 ± 0.58 c | ||
| 10 | 10.841 ± 0.020 b | 17 ± 0 d | ||
| 12 | 8.904 ± 0.023 c | 11.33 ± 0.58 a | ||
| Urea concentration | 0.25 | - | 9.212 ± 0.027 a | 9.67 ± 0.58 a |
| 0.50 | - | 9.509 ± 0.029 a | 12.33 ± 0.58 b | |
| 0.70 | - | 10.164 ± 0.023 b | 14.67 ± 0.58 c | |
| 0.80 | - | 10.812 ± 0.030 b | 16.67 ± 0.58 d | |
| 1.00 | - | 8.304 ± 0.014 c | 13.33 ± 0.58 e | |
| Different metal ions | NaCl | 0.05 | 10.134 ± 0.034 a | 13 ± 1 a |
| KCl | 0.05 | 8.013 ± 0.009 b | 9.67 ± 0.58 b | |
| MgCl2 | 0.05 | 6.106 ± 0.029 c | 10.33 ± 0.58 c | |
| CaCl2 | 0.05 | 3.269 ± 0.027 d | 11.33 ± 0.58 d | |
| NaCl concentration | 0.05 | 8.982 ± 0.003 a | 14.33± 0.0.58 a | |
| 0.1 | 10.009 ± 0.013 b | 17.98 ± 0.58 b | ||
| 0.2 | 9.016 ± 0.040 a | 15.67 ± 0.58 c | ||
| 0.3 | 8.090 ± 0.009 c | 13 ± 1 d |
| Medium Volume (mL) | Total Volume of Flask (mL) | Head Space Volume (mL) | Medium Depth (cm) | Surface Area (cm) | Biomass (g/L) | Antibacterial Activity (ZOI, mm) |
|---|---|---|---|---|---|---|
| 30 | 320 | 290 | 1.2 | 3.19 | 3.964 ± 0.123 a | 6.6733 ±0.58 a |
| 60 | 320 | 260 | 1.8 | 2.68 | 4.976 ± 0.017 b | 8 ± 0.58 b |
| 90 | 320 | 230 | 2.4 | 2.13 | 5.412 ± 0.170 b | 9.33 ±0.58 c |
| 120 | 320 | 200 | 3 | 1.8 | 5.009 ± 0.119 b | 4.16 ±0.58 d |
| Wave Numbers (cm−1) | Peak Assignment | Mode of Vibration | Functional Group |
|---|---|---|---|
| 3373.61 | O–H stretching | Medium | Phenol |
| 2941.47 | C–H stretching | Medium | Alkenes |
| 1659.61 | C=C stretching | Medium | Alkane |
| 1411.79 | C–O bending | Medium | Inorganic carbonate |
| 1261.36 | C–O stretching | Medium | Alkyl aryl ether |
| 1037.39 | C–N stretching | Strong | Alkyl amine |
| 911.75 | O–H | Medium | Alcohol |
| Sl. No. | Name of the Compound | RT (min) | Area % | Ch. Formula | MW (g mol−1) |
|---|---|---|---|---|---|
| 1 | Phthalic acid | 2.86 | 17.64 | C8H6O4 | 166.14 |
| 2 | 2-((Z)-[(6-Bromo-8-quinolinyl)amino]methyl)phenol | 3.34 | 9.15 | C16H11BrN2O | 326 |
| 3 | 3-methyl-1-butanol | 4 | 0.64 | C5H12O | 88.14 |
| 4 | 1-ethylidene-1H-indene | 11 | 0.64 | C11H10 | 142 |
| 5 | 2-ethyl-p-cresol | 12.40 | 0.64 | C9H12O | 136 |
| 6 | 2,6-dimethyl-7-octen-2-ol (Dihydromyrcenol) | 12.74 | 6.49 | C10H20O | 156 |
| 7 | 2-ethyl-1-hexanol | 13.98 | 064 | C8H18O | 130 |
| 8 | 2-mthyl-1-propanol | 14 | 0.64 | C4H10O | 74.122 |
| 9 | 7-hexadecanal | 14.1 | 0.64 | C16H30O | 238 |
| 10 | 2-cis-9-octadecenyloxyethanol | 14.2 | 0.64 | C10H40O2 | 312 |
| 11 | Propanoic acid | 15 | 0.64 | C3H6O2 | 74.08 |
| 12 | Methyl 2-ethylhexanoate | 16.82 | 1.29 | C9H18O2 | 158 |
| 13 | Decyldecanoate | 17.53 | 10.38 | C20H40O2 | 312.5 |
| 14 | (2-Aziridinylethyl)amine | 18.31 | 64.93 | C4H10N2 | 86 |
| 15 | 1,3-benzodioxan | 21.28 | 1.94 | C8H8O2 | 136 |
| 16 | 10 methyl eicosane | 22.37 | 4.5 | C21H44 | 296 |
| 17 | 1-methylene-1H-indene | 26.77 | 0.64 | C10H8 | 128 |
| 18 | Undecane | 30.02 | 1.29 | C11H24 | 156 |
| 19 | Nonanol | 35.58 | 0.64 | C9H18O | 142 |
| 20 | 2-benzothiophene | 36 | 0.64 | C8H6S | 134 |
| 21 | 2,4-dimethyl-3-hexanone | 36.45 | 0.64 | C8H16O | 128 |
| 22 | Tridecane | 36.59 | 0.64 | C13H28 | 184 |
| 23 | Tetradecane | 37 | 0.64 | C14H30 | 198 |
| 24 | Hexahydropseudoionone | 37.44 | 0.64 | C13H26O | 198 |
| 25 | 1,3-di-iso-propylnapthalene | 45.09 | 0.64 | C16H20 | 212 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santra, H.K.; Maity, S.; Banerjee, D. Production of Bioactive Compounds with Broad Spectrum Bactericidal Action, Bio-Film Inhibition and Antilarval Potential by the Secondary Metabolites of the Endophytic Fungus Cochliobolus sp. APS1 Isolated from the Indian Medicinal Herb Andrographis paniculata. Molecules 2022, 27, 1459. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27051459
Santra HK, Maity S, Banerjee D. Production of Bioactive Compounds with Broad Spectrum Bactericidal Action, Bio-Film Inhibition and Antilarval Potential by the Secondary Metabolites of the Endophytic Fungus Cochliobolus sp. APS1 Isolated from the Indian Medicinal Herb Andrographis paniculata. Molecules. 2022; 27(5):1459. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27051459
Chicago/Turabian StyleSantra, Hiran Kanti, Santanu Maity, and Debdulal Banerjee. 2022. "Production of Bioactive Compounds with Broad Spectrum Bactericidal Action, Bio-Film Inhibition and Antilarval Potential by the Secondary Metabolites of the Endophytic Fungus Cochliobolus sp. APS1 Isolated from the Indian Medicinal Herb Andrographis paniculata" Molecules 27, no. 5: 1459. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27051459