PEI-Mediated Transient Transfection of High Five Cells at Bioreactor Scale for HIV-1 VLP Production

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture Conditions
2.2. Construction of Plasmid DNA
2.3. Transient Gene Expression in Erlenmeyer Flask
2.4. Transient Gene Expression in Bioreactor
2.5. Flow Cytometry
2.6. Fluorescence Confocal Microscopy
2.7. HPLC Analyses
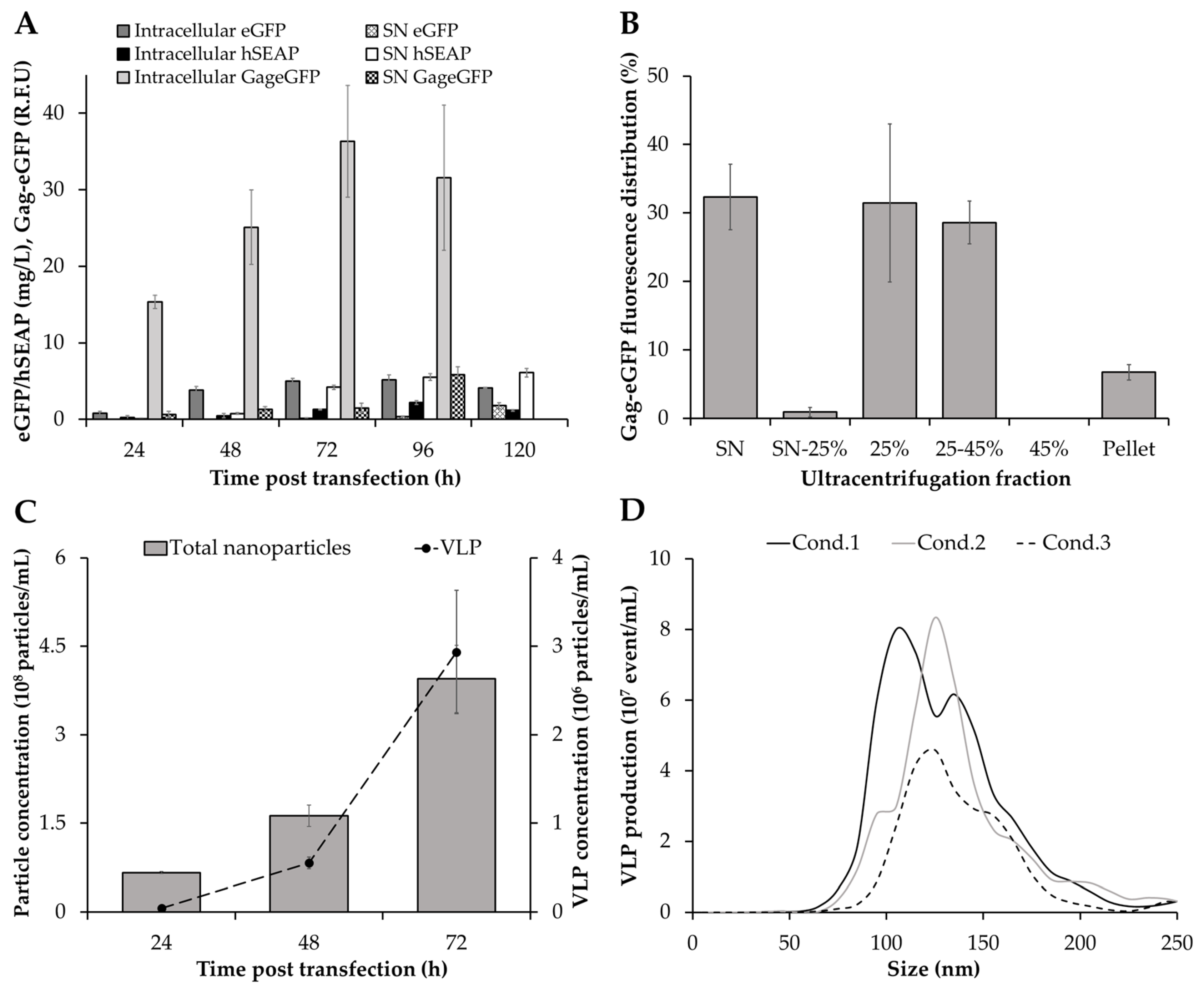
2.8. Analysis of Nanoparticle Production
2.8.1. Nanoparticle Tracking Analysis
2.8.2. Flow Virometry
2.9. eGFP/Gag-eGFP Measurement by Spectrofluorometry
2.10. hSEAP Quantification
2.11. Gag-eGFP Quantification using p24 Enzyme-Linked ImmunoSorbent Assay (ELISA)
2.12. Analytical Ultracentrifugation
2.13. Statistical Analyses
3. Results and Discussion
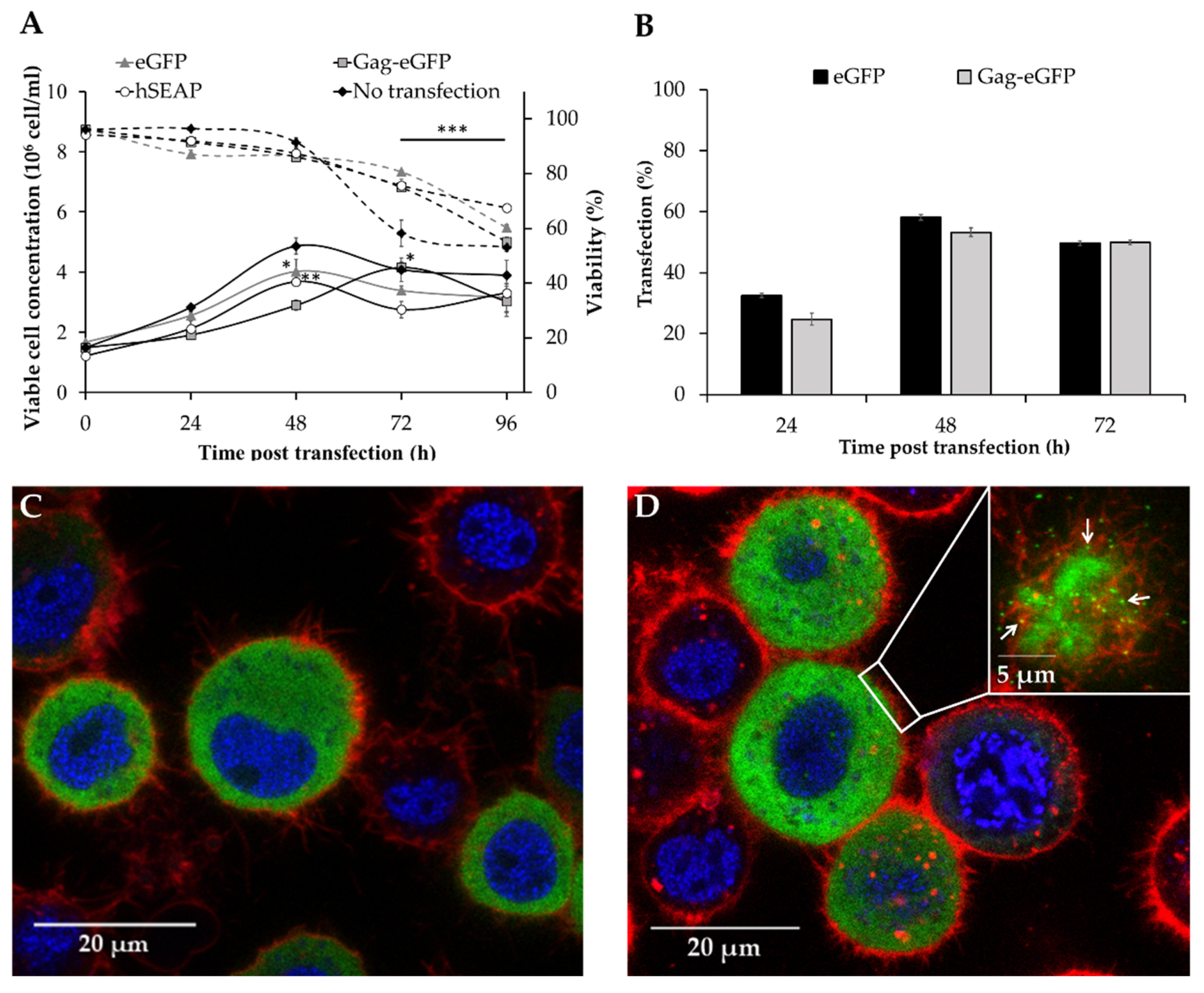
3.1. Production of Different Recombinant Products
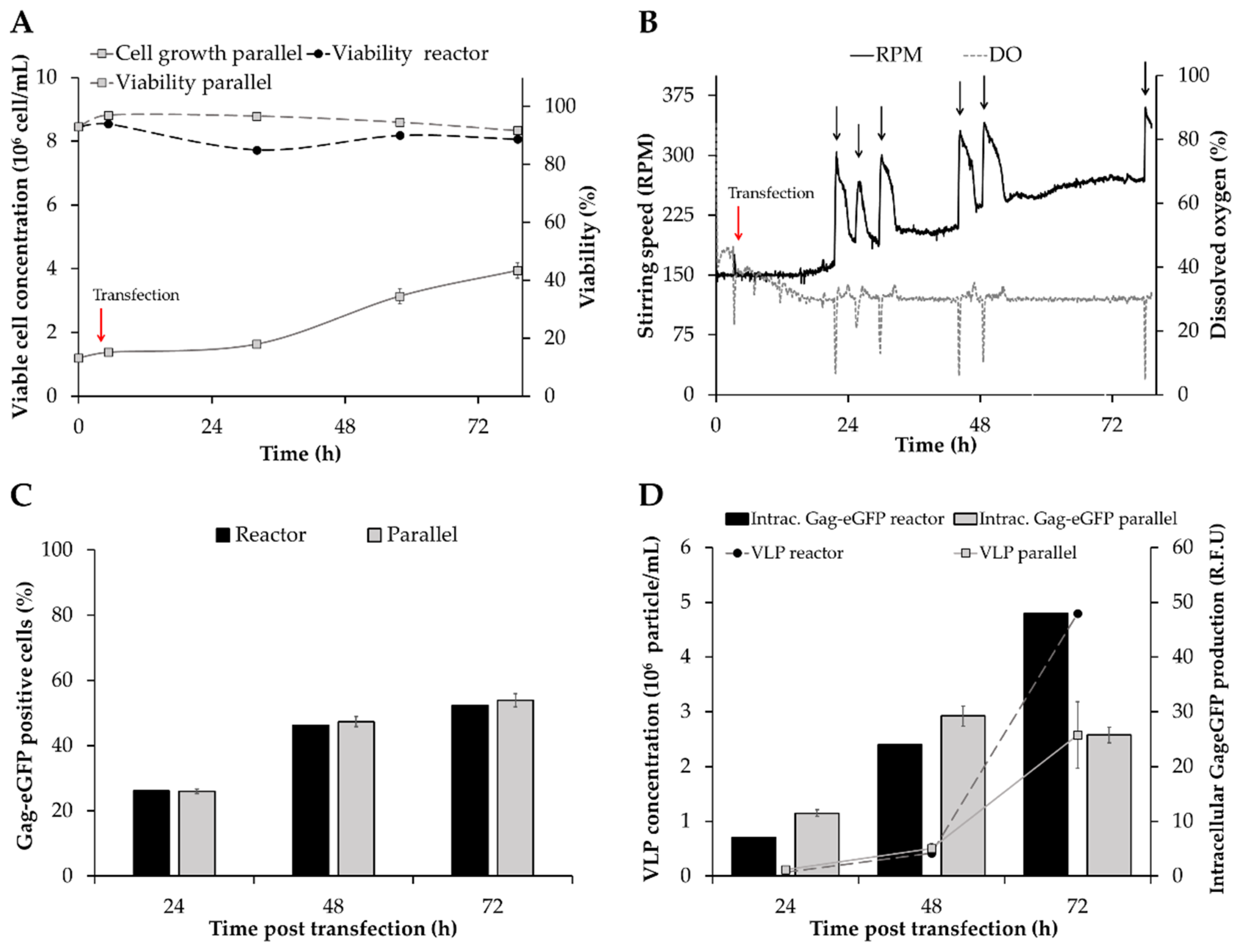
3.2. Transferability of VLP Production to Bioreactor
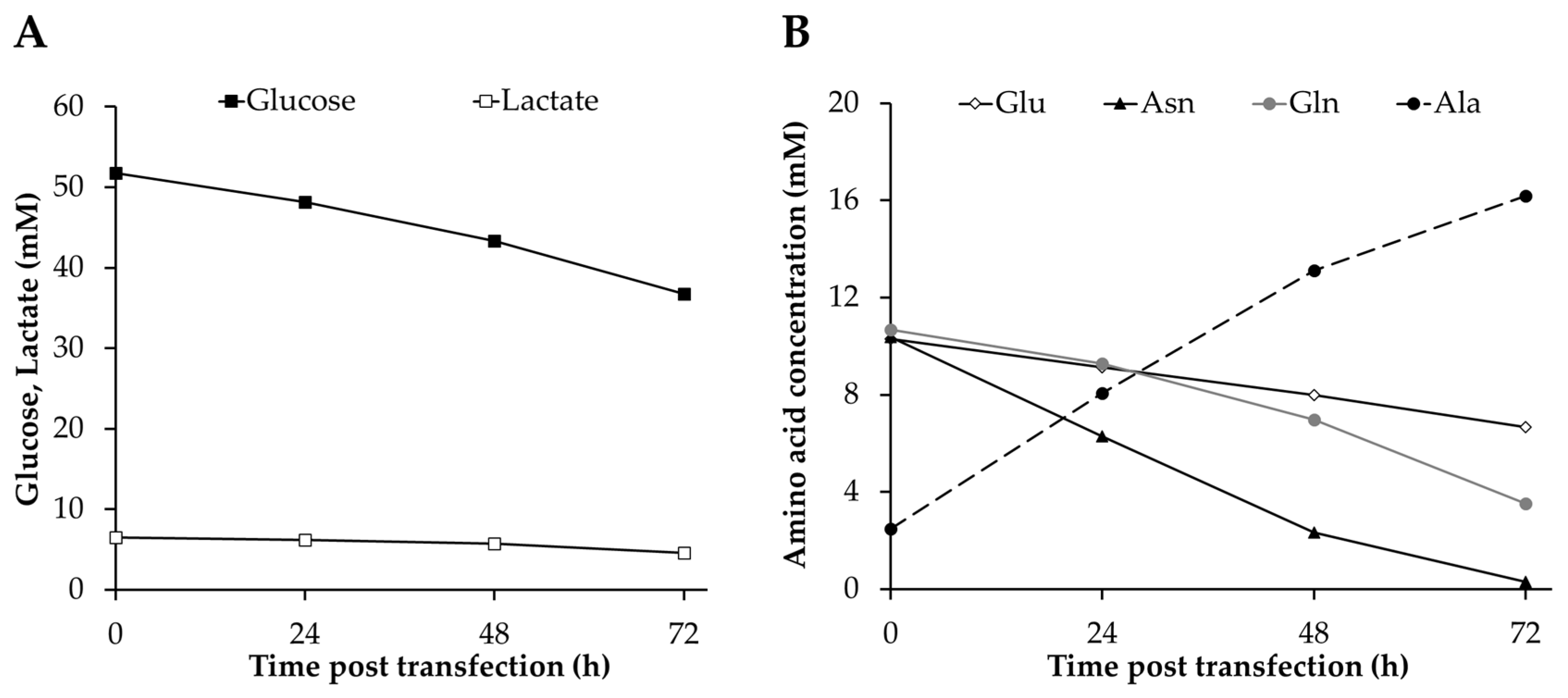
3.3. Analysis of Metabolites
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Guttieri, M.C.; Liang, M. Human antibody production using insect-cell expression systems. In Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2004; Volume 248, pp. 269–299. [Google Scholar]
- Brank, A.S.; Van Bemmel, D.M.; Christman, J.K. Optimization of Baculovirus-Mediated Expression and Purification of Hexahistidine-Tagged Murine DNA (Cytosine-C5)-Methyltransferase-1 in Spodoptera frugiperda 9 Cells. Protein Expr. Purif. 2002, 25, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Sarafanov, A.G.; Makogonenko, E.M.; Pechik, I.V.; Radtke, K.-P.; Khrenov, A.V.; Ananyeva, N.M.; Strickland, D.K.; Saenko, E.L. Identification of Coagulation Factor VIII A2 Domain Residues Forming the Binding Epitope for Low-Density Lipoprotein Receptor-Related Protein. Biochemistry 2006, 45, 1829–1840. [Google Scholar] [CrossRef] [PubMed]
- Cruz, P.E.; Cunha, A.; Peixoto, C.C.; Clemente, J.; Moreira, J.L.; Carrondo, M.J.T. Optimization of the production of virus-like particles in insect cells. Biotechnol. Bioeng. 1998, 60, 408–418. [Google Scholar] [CrossRef]
- Gheysen, D.; Jacobs, E.; de Foresta, F.; Thiriart, C.; Francotte, M.; Thines, D.; De Wilde, M. Assembly and release of HIV-1 precursor Pr55gag virus-like particles from recombinant baculovirus-infected insect cells. Cell 1989, 59, 103–112. [Google Scholar] [CrossRef]
- Mena, J.A.; Kamen, A.A. Insect cell technology is a versatile and robust vaccine manufacturing platform. Expert Rev. Vaccines 2011, 10, 1063–1081. [Google Scholar] [CrossRef]
- Roldão, A.; Mellado, M.C.M.; Castilho, L.R.; Carrondo, M.J.T.; Alves, P.M. Virus-like particles in vaccine development. Expert Rev. Vaccines 2010, 9, 1149–1176. [Google Scholar] [CrossRef]
- Nika, L.; Cuadrado-Castano, S.; Arunkumar, G.A.; Grünwald-Gruber, C.; McMahon, M.; Koczka, K.; García-Sastre, A.; Krammer, F.; Grabherr, R. An HER2-displaying virus-like particle vaccine protects from challenge with mammary carcinoma cells in a mouse model. Vaccines 2019, 7, 41. [Google Scholar] [CrossRef] [Green Version]
- Lua, L.H.L.; Connors, N.K.; Sainsbury, F.; Chuan, Y.P.; Wibowo, N.; Middelberg, A.P.J. Bioengineering virus-like particles as vaccines. Biotechnol. Bioeng. 2014, 111, 425–440. [Google Scholar] [CrossRef]
- Buonaguro, L.; Buonaguro, F.M.; Tornesello, M.L.; Mantas, D.; Beth-Giraldo, E.; Wagner, R.; Michelson, S.; Prevost, M.C.; Wolf, H.; Giraldo, G. High efficient production of Pr55gag virus-like particles expressing multiple HIV-1 epitopes, including a gp120 protein derived from an Ugandan HIV-1 isolate of subtype A. Antivir. Res. 2001, 49, 35–47. [Google Scholar] [CrossRef]
- Reiter, K.; Aguilar, P.P.; Grammelhofer, D.; Joseph, J.; Steppert, P.; Jungbauer, A. Separation of influenza virus-like particles from baculovirus by polymer grafted anion-exchanger. J. Sep. Sci. 2020, 43, 2270–2278. [Google Scholar] [CrossRef] [Green Version]
- Moleirinho, M.G.; Fernandes, R.P.; Carvalho, S.B.; Bezemer, S.; Detmers, F.; Hermans, P.; Silva, R.J.S.; Alves, P.M.; Carrondo, M.J.T.; Peixoto, C. Baculovirus affinity removal in viral-based bioprocesses. Sep. Purif. Technol. 2020, 241, 116693. [Google Scholar] [CrossRef]
- Geisse, S. Reflections on more than 10 years of TGE approaches. Protein Expr. Purif. 2009, 64, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, F.; Vidigal, J.; Dias, M.M.; Prather, K.L.J.; Coroadinha, A.S.; Teixeira, A.P.; Alves, P.M. Flipase-mediated cassette exchange in Sf9 insect cells for stable gene expression. Biotechnol. Bioeng. 2012, 109, 2836–2844. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Pitol, A.K.; Bachmann, V.; Hacker, D.L.; Baldi, L.; Wurm, F.M. A simple plasmid-based transient gene expression method using High Five cells. J. Biotechnol. 2015, 216, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Puente-Massaguer, E.; Lecina, M.; Gòdia, F. Nanoscale characterization coupled to multi-parametric optimization of Hi5 cell transient gene expression. Appl. Microbiol. Biotechnol. 2018, 102, 10495–10510. [Google Scholar] [CrossRef] [PubMed]
- Bleckmann, M.; Fritz, M.H.Y.; Bhuju, S.; Jarek, M.; Schürig, M.; Geffers, R.; Benes, V.; Besir, H.; Van Den Heuvel, J.; Li, Y. Genomic analysis and isolation of RNA polymerase II dependent promoters from Spodoptera frugiperda. PLoS ONE 2015, 10, e0132898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, X.; Hacker, D.L.; Baldi, L.; Wurm, F.M. Virus-free transient protein production in Sf9 cells. J. Biotechnol. 2013, 171, 61–70. [Google Scholar] [CrossRef]
- Mori, K.; Hamada, H.; Ogawa, T.; Ohmuro-Matsuyama, Y.; Katsuda, T.; Yamaji, H. Efficient production of antibody Fab fragment by transient gene expression in insect cells. J. Biosci. Bioeng. 2017, 124, 221–226. [Google Scholar] [CrossRef] [Green Version]
- Mizote, Y.; Masumi-Koizumi, K.; Katsuda, T.; Yamaji, H. Production of an antibody Fab fragment using 2A peptide in insect cells. J. Biosci. Bioeng. 2020, 130, 205–211. [Google Scholar] [CrossRef]
- Bleckmann, M.; Schürig, M.; Endres, M.; Samuels, A.; Gebauer, D.; Konisch, N.; van den Heuvel, J. Identifying parameters to improve the reproducibility of transient gene expression in High Five cells. PLoS ONE 2019, 14, e0217878. [Google Scholar] [CrossRef]
- Gutiérrez-Granados, S.; Cervera, L.; Kamen, A.A.; Gòdia, F. Advancements in mammalian cell transient gene expression (TGE) technology for accelerated production of biologics. Crit. Rev. Biotechnol. 2018, 38, 918–940. [Google Scholar] [CrossRef] [PubMed]
- Agathos, S.N. Insect cell bioreactors. Cytotechnology 1996, 20, 173–189. [Google Scholar] [CrossRef]
- Elias, C.B.; Jardin, B.; Kamen, A. Recombinant Protein Production in Large-Scale Agitated Bioreactors Using the Baculovirus Expression Vector System. In Baculovirus and Insect Cell Expression Protocols; Humana Press: Totowa, NJ, USA, 2007; pp. 225–245. [Google Scholar]
- Ghasemi, A.; Bozorg, A.; Rahmati, F.; Mirhassani, R.; Hosseininasab, S. Comprehensive study on Wave bioreactor system to scale up the cultivation of and recombinant protein expression in baculovirus-infected insect cells. Biochem. Eng. J. 2019, 143, 121–130. [Google Scholar] [CrossRef]
- Weber, W.; Weber, E.; Geisse, S.; Memmert, K. Optimisation of protein expression and establishment of the Wave Bioreactor for Baculovirus/insect cell culture. Cytotechnology 2002, 38, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Cronin, C.N. High-volume shake flask cultures as an alternative to cellbag technology for recombinant protein production in the baculovirus expression vector system (BEVS). Protein Expr. Purif. 2020, 165, 105496. [Google Scholar] [CrossRef] [PubMed]
- Hermida-Matsumoto, L.; Resh, M.D. Localization of Human Immunodeficiency Virus Type 1 Gag and Env at the Plasma Membrane by Confocal Imaging. J. Virol. 2000, 74, 8670–8679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pappenreiter, M.; Sissolak, B.; Sommeregger, W.; Striedner, G. Oxygen uptake rate soft-sensing via dynamic kl a computation: Cell volume and metabolic transition prediction in mammalian bioprocesses. Front. Bioeng. Biotechnol. 2019, 7, 195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puente-Massaguer, E.; Gòdia, F.; Lecina, M. Development of a non-viral platform for rapid virus-like particle production in Sf9 cells. J. Biotechnol. 2020, 322, 43–53. [Google Scholar] [CrossRef]
- Gutiérrez-Granados, S.; Cervera, L.; Gòdia, F.; Carrillo, J.; Segura, M.M. Development and validation of a quantitation assay for fluorescently tagged HIV-1 virus-like particles. J. Virol. Methods 2013, 193, 85–95. [Google Scholar] [CrossRef]
- Reiter, K.; Aguilar, P.P.; Wetter, V.; Steppert, P.; Tover, A.; Jungbauer, A. Separation of virus-like particles and extracellular vesicles by flow-through and heparin affinity chromatography. J. Chromatogr. A 2019, 1588, 77–84. [Google Scholar] [CrossRef]
- Puente-Massaguer, E.; Lecina, M.; Gòdia, F. Application of advanced quantification techniques in nanoparticle-based vaccine development with the Sf9 cell baculovirus expression system. Vaccine 2020, 38, 1849–1859. [Google Scholar] [CrossRef] [PubMed]
- Puente-Massaguer, E.; Lecina, M.; Gòdia, F. Integrating nanoparticle quantification and statistical design of experiments for efficient HIV-1 virus-like particle production in High Five cells. Appl. Microbiol. Biotechnol. 2020, 104, 1569–1582. [Google Scholar] [CrossRef] [PubMed]
- Cervera, L.; Gutiérrez-Granados, S.; Martínez, M.; Blanco, J.; Gòdia, F.; Segura, M.M. Generation of HIV-1 Gag VLPs by transient transfection of HEK 293 suspension cell cultures using an optimized animal-derived component free medium. J. Biotechnol. 2013, 166, 152–165. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Granados, S.; Cervera, L.; Segura, M.d.l.M.; Wölfel, J.; Gòdia, F. Optimized production of HIV-1 virus-like particles by transient transfection in CAP-T cells. Appl. Microbiol. Biotechnol. 2016, 100, 3935–3947. [Google Scholar] [CrossRef] [PubMed]
- Cervera, L.; González-Domínguez, I.; Segura, M.M.; Gòdia, F. Intracellular characterization of Gag VLP production by transient transfection of HEK 293 cells. Biotechnol. Bioeng. 2017, 114, 2507–2517. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, B.; Vidigal, J.; Correia, R.; Carrondo, M.J.T.; Alves, P.M.; Teixeira, A.P.; Roldão, A. Adaptive laboratory evolution of stable insect cell lines for improved HIV-Gag VLPs production. J. Biotechnol. 2020, 307, 139–147. [Google Scholar] [CrossRef]
- Van Der Pol, E.; Van Gemert, M.J.C.; Sturk, A.; Nieuwland, R.; Van Leeuwen, T.G. Single vs. swarm detection of microparticles and exosomes by flow cytometry. J. Thromb. Haemost. 2012, 10, 919–930. [Google Scholar] [CrossRef]
- González-Domínguez, I.; Puente-Massaguer, E.; Cervera, L.; Gòdia, F. Quality Assessment of Virus-Like Particles at Single Particle Level: A Comparative Study. Viruses 2020, 12, 223. [Google Scholar] [CrossRef] [Green Version]
- Théry, C.; Zitvogel, L.; Amigorena, S. Exosomes: Composition, biogenesis and function. Nat. Rev. Immunol. 2002, 2, 569–579. [Google Scholar] [CrossRef]
- Li, J.; Chen, X.; Yi, J.; Liu, Y.; Li, D.; Wang, J.; Hou, D.; Jiang, X.; Zhang, J.; Wang, J.; et al. Identification and characterization of 293t cell-derived exosomes by profiling the protein, mRNA and microrna components. PLoS ONE 2016, 11, e0163043. [Google Scholar] [CrossRef]
- Puente-Massaguer, E.; Saccardo, P.; Ferrer-Miralles, N.; Lecina, M.; Gòdia, F.; Ferrer-Miralles, N. Coupling Microscopy and Flow Cytometry for a Comprehensive Characterization of Nanoparticle Production in Insect Cells. Cytom. Part A 2020. [Google Scholar] [CrossRef] [PubMed]
- Steppert, P.; Burgstaller, D.; Klausberger, M.; Berger, E.; Aguilar, P.P.; Schneider, T.A.; Kramberger, P.; Tover, A.; Nöbauer, K.; Razzazi-Fazeli, E.; et al. Purification of HIV-1 gag virus-like particles and separation of other extracellular particles. J. Chromatogr. A 2016, 1455, 93–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pham, P.L.; Kamen, A.; Durocher, Y. Large-scale transfection of mammalian cells for the fast production of recombinant protein. Mol. Biotechnol. 2006, 34, 225–237. [Google Scholar] [CrossRef]
- Rhiel, M.; Mitchell-Logean, C.M.; Murhammer, D.W. Comparison of Trichoplusia ni BTI-Tn-5b1-4 (high five(TM) and Spodoptera frugiperda Sf-9 insect cell line metabolism in suspension cultures. Biotechnol. Bioeng. 1997, 55, 909–920. [Google Scholar] [CrossRef]
- Weidner, T.; Druzinec, D.; Mühlmann, M.; Buchholz, R.; Czermak, P. The components of shear stress affecting insect cells used with the baculovirus expression vector system. Z. Naturforsch. C 2017, 72, 429–439. [Google Scholar] [CrossRef] [PubMed]
- Wickham, T.J.; Nemerow, G.R. Optimization of Growth Methods and Recombinant Protein Production in BTI-Tn-5B1-4 Insect Cells Using the Baculovirus Expression System. Biotechnol. Prog. 1993, 9, 25–30. [Google Scholar] [CrossRef]
- Fuenmayor, J.; Cervera, L.; Gòdia, F.; Kamen, A. Extended gene expression for Gag VLP production achieved at bioreactor scale. J. Chem. Technol. Biotechnol. 2019, 94, 302–308. [Google Scholar] [CrossRef] [Green Version]
- Strobl, F.; Ghorbanpour, S.M.; Palmberger, D.; Striedner, G. Evaluation of screening platforms for virus-like particle production with the baculovirus expression vector system in insect cells. Sci. Rep. 2020, 10, 1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monteiro, F.; Bernal, V.; Saelens, X.; Lozano, A.B.; Bernal, C.; Sevilla, A.; Carrondo, M.J.T.; Alves, P.M. Metabolic profiling of insect cell lines: Unveiling cell line determinants behind system’s productivity. Biotechnol. Bioeng. 2014, 111, 816–828. [Google Scholar] [CrossRef]
- Drugmand, J.C.; Schneider, Y.J.; Agathos, S.N. Insect cells as factories for biomanufacturing. Biotechnol. Adv. 2012, 30, 1140–1157. [Google Scholar] [CrossRef] [Green Version]
- Monteiro, F.; Bernal, V.; Alves, P.M. The role of host cell physiology in the productivity of the baculovirus-insect cell system: Fluxome analysis of Trichoplusia ni and Spodoptera frugiperda cell lines. Biotechnol. Bioeng. 2017, 114, 674–684. [Google Scholar] [CrossRef] [PubMed]
- Elias, C.B.; Carpentier, E.; Durocher, Y.; Bisson, L.; Wagner, R.; Kamen, A. Improving glucose and glutamine metabolism of human HEK 293 and Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase enzyme. Biotechnol. Prog. 2003, 19, 90–97. [Google Scholar] [CrossRef]
- Benslimane, C.; Elias, C.B.; Hawari, J.; Kamen, A. Insights into the central metabolism of Spodoptera frugiperda (Sf-9) and Trichoplusia ni BTI-Tn-5B1-4 (Tn-5) insect cells by radiolabeling studies. Biotechnol. Prog. 2005, 21, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Sugiura, T.; Amann, E. Properties of two insect cell lines useful for the baculovirus expression system in serum-free culture. Biotechnol. Bioeng. 2000, 51, 494–499. [Google Scholar] [CrossRef]
- Drews, M.; Paalme, T.; Vilu, R. The growth and nutrient utilization of the insect cell line Spodoptera frugiperda Sf9 in batch and continuous culture. J. Biotechnol. 1995, 40, 187–198. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Quantification Method | Fluorescent Particles/mL | Total Particles/mL | Supernatant | Intracellular |
|---|---|---|---|---|
| NTA (particles/mL) | 3.6 ± 1.0· | 2.4 ± 0.3· | - | - |
| Flow virometry (particles/mL) | 2.9 ± 0.7· | 4.0 ± 0.6· | - | - |
| ELISA (ng/mL) | - | - | 17.1 | 238.4 |
| Fluorometry (R.F.U.) | 1.4 ± 0.6·a | - | 1.5 ± 0.6 | 36.3 ± 7.3 |
| Metabolite | Time Post Transfection (h) | ||
|---|---|---|---|
| 0–24 | 24–48 | 48–72 | |
| Glucose | −90.3 | −83.0 | −75.6 |
| Lactate | −8.4 | −7.8 | −13.1 |
| Phosphate | −48.5 | −53.2 | −17.6 |
| Aspartic acid | 25.8 | 7.1 | −5.3 |
| Glutamic acid | −29.0 | −19.9 | −15.0 |
| Asparagine | −103.8 | −68.0 | −23.4 |
| Serine | 30.0 | −0.3 | 1.8 |
| Glutamine | −35.4 | −39.8 | −39.8 |
| Histidine | −1.1 | −2.2 | −2.0 |
| Glycine | −6.2 | −7.8 | −2.8 |
| Threonine | 1.5 | −0.8 | −2.0 |
| Arginine | −0.7 | −5.1 | −1.7 |
| Alanine | 140.7 | 86.9 | 35.3 |
| Tyrosine | 0.3 | −1.7 | −3.0 |
| Valine | 0.2 | −4.5 | −5.7 |
| Methionine | −5.6 | −6.2 | −5.2 |
| Tryptophan | −0.1 | −1.4 | −0.9 |
| Phenylalanine | −2.0 | −3.7 | −3.6 |
| Isoleucine | −1.2 | −4.8 | −4.4 |
| Leucine | −0.9 | −6.1 | −5.9 |
| Lysine | −8.0 | −17.9 | −5.9 |
| Proline | −2.6 | −5.1 | −4.6 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puente-Massaguer, E.; Strobl, F.; Grabherr, R.; Striedner, G.; Lecina, M.; Gòdia, F. PEI-Mediated Transient Transfection of High Five Cells at Bioreactor Scale for HIV-1 VLP Production. Nanomaterials 2020, 10, 1580. https://0-doi-org.brum.beds.ac.uk/10.3390/nano10081580
Puente-Massaguer E, Strobl F, Grabherr R, Striedner G, Lecina M, Gòdia F. PEI-Mediated Transient Transfection of High Five Cells at Bioreactor Scale for HIV-1 VLP Production. Nanomaterials. 2020; 10(8):1580. https://0-doi-org.brum.beds.ac.uk/10.3390/nano10081580
Chicago/Turabian StylePuente-Massaguer, Eduard, Florian Strobl, Reingard Grabherr, Gerald Striedner, Martí Lecina, and Francesc Gòdia. 2020. "PEI-Mediated Transient Transfection of High Five Cells at Bioreactor Scale for HIV-1 VLP Production" Nanomaterials 10, no. 8: 1580. https://0-doi-org.brum.beds.ac.uk/10.3390/nano10081580