
Positive Effect of Ecological Restoration with Fabaceous Species on Microbial Activities of Former Guyanese Mining Sites

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Soil Properties
2.2. C, N, P Mineralization and Microbial Biomass
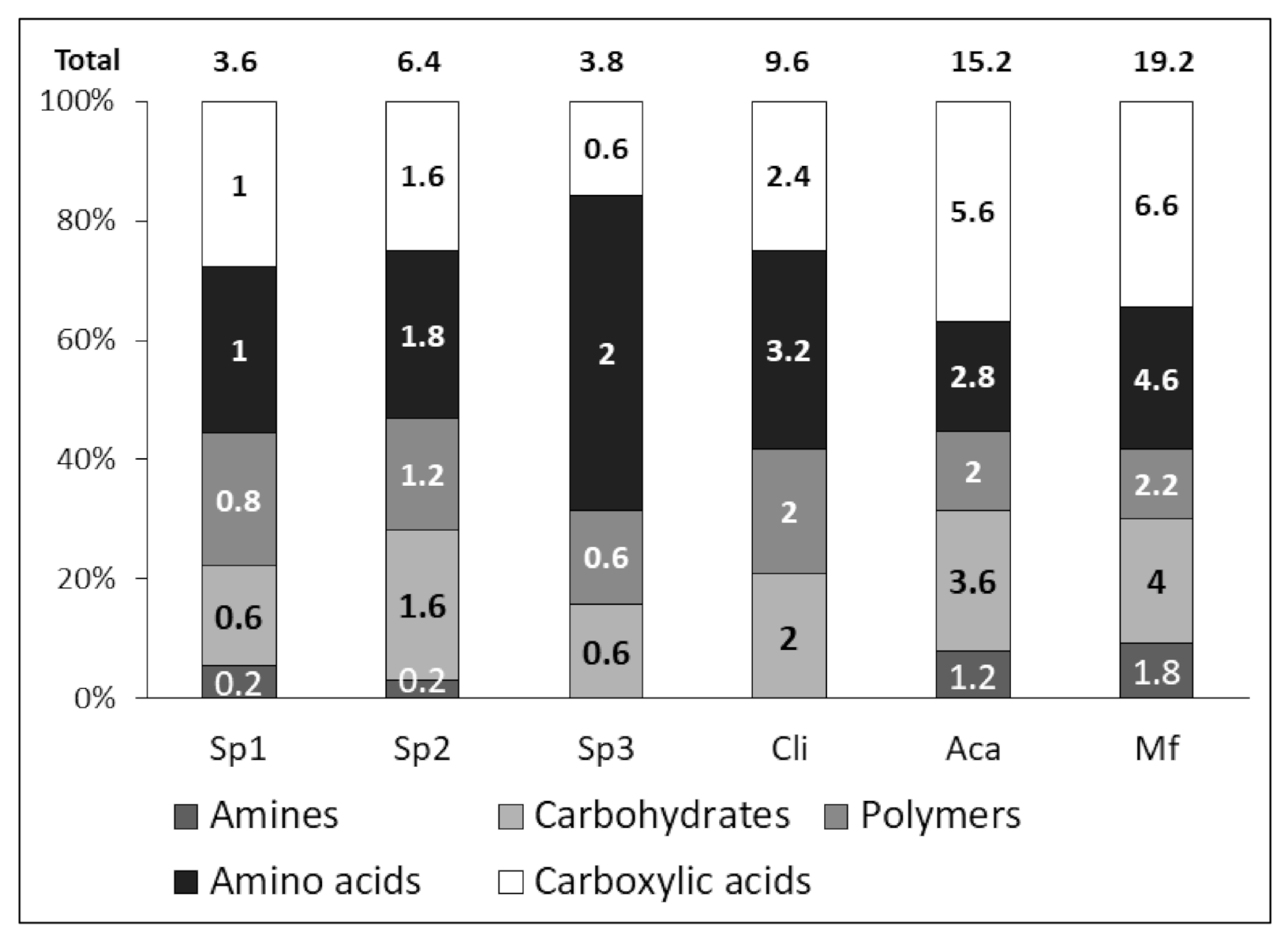
2.3. Catabolic Diversity
2.4. Effect of Ecological Restoration on Microbial Parameters
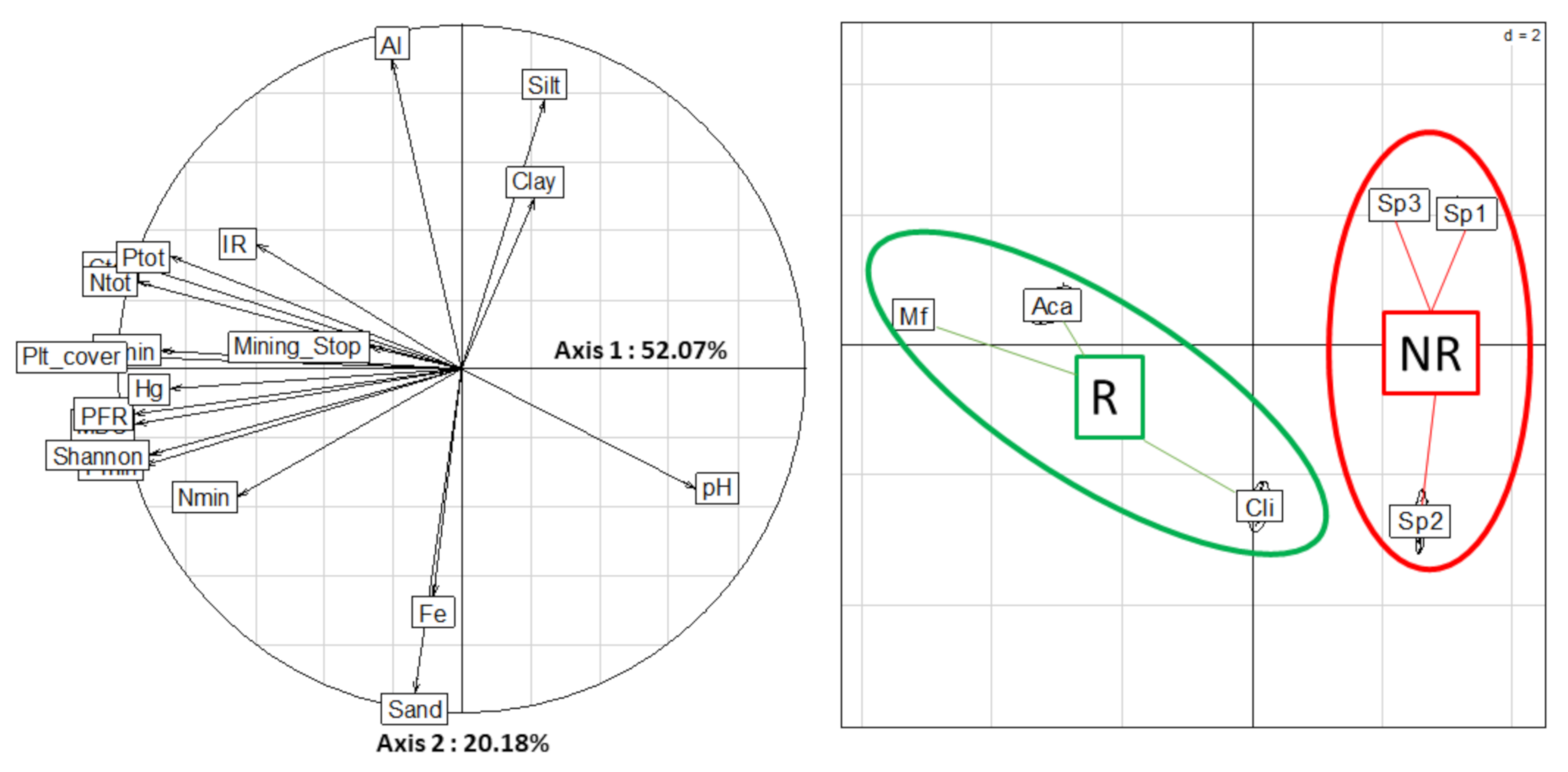
2.5. Overall Variability of the Database
3. Discussion
3.1. Positive Effect of Ecological Restoration on Nutrient Content in Soil
3.2. Positive Effect of Ecological Restoration on Soil Respiration and Organic Matter Mineralization
3.3. Positive Effect of Ecological Restoration on Catabolic Diversity and Diversity Index of Soil Microbial Communities
3.4. Effect of Restoration with Legumes
4. Materials and Methods
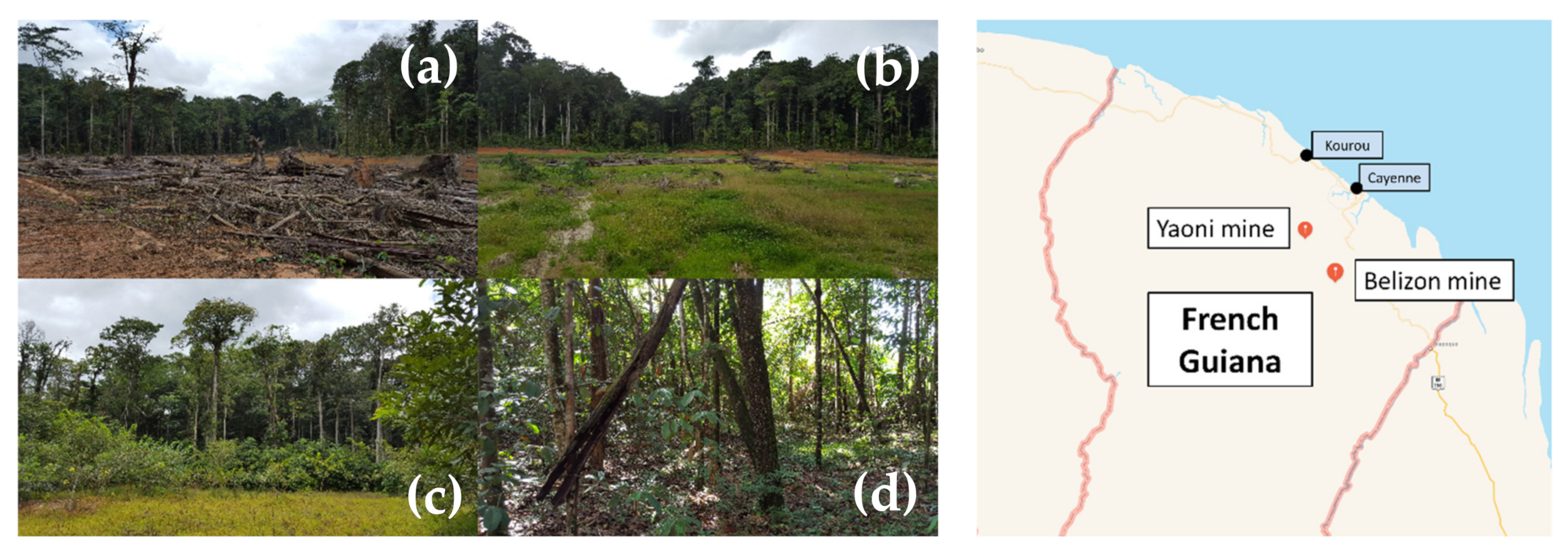
4.1. Site Description and Soil Sampling
4.2. Initial Soil Sample Characterization
4.3. Soil Granulometry Determination
4.4. Soil Total Carbon, Nitrogen, and Phosphorous Measurement
4.5. Soil Total Iron, Aluminum and Mercury Content
4.6. Soil Microbial Biomass
4.7. Catabolic Diversity Using Biolog EcoPlates
4.8. Mineralization Rate of Microbial Communities
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Carmouze, J.-P.; Lucotte, M.; Boudou, A. Le Mercure en Amazonie, 1st ed.; IRD Éditions: Bondy, France, 2001; ISBN 9782709914673. [Google Scholar]
- Hou, H.; Wang, C.; Ding, Z.; Zhang, S.; Yang, Y.; Ma, J.; Chen, F.; Li, J. Variation in the soil microbial community of reclaimed land over different reclamation periods. Sustainability 2018, 10, 2286. [Google Scholar] [CrossRef] [Green Version]
- Cardinale, B.J.; Srivastava, D.S.; Emmett Duffy, J.; Wright, J.P.; Downing, A.L.; Sankaran, M.; Jouseau, C. Effects of biodiversity on the functioning of trophic groups and ecosystems. Nature 2006, 443, 989–992. [Google Scholar] [CrossRef] [PubMed]
- Loubry, D. Livret Technique Pour la Conduite de la Revégétalisation sur les Surfaces Minières Alluvionanaires de Guyane. Available online: http://www.documentation.ird.fr/hor/fdi:010029513 (accessed on 22 September 2017).
- Chaer, G.M.; Resende, A.S.; Campello, E.F.C.; De Faria, S.M.; Boddey, R.M.; Schmidt, S. Nitrogen-fixing legume tree species for the reclamation of severely degraded lands in Brazil. Tree Physiol. 2011, 31, 139–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butterfield, R.P. Early species selection for tropical reforestation: A consideration of stability. For. Ecol. Manag. 1996, 81, 161–168. [Google Scholar] [CrossRef]
- Norisada, M.; Hitsuma, G.; Kuroda, K.; Yamanoshita, T.; Masumori, M.; Tange, T.; Yagi, H.; Nuyim, T.; Sasaki, S.; Kojima, K. Acacia mangium, a nurse tree candidate for reforestation on degraded sandy soils in the Malay Peninsula. For. Sci. 2005, 51, 498–510. [Google Scholar]
- Hulugalle, N.R. Amelioration of a highly degraded tropical alfisol by planting I. Changes in soil physical and chemical properties 1989–1891. Land Degrad. Dev. 1992, 3, 141–152. [Google Scholar] [CrossRef]
- Hutton, E.M. Selection and breeding of Leucaena for very acid soils. In Proceedings of the Leucaena Research in the Asian Pacific Region, Singapore, 23–26 November 1983. [Google Scholar]
- Guedron, S.; Grimaldi, M.; Grimaldi, C.; Cossa, D.; Tisserand, D.; Charlet, L. Amazonian former gold mined soils as a source of methylmercury: Evidence from a small scale watershed in French Guiana. Water Res. 2011, 45, 2659–2669. [Google Scholar] [CrossRef] [Green Version]
- Logsdon, S.D.; Karlen, D.L. Bulk density as a soil quality indicator during conversion to no-tillage. Soil Tillage Res. 2004, 78, 143–149. [Google Scholar] [CrossRef]
- Angers, D.A.; Pesant, A.; Vigneux, J. Early Cropping-Induced Changes in Soil Aggregation, Organic Matter, and Microbial Biomass. Soil Sci. Soc. Am. J. 1992, 56, 115. [Google Scholar] [CrossRef]
- Smith, J.L.; Doran, J.W.; Jones, A.J. Measurement and Use of pH and Electrical Conductivity for Soil Quality Analysis. Methods Assess. Soil Qual. 1996, 49, 169–185. [Google Scholar] [CrossRef]
- Karlen, D.; Andrews, S.; Wienhold, B.J.; Zobeck, T. Soil Quality Assessment: Past, Present and Future. Electron. J. Integr. Biosci. 2008, 6, 3–14. [Google Scholar]
- Ekenler, M.; Tabatabai, M.A. Tillage and residue management effects on β–glucosaminidase activity in soils. Soil Biol. Biochem. 2003, 35, 871–874. [Google Scholar] [CrossRef]
- Jordan, D.; Kremer, R.J.; Bergfield, W.A.; Kim, K.Y.; Cacnio, V.N. Evaluation of microbial methods as potential indicators of soil quality in historical agricultural fields. Biol. Fertil. Soils 1995, 19, 297–302. [Google Scholar] [CrossRef]
- Harris-Hellal, J.; Vallaeys, T.; Garnier-Zarli, E.; Bousserrhine, N. Effects of mercury on soil microbial communities in tropical soils of French Guyana. Appl. Soil Ecol. 2009, 41, 59–68. [Google Scholar] [CrossRef]
- Harris-Hellal, J.; Grimaldi, M.; Garnier-Zarli, E.; Bousserrhine, N. Mercury mobilization by chemical and microbial iron oxide reduction in soils of French Guyana. Biogeochemistry 2011, 103, 223–234. [Google Scholar] [CrossRef]
- Da Silva, E.; Nahmani, J.; Lapied, E.; Alphonse, V.; Garnier-Zarli, E.; Bousserrhine, N. Toxicity of mercury to the earthworm Pontoscolex corethrurus in a tropical soil of French Guiana. Appl. Soil Ecol. 2016, 104, 79–84. [Google Scholar] [CrossRef]
- Ekyastuti, W.; Ekamawanti, H.A. Short Communication: The role of microbial rhizosphere in enhancing plant growth of Jatropha curcas in soil contaminated mercury. Biodiversitas 2018, 19, 621–625. [Google Scholar] [CrossRef]
- Wong, M. Ecological restoration of mine degraded soils, with emphasis on metal contaminated soils. Chemosphere 2003, 50, 775–780. [Google Scholar] [CrossRef]
- Finžgar, N.; Kos, B.; Leštan, D. Bioavailability and mobility of Pb after soil treatment with different remediation methods. Plant Soil Environ. 2006, 52, 25. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zhou, X.; Yan, J.; Li, H.; He, J. Effects of regenerating vegetation on soil enzyme activity and microbial structure in reclaimed soils on a surface coal mine site. Appl. Soil Ecol. 2015, 87, 56–62. [Google Scholar] [CrossRef]
- Yang, D.; Zeng, D.; Mao, R.; Li, L. Restoration effect of young plantations on magnesite mine spoil in Northeast China. In Proceedings of the 2010 4th International Conference on Bioinformatics and Biomedical Engineering (iCBBE 2010), Chengdu, China, 18–20 June 2010. [Google Scholar]
- Singh, A.N.; Raghubanshi, A.S.; Singh, J.S. Impact of native tree plantations on mine spoil in a dry tropical environment. For. Ecol. Manag. 2004, 187, 49–60. [Google Scholar] [CrossRef]
- Tripathi, N.; Singh, R.S. Ecological restoration of mined-out areas of dry tropical environment, India. Environ. Monit. Assess. 2008, 146, 325–337. [Google Scholar] [CrossRef] [PubMed]
- Schimann, H.; Joffre, R.; Roggy, J.C.; Lensi, R.; Domenach, A.M. Evaluation of the recovery of microbial functions during soil restoration using near-infrared spectroscopy. Appl. Soil Ecol. 2007, 37, 223–232. [Google Scholar] [CrossRef]
- Couic, E.; Grimaldi, M.; Alphonse, V.; Balland-Bolou-Bi, C.; Livet, A.; Giusti-Miller, S.; Sarrazin, M.; Bousserrhine, N. Mercury behaviour and C, N, and P biogeochemical cycles during ecological restoration processes of old mining sites in French Guiana. Environ. Sci. Process. Impacts 2018, 20, 657–672. [Google Scholar] [CrossRef]
- Ross, D.J. Microbial biomass in a stored soil: A comparison of different estimation procedures. Soil Biol. Biochem. 1991, 23, 1005–1007. [Google Scholar] [CrossRef]
- Heděnec, P.; Vindušková, O.; Kukla, J.; Šnajdr, J.; Baldrian, P.; Frouz, J.; Frouz, J. Enzyme activity of topsoil layer on reclaimed and unreclaimed post-mining sites. Biol. Commun. 2017, 62, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Hobbs, R.J.; Harris, J.A. Restoration ecology: Repairing the earth’s ecosystems in the new millennium. Restor. Ecol. 2001, 9, 239–246. [Google Scholar] [CrossRef] [Green Version]
- Viana, R.M.; Ferraz, J.B.S.; Neves, A.F.; Vieira, G.; Pereira, B.F.F. Soil quality indicators for different restoration stages on Amazon rainforest. Soil Tillage Res. 2014, 140, 1–7. [Google Scholar] [CrossRef]
- Asmar, F. Variation in activity of root extracellular phytase between genotypes of barley. Plant Soil 1997, 195, 61–64. [Google Scholar] [CrossRef]
- Elfstrand, S.; Båth, B.; Mårtensson, A. Influence of various forms of green manure amendment on soil microbial community composition, enzyme activity and nutrient levels in leek. Appl. Soil Ecol. 2007, 36, 70–82. [Google Scholar] [CrossRef]
- Kang, H.; Freeman, C. Interactions of Marsh Orchid (Dactylorhiza spp.) and Soil Microorganisms in Relation to Extracellular Enzyme Activities in a Peat Soil. Pedosphere 2007, 17, 681–687. [Google Scholar] [CrossRef]
- Sonja, B.; Mariana Rodriguez, M.; Jacques, B.; Kela, P.W. Catabolic profiles of microbial communities in relation to plant identity and diversity in floating plant treatment wetlands. Ecol. Eng. 2014, 67, 190–197. [Google Scholar] [CrossRef]
- Schimann, H.; Petit-Jean, C.; Guitet, S.; Reis, T.; Domenach, A.M.; Roggy, J.C. Microbial bioindicators of soil functioning after disturbance: The case of gold mining in tropical rainforests of French Guiana. Ecol. Indic. 2012, 20, 34–41. [Google Scholar] [CrossRef]
- Zhu, L.X.; Xiao, Q.; Shen, Y.F.; Li, S.Q. Microbial functional diversity responses to 2 years since biochar application in silt-loam soils on the Loess Plateau. Ecotoxicol. Environ. Saf. 2017, 144, 578–584. [Google Scholar] [CrossRef]
- Asgharipour, M.R.; Rafiei, M. The effects of land use on biomass and catabolic diversity of soil microbial communities. Afr. J. Agric. Res. 2015, 6, 4607–4612. [Google Scholar] [CrossRef]
- Couic, E.; Alphonse, V.; Livet, A.; Giusti-Miller, S.; Bousserrhine, N. Influence of ecological restoration on mercury mobility and microbial activities on former guyanese mining sites. Appl. Sci. 2021, 11, 2231. [Google Scholar] [CrossRef]
- Lamb, D.; Lawrence, P. Mixed species plantations using high value rainforest trees in Australia. Restor. Trop. For. Ecosyst. 1993, 30, 101–108. [Google Scholar] [CrossRef]
- Cattanio, J.H.; Kuehne, R.; Vlek, P.L.G. Organic material decomposition and nutrient dynamics in a mulch system enriched with leguminous trees in the Amazon. Rev. Bras. Ciência Solo 2008, 32, 1073–1086. [Google Scholar] [CrossRef] [Green Version]
- Jaquetti, R.; Gonçalves, J.; Ferraz, J.; Ferreira, M.; Santos Junior, U. Ecofunctional Traits and Biomass Production in Leguminous Tree Species under Fertilization Treatments during Forest Restoration in Amazonia. Forests 2016, 7, 76. [Google Scholar] [CrossRef] [Green Version]
- Siddique, I.; Engel, V.L.; Parrotta, J.A.; Lamb, D.; Nardoto, G.B.; Ometto, J.P.H.B.; Martinelli, L.A.; Schmidt, S. Dominance of legume trees alters nutrient relations in mixed species forest restoration plantings within seven years. Biogeochemistry 2008, 88, 89–101. [Google Scholar] [CrossRef]
- Fierer, N.; Strickland, M.S.; Liptzin, D.; Bradford, M.A.; Cleveland, C.C. Global patterns in belowground communities. Ecol. Lett. 2009, 12, 1238–1249. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.P.E.; Domsch, K.H. A physiological method for the quantitative measurement of microbial biomass in soils. Soil Biol. Biochem. 1978, 10, 215–221. [Google Scholar] [CrossRef]
- Garland, J.L.; Mills, A.L. Classification and characterization of heterotrophic microbial communities on the basis of patterns of community-level sole-carbon-source utilization. Appl. Environ. Microbiol. 1991, 57, 2351–2359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Sites | Restored | Total | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ctot | DOC | Ntot | Ptot | C/N | pH-H2O | Fe | Al | Hg | Clay | Silt | Sand | ||
| g·kg−1 | g·kg−1 | g·kg−1 | g·kg−1 | g·kg−1 | g·kg−1 | µg·kg−1 | % | % | % | ||||
| Sp1 | NR | 6.01b ± 0.32 | 3.88ab ± 0.04 | 0.40b ± 0.03 | 0.21ab ± 0.04 | 15.02b ± 0.30 | 5.26ab ± 0.23 | 65.15b ± 3.17 | 53.88b ± 1.53 | 0.23a ± 0.01 | 46.4c ± 3.6 | 41.5b ± 2.8 | 3.3a ± 0.2 |
| Sp2 | NR | 3.52a ± 0.37 | 2.47a ± 0.23 | 0.14a ± 0.02 | 0.13a ± 0.01 | 25.14c ± 1.08 | 5.3b ± 0.13 | 66.2b ± 2.1 | 16.4a ± 0.6 | 0.27a ± 0.02 | 7.5a ± 2.7 | 21a ± 2 | 71.1d ± 1.6 |
| Sp3 | NR | 9.55b ± 0.73 | 5.84b ± 0.35 | 0.45b ± 0.01 | 0.27b ± 0.07 | 21.2c ± 1.27 | 4.66ab ± 0.07 | 44.56a ± 0.77 | 41.33b ± 1.77 | 0.23a ± 0.07 | 22.2b ± 5.1 | 65.8c ± 3.2 | 11.8b ± 1.2 |
| Cli | R | 7.01b ± 0.83 | 5.0b ± 0.4 | 0.40b ± 0.04 | 0.17a ± 0.02 | 17.52b ± 1.35 | 5.0ab ± 0.03 | 113.3c ± 4.2 | 25.5a ± 2.15 | 0.28a ± 0.03 | 21.3b ± 1.6 | 24.8a ± 3.7 | 53.9c ± 2.5 |
| Aca | R | 14.11c ± 1.2 | 10.55c ± 0.62 | 1.12c ± 0.07 | 0.37b ± 0.12 | 12.6a ± 0.54 | 4.21a ± 0.07 | 53.69ab ± 4.15 | 45.31b ± 6.51 | 0.48b ± 0.04 | 18.4b ± 2.5 | 35.6b ± 3.9 | 41.6c ± 4.4 |
| Mf | R | 22.88d ± 1.5 | 15.58d ± 1.41 | 1.81d ± 0.10 | 0.76c ± 0.15 | 12.64a ± 0.41 | 4.66ab ± 0.11 | 71.02b ± 1.37 | 43.76b ± 1.21 | 0.41b ± 0.05 | 19.8b ± 3.8 | 32.4b ± 4.9 | 36.4c ± 5.9 |
| Sites | Restored | MBC | Cmin | NO3−-Nmin | NH4+-Nmin | Ntotmin | Pmin |
|---|---|---|---|---|---|---|---|
| mg kg−1 | µg g−1day−1 | µg g−1day−1 | µg g−1day−1 | µg g−1day−1 | µg g−1day−1 | ||
| Sp1 | NR | 224.5a ± 13.2 | 4.45a ± 0.32 | 0.008a ± 0.001 | 0.13a ± 0.06 | 0.14a ± 0.07 | 0.017a ± 0.003 |
| Sp2 | NR | 390.5a ± 77.3 | 4.96a ± 1.1 | 0.07b ± 0.006 | 0.8b ± 0.2 | 0.88c ± 0.2 | 0.024a ± 0.002 |
| Sp3 | NR | 312.6a ± 10.1 | 5.77a ± 0.37 | 0.004a ± 0.0003 | 0.45ab ± 0.05 | 0.46b ± 0.05 | 0.03b ± 0.002 |
| Cli | R | 845.2b ± 38.5 | 35.37b ± 1.4 | 1.56d ± 0.35 | 1.15c ± 0.15 | 2.7e ± 0.44 | 0.047c ± 0.007 |
| Aca | R | 909b ± 69 | 39.14bc ± 1.9 | 0.33c ± 0.12 | 1.2c ± 0.32 | 1.55d ± 0.33 | 0.058cd ± 0.006 |
| Mf | R | 1103b ± 88 | 46.58c ± 1.5 | 0.35c ± 0.11 | 1.7d ± 0.14 | 2.03e ± 0.07 | 0.065d ± 0.008 |
| Sites | Restored | AWCD | PFR | Catabolic Diversity | H | EH |
|---|---|---|---|---|---|---|
| Sp1 | NR | 0.08a ± 0.01 | 3.6a ± 0.9 | 0.12a ± 0.02 | 1.2a ± 0.11 | 0.76a ± 0.05 |
| Sp2 | NR | 0.18b ± 0.1 | 6.6ab ± 2.1 | 0.21b ± 0.07 | 1.9ab ± 0.47 | 0.53a ± 0.25 |
| Sp3 | NR | 0.09a ± 0.02 | 3.8a ± 0.8 | 0.12a ± 0.03 | 1.3a ± 0.44 | 0.88b ± 0.036 |
| Cli | R | 0.34c ± 0.13 | 9.6b ± 1.9 | 0.31c± 0.06 | 2.1b ± 0.12 | 0.92b ± 0.015 |
| Aca | R | 0.32c ± 0.1 | 15.2c ± 3.2 | 0.49d ± 0.11 | 2.6c ± 0.22 | 0.95b ± 0.027 |
| Mf | R | 0.49d ± 0.09 | 19.2c ± 1.1 | 0.62d ± 0.04 | 3.3c ± 0.11 | 0.98b ± 0.01 |
| Sites | Level of Restoration | ||
|---|---|---|---|
| Df | 5 | 1 | |
| Total organic carbon | F-value | 4472 ** | 25 ** |
| Total nitrogen | 1529 ** | 22 ** | |
| Total phosphorus | 224 ** | 11 * | |
| Microbial biomass carbon | 1330 ** | 322 ** | |
| Carbon mineralization | 480 ** | 20 ** | |
| Nitrate mineralization (NO3−-Nmin) | 67 ** | 18 ** | |
| Ammonium mineralization | 49 ** | 60 ** | |
| Phosphorus mineralization | 58 ** | 94 ** | |
| Average well color development (AWCD) | 15 ** | 47 * | |
| Potential functional richness | 56 ** | 60 ** | |
| Catabolic diversity | 5 ** | 60 ** | |
| Shannon Index | 37 ** | 44 ** | |
| Regularity index | 11 * | 17 * |
| Variables | MBC | C_min | NO3_min | NH4_min | Ntot_min | P_min | PFR | Diversity | H | EH | AWCD |
|---|---|---|---|---|---|---|---|---|---|---|---|
| MBC | 1.00 | 0.80 | 0.50 | 0.92 | 0.84 | 0.93 | 0.89 | 0.89 | 0.85 | 0.64 | 0.83 |
| C_min | - | 1.00 | 0.11 | 0.79 | 0.53 | 0.75 | 0.82 | 0.82 | 0.82 | 0.47 | 0.74 |
| NO3_min | - | - | 1.00 | 0.40 | 0.84 | 0.36 | 0.20 | 0.20 | 0.22 | 0.35 | 0.35 |
| NH4_min | - | - | - | 1.00 | 0.83 | 0.80 | 0.77 | 0.77 | 0.77 | 0.59 | 0.72 |
| Ntot_min | - | - | - | - | 1.00 | 0.69 | 0.57 | 0.57 | 0.58 | 0.56 | 0.64 |
| P_min | - | - | - | - | - | 1.00 | 0.93 | 0.93 | 0.89 | 0.48 | 0.85 |
| PFR | - | - | - | - | - | - | 1.00 | 1.00 | 0.95 | 0.43 | 0.90 |
| Diversity | - | - | - | - | - | - | - | 1.00 | 0.95 | 0.43 | 0.90 |
| H | - | - | - | - | - | - | - | - | 1.00 | 0.28 | 0.87 |
| EH | - | - | - | - | - | - | - | - | - | 1.00 | 0.35 |
| AWCD | - | - | - | - | - | - | - | - | - | - | 1.00 |
| Sites | Sp1 | Sp2 | Sp3 | Cli | Aca | Mf |
|---|---|---|---|---|---|---|
| level of restoration | Non-restored | Non-restored | Non-restored | Restored | Restored | Restored |
| Pioneer species | Herbaceous species | Cyperus spp. and Carex spp. | Lycopodiella spp. | Clitoria racemosa | Acacia mangium | C. racemosa and A. mangium |
| Type of vegetation cover | Almost bare soil | Grassland | Grassland | Coppiced forest | Dense forest | Dense forest |
| End of mining (year) | 1998 | 1998 | 2013 | 2013 | 1998 | 1998 |
| Geographical coordinates | N 04°30.311′/ W 052°26.919′ | N 04°30.311′/ W 052°26.919′ | N 04°22.509′/ W 052°20.739′ | N 04°29.860′/ W 052°26.966′ | N 04°29.860′/ W 052°26.966′ | N 04°29.860′/ W 052°26.966′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Couic, E.; Tribondeau, A.; Alphonse, V.; Livet, A.; Bousserrhine, N. Positive Effect of Ecological Restoration with Fabaceous Species on Microbial Activities of Former Guyanese Mining Sites. Molecules 2022, 27, 1768. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27061768
Couic E, Tribondeau A, Alphonse V, Livet A, Bousserrhine N. Positive Effect of Ecological Restoration with Fabaceous Species on Microbial Activities of Former Guyanese Mining Sites. Molecules. 2022; 27(6):1768. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27061768
Chicago/Turabian StyleCouic, Ewan, Alicia Tribondeau, Vanessa Alphonse, Alexandre Livet, and Noureddine Bousserrhine. 2022. "Positive Effect of Ecological Restoration with Fabaceous Species on Microbial Activities of Former Guyanese Mining Sites" Molecules 27, no. 6: 1768. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27061768