
Protective Effects of a Polyphenol-Rich Blueberry Extract on Adult Human Neural Progenitor Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
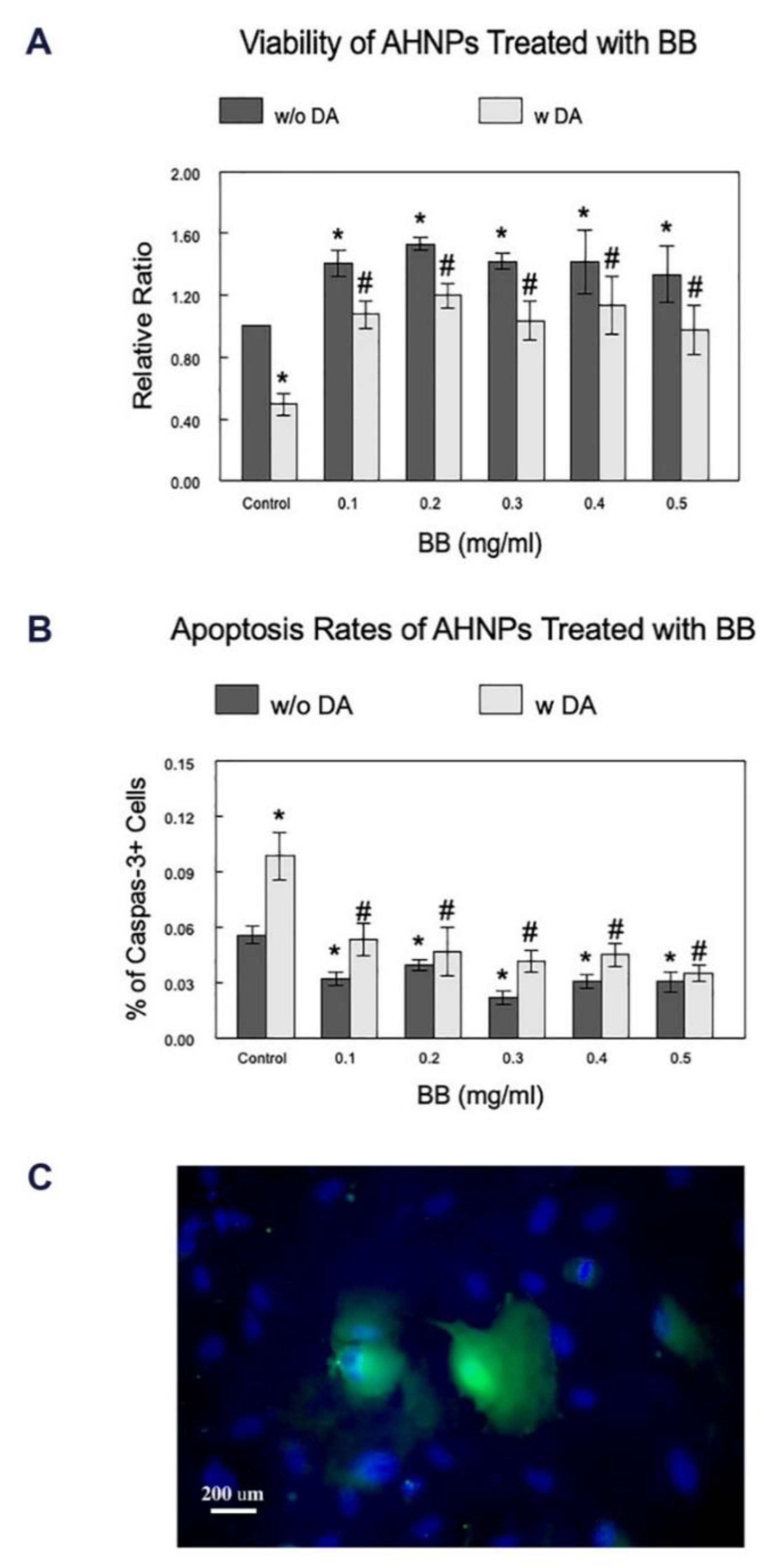
2.1. Effects of BB on the Viability of AHNPs
2.2. Effects of BB on the Apoptosis Rates of AHNPs
2.3. Effects of BB on the Proliferation of AHNPs
2.4. Effects of BB on the Phenotyping of AHNPs
2.5. Effects of BB on the Calcium Buffering Ability of AHNPs
2.6. Effects of BB on Expression of Oxidative and Inflammatory Markers of the AHNPs
3. Materials and Methods
3.1. Cell Culture
3.2. Blueberry Extract Preparation and Treatments
3.3. Cellular Stressor Treatments
3.4. Cell Viability Assay
3.5. Proliferation Assay
3.6. Calcium (Ca2+) Imaging
3.7. Phenotyping
3.8. Western Blot
3.9. Statistical Analyses
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AHNPs | Adult hippocampal human neural progenitor cells |
| ANOVA | Analyses of variance |
| BBs | Blueberries |
| bFGF | Basic fibroblast growth factor |
| Ca2+ | Calcium |
| CNS | Central nervous system |
| DA | Dopamine |
| DMEM | Dulbecco’s modified Eagle’s medium |
| ECL | Electrochemiluminescence |
| EdU | 5-ethynyl-2′-deoxyuridine |
| EGF | Epidermal growth factor |
| FBS | Fetal bovine serum |
| iNOS | Inducible nitric oxide synthase |
| LPS | Lipopolysaccharide |
| NOX-2 | NADPH oxidase 2 |
| NSC | Neuronal stem cell |
| PBSt | PBS supplemented with Tween-20 |
| SGZ | Subgranular zone |
| SVZ | Subventricular zone |
References
- Ming, G.-L.; Song, H. Adult neurogenesis in the mammalian brain: Significant answers and significant questions. Neuron 2011, 70, 687–702. [Google Scholar] [CrossRef]
- Berdugo-Vega, G.; Arias-Gil, G.; Lopez-Fernandez, A.; Artegiani, B.; Wasielewska, J.M.; Lee, C.C.; Lippert, M.T.; Kempermann, G.; Takagaki, K.; Calegari, F. Increasing neurogenesis refines hippocampal activity rejuvenating navigational learning strategies and contextual memory throughout life. Nat. Commun. 2020, 11, 135. [Google Scholar] [CrossRef]
- Shohayeb, B.; Diab, M.; Ahmed, M.; Ng, D.C.H. Factors that influence adult neurogenesis as potential therapy. Transl. Neurodegener. 2018, 7, 4. [Google Scholar] [CrossRef] [PubMed]
- Galvan, V.; Jin, K. Neurogenesis in the aging brain. Clin. Interv. Aging. 2007, 2, 605–610. [Google Scholar]
- Kuhn, H.-G.; Dickinson-Anson, H.; Gage, F.H. Neurogenesis in the dentate gyrus of the adult rat: Age-related decrease of neuronal progenitor proliferation. J. Neurosci. 1996, 16, 2027–2033. [Google Scholar] [CrossRef]
- Foster, T.C. Biological markers of age-related memory deficits: Treatment of senescent physiology. CNS Drugs 2006, 20, 153–166. [Google Scholar] [CrossRef]
- Urbán, N.; Blomfield, I.M.; Guillemot, F. Quiescence of adult mammalian neural stem cells: A highly regulated rest. Neuron 2019, 104, 834–848. [Google Scholar] [CrossRef]
- Ojo, J.O.; Rezaie, P.; Gabbott, P.L.; Stewart, M.G. Impact of age-related neuroglial cell responses on hippocampal deterioration. Front. Aging Neurosci. 2015, 7, 57. [Google Scholar] [CrossRef]
- Solano Fonseca, R.; Mahesula, S.; Apple, D.M.; Raghunathan, R.; Dugan, A.; Cardona, A.; O’Connor, J.; Kokovay, E. Neurogenic niche microglia undergo positional remodeling and progressive activation contributing to age-associated reductions in neurogenesis. Stem Cells Dev. 2016, 25, 542–555. [Google Scholar] [CrossRef]
- Ekdahl, C.T.; Claasen, J.-H.; Bonde, S.; Kokaia, Z.; Lindvall, O. Inflammation is detrimental for neurogenesis in adult brain. Proc. Natl. Acad. Sci. USA 2003, 100, 13632–13637. [Google Scholar] [CrossRef]
- Chesnokova, V.; Pechnick, R.N.; Wawrowsky, K. Chronic peripheral inflammation, hippocampal neurogenesis, and behavior. Brain Behav. Immun. 2016, 58, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Steindler, D.A.; Reynolds, B.A. Perspective: Neuroregenerative nutrition. Adv. Nutr. Int. Rev. J. 2017, 8, 546–557. [Google Scholar] [CrossRef]
- Kelly, E.; Vyas, P.; Weber, J.T. Biochemical properties and neuroprotective effects of compounds in various species of berries. Molecules 2017, 23, 26. [Google Scholar] [CrossRef]
- Ma, L.; Sun, Z.; Zeng, Y.; Luo, M.; Yang, J. Molecular mechanism and health role of functional ingredients in blueberry for chronic disease in human beings. Int. J. Mol. Sci. 2018, 19, 2785. [Google Scholar] [CrossRef]
- Andres-Lacueva, C.; Shukitt-Hale, B.; Galli, R.L.; Jauregui, O.; Lamuela-Raventos, R.M.; Joseph, J.A. Anthocyanins in aged blueberry-fed rats are found centrally and may enhance memory. Nutr. Neurosci. 2005, 8, 111–120. [Google Scholar] [CrossRef]
- Bensalem, J.; Dudonné, S.; Gaudout, D.; Servant, L.; Calon, F.; Desjardins, Y.; Layé, S.; Lafenetre, P.; Pallet, V. Polyphenol-rich extract from grape and blueberry attenuates cognitive decline and improves neuronal function in aged mice. J. Nutr. Sci. 2018, 7, e19. [Google Scholar] [CrossRef]
- Heitman, E.; Ingram, D.K. Cognitive and neuroprotective effects of chlorogenic acid. Nutr. Neurosci. 2017, 20, 32–39. [Google Scholar] [CrossRef]
- Özduran, G.; Becer, E.; Vatansever, H.S. The role and mechanisms of action of catechins in neurodegenerative diseases. J. Am. Coll. Nutr. 2021, 1–8. [Google Scholar] [CrossRef]
- Joseph, J.A.; Shukitt-Hale, B.; Brewer, G.J.; Weikel, K.A.; Kalt, W.; Fisher, D.R. Differential protection among fractionated blueberry polyphenolic families against DA-, Aβ42- and LPS-induced decrements in Ca2+ buffering in primary hippocampal cells. J. Agric. Food Chem. 2010, 58, 8196–8204. [Google Scholar] [CrossRef]
- Ma, H.; Johnson, S.L.; Liu, W.; DaSilva, N.A.; Meschwitz, S.; Dain, J.A.; Seeram, N.P. Evaluation of polyphenol anthocyanin-enriched extracts of blackberry, black raspberry, blueberry, cranberry, red raspberry, and strawberry for free radical scavenging, reactive carbonyl species trapping, anti-glycation, anti-β-amyloid aggregation, and microglial neuroprotective effects. Int. J. Mol. Sci. 2018, 19, 461. [Google Scholar] [CrossRef]
- Shukitt-Hale, B.; Bielinski, D.F.; Lau, F.C.; Willis, L.M.; Carey, A.N.; Joseph, J.A. The beneficial effects of berries on cognition, motor behaviour and neuronal function in ageing. Br. J. Nutr. 2015, 114, 1542–1549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.; Hamilton, D.A.; Joseph, J.A.; Shukitt-Hale, B. Dietary blueberry improves cognition among older adults in a randomized, double-blind, placebo-controlled trial. Eur. J. Nutr. 2018, 57, 1169–1180. [Google Scholar] [CrossRef]
- Shishtar, E.; Rogers, G.T.; Blumberg, J.B.; Au, R.; Jacques, P.F. Long-term dietary flavonoid intake and change in cognitive function in the Framingham Offspring cohort. Public Health Nutr. 2020, 23, 1576–1588. [Google Scholar] [CrossRef]
- Joseph, J.A.; Fisher, D.R.; Carey, A.N. Fruit extracts antagonize Abeta- or DA-induced deficits in Ca2+ flux in M1-transfected COS-7 cells. J. Alzheimers Dis. 2004, 6, 403–411; discussion 443–449. [Google Scholar] [CrossRef]
- Galli, R.L.; Bielinski, D.F.; Szprengiel, A.; Shukitt-Hale, B.; Joseph, J.A. Blueberry supplemented diet reverses age-related decline in hippocampal HSP70 neuroprotection. Neurobiol. Aging 2006, 27, 344–350. [Google Scholar] [CrossRef]
- Duffy, K.B.; Spangler, E.L.; Devan, B.D.; Guo, Z.; Bowker, J.L.; Janas, A.M.; Hagepanos, A.; Minor, R.K.; DeCabo, R.; Mouton, P.R.; et al. A blueberry-enriched diet provides cellular protection against oxidative stress and reduces a kainate-induced learning impairment in rats. Neurobiol. Aging 2008, 29, 1680–1689. [Google Scholar] [CrossRef]
- Olguín, V.; Durán, A.; Heras, M.L.; Rubilar, J.C.; Cubillos, F.A.; Olguín, P.; Klein, A.D. Genetic background matters: Population-based studies in model organisms for translational research. Int. J. Mol. Sci. 2022, 23, 7570. [Google Scholar] [CrossRef]
- Beaulieu-Laroche, L.; Brown, N.J.; Hansen, M.; Toloza, E.H.S.; Sharma, J.; Williams, Z.M.; Frosch, M.P.; Cosgrove, G.R.; Cash, S.S.; Harnett, M.T. Allometric rules for mammalian cortical layer 5 neuron biophysics. Nature 2021, 600, 274–278. [Google Scholar] [CrossRef]
- Soukup, S.T.; Helppi, J.; Müller, D.R.; Zierau, O.; Watzl, B.; Vollmer, G.; Diel, P.; Bub, A.; Kulling, S.E. Phase II metabolism of the soy isoflavones genistein and daidzein in humans, rats and mice: A cross-species and sex comparison. Arch. Toxicol. 2016, 90, 1335–1347. [Google Scholar] [CrossRef]
- Spalding, K.L.; Bergmann, O.; Alkass, K.; Bernard, S.; Salehpour, M.; Huttner, H.B.; Boström, E.; Westerlund, I.; Vial, C.; Buchholz, B.A.; et al. Dynamics of hippocampal neurogenesis in adult humans. Cell 2013, 153, 1219–1227. [Google Scholar] [CrossRef]
- Walton, N.M.; Sutter, B.M.; Chen, H.-X.; Chang, L.-J.; Roper, S.N.; Scheffler, B.; Steindler, D.A. Derivation and large-scale expansion of multipotent astroglial neural progenitors from adult human brain. Development 2006, 133, 3671–3681. [Google Scholar] [CrossRef] [Green Version]
- Joseph, J.A.; Shukitt-Hale, B.; Denisova, N.A.; Bielinski, D.; Martin, A.; McEwen, J.J.; Bickford, P. Reversals of age-related declines in neuronal signal transduction, cognitive, and motor behavioral deficits with blueberry, spinach, or strawberry dietary supplementation. J. Neurosci. 1999, 19, 8114–8121. [Google Scholar] [CrossRef]
- Ganguly, U.; Ganguly, A.; Sen, O.; Ganguly, G.; Cappai, R.; Sahoo, A.; Chakrabarti, S. Dopamine Cytotoxicity on SH-SY5Y Cells: Involvement of α-Synuclein and Relevance in the Neurodegeneration of Sporadic Parkinson’s Disease. Neurotox. Res. 2019, 35, 898–907. [Google Scholar] [CrossRef]
- Strober, W. Trypan blue exclusion test of cell viability. Curr. Protoc. Immunol. 2015, 111, A3 B 1–A3 B 3. [Google Scholar] [CrossRef]
- Salic, A.; Mitchison, T.J. A chemical method for fast and sensitive detection of DNA synthesis in vivo. Proc. Natl. Acad. Sci. USA 2008, 105, 2415–2420. [Google Scholar] [CrossRef]
- Poulose, S.M.; Fisher, D.R.; Larson, J.; Bielinski, D.F.; Rimando, A.M.; Carey, A.N.; Schauss, A.G.; Shukitt-Hale, B. Anthocyanin-rich açai (Euterpe oleracea Mart.) fruit pulp fractions attenuate inflammatory stress signaling in mouse brain BV-2 microglial cells. J. Agric. Food Chem. 2012, 60, 1084–1093. [Google Scholar] [CrossRef]
- Adusumilli, V.S.; Walker, T.L.; Overall, R.W.; Klatt, G.M.; Zeidan, S.A.; Zocher, S.; Kirova, D.G.; Ntitsias, K.; Fischer, T.J.; Sykes, A.M.; et al. ROS dynamics delineate functional states of hippocampal neural stem cells and link to their activity-dependent exit from quiescence. Cell Stem Cell 2020, 28, 300–314.e6. [Google Scholar] [CrossRef]
- Carey, A.N.; Gildawie, K.; Rovnak, A.; Thangthaeng, N.; Fisher, D.R.; Shukitt-Hale, B. Blueberry supplementation attenuates microglia activation and increases neuroplasticity in mice consuming a high-fat diet. Nutr. Neurosci. 2019, 22, 253–263. [Google Scholar] [CrossRef]
- Wang, S.; Okun, M.S.; Suslov, O.; Zheng, T.; McFarland, N.R.; Vedam-Mai, V.; Foote, K.D.; Roper, S.N.; Yachnis, A.T.; Siebzehnrubl, F.A.; et al. Neurogenic potential of progenitor cells isolated from postmortem human Parkinsonian brains. Brain Res. 2012, 1464, 61–72. [Google Scholar] [CrossRef]
- Scopa, C.; Marrocco, F.; Latina, V.; Ruggeri, F.; Corvaglia, V.; La Regina, F.; Ammassari-Teule, M.; Middei, S.; Amadoro, G.; Meli, G.; et al. Impaired adult neurogenesis is an early event in Alzheimer’s disease neurodegeneration, mediated by intracellular Abeta oligomers. Cell Death Differ. 2020, 27, 934–948. [Google Scholar] [CrossRef]
- Larson, T.A.; Thatra, N.M.; Lee, B.H.; Brenowitz, E.A. Reactive neurogenesis in response to naturally occurring apoptosis in an adult brain. J. Neurosci. 2014, 34, 13066–13076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, W.; Jiang, Y.-G.; Yang, H.-P.; Cheng, D.-M.; Lu, H.; Lu, S.-J. Cytoprotective effect of blueberry extracts against oxidative damage of rat hippocampal neurons induced by H2O2. Zhongguo Yingyong Shenglixue Zazhi = Chin. J. Appl. Physiol. 2010, 26, 51–54. [Google Scholar]
- Bickford, P.C.; Tan, J.; Shytle, R.D.; Sanberg, C.D.; El-Badri, N.; Sanberg, P.R. Nutraceuticals synergistically promote proliferation of human stem cells. Stem Cells Dev. 2006, 15, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Kuzumaki, N.; Ikegami, D.; Imai, S.; Narita, M.; Tamura, R.; Yajima, M.; Suzuki, A.; Miyashita, K.; Niikura, K.; Takeshima, H.; et al. Enhanced IL-1beta production in response to the activation of hippocampal glial cells impairs neurogenesis in aged mice. Synapse 2010, 64, 721–728. [Google Scholar] [PubMed]
- Spitzer, N. Calcium: First messenger. Nat. Neurosci. 2008, 11, 243–244. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.R.; Seo, J.T. Calcium overload is essential for the acceleration of staurosporine-induced cell death following neuronal differentiation in PC12 cells. Exp. Mol. Med. 2009, 41, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Shukitt-Hale, B. Blueberries and neuronal aging. Gerontology 2012, 58, 518–523. [Google Scholar] [CrossRef] [PubMed]
- Shukitt-Hale, B.; Lau, F.C.; Carey, A.N.; Galli, R.L.; Spangler, E.L.; Ingram, D.K.; Joseph, J.A. Blueberry polyphenols attenuate kainic acid-induced decrements in cognition and alter inflammatory gene expression in rat hippocampus. Nutr. Neurosci. 2008, 11, 172–182. [Google Scholar] [CrossRef]
- Kino, T. Stress, glucocorticoid hormones, and hippocampal neural progenitor cells: Implications to mood disorders. Front. Physiol. 2015, 6, 230. [Google Scholar] [CrossRef] [PubMed]
- de Miranda, A.S.; Zhang, C.J.; Katsumoto, A.; Teixeira, A.L. Hippocampal adult neurogenesis: Does the immune system matter? J. Neurol. Sci. 2017, 372, 482–495. [Google Scholar] [CrossRef]
- Cahoon, D.S.; Fisher, D.R.; Lamon-Fava, S.; Wu, D.; Zheng, T.; Shukitt-Hale, B. Blueberry treatment administered before and/or after lipopolysaccharide stimulation attenuates inflammation and oxidative stress in rat microglial cells. Nutr. Neurosci. 2022, 1–11. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, T.; Bielinski, D.F.; Fisher, D.R.; Zhang, J.; Shukitt-Hale, B. Protective Effects of a Polyphenol-Rich Blueberry Extract on Adult Human Neural Progenitor Cells. Molecules 2022, 27, 6152. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27196152
Zheng T, Bielinski DF, Fisher DR, Zhang J, Shukitt-Hale B. Protective Effects of a Polyphenol-Rich Blueberry Extract on Adult Human Neural Progenitor Cells. Molecules. 2022; 27(19):6152. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27196152
Chicago/Turabian StyleZheng, Tong, Donna F. Bielinski, Derek R. Fisher, Jianyi Zhang, and Barbara Shukitt-Hale. 2022. "Protective Effects of a Polyphenol-Rich Blueberry Extract on Adult Human Neural Progenitor Cells" Molecules 27, no. 19: 6152. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27196152