Reconfiguring Nature’s Cholesterol Accepting Lipoproteins as Nanoparticle Platforms for Transport and Delivery of Therapeutic and Imaging Agents

Abstract
:1. Introduction and Historical Perspective
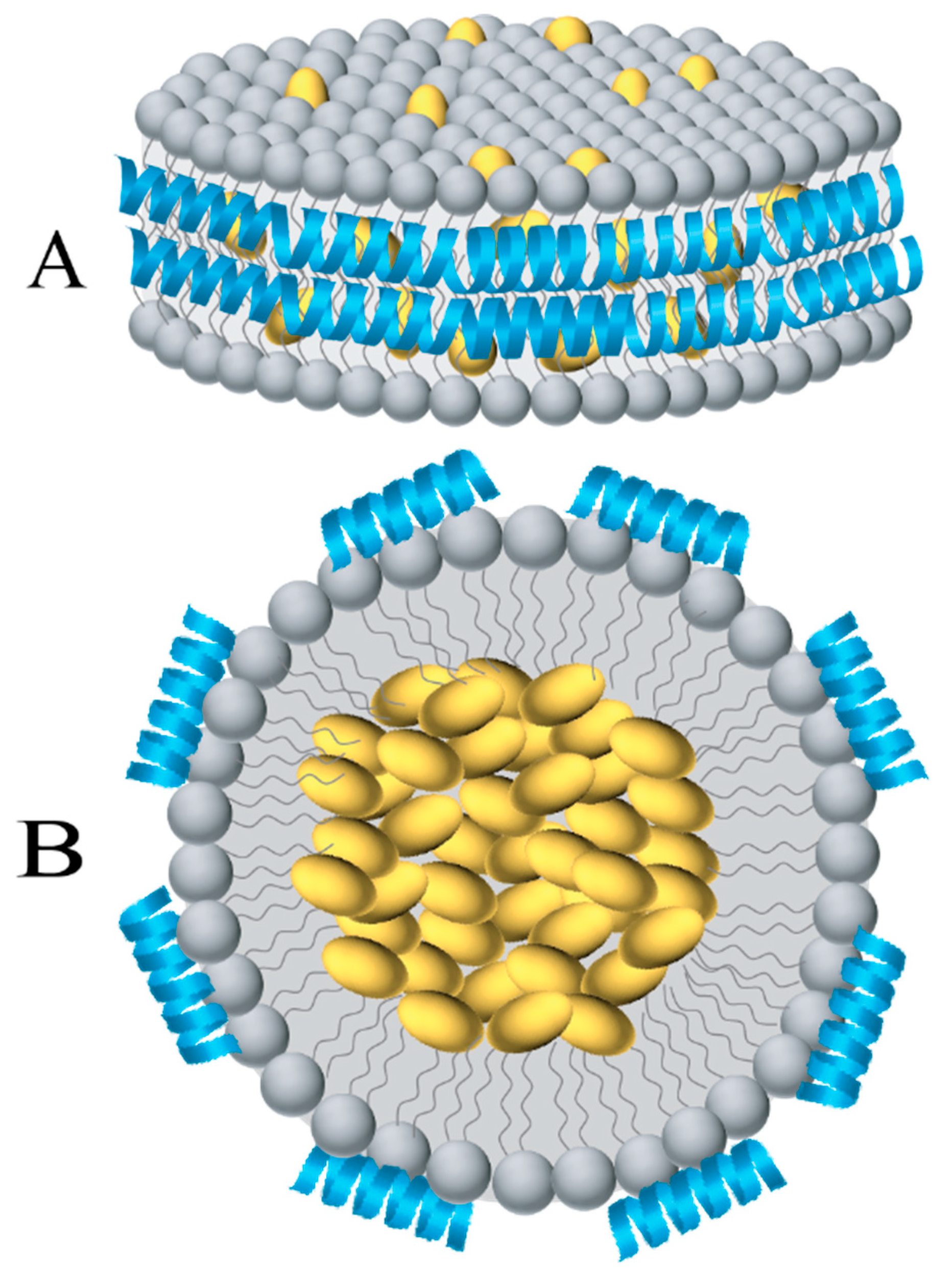
2. Structural Organization of the Nanodisc Platform
2.1. Structural Analysis of Membrane Proteins
2.2. Acceptors of Cholesterol Efflux for Improved Cardiovascular Outcomes
3. The Protein Scaffold of Apolipoprotein-Based Nanostructures
3.1. Repurposed LDL and its Derivatives
3.2. ApoAI and ApoE3
3.3. ApoJ
3.4. Human Serum Albumin Coated with ApoAI or ApoE3
3.5. Peptide Mimetics
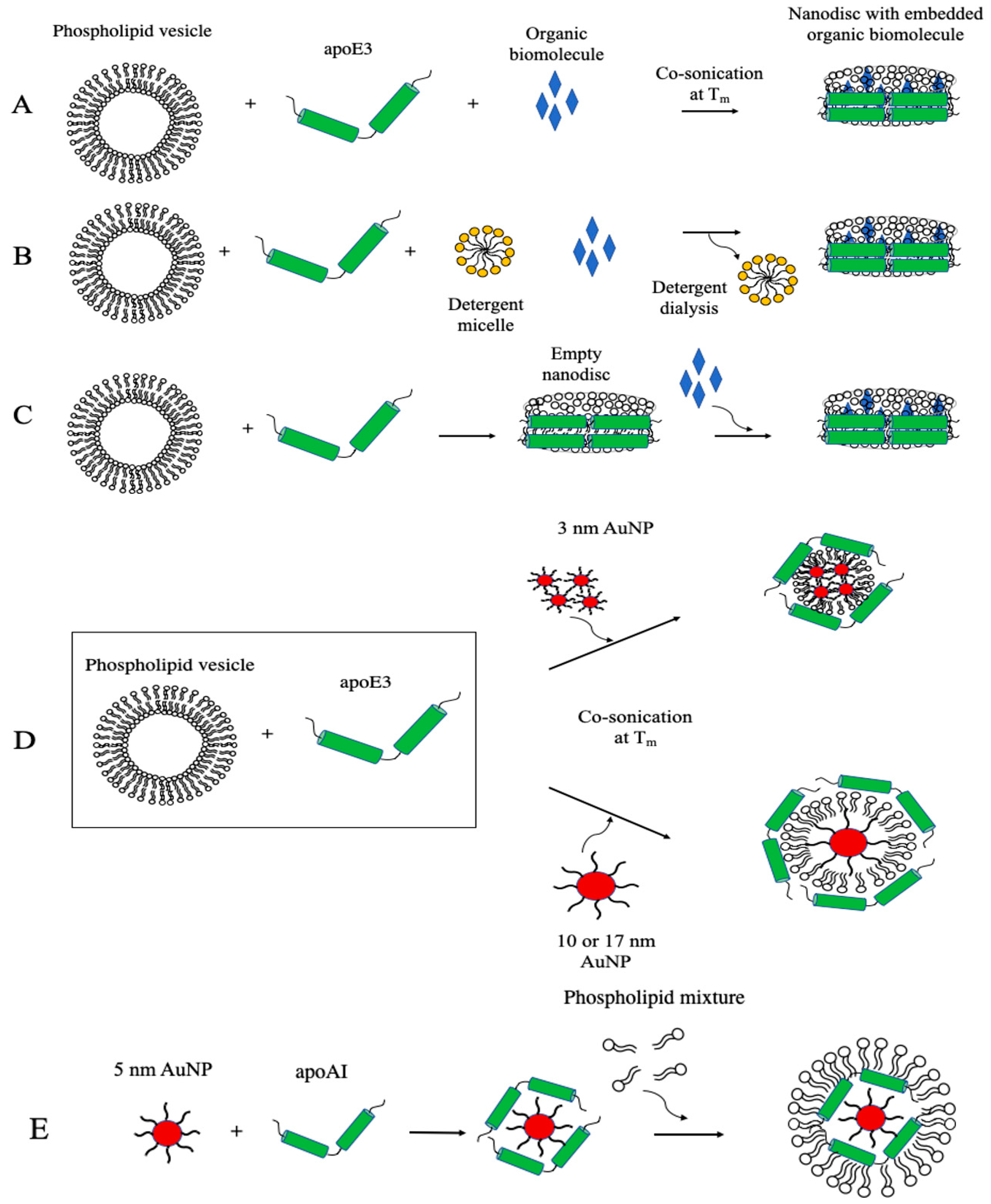
4. Mechanism of Assembly of the Lipid Platform of Apolipoprotein-Based Nanostructures
5. Synthesis of Apolipoprotein-Based Nanoparticles
6. Organic Biomolecules Payload in Apolipoprotein-Based Nanostructures
6.1. Antioxidant, Anti-Inflammatory, and Anti-Atherogenic Agents
6.2. Nucleic Acid Agents
6.3. Anti-Cancer and Antimicrobial Therapeutics
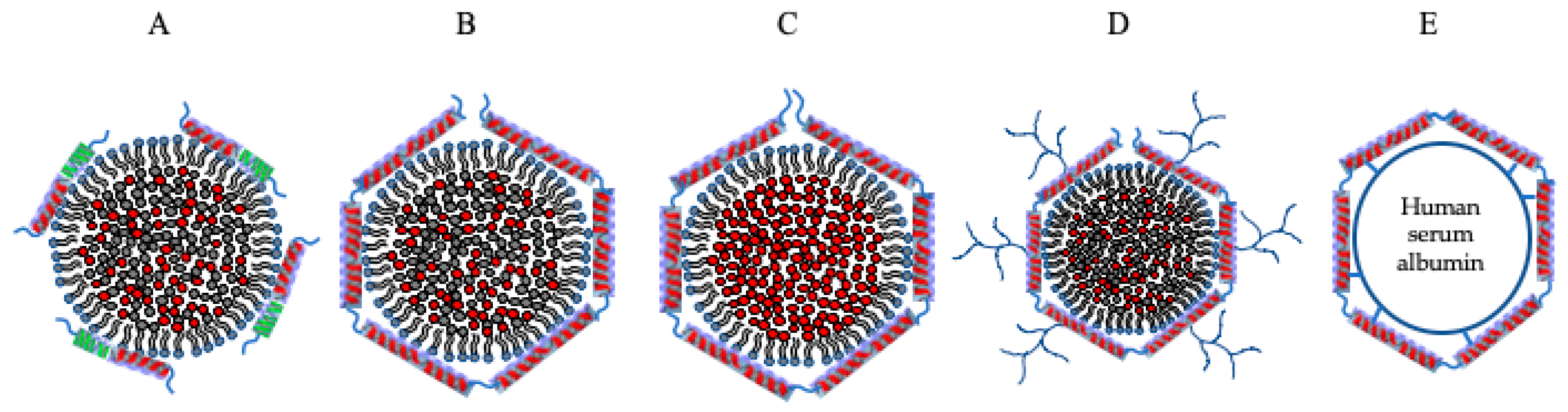
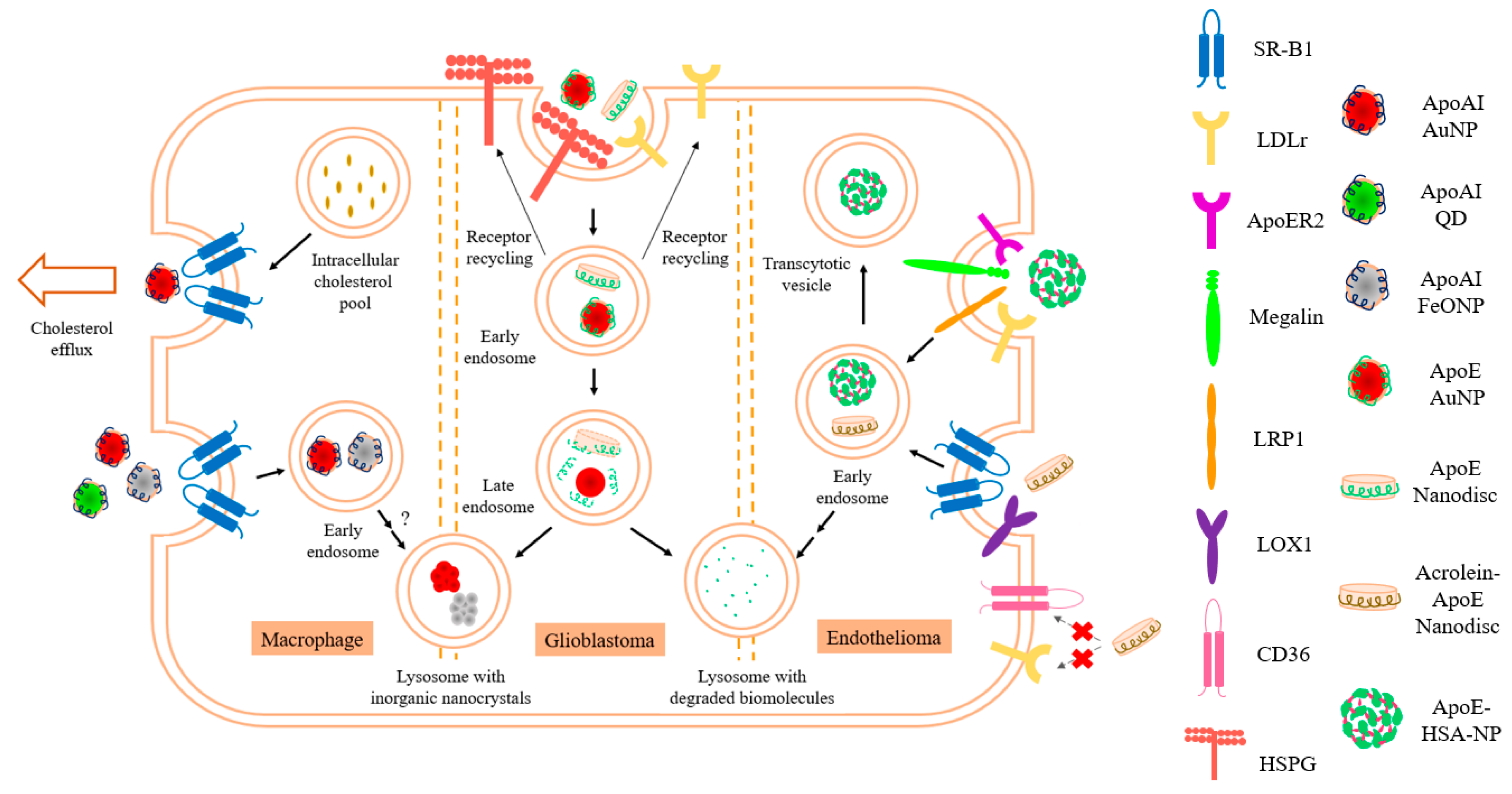
7. Metal Core in Apolipoprotein- Based Nanostructures for Imaging, Diagnostics, and Therapeutics
7.1. Metal Core in Nanolipoproteins as Diagnostic Tool
7.2. Metal Core in Nanolipoproteins as Therapeutic Agents
8. Summary and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Counsell, R.E.; Pohland, R.C. Lipoproteins as potential site-specific delivery systems for diagnostic and therapeutic agents. J. Med. Chem. 1982, 25, 1115–1120. [Google Scholar] [CrossRef] [PubMed]
- Firestone, R.A. Low-density lipoprotein as a vehicle for targeting antitumor compounds to cancer cells. Bioconjug. Chem. 1994, 5, 105–113. [Google Scholar] [CrossRef]
- Segrest, J.P.; Jones, M.K.; De Loof, H.; Brouillette, C.G.; Venkatachalapathi, Y.V.; Anantharamaiah, G.M. The amphipathic helix in the exchangeable apolipoproteins: A review of secondary structure and function. J. Lipid Res. 1992, 33, 141–166. [Google Scholar]
- Raussens, V.; Narayanaswami, V.; Goormaghtigh, E.; Ryan, R.O.; Ruysschaert, J.M. Alignment of the apolipophorin-III α-helices in complex with dimyristoylphosphatidylcholine: A unique spatial orientation. J. Biol. Chem. 1995, 270, 12542–12547. [Google Scholar] [CrossRef] [Green Version]
- Wientzek, M.; Kay, C.M.; Oikawa, K.; Ryan, R.O. Binding of insect apolipophorin III to dimyristoylphosphatidylcholine vesicles. Evidence for a conformational change. J. Biol. Chem. 1994, 269, 4605–4612. [Google Scholar]
- Brasseur, R.; De Meutter, J.; Vanloo, B.; Goormaghtigh, E.; Ruysschaert, J.M.; Rosseneu, M. Mode of assembly of amphipathic helical segments in model high-density lipoproteins. Biochim. Biophys. Acta BBA Lipids Lipid Metab. 1990, 1043, 245–252. [Google Scholar] [CrossRef]
- Wald, J.H.; Goormaghtigh, E.; De Meutter, J.; Ruysschaert, J.M.; Jonas, A. Investigation of the lipid domains and apolipoprotein orientation in reconstituted high density lipoproteins by fluorescence and IR methods. J. Biol. Chem. 1990, 265, 20044–20050. [Google Scholar]
- Wald, J.H.; Krul, E.S.; Jonas, A. Structure of apolipoprotein A-I in three homogeneous, reconstituted high density lipoprotein particles. J. Biol. Chem. 1990, 265, 20037–20043. [Google Scholar] [PubMed]
- De Pauw, M.; Vanloo, B.; Weisgraber, K.; Rosseneu, M. Comparison of lipid-binding and lecithin: Cholesterol acyltransferase activation of the amino- and carboxyl-terminal domains of human apolipoprotein E3. Biochemistry 1995, 34, 10953–10960. [Google Scholar] [CrossRef] [PubMed]
- Islam, R.M.; Pourmousa, M.; Sviridov, D.; Gordon, S.M.; Neufeld, E.B.; Freeman, L.A.; Perrin, B.S.; Pastor, R.W.; Remaley, A.T. Structural properties of apolipoprotein A-I mimetic peptides that promote ABCA1-dependent cholesterol efflux. Sci. Rep. 2018, 8, 2956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raussens, V.; Drury, J.; Forte, T.M.; Choy, N.; Goormaghtigh, E.; Ruysschaert, J.M.; Narayanaswami, V. Orientation and mode of lipid-binding interaction of human apolipoprotein E C-terminal domain. Biochem. J. 2005, 387, 747–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raussens, V.; Fisher, C.A.; Goormaghtigh, E.; Ryan, R.O.; Ruysschaert, J.M. The low density lipoprotein receptor active conformation of apolipoprotein E: Helix organization in N-terminal domain-phospholipid disc particles. J. Biol. Chem. 1998, 273, 25825–25830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koppaka, V.; Silvestro, L.; Engler, J.A.; Brouillette, C.G.; Axelsen, P.H. The structure of human lipoprotein A-I: Evidence for the “belt” model. J. Biol. Chem. 1999, 274, 14541–14544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidson, W.S.; D Silva, R.G. Apolipoprotein structural organization in high density lipoproteins: Belts, bundles, hinges and hairpins. Curr. Opin. Lipidol. 2005, 16, 295–300. [Google Scholar] [CrossRef]
- Borhani, D.W.; Rogers, D.P.; Engler, J.A.; Brouillette, C.G. Crystal structure of truncated human apolipoprotein A-I suggests a lipid-bound conformation. Proc. Natl. Acad. Sci. USA 1997, 94, 12291–12296. [Google Scholar] [CrossRef] [Green Version]
- Chetty, P.S.; Nguyen, D.; Nickel, M.; Lund-Katz, S.; Mayne, L.; Englander, S.W.; Phillips, M.C. Comparison of apoA-I helical structure and stability in discoidal and spherical HDL particles by HX and mass spectrometry. J. Lipid Res. 2013, 54, 1589–1597. [Google Scholar] [CrossRef] [Green Version]
- Chetty, P.S.; Mayne, L.; Kan, Z.-Y.; Lund-Katz, S.; Englander, S.W.; Phillips, M.C. Apolipoprotein A-I helical structure and stability in discoidal high-density lipoprotein (HDL) particles by hydrogen exchange and mass spectrometry. Proc. Natl. Acad. Sci. USA 2012, 109, 11687–11692. [Google Scholar] [CrossRef] [Green Version]
- Khumsupan, P.; Ramirez, R.; Khumsupan, D.; Narayanaswami, V. Apolipoprotein E LDL receptor-binding domain-containing high-density lipoprotein: A nanovehicle to transport curcumin, an antioxidant and anti-amyloid bioflavonoid. Biochim. Biophys. Acta BBA Biomembr. 2011, 1808, 352–359. [Google Scholar] [CrossRef] [Green Version]
- Borch, J.; Hamann, T. The nanodisc: A novel tool for membrane protein studies. Biol. Chem. 2009, 390, 805–814. [Google Scholar] [CrossRef]
- Viegas, A.; Viennet, T.; Etzkorn, M. The power, pitfalls and potential of the nanodisc system for NMR-based studies. Biol. Chem. 2016, 397, 1335–1354. [Google Scholar] [CrossRef]
- Denisov, I.G.; Sligar, S.G. Nanodiscs in Membrane Biochemistry and Biophysics. Chem. Rev. 2017, 117, 4669–4713. [Google Scholar] [CrossRef] [PubMed]
- McLean, M.A.; Gregory, M.C.; Sligar, S.G. Nanodiscs: A Controlled Bilayer Surface for the Study of Membrane Proteins. Annu. Rev. Biophys. 2018, 47, 107–124. [Google Scholar] [CrossRef]
- Ritchie, T.K.; Grinkova, Y.V.; Bayburt, T.H.; Denisov, I.G.; Zolnerciks, J.K.; Atkins, W.M.; Sligar, S.G. Chapter 11—Reconstitution of membrane proteins in phospholipid bilayer nanodiscs. Methods Enzymol. 2009, 464, 211–231. [Google Scholar] [PubMed] [Green Version]
- Bayburt, T.H.; Sligar, S.G. Membrane protein assembly into Nanodiscs. FEBS Lett. 2010, 584, 1721–1727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denisov, I.G.; Sligar, S.G. Nanodiscs for structural and functional studies of membrane proteins. Nat. Struct. Mol. Biol. 2016, 23, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Tsujita, M.; Wolska, A.; Gutmann, D.A.P.; Remaley, A.T. Reconstituted discoidal high-density lipoproteins: Bioinspired nanodiscs with many unexpected applications. Curr. Atheroscler. Rep. 2018, 20, 59. [Google Scholar] [CrossRef] [PubMed]
- Brouillette, C.G.; Anantharamaiah, G.M.; Engler, J.A.; Borhani, D.W. Structural models of human apolipoprotein A-I: A critical analysis and review. Biochim. Biophys. Acta 2001, 1531, 4–46. [Google Scholar] [CrossRef]
- Rothblat, G.H.; Phillips, M.C. High-density lipoprotein heterogeneity and function in reverse cholesterol transport. Curr. Opin. Lipidol. 2010, 21, 229–238. [Google Scholar] [CrossRef]
- Chi, G. High-density lipoprotein-targeted therapy for coronary heart disease: Is the HDL hypothesis operational or defunct? Curr. Trends Biomed. Eng. Biosci. 2018, 12, 555836. [Google Scholar] [CrossRef]
- Tardy, C.; Goffinet, M.; Boubekeur, N.; Ackermann, R.; Sy, G.; Bluteau, A.; Cholez, G.; Keyserling, C.; Lalwani, N.; Paolini, J.F.; et al. CER-001, a HDL-mimetic, stimulates the reverse lipid transport and atherosclerosis regression in high cholesterol diet-fed LDL-receptor deficient mice. Atherosclerosis 2014, 232, 110–118. [Google Scholar] [CrossRef]
- Nanjee, M.N.; Cooke, C.J.; Garvin, R.; Semeria, F.; Lewis, G.; Olszewski, W.L.; Miller, N.E. Intravenous apoA-I/lecithin discs increase pre-beta-HDL concentration in tissue fluid and stimulate reverse cholesterol transport in humans. J. Lipid Res. 2001, 42, 1586–1593. [Google Scholar] [PubMed]
- Spieker, L.E.; Sudano, I.; Hürlimann, D.; Lerch, P.G.; Lang, M.G.; Binggeli, C.; Corti, R.; Ruschitzka, F.; Lüscher, T.F.; Noll, G. High-density lipoprotein restores endothelial function in hypercholesterolemic men. Circulation 2002, 105, 1399–1402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tardif, J.C.; Gregoire, J.; L’Allier, P.L.; Ibrahim, R.; Lesperance, J.; Heinonen, T.M.; Kouz, S.; Berry, C.; Basser, R.; Lavoie, M.A.; et al. Effects of reconstituted high-density lipoprotein infusions on coronary atherosclerosis: A randomized controlled trial. JAMA 2007, 297, 1675–1682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tricoci, P.; D’Andrea, D.M.; Gurbel, P.A.; Yao, Z.; Cuchel, M.; Winston, B.; Schott, R.; Weiss, R.; Blazing, M.A.; Cannon, L.; et al. Infusion of reconstituted high-density lipoprotein, CSL112, in patients with atherosclerosis: Safety and pharmacokinetic results from a phase 2a randomized clinical trial. J. Am. Heart Assoc. 2015, 4, e002171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michael Gibson, C.; Korjian, S.; Tricoci, P.; Daaboul, Y.; Yee, M.; Jain, P.; Alexander, J.H.; Steg, P.G.; Lincoff, A.M.; Kastelein, J.J.P.; et al. Safety and tolerability of CSL112, a reconstituted, infusible, plasma-derived apolipoprotein A-I, after acute myocardial infarction: The AEGIS-I trial (ApoA-I Event Reducing in Ischemic Syndromes I). Circulation 2016, 134, 1918–1930. [Google Scholar] [CrossRef]
- Reijers, J.A.A.; Kallend, D.G.; Malone, K.E.; Jukema, J.W.; Wijngaard, P.L.J.; Burggraaf, J.; Moerland, M. MDCO-216 does not induce adverse immunostimulation, in contrast to its predecessor ETC-216. Cardiovasc. Drugs Ther. 2017, 31, 381–389. [Google Scholar] [CrossRef] [Green Version]
- Cho, K.H.; Kim, J.R. A reconstituted HDL containing V156K or R173C apoA-I exhibited anti-inflammatory activity in apo-E deficient mice and showed resistance to myeloperoxidase-mediated oxidation. Exp. Mol. Med. 2009, 41, 417–428. [Google Scholar] [CrossRef] [Green Version]
- Annema, W.; von Eckardstein, A. High-density lipoproteins. Multifunctional but vulnerable protections from atherosclerosis. Circ. J. Off. J. Jpn. Circ. Soc. 2013, 77, 2432–2448. [Google Scholar]
- Ghosh, M.; Ryan, R.O. ApoE enhances nanodisk-mediated curcumin delivery to glioblastoma multiforme cells. Nanomedicine 2014, 9, 763–771. [Google Scholar] [CrossRef] [Green Version]
- McConathy, W.J.; Nair, M.P.; Paranjape, S.; Mooberry, L.; Lacko, A.G. Evaluation of synthetic/reconstituted high-density lipoproteins as delivery vehicles for paclitaxel. Anticancer Drugs 2008, 19, 183–188. [Google Scholar] [CrossRef]
- Mooberry, L.K.; Nair, M.; Paranjape, S.; McConathy, W.J.; Lacko, A.G. Receptor mediated uptake of paclitaxel from a synthetic high density lipoprotein nanocarrier. J. Drug Target. 2010, 18, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.-Y.; Yang, Y.; Heo, P.; Lee, J.-C.; Kong, B.; Cho, J.Y.; Yoon, K.; Shin, C.-S.; Seo, J.-H.; Kim, S.-G.; et al. pH-responsive high-density lipoprotein-like nanoparticles to release paclitaxel at acidic pH in cancer chemotherapy. Int. J. Nanomed. 2012, 7, 2805–2816. [Google Scholar]
- Yuan, Y.; Wang, W.; Wang, B.; Zhu, H.; Zhang, B.; Feng, M. Delivery of hydrophilic drug doxorubicin hydrochloride-targeted liver using apoAI as carrier. J. Drug Target. 2013, 21, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Chen, B. Recombinant high density lipoprotein reconstituted with apolipoprotein AI cysteine mutants as delivery vehicles for 10-hydroxycamptothecin. Cancer Lett. 2010, 298, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Kuai, R.; Subramanian, C.; White, P.T.; Timmermann, B.N.; Moon, J.J.; Cohen, M.S.; Schwendeman, A. Synthetic high-density lipoprotein nanodisks for targeted withalongolide delivery to adrenocortical carcinoma. Int. J. Nanomed. 2017, 12, 6581–6594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahzad, M.M.K.; Mangala, L.S.; Han, H.D.; Lu, C.; Bottsford-Miller, J.; Nishimura, M.; Mora, E.M.; Lee, J.-W.; Stone, R.L.; Pecot, C.V.; et al. Targeted Delivery of Small Interfering RNA Using Reconstituted High-Density Lipoprotein Nanoparticles. Neoplasia 2011, 13, 309–319. [Google Scholar] [CrossRef] [Green Version]
- Shah, S.; Chib, R.; Raut, S.; Bermudez, J.; Sabnis, N.; Duggal, D.; Kimball, J.D.; Lacko, A.G.; Gryczynski, Z.; Gryczynski, I. Photophysical characterization of anticancer drug valrubicin in rHDL nanoparticles and its use as an imaging agent. J. Photochem. Photobiol. B 2016, 155, 60–65. [Google Scholar] [CrossRef] [Green Version]
- Sabnis, N.; Nair, M.; Israel, M.; McConathy, W.J.; Lacko, A.G. Enhanced solubility and functionality of valrubicin (AD-32) against cancer cells upon encapsulation into biocompatible nanoparticles. Int. J. Nanomed. 2012, 7, 975–983. [Google Scholar]
- Kreuter, J.; Shamenkov, D.; Petrov, V.; Ramge, P.; Cychutek, K.; Koch-Brandt, C.; Alyautdin, R. Apolipoprotein-mediated transport of nanoparticle-bound drugs across the blood-brain barrier. J. Drug Target. 2002, 10, 317–325. [Google Scholar] [CrossRef]
- Dubowchik, G.M.; Firestone, R.A. Improved cytotoxicity of antitumor compounds deliverable by the LDL pathway. Bioconjug. Chem. 1995, 6, 427–439. [Google Scholar] [CrossRef]
- Chu, A.C.Y.; Tsang, S.Y.; Lo, E.H.K.; Fung, K.P. Low density lipoprotein as a targeted carrier for doxorubicin in nude mice bearing human hepatoma HepG2 cells. Life Sci. 2001, 70, 591–601. [Google Scholar] [CrossRef]
- Shaw, J.M.; Shaw, K.V.; Yanovich, S.; Iwanik, M.; Futch, W.S.; Rosowsky, A.; Schook, L.B. Delivery of lipophilic drugs using lipoproteins. Ann. N. Y. Acad. Sci. 1987, 507, 252–271. [Google Scholar] [CrossRef] [PubMed]
- Vitols, S.; Söderberg-Reid, K.; Masquelier, M.; Sjöström, B.; Peterson, C. Low density lipoprotein for delivery of a water-insoluble alkylating agent to malignant cells. In vitro and in vivo studies of a drug-lipoprotein complex. Br. J. Cancer 1990, 62, 724–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callahan, D.E.; Forte, T.M.; Javed Afzal, S.M.; Deen, D.F.; Kahl, S.B.; Bjornstad, K.A.; Bauer, W.F.; Blakely, E.A. Boronated protoporphyrin (BOPP): Localization in lysosomes of the human glioma cell line SF-767 with uptake modulated by lipoprotein levels. Int. J. Radiat. Oncol. 1999, 45, 761–771. [Google Scholar] [CrossRef]
- Jin, H.; Lovell, J.F.; Chen, J.; Ng, K.; Cao, W.; Ding, L.; Zhang, Z.; Zheng, G. Cytosolic delivery of LDL nanoparticle cargo using photochemical internalization. Photochem. Photobiol. Sci. 2011, 10, 810–816. [Google Scholar] [CrossRef]
- Firestone, R.A.; Pisano, J.M.; Falck, J.R.; McPhaul, M.M.; Krieger, M. Selective delivery of cytotoxic compounds to cells by the LDL pathway. J. Med. Chem. 1984, 27, 1037–1043. [Google Scholar] [CrossRef]
- Rensen, P.C.N.; de Vrueh, R.L.A.; Kuiper, J.; Bijsterbosch, M.K.; Biessen, E.A.L.; van Berkel, T.J.C. Recombinant lipoproteins: Lipoprotein-like lipid particles for drug targeting. Adv. Drug Deliv. Rev. 2001, 47, 251–276. [Google Scholar] [CrossRef]
- Vitols, S.G.; Masquelier, M.; Peterson, C.O. Selective uptake of a toxic lipophilic anthracycline derivative by the low density lipoprotein receptor pathway in cultured fibroblasts. J. Med. Chem. 1985, 28, 451–454. [Google Scholar] [CrossRef]
- Lundberg, B. Cytotoxic activity of two new lipophilic steroid nitrogen carbamates incorporated into low-density lipoprotein. Anticancer. Drug Des. 1994, 9, 471–476. [Google Scholar]
- Lundberg, B. Preparation of drug-low density lipoprotein complexes for delivery of antitumoral drugs via the low density lipoprotein pathway. Cancer Res. 1987, 47, 4105–4108. [Google Scholar]
- Allijn, I.E.; Leong, W.; Tang, J.; Gianella, A.; Mieszawska, A.J.; Fay, F.; Ma, G.; Russell, S.; Callo, C.B.; Gordon, R.E.; et al. Gold nanocrystal labeling allows low-density lipoprotein imaging from the subcellular to macroscopic level. ACS Nano 2013, 7, 9761–9770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, M.L.; Corbin, I.R.; Levitin, R.B.; Cao, W.; Mainprize, J.G.; Yaffe, M.J.; Zheng, G. In vitro assessment of poly-iodinated triglyceride reconstituted low-density lipoprotein. Acad. Radiol. 2010, 17, 1359–1365. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Adhikari, B.B.; Cruz, S.; Schramm, M.P.; Vinson, J.A.; Narayanaswami, V. Targeted Intracellular Delivery of Resveratrol to Glioblastoma Cells Using Apolipoprotein E-Containing Reconstituted HDL as a Nanovehicle. PLoS ONE 2015, 10, e0135130. [Google Scholar] [CrossRef] [PubMed]
- Chuang, S.; Shon, Y.-S.; Narayanaswami, V. Apolipoprotein E3-mediated cellular uptake of reconstituted high-density lipoprotein bearing core 3, 10, or 17 nm hydrophobic gold nanoparticles. Int. J. Nanomed. 2017, 12, 8495–8510. [Google Scholar] [CrossRef] [Green Version]
- Zlokovic, B.V. Cerebrovascular transport of Alzheimer’s amyloid beta and apolipoproteins J and E: Possible anti-amyloidogenic role of the blood-brain barrier. Life Sci. 1996, 59, 1483–1497. [Google Scholar] [CrossRef]
- Calero, M.; Rostagno, A.; Matsubara, E.; Zlokovic, B.; Frangione, B.; Ghiso, J. Apolipoprotein J (clusterin) and Alzheimer’s disease. Microsc. Res. Tech. 2000, 50, 305–315. [Google Scholar] [CrossRef]
- Nikanjam, M.; Blakely, E.A.; Bjornstad, K.A.; Shu, X.; Budinger, T.F.; Forte, T.M. Synthetic nano-low density lipoprotein as targeted drug delivery vehicle for glioblastoma multiforme. Int. J. Pharm. 2007, 328, 86–94. [Google Scholar] [CrossRef] [Green Version]
- Nikanjam, M.; Gibbs, A.R.; Hunt, C.A.; Budinger, T.F.; Forte, T.M. Synthetic nano-LDL with paclitaxel oleate as a targeted drug delivery vehicle for glioblastoma multiforme. J. Control. Release 2007, 124, 163–171. [Google Scholar] [CrossRef]
- Baillie, G.; Owens, M.D.; Halbert, G.W. A synthetic low density lipoprotein particle capable of supporting U937 proliferation in vitro. J. Lipid Res. 2002, 43, 69–73. [Google Scholar]
- Hayavi, S.; Halbert, G.W. Synthetic low-density lipoprotein, a novel biomimetic lipid supplement for serum-free tissue culture. Biotechnol. Prog. 2008, 21, 1262–1268. [Google Scholar] [CrossRef]
- Ji, B.; Peacock, G.; Lu, D.R. Synthesis of cholesterol–carborane conjugate for targeted drug delivery. Bioorg. Med. Chem. Lett. 2002, 12, 2455–2458. [Google Scholar] [CrossRef]
- Masquelier, M.; Lundberg, B.; Peterson, C.; Vitols, S. Cytotoxic effect of a lipophilic alkylating agent after incorporation into low density lipoprotein or emulsions: Studies in human leukemic cells. Leuk. Res. 2006, 30, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Huntosova, V.; Buzova, D.; Petrovajova, D.; Kasak, P.; Nadova, Z.; Jancura, D.; Sureau, F.; Miskovsky, P. Development of a new LDL-based transport system for hydrophobic/amphiphilic drug delivery to cancer cells. Int. J. Pharm. 2012, 436, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.; Jain, K.; Mehra, N.K.; Jain, N.K. Lipoproteins tethered dendrimeric nanoconstructs for effective targeting to cancer cells. J. Nanopart. Res. 2013, 15, 2003. [Google Scholar] [CrossRef]
- Lerch, P.G.; Fortsch, V.; Hodler, G.; Bolli, R. Production and characterization of a reconstituted high density lipoprotein for therapeutic applications. Vox Sang. 1996, 71, 155–164. [Google Scholar] [CrossRef]
- Newton, R.S.; Krause, B.R. HDL therapy for the acute treatment of atherosclerosis. Atheroscler. Suppl. 2002, 3, 31–38. [Google Scholar] [CrossRef]
- Damiano, M.G.; Mutharasan, R.K.; Tripathy, S.; McMahon, K.M.; Thaxton, C.S. Templated high density lipoprotein nanoparticles as potential therapies and for molecular delivery. Adv. Drug Deliv. Rev. 2013, 65, 649–662. [Google Scholar] [CrossRef]
- Kuai, R.; Li, D.; Chen, Y.E.; Moon, J.J.; Schwendeman, A. High-density lipoproteins: Nature’s multifunctional nanoparticles. ACS Nano 2016, 10, 3015–3041. [Google Scholar] [CrossRef] [Green Version]
- Simonsen, J.B. Evaluation of reconstituted high-density lipoprotein (rHDL) as a drug delivery platform—A detailed survey of rHDL particles ranging from biophysical properties to clinical implications. Nanomed. Nanotechnol. Biol. Med. 2016, 12, 2161–2179. [Google Scholar] [CrossRef]
- Mooberry, L.K.; Sabnis, N.A.; Panchoo, M.; Nagarajan, B.; Lacko, A.G. Targeting the SR-B1 receptor as a gateway for cancer therapy and imaging. Front. Pharmacol. 2016, 7, 466. [Google Scholar] [CrossRef]
- Huang, H.; Cruz, W.; Chen, J.; Zheng, G. Learning from biology: Synthetic lipoproteins for drug delivery. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2015, 7, 298–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thaxton, C.S.; Rink, J.S.; Naha, P.C.; Cormode, D.P. Lipoproteins and lipoprotein mimetics for imaging and drug delivery. Adv. Drug Deliv. Rev. 2016, 106, 116–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raut, S.; Mooberry, L.; Sabnis, N.; Garud, A.; Dossou, A.S.; Lacko, A. Reconstituted HDL: Drug delivery platform for overcoming biological barriers to cancer therapy. Front. Pharmacol. 2018, 9, 1154. [Google Scholar] [CrossRef] [PubMed]
- Melchior, J.T.; Walker, R.G.; Cooke, A.L.; Morris, J.; Castleberry, M.; Thompson, T.B.; Jones, M.K.; Song, H.D.; Rye, K.-A.; Oda, M.N.; et al. A consensus model of human apolipoprotein A-I in its monomeric and lipid-free state. Nat. Struct. Mol. Biol. 2017, 24, 1093–1099. [Google Scholar] [CrossRef] [PubMed]
- Phillips, M.C. Is ABCA1 a lipid transfer protein? J. Lipid Res. 2018, 59, 749–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, W.J.; Azhar, S.; Kraemer, F.B. SR-B1: A unique multifunctional receptor for cholesterol influx and efflux. Annu. Rev. Physiol. 2018, 80, 95–116. [Google Scholar] [CrossRef] [PubMed]
- de Gonzalo-Calvo, D.; López-Vilaró, L.; Nasarre, L.; Perez-Olabarria, M.; Vázquez, T.; Escuin, D.; Badimon, L.; Barnadas, A.; Lerma, E.; Llorente-Cortés, V. Intratumor cholesteryl ester accumulation is associated with human breast cancer proliferation and aggressive potential: A molecular and clinicopathological study. BMC Cancer 2015, 15, 460. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Wang, J.; Li, M.; Yin, L.; Li, X.-A.; Zhang, T.-G. Up-regulated expression of scavenger receptor class B type 1 (SR-B1) is associated with malignant behaviors and poor prognosis of breast cancer. Pathol. Res. Pract. 2016, 212, 555–559. [Google Scholar] [CrossRef]
- Vasquez, M.; Simões, I.; Consuegra-Fernández, M.; Aranda, F.; Lozano, F.; Berraondo, P. Exploiting scavenger receptors in cancer immunotherapy: Lessons from CD5 and SR-B1. Eur. J. Immunol. 2017, 47, 1108–1118. [Google Scholar] [CrossRef] [Green Version]
- Cruz, P.M.R.; Mo, H.; McConathy, W.J.; Sabnis, N.; Lacko, A.G. The role of cholesterol metabolism and cholesterol transport in carcinogenesis: A review of scientific findings, relevant to future cancer therapeutics. Front. Pharmacol. 2013, 4, 119. [Google Scholar] [CrossRef] [Green Version]
- Williams, D.L.; de la Llera-Moya, M.; Thuahnai, S.T.; Lund-Katz, S.; Connelly, M.A.; Azhar, S.; Anantharamaiah, G.M.; Phillips, M.C. Binding and cross-linking studies show that scavenger receptor BI interacts with multiple sites in apolipoprotein A-I and identify the class A amphipathic α-helix as a recognition motif. J. Biol. Chem. 2000, 275, 18897–18904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillard, B.K.; Bassett, G.R.; Gotto, A.M.; Rosales, C.; Pownall, H.J. Scavenger receptor B1 (SR-B1) profoundly excludes high density lipoprotein (HDL) apolipoprotein AII as it nibbles HDL-cholesteryl ester. J. Biol. Chem. 2017, 292, 8864–8873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edelstein, C.; Halari, M.; Scanu, A.M. On the mechanism of the displacement of apolipoprotein A-I by apolipoprotein A-II from the high density lipoprotein surface. Effect of concentration and molecular forms of apolipoprotein A-II. J. Biol. Chem. 1982, 257, 7189–7195. [Google Scholar] [PubMed]
- Liang, H.Q.; Rye, K.A.; Barter, P.J. Remodelling of reconstituted high density lipoproteins by lecithin: Cholesterol acyltransferase. J. Lipid Res. 1996, 37, 1962–1970. [Google Scholar]
- Mehta, R.; Gantz, D.L.; Gursky, O. Human Plasma High-density Lipoproteins are Stabilized by Kinetic Factors. J. Mol. Biol. 2003, 328, 183–192. [Google Scholar] [CrossRef]
- Gillard, B.K.; Courtney, H.S.; Massey, J.B.; Pownall, H.J. Serum Opacity Factor Unmasks Human Plasma High-Density Lipoprotein Instability via Selective Delipidation and Apolipoprotein A-I Desorption. Biochemistry 2007, 46, 12968–12978. [Google Scholar] [CrossRef] [Green Version]
- Nakayama, T.; Butler, J.S.; Sehgal, A.; Severgnini, M.; Racie, T.; Sharman, J.; Ding, F.; Morskaya, S.S.; Brodsky, J.; Tchangov, L.; et al. Harnessing a Physiologic Mechanism for siRNA Delivery With Mimetic Lipoprotein Particles. Mol. Ther. 2012, 20, 1582–1589. [Google Scholar] [CrossRef] [Green Version]
- Rudenko, G.; Deisenhofer, J. The low-density lipoprotein receptor: Ligands, debates and lore. Curr. Opin. Struct. Biol. 2003, 13, 683–689. [Google Scholar] [CrossRef]
- Herz, J.; Strickland, D.K. LRP: A multifunctional scavenger and signaling receptor. J. Clin. Investig. 2001, 108, 779–784. [Google Scholar] [CrossRef]
- Chen, J.; Li, Q.; Wang, J. Topology of human apolipoprotein E3 uniquely regulates its diverse biological functions. Proc. Natl. Acad. Sci. USA 2011, 108, 14813–14818. [Google Scholar] [CrossRef] [Green Version]
- Weisgraber, K.H. Apolipoprotein E: Structure-function relationships. Adv. Protein Chem. 1994, 45, 249–302. [Google Scholar] [PubMed]
- Fisher, C.; Abdul-Aziz, D.; Blacklow, S.C. A Two-Module Region of the Low-Density Lipoprotein Receptor Sufficient for Formation of Complexes with Apolipoprotein E Ligands. Biochemistry 2004, 43, 1037–1044. [Google Scholar] [CrossRef]
- Fass, D.; Blacklow, S.; Kim, P.S.; Berger, J.M. Molecular basis of familial hypercholesterolaemia from structure of LDL receptor module. Nature 1997, 388, 691–693. [Google Scholar] [CrossRef] [PubMed]
- Lund-Katz, S.; Zaiou, M.; Wehrli, S.; Dhanasekaran, P.; Baldwin, F.; Weisgraber, K.H.; Phillips, M.C. Effects of Lipid Interaction on the Lysine Microenvironments in Apolipoprotein E. J. Biol. Chem. 2000, 275, 34459–34464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guttman, M.; Prieto, J.H.; Handel, T.M.; Domaille, P.J.; Komives, E.A. Structure of the Minimal Interface Between ApoE and LRP. J. Mol. Biol. 2010, 398, 306–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, C.; Wardell, M.; Weisgraber, K.; Mahley, R.; Agard, D. Three-dimensional structure of the LDL receptor-binding domain of human apolipoprotein E. Science 1991, 252, 1817–1822. [Google Scholar] [CrossRef]
- Sivashanmugam, A.; Wang, J. A Unified Scheme for Initiation and Conformational Adaptation of Human Apolipoprotein E N-terminal Domain upon Lipoprotein Binding and for Receptor Binding Activity. J. Biol. Chem. 2009, 284, 14657–14666. [Google Scholar] [CrossRef] [Green Version]
- Thuahnai, S.T.; Lund-Katz, S.; Williams, D.L.; Phillips, M.C. Scavenger Receptor Class B, Type I-mediated Uptake of Various Lipids into Cells: Influence of the Nature of the Donor Particle Interaction with the Receptor. J. Biol. Chem. 2001, 276, 43801–43808. [Google Scholar] [CrossRef] [Green Version]
- Pohlkamp, T.; Wasser, C.R.; Herz, J. Functional Roles of the Interaction of APP and Lipoprotein Receptors. Front. Mol. Neurosci. 2017, 10, 54. [Google Scholar] [CrossRef]
- Maletínská, L.; Blakely, E.A.; Bjornstad, K.A.; Deen, D.F.; Knoff, L.J.; Forte, T.M. Human glioblastoma cell lines: Levels of low-density lipoprotein receptor and low-density lipoprotein receptor-related protein. Cancer Res. 2000, 60, 2300–2303. [Google Scholar]
- Vitols, S.; Peterson, C.; Larsson, O.; Holm, P.; Aberg, B. Elevated uptake of low density lipoproteins by human lung cancer tissue in vivo. Cancer Res. 1992, 52, 6244–6247. [Google Scholar] [PubMed]
- Twiddy, A.L.; Cox, M.E.; Wasan, K.M. Knockdown of scavenger receptor Class B Type I reduces prostate specific antigen secretion and viability of prostate cancer cells. The Prostate 2012, 72, 955–965. [Google Scholar] [CrossRef] [PubMed]
- Danilo, C.; Gutierrez-Pajares, J.L.; Mainieri, M.A.; Mercier, I.; Lisanti, M.P.; Frank, P.G. Scavenger receptor class B type I regulates cellular cholesterol metabolism and cell signaling associated with breast cancer development. Breast Cancer Res. BCR 2013, 15, R87. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Liu, Y.; Jin, H.; Pan, S.; Qian, Y.; Huang, C.; Zeng, Y.; Luo, Q.; Zeng, M.; Zhang, Z. Scavenger receptor B1 is a potential biomarker of human nasopharyngeal carcinoma and its growth is inhibited by HDL-mimetic nanoparticles. Theranostics 2013, 3, 477–486. [Google Scholar] [CrossRef]
- Lek, M.T.; Cruz, S.; Ibe, N.U.; Beck, W.H.J.; Bielicki, J.K.; Weers, P.M.M.; Narayanaswami, V. Swapping the N- and C-terminal domains of human apolipoprotein E3 and AI reveals insights into their structure/activity relationship. PLoS ONE 2017, 12, e0178346. [Google Scholar] [CrossRef]
- Lillis, A.P.; Mikhailenko, I.; Strickland, D.K. Beyond endocytosis: LRP function in cell migration, proliferation and vascular permeability. J. Thromb. Haemost. 2005, 3, 1884–1893. [Google Scholar] [CrossRef]
- Yeh, F.L.; Wang, Y.; Tom, I.; Gonzalez, L.C.; Sheng, M. TREM2 Binds to Apolipoproteins, Including APOE and CLU/APOJ, and Thereby Facilitates Uptake of Amyloid-Beta by Microglia. Neuron 2016, 91, 328–340. [Google Scholar] [CrossRef]
- Jin, S.C.; Carrasquillo, M.M.; Benitez, B.A.; Skorupa, T.; Carrell, D.; Patel, D.; Lincoln, S.; Krishnan, S.; Kachadoorian, M.; Reitz, C.; et al. TREM2 is associated with increased risk for Alzheimer’s disease in African Americans. Mol. Neurodegener. 2015, 10, 19. [Google Scholar] [CrossRef] [Green Version]
- Gratuze, M.; Leyns, C.E.G.; Holtzman, D.M. New insights into the role of TREM2 in Alzheimer’s disease. Mol. Neurodegener. 2018, 13, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Saito, H.; Dhanasekaran, P.; Nguyen, D.; Baldwin, F.; Weisgraber, K.H.; Wehrli, S.; Phillips, M.C.; Lund-Katz, S. Characterization of the Heparin Binding Sites in Human Apolipoprotein E. J. Biol. Chem. 2003, 278, 14782–14787. [Google Scholar] [CrossRef] [Green Version]
- Mahley, R.W.; Rall, S.C. Apolipoprotein E: Far more than a lipid transport protein. Annu. Rev. Genomics Hum. Genet. 2000, 1, 507–537. [Google Scholar] [CrossRef] [PubMed]
- Levitan, I.; Volkov, S.; Subbaiah, P.V. Oxidized LDL: Diversity, Patterns of Recognition, and Pathophysiology. Antioxid. Redox Signal. 2010, 13, 39–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parthasarathy, S.; Raghavamenon, A.; Garelnabi, M.O.; Santanam, N. Oxidized Low-Density Lipoprotein. In Free Radicals and Antioxidant Protocols; Uppu, R., Murthy, S., Pryor, W., Parinandi, N., Eds.; Humana Press: Totowa, NJ, USA, 2010; Volume 610, pp. 403–417. [Google Scholar]
- Chadwick, A.C.; Holme, R.L.; Chen, Y.; Thomas, M.J.; Sorci-Thomas, M.G.; Silverstein, R.L.; Pritchard, K.A.; Sahoo, D. Acrolein Impairs the Cholesterol Transport Functions of High Density Lipoproteins. PLoS ONE 2015, 10, e0123138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz, S.; Narayanaswami, V. Cellular Uptake and Clearance of Oxidatively-modified Apolipoprotein E3 by Cerebral Cortex Endothelial Cells. Int. J. Mol. Sci. 2019, 20, 4582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valacchi, G.; Sticozzi, C.; Lim, Y.; Pecorelli, A. Scavenger receptor class B type I: A multifunctional receptor: Valacchi et al. Ann. N. Y. Acad. Sci. 2011, 1229, E1–E7. [Google Scholar] [CrossRef]
- Bachmeier, C.; Shackleton, B.; Ojo, J.; Paris, D.; Mullan, M.; Crawford, F. Apolipoprotein E isoform-specific effects on lipoprotein receptor processing. Neuromolecular Med. 2014, 16, 686–696. [Google Scholar] [CrossRef] [Green Version]
- Rebeck, G.W.; LaDu, M.J.; Estus, S.; Bu, G.; Weeber, E.J. The generation and function of soluble apoE receptors in the CNS. Mol. Neurodegener. 2006, 1, 15. [Google Scholar] [CrossRef] [Green Version]
- Fernández-de-Retana, S.; Cano-Sarabia, M.; Marazuela, P.; Sánchez-Quesada, J.L.; Garcia-Leon, A.; Montañola, A.; Montaner, J.; Maspoch, D.; Hernández-Guillamon, M. Characterization of ApoJ-reconstituted high-density lipoprotein (rHDL) nanodisc for the potential treatment of cerebral β-amyloidosis. Sci. Rep. 2017, 7, 14637. [Google Scholar] [CrossRef]
- Zensi, A.; Begley, D.; Pontikis, C.; Legros, C.; Mihoreanu, L.; Wagner, S.; Büchel, C.; von Briesen, H.; Kreuter, J. Albumin nanoparticles targeted with Apo E enter the CNS by transcytosis and are delivered to neurones. J. Control. Release 2009, 137, 78–86. [Google Scholar] [CrossRef]
- Park, K. Transport across the blood-brain barrier using albumin nanoparticles. J. Control. Release 2009, 137, 1. [Google Scholar] [CrossRef]
- Wagner, S.; Zensi, A.; Wien, S.L.; Tschickardt, S.E.; Maier, W.; Vogel, T.; Worek, F.; Pietrzik, C.U.; Kreuter, J.; von Briesen, H. Uptake Mechanism of ApoE-Modified Nanoparticles on Brain Capillary Endothelial Cells as a Blood-Brain Barrier Model. PLoS ONE 2012, 7, e32568. [Google Scholar] [CrossRef] [PubMed]
- Kreuter, J.; Hekmatara, T.; Dreis, S.; Vogel, T.; Gelperina, S.; Langer, K. Covalent attachment of apolipoprotein A-I and apolipoprotein B-100 to albumin nanoparticles enables drug transport into the brain. J. Control. Release 2007, 118, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Michaelis, K.; Hoffmann, M.M.; Dreis, S.; Herbert, E.; Alyautdin, R.N.; Michaelis, M.; Kreuter, J.; Langer, K. Covalent linkage of apolipoprotein E to albumin nanoparticles strongly enhances drug transport into the brain. J. Pharmacol. Exp. Ther. 2006, 317, 1246–1253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, S.T.; Navab, M.; Anantharamaiah, G.M.; Fogelman, A.M. Apolipoprotein A-I mimetics. Curr. Opin. Lipidol. 2014, 25, 304–308. [Google Scholar] [CrossRef] [Green Version]
- Getz, G.S.; Reardon, C.A. Apolipoprotein A-I and A-I mimetic peptides: A role in atherosclerosis. J. Inflamm. Res. 2011, 4, 83. [Google Scholar] [CrossRef]
- White, C.R.; Garber, D.W.; Anantharamaiah, G.M. Anti-inflammatory and cholesterol-reducing properties of apolipoprotein mimetics: A review. J. Lipid Res. 2014, 55, 2007–2021. [Google Scholar] [CrossRef] [Green Version]
- Imura, T.; Tsukui, Y.; Taira, T.; Aburai, K.; Sakai, K.; Sakai, H.; Abe, M.; Kitamoto, D. Surfactant-like Properties of an Amphiphilic α-Helical Peptide Leading to Lipid Nanodisc Formation. Langmuir 2014, 30, 4752–4759. [Google Scholar] [CrossRef]
- Smythies, L.E.; White, C.R.; Maheshwari, A.; Palgunachari, M.N.; Anantharamaiah, G.M.; Chaddha, M.; Kurundkar, A.R.; Datta, G. Apolipoprotein A-I mimetic 4F alters the function of human monocyte-derived macrophages. Am. J. Physiol. Cell Physiol. 2010, 298, C1538–C1548. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Kim, S.H.; Patel, A.B.; Narayanaswami, V.; Iavarone, A.T.; Hura, G.L.; Bielicki, J.K. The positional specificity of EXXK motifs within an amphipathic α-helix dictates preferential lysine modification by acrolein: Implications for the design of high-density lipoprotein mimetic peptides. Biochemistry 2012, 51, 6400–6412. [Google Scholar] [CrossRef]
- Kariyazono, H.; Nadai, R.; Miyajima, R.; Takechi-Haraya, Y.; Baba, T.; Shigenaga, A.; Okuhira, K.; Otaka, A.; Saito, H. Formation of stable nanodiscs by bihelical apolipoprotein A-I mimetic peptide. J. Pep. Sci. 2016, 22, 116–122. [Google Scholar] [CrossRef]
- Carlson, M.L.; Young, J.W.; Zhao, Z.; Fabre, L.; Jun, D.; Li, J.; Li, J.; Dhupar, H.S.; Wason, I.; Mills, A.T.; et al. The Peptidisc, a simple method for stabilizing membrane proteins in detergent-free solution. eLife 2018, 7, e34085. [Google Scholar] [CrossRef] [PubMed]
- Patel, H.; Ding, B.; Ernst, K.; Shen, L.; Yuan, W.; Tang, J.; Drake, L.R.; Kang, J.; Li, Y.; Chen, Z.; et al. Characterization of apolipoprotein A-I peptide phospholipid interaction and its effect on HDL nanodisc assembly. Int. J. Nanomed. 2019, 14, 3069–3086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bielicki, J.K. ABCA1 agonist peptides for the treatment of disease. Curr. Opin. Lipidol. 2016, 27, 40–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bielicki, J.K.; Zhang, H.; Cortez, Y.; Zheng, Y.; Narayanaswami, V.; Patel, A.; Johansson, J.; Azhar, S. A new HDL mimetic peptide that stimulates cellular cholesterol efflux with high efficiency greatly reduces atherosclerosis in mice. J. Lipid Res. 2010, 51, 1496–1503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anantharamaiah, G.M.; Jones, J.L.; Brouillette, C.G.; Schmidt, C.F.; Chung, B.H.; Hughes, T.A.; Bhown, A.S.; Segrest, J.P. Studies of synthetic peptide analogs of the amphipathic helix. Structure of complexes with dimyristoyl phosphatidylcholine. J. Biol. Chem. 1985, 260, 10248–10255. [Google Scholar] [PubMed]
- Re, F.; Cambianica, I.; Zona, C.; Sesana, S.; Gregori, M.; Rigolio, R.; La Ferla, B.; Nicotra, F.; Forloni, G.; Cagnotto, A.; et al. Functionalization of liposomes with ApoE-derived peptides at different density affects cellular uptake and drug transport across a blood-brain barrier model. Nanomed. Nanotechnol. Biol. Med. 2011, 7, 551–559. [Google Scholar] [CrossRef]
- Sauer, I.; Nikolenko, H.; Keller, S.; Abu Ajaj, K.; Bienert, M.; Dathe, M. Dipalmitoylation of a cellular uptake-mediating apolipoprotein E-derived peptide as a promising modification for stable anchorage in liposomal drug carriers. Biochim. Biophys. Acta 2006, 1758, 552–561. [Google Scholar] [CrossRef] [Green Version]
- Jonas, A. Reconstitution of high-density lipoproteins. Methods Enzymol. 1986, 128, 553–582. [Google Scholar]
- Wan, C.-P.L.; Chiu, M.H.; Wu, X.; Lee, S.K.; Prenner, E.J.; Weers, P.M.M. Apolipoprotein-induced conversion of phosphatidylcholine bilayer vesicles into nanodisks. Biochim. Biophys. Acta BBA Biomembr. 2011, 1808, 606–613. [Google Scholar] [CrossRef] [Green Version]
- Surewicz, W.K.; Epand, R.M.; Pownall, H.J.; Hui, S.W. Human apolipoprotein A-I forms thermally stable complexes with anionic but not with zwitterionic phospholipids. J. Biol. Chem. 1986, 261, 16191–16197. [Google Scholar]
- Her, C.; Filoti, D.I.; McLean, M.A.; Sligar, S.G.; Alexander Ross, J.B.; Steele, H.; Laue, T.M. The Charge Properties of Phospholipid Nanodiscs. Biophys. J. 2016, 111, 989–998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narayanaswami, V.; Wang, J.; Schieve, D.; Kay, C.M.; Ryan, R.O. A molecular trigger of lipid binding-induced opening of a helix bundle exchangeable apolipoprotein. Proc. Natl. Acad. Sci. USA 1999, 96, 4366–4371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weers, P.M.M.; Narayanaswami, V.; Kay, C.M.; Ryan, R.O. Interaction of an Exchangeable Apolipoprotein with Phospholipid Vesicles and Lipoprotein Particles: Role of Leucines 32, 34, and 95 in Locusta migratoria Apolipophorin III. J. Biol. Chem. 1999, 274, 21804–21810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redmond, K.A.; Murphy, C.; Narayanaswami, V.; Kiss, R.S.; Hauser, P.; Guigard, E.; Kay, C.M.; Ryan, R.O. Replacement of helix 1’ enhances the lipid binding activity of apoE3 N-terminal domain. FEBS J. 2006, 273, 558–567. [Google Scholar] [CrossRef] [PubMed]
- Narayanaswami, V.; Kiss, R.S.; Weers, P.M.M. The helix bundle: A reversible lipid binding motif. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2010, 155, 123–133. [Google Scholar] [CrossRef] [Green Version]
- Weers, P.M.; Van Der Horst, D.J.; Ryan, R.O. Interaction of locust apolipophorin III with lipoproteins and phospholipid vesicles: Effect of glycosylation. J. Lipid Res. 2000, 41, 416–423. [Google Scholar]
- Wetterau, J.R.; Aggerbeck, L.P.; Rall, S.C.; Weisgraber, K.H. Human apolipoprotein E3 in aqueous solution. I. Evidence for two structural domains. J. Biol. Chem. 1988, 263, 6240–6248. [Google Scholar]
- Kiss, R.S.; Weers, P.M.M.; Narayanaswami, V.; Cohen, J.; Kay, C.M.; Ryan, R.O. Structure-guided protein engineering modulates helix bundle exchangeable apolipoprotein properties. J. Biol. Chem. 2003, 278, 21952–21959. [Google Scholar] [CrossRef] [Green Version]
- Nichols, A.V.; Gong, E.L.; Blanche, P.J.; Forte, T.M.; Shore, V.G. Pathways in the formation of human plasma high density lipoprotein subpopulations containing apolipoprotein A-I without apolipoprotein A-II. J. Lipid Res. 1987, 28, 719–732. [Google Scholar]
- Ghosh, M.; Singh, A.T.K.; Xu, W.; Sulchek, T.; Gordon, L.I.; Ryan, R.O. Curcumin nanodisks: Formulation and characterization. Nanomed. Nanotechnol. Biol. Med. 2011, 7, 162–167. [Google Scholar] [CrossRef] [Green Version]
- Matz, C.E.; Jonas, A. Micellar complexes of human apolipoprotein A-I with phosphatidylcholines and cholesterol prepared from cholate-lipid dispersions. J. Biol. Chem. 1982, 257, 4535–4540. [Google Scholar] [PubMed]
- Kim, S.H.; Kothari, S.; Patel, A.B.; Bielicki, J.K.; Narayanaswami, V. A pyrene based fluorescence approach to study conformation of apolipoprotein E3 in macrophage-generated nascent high density lipoprotein. Biochem. Biophys. Res. Commun. 2014, 450, 124–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Fay, F.; Cormode, D.P.; Sanchez-Gaytan, B.L.; Tang, J.; Hennessy, E.J.; Ma, M.; Moore, K.; Farokhzad, O.C.; Fisher, E.A.; et al. Single step reconstitution of multifunctional high-density lipoprotein-derived nanomaterials using microfluidics. ACS Nano 2013, 7, 9975–9983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cormode, D.P.; Roessl, E.; Thran, A.; Skajaa, T.; Gordon, R.E.; Schlomka, J.-P.; Fuster, V.; Fisher, E.A.; Mulder, W.J.M.; Proksa, R.; et al. Atherosclerotic plaque composition: Analysis with multicolor CT and targeted gold nanoparticles. Radiology 2010, 256, 774–782. [Google Scholar] [CrossRef] [PubMed]
- Thaxton, C.S.; Daniel, W.L.; Giljohann, D.A.; Thomas, A.D.; Mirkin, C.A. Templated spherical high density lipoprotein nanoparticles. J. Am. Chem. Soc. 2009, 131, 1384–1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luthi, A.J.; Zhang, H.; Kim, D.; Giljohann, D.A.; Mirkin, C.A.; Thaxton, C.S. Tailoring of biomimetic high-density lipoprotein nanostructures changes cholesterol binding and efflux. ACS Nano 2012, 6, 276–285. [Google Scholar] [CrossRef] [Green Version]
- McMahon, K.M.; Mutharasan, R.K.; Tripathy, S.; Veliceasa, D.; Bobeica, M.; Shumaker, D.K.; Luthi, A.J.; Helfand, B.T.; Ardehali, H.; Mirkin, C.A.; et al. Biomimetic high density lipoprotein nanoparticles for nucleic acid delivery. Nano Lett. 2011, 11, 1208–1214. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.T.K.; Ghosh, M.; Forte, T.M.; Ryan, R.O.; Gordon, L.I. Curcumin nanodisk-induced apoptosis in mantle cell lymphoma. Leuk. Lymphoma 2011, 52, 1537–1543. [Google Scholar] [CrossRef]
- Duivenvoorden, R.; Tang, J.; Cormode, D.P.; Mieszawska, A.J.; Izquierdo-Garcia, D.; Ozcan, C.; Otten, M.J.; Zaidi, N.; Lobatto, M.E.; van Rijs, S.M.; et al. A statin-loaded reconstituted high-density lipoprotein nanoparticle inhibits atherosclerotic plaque inflammation. Nat. Commun. 2014, 5, 3065. [Google Scholar] [CrossRef] [Green Version]
- Neves, A.R.; Queiroz, J.F.; Reis, S. Brain-targeted delivery of resveratrol using solid lipid nanoparticles functionalized with apolipoprotein E. J. Nanobiotechnol. 2016, 14, 27. [Google Scholar] [CrossRef] [Green Version]
- Martins, P.A.T.; Alsaiari, S.; Julfakyan, K.; Nie, Z.; Khashab, N.M. Self-assembled lipoprotein based gold nanoparticles for detection and photothermal disaggregation of β-amyloid aggregates. Chem. Commun. 2017, 53, 2102–2105. [Google Scholar] [CrossRef]
- Kontush, A.; Lhomme, M.; Chapman, M.J. Unraveling the complexities of the HDL lipidome. J. Lipid Res. 2013, 54, 2950–2963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poloni, D.M.; Dangles, O.; Vinson, J.A. Binding of plant polyphenols to serum albumin and LDL: Healthy implications for heart disease. J. Agric. Food Chem. 2019, 67, 9139–9147. [Google Scholar] [CrossRef]
- Sharma, R.A.; Gescher, A.J.; Steward, W.P. Curcumin: The story so far. Eur. J. Cancer 2005, 41, 1955–1968. [Google Scholar] [CrossRef] [PubMed]
- Ganjali, S.; Blesso, C.N.; Banach, M.; Pirro, M.; Majeed, M.; Sahebkar, A. Effects of curcumin on HDL functionality. Pharmacol. Res. 2017, 119, 208–218. [Google Scholar] [CrossRef] [PubMed]
- Ono, K.; Hasegawa, K.; Naiki, H.; Yamada, M. Curcumin has potent anti-amyloidogenic effects for Alzheimer’s beta-amyloid fibrils in vitro. J. Neurosci. Res. 2004, 75, 742–750. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Lim, G.P.; Begum, A.N.; Ubeda, O.J.; Simmons, M.R.; Ambegaokar, S.S.; Chen, P.P.; Kayed, R.; Glabe, C.G.; Frautschy, S.A.; et al. Curcumin inhibits formation of amyloid β oligomers and fibrils, binds plaques, and reduces amyloid in vivo. J. Biol. Chem. 2005, 280, 5892–5901. [Google Scholar] [CrossRef] [Green Version]
- Lim, G.P.; Chu, T.; Yang, F.; Beech, W.; Frautschy, S.A.; Cole, G.M. The curry spice curcumin reduces oxidative damage and amyloid pathology in an Alzheimer transgenic mouse. J. Neurosci. 2001, 21, 8370–8377. [Google Scholar] [CrossRef]
- Pal, S.; Ho, N.; Santos, C.; Dubois, P.; Mamo, J.; Croft, K.; Allister, E. Red wine polyphenolics increase LDL receptor expression and activity and suppress the secretion of ApoB100 from human HepG2 cells. J. Nutr. 2003, 133, 700–706. [Google Scholar] [CrossRef]
- Frankel, E.N.; Waterhouse, A.L.; Kinsella, J.E. Inhibition of human LDL oxidation by resveratrol. Lancet 1993, 341, 1103–1104. [Google Scholar] [CrossRef]
- Khadem-Ansari, M.H.; Rasmi, Y.; Ramezani, F. Effects of red grape juice consumption on high density lipoprotein-cholesterol, apolipoprotein AI, apolipoprotein B and homocysteine in healthy human volunteers. Open Biochem. J. 2010, 4, 96–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vang, O.; Ahmad, N.; Baile, C.A.; Baur, J.A.; Brown, K.; Csiszar, A.; Das, D.K.; Delmas, D.; Gottfried, C.; Lin, H.-Y.; et al. What is new for an old molecule? Systematic review and recommendations on the use of resveratrol. PLoS ONE 2011, 6, e19881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de la Lastra, C.A.; Villegas, I. Resveratrol as an antioxidant and pro-oxidant agent: Mechanisms and clinical implications. Biochem. Soc. Trans. 2007, 35, 1156–1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.J.; Ahmad, F.; Philp, A.; Baar, K.; Williams, T.; Luo, H.; Ke, H.; Rehmann, H.; Taussig, R.; Brown, A.L.; et al. Resveratrol ameliorates aging-related metabolic phenotypes by inhibiting cAMP phosphodiesterases. Cell 2012, 148, 421–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walle, T.; Hsieh, F.; DeLegge, M.H.; Oatis, J.E.; Walle, U.K. High absorption but very low bioavailability of oral resveratrol in humans. Drug Metab. Dispos. 2004, 32, 1377–1382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuhnle, G.; Spencer, J.P.; Chowrimootoo, G.; Schroeter, H.; Debnam, E.S.; Srai, S.K.; Rice-Evans, C.; Hahn, U. Resveratrol is absorbed in the small intestine as resveratrol glucuronide. Biochem. Biophys. Res. Commun. 2000, 272, 212–217. [Google Scholar] [CrossRef]
- Miksits, M.; Maier-Salamon, A.; Aust, S.; Thalhammer, T.; Reznicek, G.; Kunert, O.; Haslinger, E.; Szekeres, T.; Jaeger, W. Sulfation of resveratrol in human liver: Evidence of a major role for the sulfotransferases SULT1A1 and SULT1E1. Xenobiotica Fate Foreign Compd. Biol. Syst. 2005, 35, 1101–1119. [Google Scholar] [CrossRef]
- Pangeni, R.; Sahni, J.K.; Ali, J.; Sharma, S.; Baboota, S. Resveratrol: Review on therapeutic potential and recent advances in drug delivery. Expert Opin. Drug Deliv. 2014, 11, 1285–1298. [Google Scholar] [CrossRef]
- Urpí-Sardà, M.; Jáuregui, O.; Lamuela-Raventós, R.M.; Jaeger, W.; Miksits, M.; Covas, M.-I.; Andres-Lacueva, C. Uptake of diet resveratrol into the human low-density lipoprotein. Identification and quantification of resveratrol metabolites by liquid chromatography coupled with tandem mass spectrometry. Anal. Chem. 2005, 77, 3149–3155. [Google Scholar] [CrossRef]
- Marambaud, P.; Zhao, H.; Davies, P. Resveratrol promotes clearance of Alzheimer’s disease amyloid-beta peptides. J. Biol. Chem. 2005, 280, 37377–37382. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.L.; Xiao, Y.; Liu, J.-P.; Wu, Z.-M.; Gu, X.; Xu, Y.-M.; Lu, H. Structure and remodeling behavior of drug-loaded high density lipoproteins and their atherosclerotic plaque targeting mechanism in foam cell model. Int. J. Pharm. 2011, 419, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Li, J.; Liu, J.; Wu, Z.; Xu, Y.; Wang, J. Tanshinone IIA-loaded reconstituted high density lipoproteins: Atherosclerotic plaque targeting mechanism in a foam cell model and pharmacokinetics in rabbits. Pharm. 2012, 67, 324–330. [Google Scholar]
- Ghosh, M.; Ren, G.; Simonsen, J.B.; Ryan, R.O. Cationic lipid nanodisks as an siRNA delivery vehicle. Biochem. Cell Biol. Biochim. Biol. Cell. 2014, 92, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Tamaru, M.; Akita, H.; Nakatani, T.; Kajimoto, K.; Sato, Y.; Hatakeyama, H.; Harashima, H. Application of apolipoprotein E-modified liposomal nanoparticles as a carrier for delivering DNA and nucleic acid in the brain. Int. J. Nanomed. 2014, 9, 4267–4276. [Google Scholar]
- Tamaru, M.; Akita, H.; Kajimoto, K.; Sato, Y.; Hatakeyama, H.; Harashima, H. An apolipoprotein E modified liposomal nanoparticle: Ligand dependent efficiency as a siRNA delivery carrier for mouse-derived brain endothelial cells. Int. J. Pharm. 2014, 465, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Henrich, S.E.; Hong, B.J.; Rink, J.S.; Nguyen, S.T.; Thaxton, C.S. Supramolecular assembly of high-density lipoprotein mimetic nanoparticles using lipid-conjugated core scaffolds. J. Am. Chem. Soc. 2019, 141, 9753–9757. [Google Scholar] [CrossRef]
- Soutschek, J.; Akinc, A.; Bramlage, B.; Charisse, K.; Constien, R.; Donoghue, M.; Elbashir, S.; Geick, A.; Hadwiger, P.; Harborth, J.; et al. Therapeutic silencing of an endogenous gene by systemic administration of modified siRNAs. Nature 2004, 432, 173–178. [Google Scholar] [CrossRef]
- Wolfrum, C.; Shi, S.; Jayaprakash, K.N.; Jayaraman, M.; Wang, G.; Pandey, R.K.; Rajeev, K.G.; Nakayama, T.; Charrise, K.; Ndungo, E.M.; et al. Mechanisms and optimization of in vivo delivery of lipophilic siRNAs. Nat. Biotechnol. 2007, 25, 1149–1157. [Google Scholar] [CrossRef]
- Tripathy, S.; Vinokour, E.; McMahon, K.M.; Volpert, O.V.; Thaxton, C.S. High density lipoprotein nanoparticles deliver RNAi to endothelial cells to inhibit angiogenesis. Part. Part. Syst. Charact. 2014, 31, 1141–1150. [Google Scholar] [CrossRef] [Green Version]
- Park, K.H.; Yun, C.O.; Kwon, O.J.; Kim, C.H.; Kim, J.R.; Cho, K.-H. Enhanced delivery of adenovirus, using proteoliposomes containing wildtype or V156K apolipoprotein A-I and dimyristoylphosphatidylcholine. Hum. Gene Ther. 2010, 21, 579–587. [Google Scholar] [CrossRef]
- Pottash, A.E.; Kuffner, C.; Noonan-Shueh, M.; Jay, S.M. Protein-based vehicles for biomimetic RNAi delivery. J. Biol. Eng. 2019, 13, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, J.; Xiao, Y.; Liu, J.; Zhang, W.; He, H.; Chen, L.; Zhang, M. Preparation, characterizations, and in vitro metabolic processes of paclitaxel-loaded discoidal recombinant high-density lipoproteins. J. Pharm. Sci. 2012, 101, 2900–2908. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Jia, J.; Liu, J.; He, H.; Zhang, W.; Li, Z. Tumor targeting effects of a novel modified paclitaxel-loaded discoidal mimic high density lipoproteins. Drug Deliv. 2013, 20, 356–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altucci, L.; Gronemeyer, H. The promise of retinoids to fight against cancer. Nat. Rev. Cancer 2001, 1, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Redmond, K.A.; Nguyen, T.-S.; Ryan, R.O. All-trans-retinoic acid nanodisks. Int. J. Pharm. 2007, 339, 246–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.T.K.; Evens, A.M.; Anderson, R.J.; Beckstead, J.A.; Sankar, N.; Sassano, A.; Bhalla, S.; Yang, S.; Platanias, L.C.; Forte, T.M.; et al. All trans retinoic acid nanodisks enhance retinoic acid receptor mediated apoptosis and cell cycle arrest in mantle cell lymphoma. Br. J. Haematol. 2010, 150, 158–169. [Google Scholar] [CrossRef] [Green Version]
- Berney, E.; Sabnis, N.; Panchoo, M.; Raut, S.; Dickerman, R.; Lacko, A.G. The SR-B1 Receptor as a Potential Target for Treating Glioblastoma. J. Oncol. 2019, 2019, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Su, Y.; Zhang, F.; Chen, K.; Xu, X.; Xu, L.; Zhou, J.; Wang, W. A dual-targeting reconstituted high density lipoprotein leveraging the synergy of sorafenib and antimiRNA21 for enhanced hepatocellular carcinoma therapy. Acta Biomater. 2018, 75, 413–426. [Google Scholar] [CrossRef]
- Ding, Y.; Wang, Y.; Opoku-Damoah, Y.; Wang, C.; Shen, L.; Yin, L.; Zhou, J. Dual-functional bio-derived nanoparticulates for apoptotic antitumor therapy. Biomaterials 2015, 72, 90–103. [Google Scholar] [CrossRef]
- Oda, M.N.; Hargreaves, P.L.; Beckstead, J.A.; Redmond, K.A.; van Antwerpen, R.; Ryan, R.O. Reconstituted high density lipoprotein enriched with the polyene antibiotic amphotericin B. J. Lipid Res. 2006, 47, 260–267. [Google Scholar] [CrossRef] [Green Version]
- Cho, K.-H. Enhanced delivery of rapamycin by V156K-apoA-I high-density lipoprotein inhibits cellular proatherogenic effects and senescence and promotes tissue regeneration. J. Gerontol. A 2011, 66, 1274–1285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bijsterbosch, M.K.; van de Bilt, H.; van Berkel, T.J. Specific targeting of a lipophilic prodrug of iododeoxyuridine to parenchymal liver cells using lactosylated reconstituted high density lipoprotein particles. Biochem. Pharmacol. 1996, 52, 113–121. [Google Scholar] [CrossRef]
- Bijsterbosch, M.K.; Van Berkel, T.J. Lactosylated high density lipoprotein: A potential carrier for the site-specific delivery of drugs to parenchymal liver cells. Mol. Pharmacol. 1992, 41, 404–411. [Google Scholar] [PubMed]
- Cormode, D.P.; Skajaa, T.; van Schooneveld, M.M.; Koole, R.; Jarzyna, P.; Lobatto, M.E.; Calcagno, C.; Barazza, A.; Gordon, R.E.; Zanzonico, P.; et al. Nanocrystal core high-density lipoproteins: A multimodality contrast agent platform. Nano Lett. 2008, 8, 3715–3723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skajaa, T.; Cormode, D.P.; Jarzyna, P.A.; Delshad, A.; Blachford, C.; Barazza, A.; Fisher, E.A.; Gordon, R.E.; Fayad, Z.A.; Mulder, W.J.M. The biological properties of iron oxide core high-density lipoprotein in experimental atherosclerosis. Biomaterials 2011, 32, 206–213. [Google Scholar] [CrossRef] [Green Version]
- Frias, J.C.; Williams, K.J.; Fisher, E.A.; Fayad, Z.A. Recombinant HDL-like nanoparticles: A specific contrast agent for MRI of atherosclerotic plaques. J. Am. Chem. Soc. 2004, 126, 16316–16317. [Google Scholar] [CrossRef]
- Cormode, D.P.; Briley-Saebo, K.C.; Mulder, W.J.M.; Aguinaldo, J.G.S.; Barazza, A.; Ma, Y.; Fisher, E.A.; Fayad, Z.A. An apoA-I mimetic peptide high-density-lipoprotein-based MRI contrast agent for atherosclerotic plaque composition detection. Small 2008, 4, 1437–1444. [Google Scholar] [CrossRef]
- Hardman, R. A toxicologic review of quantum dots: Toxicity depends on physicochemical and environmental factors. Environ. Health Perspect. 2006, 114, 165–172. [Google Scholar] [CrossRef]
- Skajaa, T.; Zhao, Y.; van den Heuvel, D.J.; Gerritsen, H.C.; Cormode, D.P.; Koole, R.; van Schooneveld, M.M.; Post, J.A.; Fisher, E.A.; Fayad, Z.A.; et al. Quantum dot and Cy5.5 labeled nanoparticles to investigate lipoprotein biointeractions via Förster resonance energy transfer. Nano Lett. 2010, 10, 5131–5138. [Google Scholar] [CrossRef] [Green Version]
- Crowley, J.T.; Toledo, A.M.; LaRocca, T.J.; Coleman, J.L.; London, E.; Benach, J.L. Lipid exchange between Borrelia burgdorferi and host cells. PLOS Pathog. 2013, 9, e1003109. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, L.C.; Bickford, L.R.; Lewinski, N.A.; Coughlin, A.J.; Hu, Y.; Day, E.S.; West, J.L.; Drezek, R.A. A new era for cancer treatment: Gold-nanoparticle-mediated thermal therapies. Small 2011, 7, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Hainfeld, J.F.; O’Connor, M.J.; Lin, P.; Qian, L.; Slatkin, D.N.; Smilowitz, H.M. Infrared-transparent gold nanoparticles converted by tumors to infrared absorbers cure tumors in mice by photothermal therapy. PLoS ONE 2014, 9, e88414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMahon, K.M.; Scielzo, C.; Angeloni, N.L.; Deiss-Yehiely, E.; Scarfo, L.; Ranghetti, P.; Ma, S.; Kaplan, J.; Barbaglio, F.; Gordon, L.I.; et al. Synthetic high-density lipoproteins as targeted monotherapy for chronic lymphocytic leukemia. Oncotarget 2017, 8, 11219–11227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabrilovich, D.I.; Nagaraj, S. Myeloid-derived suppressor cells as regulators of the immune system. Nat. Rev. Immunol. 2009, 9, 162–174. [Google Scholar] [CrossRef] [PubMed]
- Plebanek, M.P.; Bhaumik, D.; Bryce, P.J.; Thaxton, C.S. Scavenger Receptor Type B1 and lipoprotein nanoparticle inhibit myeloid-derived suppressor cells. Mol. Cancer Ther. 2018, 17, 686–697. [Google Scholar] [CrossRef] [Green Version]
- Crosby, N.M.; Ghosh, M.; Su, B.; Beckstead, J.A.; Kamei, A.; Simonsen, J.B.; Luo, B.; Gordon, L.I.; Forte, T.M.; Ryan, R.O. Anti-CD20 single chain variable antibody fragment-apolipoprotein A-I chimera containing nanodisks promote targeted bioactive agent delivery to CD20-positive lymphomas. Biochem. Cell Biol. 2015, 93, 343–350. [Google Scholar] [CrossRef] [Green Version]
- Herz, J. LRP: A bright beacon at the blood-brain barrier. J. Clin. Investig. 2003, 112, 1483–1485. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Apolipoprotein | Lipoprotein Association in Plasma or 1CNS | Physiological Function | Potential Applications |
|---|---|---|---|
| 2ApoAI | 3HDL, chylomicrons, 3VLDL | Structural protein in HDL; 4LCAT activator; promotes cholesterol efflux; binds 4SR-B1 | Anti-atherogenic and drug delivery [39,40,41,42,43,44,45], siRNA delivery [46], imaging agent delivery [47,48] |
| ApoAII | HDL, chylomicrons, VLDL | Structural protein in HDL; activates hepatic lipase | Drug delivery [49] |
| ApoB-100 | VLDL, 3IDL, 3LDL | Structural protein in VLDL, IDL, LDL; ligand for LDL receptor | Drug and imaging agent delivery [50,51,52,53,54,55,56,57,58,59,60,61,62] |
| ApoCII | Chylomicrons, VLDL, HDL | 4Lipoprotein lipase activator | Drug delivery [49] |
| ApoE3 | Chylomicron remnants, VLDL, IDL, HDL | Ligand for 4LDLr family of proteins; LCAT activator; antioxidant; promotes cholesterol efflux; binds SR-B1 | Anti-atherogenic, drug, flavonoid and imaging agent delivery [18,39,63,64] |
| ApoJ (clusterin) | HDL | Anti-amyloidogenic; chaperone; antioxidant | Reduce aggregation of Aβ [65,66] |
| Apolipoprotein | Nanoparticle Type | Nanoparticle Preparation Methodology | Comments and Perspectives |
|---|---|---|---|
| ApoAI | ApoAI–rHDL (method 1) | Thin film hydration of cargo–lipid complex followed by co-sonication with apoAI [169,170] | Conventional method of preparing rHDL |
| ApoAI–rHDL (method 2) | Thin film hydration of cargo–lipid–protein complex followed by cholate dialysis [46,162,163] | Preparation of rHDL containing phospholipids with lower Tm; presence of residual detergent a potential source of toxicity | |
| ApoAI–AuNP HDL (method 1) | Incubation of thiolated lipids and apoAI with AuNP [77,166] | Covalent conjugation of protein and lipids onto Au surface Circumvents apolipoprotein exchange | |
| ApoAI–AuNP HDL (method 2) | Nanoprecipitation to form Au–lipid NP, followed by co-sonication with apoAI [165] | Noncovalent interactions between protein and NP, allowing retention of native interactions | |
| ApoAI-coated HSA | Desolvation to form HSA NP, followed by conjugation to thiolated apoAI [133] | Non-lipidated formulation that allows trafficking of NP to the brain, possibly through SR-B1 on brain endothelial cells | |
| μHDL | Controlled mixing of lipids, payloads, and apoAI using a microfluidics device [164] | Suitable for scaled-up production of HDL as drug delivery vehicles | |
| ApoAII | ApoAII–poly(butyl cyanoacrylate (PBCA) | Incubation of apoAII with PBCA NP, with or without polysorbate 80 [49] | Lipid-free polymeric formulation of apoAII |
| ApoB-100 | ApoB-100-rLDL | Plasma-derived LDL used to reconstitute with therapeutic agents directly [50,51,52,53,54,55] | Templated approach that directly incorporates drug molecules with LDL |
| ApoB-100 coated HSA | Desolvation to form HSA NP, followed by conjugation to thiolated apoB-100 [133] | Non-lipidated formulation that allows trafficking of NP to the brain, possibly through LDLr and LRP1 on brain endothelial cells | |
| ApoB–AuNP HDL | Nanoprecipitation to form Au–lipid NP, followed by co-sonication with apoB-100 [61] | Noncovalent interactions between protein and NP, allowing retention of native interactions | |
| ApoB–PBCA | Incubation of apoB with PBCA NP, with or without polysorbate 80 [49] | Lipid-free polymeric formulation of apoB; capable of crossing the BBB | |
| Dextran coated LDL | Incubation of dextran with LDL [73] | Polymeric formulation of drug/LDL complex; decreases exchange with other serum apolipoproteins | |
| Dendrimer coated LDL | Conjugation of G5.0 PPI dendrimers to LDL via –OH to –NH2 linkage [74] | Dendrimer grafting allows drug loading and controlled release on the surface of LDL | |
| ApoCII | ApoCII–PBCA | Incubation of apoCII with PBCA NP, with or without polysorbate 80 [49] | Lipid free polymeric formulation of apoCII |
| ApoE3 | ApoE3–rHDL | Thin film hydration of cargo–lipid complex followed by co-sonication with apoE3 [18,39,63,171] | Conventional method of preparing rHDL |
| ApoE3–AuNP HDL | Thin film hydration of AuNP–lipid complex followed by co-sonication with apoE3 [64,172] | Noncovalent interactions between protein and NP, allowing retention of native interactions | |
| ApoE3 coated HSA | Desolvation to form HSA NP, followed by conjugation with apoE3 [130,131,132,134] | Non-lipidated formulation that allows trafficking of NP to the brain, possibly through LDLr and LRP1 on brain endothelial cells | |
| ApoE3–PBCA | Incubation of apoE3 with PBCA NP, with or without polysorbate 80 [49] | Lipid-free polymeric formulation of apoE3; capable of crossing the BBB | |
| ApoJ | ApoJ–PBCA | Incubation of apoJ with PBCA NP, with or without polysorbate 80 [49] | Lipid-free polymeric formulation of apoJ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chuang, S.T.; Cruz, S.; Narayanaswami, V. Reconfiguring Nature’s Cholesterol Accepting Lipoproteins as Nanoparticle Platforms for Transport and Delivery of Therapeutic and Imaging Agents. Nanomaterials 2020, 10, 906. https://0-doi-org.brum.beds.ac.uk/10.3390/nano10050906
Chuang ST, Cruz S, Narayanaswami V. Reconfiguring Nature’s Cholesterol Accepting Lipoproteins as Nanoparticle Platforms for Transport and Delivery of Therapeutic and Imaging Agents. Nanomaterials. 2020; 10(5):906. https://0-doi-org.brum.beds.ac.uk/10.3390/nano10050906
Chicago/Turabian StyleChuang, Skylar T., Siobanth Cruz, and Vasanthy Narayanaswami. 2020. "Reconfiguring Nature’s Cholesterol Accepting Lipoproteins as Nanoparticle Platforms for Transport and Delivery of Therapeutic and Imaging Agents" Nanomaterials 10, no. 5: 906. https://0-doi-org.brum.beds.ac.uk/10.3390/nano10050906