Commitment of Autologous Human Multipotent Stem Cells on Biomimetic Poly-L-Lactic Acid-Based Scaffolds Is Strongly Influenced by Structure and Concentration of Carbon Nanomaterial

 , ,
, ,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Nanofiller Feature, and Scaffold Preparation and Sterilization
2.2. SH-SY5Y, Primary Myoblast, and Adipocyte Culture
2.3. hCMC Isolation, Culture, and Differentiation
2.4. Morphological Analysis
2.5. RNA Extraction and qPCR
- -
- Leptin, NM_000230.2, A: 259bp,F: CCATAACAGCCAACAGGTG, R: CCTCTCGCTGTAACTCACTGC;
- -
- MAP2, NM_002374.3, A: 253bp,F: ATAGACCTAAGCCATGTG, R: GGGACTGTGTAATGATCTC;
- -
- MYOD1, NM_000230.2, A: 269bp,F: GAGGCGGGAGAACTGAAG, R: CTGCTACATTTGGGACCG;
- -
- MYOG, NM_002479.6, A: 259bp,F: GGACAGCATCACAGTGGAAG, R: GAATGAGGGCGTCCAGTC;
- -
- Nestin, NM_006617.1, A: 257bp,F: CAGGGGAGGACTAGGAAAAGA, R: GAGATGGAGCAGGCAAGAG;
- -
- Pax7, NM_001135254.2, A: 259bp,F: CTTGAGAACAGGACGGGTC, R: GTCTTGGTTTTGGTGCCTC;
- -
- Plin1, NM_002666.5, A: 242bp,F: CACAGCCACATTTCCATTTG, R: CAATGAAGGGGAACAGGG;
- -
- Ribosomal protein S13 (S13), NM_001017.2, A: 259bp,F: TACAAACTGGCCAAGAAGGG, R: GGTGAATCCGGCTCTCTATTAG;
- -
- TUBß3, NM_001197181, A: 259bp,F: AGGAAGAGGGCGAGATGTA, R: CAATAAGACAGAGACAGGAGCAG.
2.6. Immunofluorescence
2.7. Statistical Analysis
3. Results and Discussion
3.1. Scaffold Effect on Neuronal Differentiation
3.2. Selection of Non-Neuronal Markers
3.3. Scaffold Effect on Marker-Gene Modulation
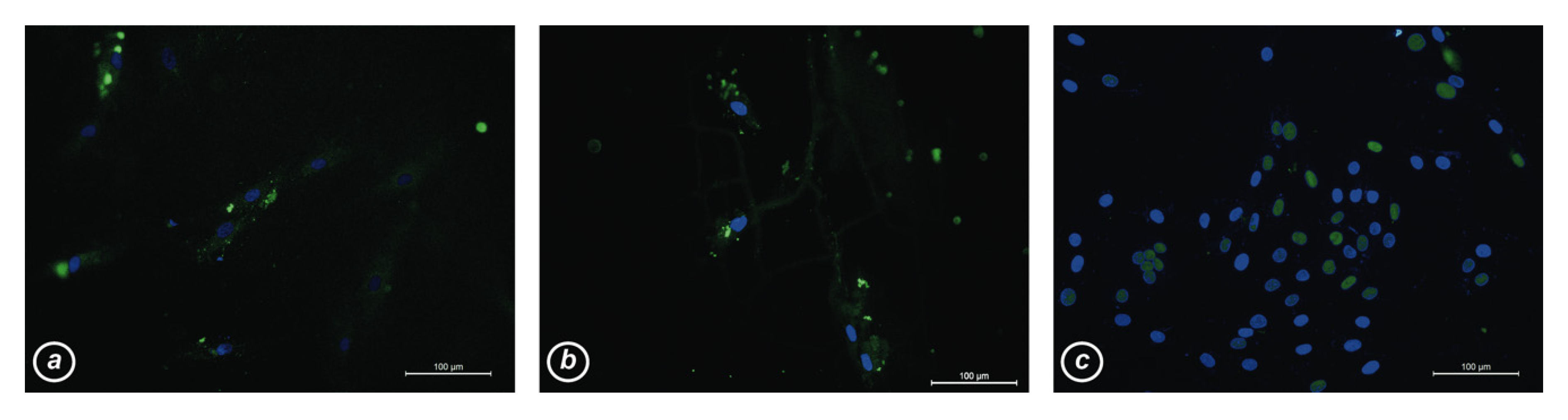
3.4. Production and Subcellular Localization of MyoD1 Protein in hCMCs and Control Myoblasts
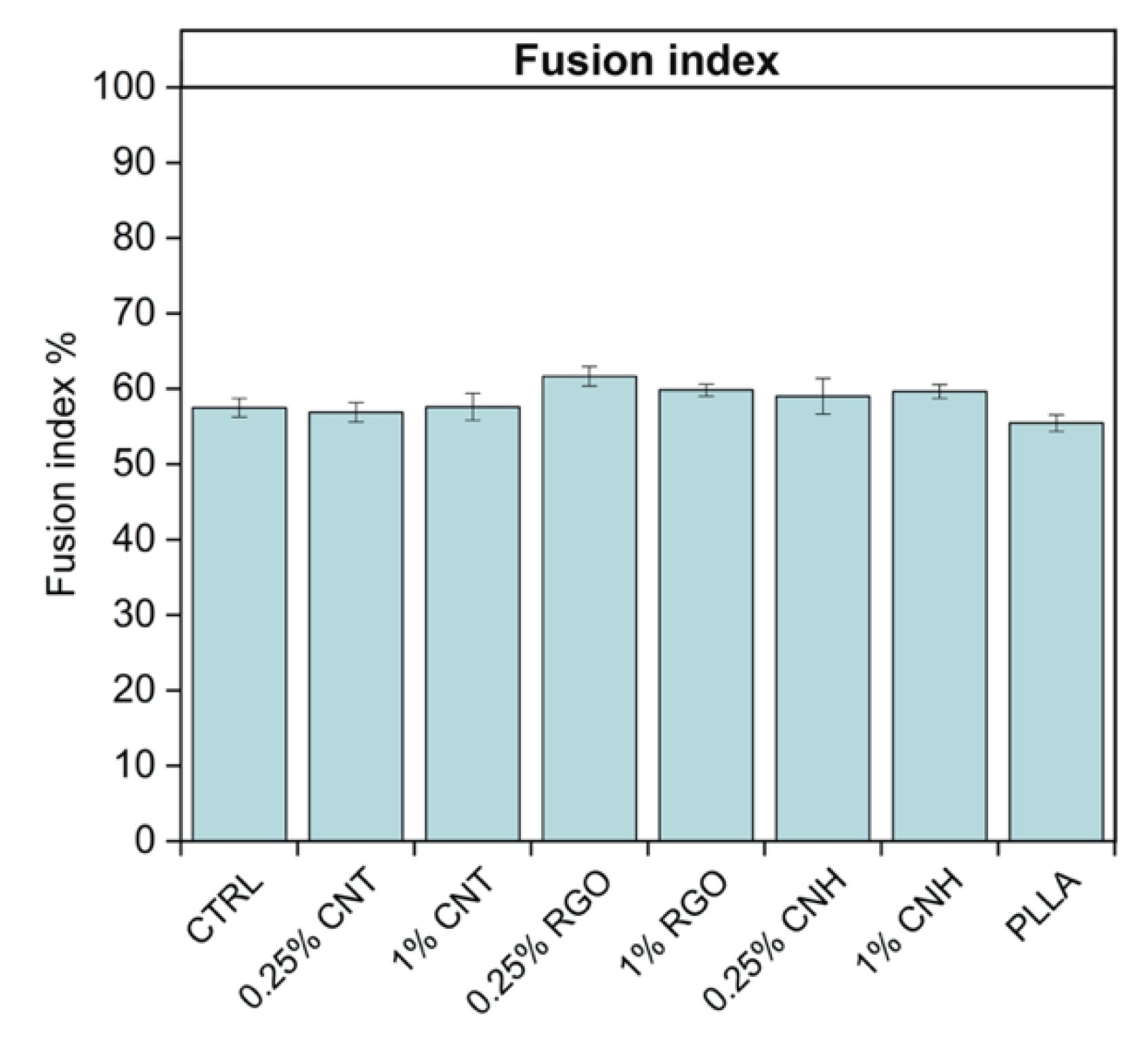
3.5. Effect of CNM@PLLA Scaffolds on Myoblast Differentiation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhou, Y.; Notterpek, L. Promoting peripheral myelin repair. Exp. Neurol. 2016, 283, 573–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, L.; Prabhakaran, M.P.; Ramakrishna, S. Strategies for regeneration of components of nervous system: Scaffolds, cells and biomolecules. Regen. Biomater. 2015, 2, 31–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiono, V.; Tonda-Turo, C. Trends in the design of nerve guidance channels in peripheral nerve tissue engineering. Prog. Neurobiol. 2015, 131, 87–104. [Google Scholar] [CrossRef] [PubMed]
- Gilbert-Honick, J.; Grayson, W. Vascularized and innervated skeletal muscle tissue engineering. Adv. Healthc. Mater. 2020, 9, e1900626. [Google Scholar] [CrossRef]
- Han, Y.; Li, X.; Zhang, Y.; Han, Y.; Chang, F.; Ding, J. Mesenchymal stem cells for regenerative medicine. Cells 2019, 8, 886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.; Lim, D.J.; Lee, S.H.; Park, H. Nanofibrous mineralized electrospun scaffold as a substrate for bone tissue regeneration. J. Biomed. Nanotechnol. 2016, 12, 2076–2082. [Google Scholar] [CrossRef]
- Park, K. Adipose-derived stem cells combined with neuregulin microparticles for efficient cardiac repair. J. Control Release 2017, 249, 196. [Google Scholar] [CrossRef]
- Scapin, G.; Bertalot, T.; Vicentini, N.; Gatti, T.; Tescari, S.; De Filippis, V.; Marega, C.; Menna, E.; Gasparella, M.; Parnigotto, P.P.; et al. Neuronal commitment of human circulating multipotent cells by carbon nanotube-polymer scaffolds and biomimetic peptides. Nanomedicine 2016, 11, 1929–1946. [Google Scholar] [CrossRef]
- Di Liddo, R.; Bertalot, T.; Borean, A.; Pirola, I.; Argentoni, A.; Schrenk, S.; Cenzi, C.; Capelli, S.; Conconi, M.T.; Parnigotto, P.P. Leucocyte and Platelet-rich Fibrin: A carrier of autologous multipotent cells for regenerative medicine. J. Cell Mol. Med. 2018, 22, 1840–1854. [Google Scholar] [CrossRef] [Green Version]
- Mao, A.S.; Shin, J.W.; Mooney, D.J. Effects of substrate stiffness and cell-cell contact on mesenchymal stem cell differentiation. Biomaterials 2016, 98, 184–191. [Google Scholar] [CrossRef] [Green Version]
- Saha, K.; Keung, A.J.; Irwin, E.F.; Li, Y.; Little, L.; Schaffer, D.V.; Healy, K.E. Substrate modulus directs neural stem cell behavior. Biophys. J. 2008, 95, 4426–4438. [Google Scholar] [CrossRef] [Green Version]
- Keung, A.J.; de Juan-Pardo, E.M.; Schaffer, D.V.; Kumar, S. Rho GTPases mediate the mechano-sensitive lineage commitment of neural stem cells. Stem. Cells 2011, 29, 1886–1897. [Google Scholar] [CrossRef] [PubMed]
- Jo, H.; Sim, M.; Kim, S.; Yang, S.; Yoo, Y.; Park, J.H.; Yoon, T.H.; Kim, M.G.; Lee, J.Y. Electrically conductive graphene/polyacrylamide hydrogels produced by mild chemical reduction for enhanced myoblast growth and differentiation. Acta Biomater. 2017, 48, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Shao, H.; Li, T.; Zhu, R.; Xu, X.; Yu, J.; Chen, S.; Song, L.; Ramakrishna, S.; Lei, Z.; Ruan, Y.; et al. Carbon nanotube multilayered nanocomposites as multifunctional substrates for actuating neuronal differentiation and functions of neural stem cells. Biomaterials 2018, 175, 93–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lovat, V.; Pantarotto, D.; Lagostena, L.; Cacciari, B.; Grandolfo, M.; Righi, M.; Spalluto, G.; Prato, M.; Ballerini, L. Carbon nanotube substrates boost neuronal electrical signaling. Nano Lett. 2005, 5, 1107–1110. [Google Scholar] [CrossRef] [PubMed]
- Sucapane, A.; Cellot, G.; Prato, M.; Giugliano, M.; Parpura, V.; Ballerini, L. Interactions between cultured neurons and carbon nanotubes: A nanoneuroscience vignette. J. Nanoneurosci. 2009, 1, 10–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cellot, G.; Cilia, E.; Cipollone, S.; Rancic, V.; Sucapane, A.; Giordani, S.; Gambazzi, L.; Markram, H.; Grandolfo, M.; Scaini, D.; et al. Carbon nanotubes might improve neuronal performance by favouring electrical shortcuts. Nat. Nanotechnol. 2009, 4, 126–133. [Google Scholar] [CrossRef]
- Oprych, K.M.; Whitby, R.L.; Mikhalovsky, S.V.; Tomlins, P.; Adu, J. Repairing peripheral nerves: is there a role for carbon nanotubes? Adv. Healthc. Mater. 2016, 5, 1253–1271. [Google Scholar] [CrossRef] [Green Version]
- Fabbro, A.; Prato, M.; Ballerini, L. Carbon nanotubes in neuroregeneration and repair. Adv. Drug. Deliv. Rev. 2013, 65, 2034–2044. [Google Scholar] [CrossRef]
- Fabbro, A.; Bosi, S.; Ballerini, L.; Prato, M. Carbon nanotubes: Artificial nanomaterials to engineer single neurons and neuronal networks. ACS Chem. Neurosci. 2012, 3, 611–618. [Google Scholar] [CrossRef] [Green Version]
- Fattahi, P.; Yang, G.; Kim, G.; Abidian, M.R. A review of organic and inorganic biomaterials for neural interfaces. Adv. Mater. 2014, 26, 1846–1885. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhao, T.; Sun, L.; Aifantis, K.E.; Fan, Y.; Feng, Q.; Cui, F.; Watari, F. The applications of conductive nanomaterials in the biomedical field. J. Biomed. Mater. Res. Part A 2016, 104, 322–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotov, N.A.; Winter, J.O.; Clements, I.P.; Jan, E.; Timko, B.P.; Campidelli, S.; Pathak, S.; Mazzatenta, A.; Lieber, C.M.; Prato, M.; et al. Nanomaterials for neural interfaces. Adv. Mater. 2009, 21, 3970–4004. [Google Scholar] [CrossRef] [Green Version]
- Ku, S.H.; Lee, M.; Park, C.B. Carbon-based nanomaterials for tissue engineering. Adv. Healthc. Mater. 2013, 2, 244–260. [Google Scholar] [CrossRef] [PubMed]
- Akhavan, O. Graphene scaffolds in progressive nanotechnology/stem cell-based tissue engineering of the nervous system. J. Mater. Cem. B 2016, 4, 3169–3190. [Google Scholar] [CrossRef]
- Qian, Y.; Zhao, X.; Han, Q.; Chen, W.; Li, H.; Yuan, W. An integrated multi-layer 3D-fabrication of PDA/RGD coated graphene loaded PCL nanoscaffold for peripheral nerve restoration. Nat. Commun. 2018, 9, 323. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Cheng, Y.; Chen, L.; Zhu, T.; Ye, K.; Jia, C.; Wang, H.; Zhu, M.; Fan, C.; Mo, X. In vitro and in vivo studies of electroactive reduced graphene oxide-modified nanofiber scaffolds for peripheral nerve regeneration. Acta Biomater. 2019, 84, 98–113. [Google Scholar] [CrossRef]
- Gardin, C.; Piattelli, A.; Zavan, B. Graphene in regenerative medicine: Focus on stem cells and neuronal differentiation. Trends Biotechnol. 2016, 34, 435–437. [Google Scholar] [CrossRef]
- Ma, Y.; Shen, H.; Tu, X.; Zhang, Z. Assessing in vivo toxicity of graphene materials: Current methods and future outlook. Nanomedicine 2014, 9, 1565–1580. [Google Scholar] [CrossRef]
- Lanone, S.; Andujar, P.; Kermanizadeh, A.; Boczkowski, J. Determinants of carbon nanotube toxicity. Adv. Drug. Deliv. Rev. 2013, 65, 2063–2069. [Google Scholar] [CrossRef]
- Dong, J.; Ma, Q. Advances in mechanisms and signaling pathways of carbon nanotube toxicity. Nanotoxicology 2015, 9, 658–676. [Google Scholar] [CrossRef] [Green Version]
- Migliore, L.; Uboldi, C.; Di Bucchianico, S.; Coppedè, F. Nanomaterials and neurodegeneration. Environ. Mol. Mutagen. 2015, 56, 149–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilbasmis-Tamer, S.; Ciftci, H.; Turk, M.; Degim, T.; Tamer, U. Multiwalled carbon nanotube-chitosan scaffold: Cytotoxic, apoptotic, and necrotic effects on chondrocyte cell lines. Curr. Pharm. Biotechnol. 2017, 18, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Yang, M.; Bussy, C.; Iijima, S.; Kostarelos, K.; Yudasaka, M. Biodegradation of carbon nanohorns in macrophage cells. Nanoscale 2015, 7, 2834–2840. [Google Scholar] [CrossRef] [PubMed]
- Salice, P.; Fabris, E.; Sartorio, C.; Fenaroli, D.; Figà, V.; Casaletto, M.P.; Cataldo, S.; Pignataro, B.; Menna, E. An insight into the functionalisation of carbon nanotubes by diazonium chemistry: Towards a controlled decoration. Carbon 2014, 74, 73–82. [Google Scholar] [CrossRef]
- Liu, J.; Appaix, F.; Bibari, O.; Marchand, G.; Benabid, A.L.; Sauter-Starace, F.; De Waard, M. Control of neuronal network organization by chemical surface functionalization of multi-walled carbon nanotube arrays. Nanotechnology 2011, 22, 195101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, H.; Ni, Y.; Montana, V.; Haddon, R.C.; Parpura, V. Chemically functionalized carbon nanotubes as substrates for neuronal growth. Nano Lett. 2004, 4, 507–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatti, T.; Vicentini, N.; Mba, M.; Menna, E. Organic functionalized carbon nanostructures for functional polymer-based nanocomposites. Eur. J. Org. Chem. 2016, 6, 1071–1090. [Google Scholar] [CrossRef]
- Goode, A.E.; Gonzalez Carter, D.A.; Motskin, M.; Pienaar, I.S.; Chen, S.; Hu, S.; Ruenraroengsak, P.; Ryan, M.P.; Shaffer, M.S.; Dexter, D.T.; et al. High resolution and dynamic imaging of biopersistence and bioreactivity of extra and intracellular MWNTs exposed to microglial cells. Biomaterials 2015, 70, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Haniu, H.; Saito, N.; Matsuda, Y.; Tsukahara, T.; Usui, Y.; Maruyama, K.; Takanashi, S.; Aoki, K.; Kobayashi, S.; Nomura, H.; et al. Biological responses according to the shape and size of carbon nanotubes in BEAS-2B and MESO-1 cells. Int. J. Nanomed. 2014, 17, 1979–1990. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Romain, C.; Williams, C.K. Sustainable polymers from renewable resources. Nature 2016, 540, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Irimia-Vladu, M. Green electronics: Biodegradable and biocompatible materials and devices for sustainable future. Chem. Soc. Rev. 2014, 43, 588–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nofar, M.; Sacligil, D.; Carreau, P.J.; Kamal, M.R.; Heuzey, M.C. Poly (lactic acid) blends: Processing, properties and applications. Int. J. Biol. Macromol. 2019, 125, 307–360. [Google Scholar] [CrossRef] [PubMed]
- Andreeva, T.D.; Stoichev, S.; Taneva, S.G.; Krastev, R. Hybrid graphene oxide/polysaccharide nanocomposites with controllable surface properties and biocompatibility. Carbohydr. Polym. 2018, 181, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Kingston, C.; Zepp, R.; Andrady, A.; Boverhof, D.; Fehir, R.; Hawkins, D.; Roberts, J.; Sayre, P.; Shelton, B.; Sultan, Y.; et al. Release characteristics of selected carbon nanotube polymer composites. Carbon 2014, 68, 33–57. [Google Scholar] [CrossRef]
- Weng, B.; Diao, J.; Xu, Q.; Liu, Y.; Li, C.; Ding, A.; Chen, J. Bio-interface of Conducting polymer-based materials for neuroregeneration. Adv. Mater. Interfaces 2015, 2, 1500059-1–1500059-23. [Google Scholar] [CrossRef] [Green Version]
- Ventre, M.; Causa, F.; Netti, P.A. Determinants of cell-material crosstalk at the interface: Towards engineering of cell instructive materials. J. R. Soc. Interface 2012, 9, 2017–2032. [Google Scholar] [CrossRef]
- Someya, T.; Bao, Z.; Malliaras, G.G. The rise of plastic bioelectronics. Nature. 2016, 540, 379–385. [Google Scholar] [CrossRef]
- Liao, C.; Zhang, M.; Yao, M.Y.; Hua, T.; Li, L.; Yan, F. Flexible organic electronics in biology: Materials and devices. Adv. Mater. 2015, 27, 7493–7527. [Google Scholar] [CrossRef]
- Scapin, G.; Salice, P.; Tescari, S.; Menna, E.; De Filippis, V.; Filippini, F. Enhanced neuronal cell differentiation combining biomimetic peptides and a carbon nanotube-polymer scaffold. Nanomedicine 2015, 11, 621–632. [Google Scholar] [CrossRef]
- Vicentini, N.; Gatti, T.; Salice, P.; Scapin, G.; Marega, C.; Filippini, F.; Menna, E. Covalent functionalization enables good dispersion and anisotropic orientation of multi-walled carbon nanotubes in a poly(L-lactic acid) electrospun nanofibrous matrix boosting neuronal differentiation. Carbon 2015, 95, 725–730. [Google Scholar] [CrossRef]
- Vicentini, N.; Gatti, T.; Salerno, M.; Hernandez Gomez, Y.S.; Bellon, M.; Gallio, S.; Marega, C.; Filippini, F.; Menna, E. Effect of different functionalized carbon nanostructures as fillers on the physical properties of biocompatible poly(L-lactic acid) composites. Mater. Chem. Phys. 2018, 214, 265–276. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vacca, M.; Albania, L.; Della Ragione, F.; Carpi, A.; Rossi, V.; Strazzullo, M.; De Franceschi, N.; Rossetto, O.; Filippini, F.; D’Esposito, M. Alternative splicing of the human gene SYBL1 modulates protein domain architecture of Longin VAMP7/TI-VAMP, showing both non-SNARE and synaptobrevin-like isoforms. BMC Mol. Biol. 2011, 12, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernal, A.; Arranz, L. Nestin-expressing progenitor cells: Function, identity and therapeutic implications. Cell Mol. Life Sci. 2018, 75, 2177–2195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Person, F.; Wilczak, W.; Hube-Magg, C.; Burdelski, C.; Möller-Koop, C.; Simon, R.; Noriega, M.; Sauter, G.; Steurer, S.; Burdak-Rothkamm, S.; et al. Prevalence of βIII-tubulin (TUBB3) expression in human normal tissues and cancers. Tumour. Biol. 2017, 39, 1010428317712166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardwick, L.J.; Davies, J.D.; Philpott, A. MyoD phosphorylation on multiple C terminal sites regulates myogenic conversion activity. Biochem. Biophys. Res. Commun. 2016, 481, 97–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tonellato, M.; Piccione, M.; Gasparotto, M.; Bellet, P.; Tibaudo, L.; Vicentini, N.; Bergantino, E.; Menna, E.; Vitiello, L.; Di Liddo, R.; et al. Commitment of Autologous Human Multipotent Stem Cells on Biomimetic Poly-L-Lactic Acid-Based Scaffolds Is Strongly Influenced by Structure and Concentration of Carbon Nanomaterial. Nanomaterials 2020, 10, 415. https://0-doi-org.brum.beds.ac.uk/10.3390/nano10030415
Tonellato M, Piccione M, Gasparotto M, Bellet P, Tibaudo L, Vicentini N, Bergantino E, Menna E, Vitiello L, Di Liddo R, et al. Commitment of Autologous Human Multipotent Stem Cells on Biomimetic Poly-L-Lactic Acid-Based Scaffolds Is Strongly Influenced by Structure and Concentration of Carbon Nanomaterial. Nanomaterials. 2020; 10(3):415. https://0-doi-org.brum.beds.ac.uk/10.3390/nano10030415
Chicago/Turabian StyleTonellato, Marika, Monica Piccione, Matteo Gasparotto, Pietro Bellet, Lucia Tibaudo, Nicola Vicentini, Elisabetta Bergantino, Enzo Menna, Libero Vitiello, Rosa Di Liddo, and et al. 2020. "Commitment of Autologous Human Multipotent Stem Cells on Biomimetic Poly-L-Lactic Acid-Based Scaffolds Is Strongly Influenced by Structure and Concentration of Carbon Nanomaterial" Nanomaterials 10, no. 3: 415. https://0-doi-org.brum.beds.ac.uk/10.3390/nano10030415