Assessment of the Influence of Crystalline Form on Cyto-Genotoxic and Inflammatory Effects Induced by TiO2 Nanoparticles on Human Bronchial and Alveolar Cells

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Nanoparticles
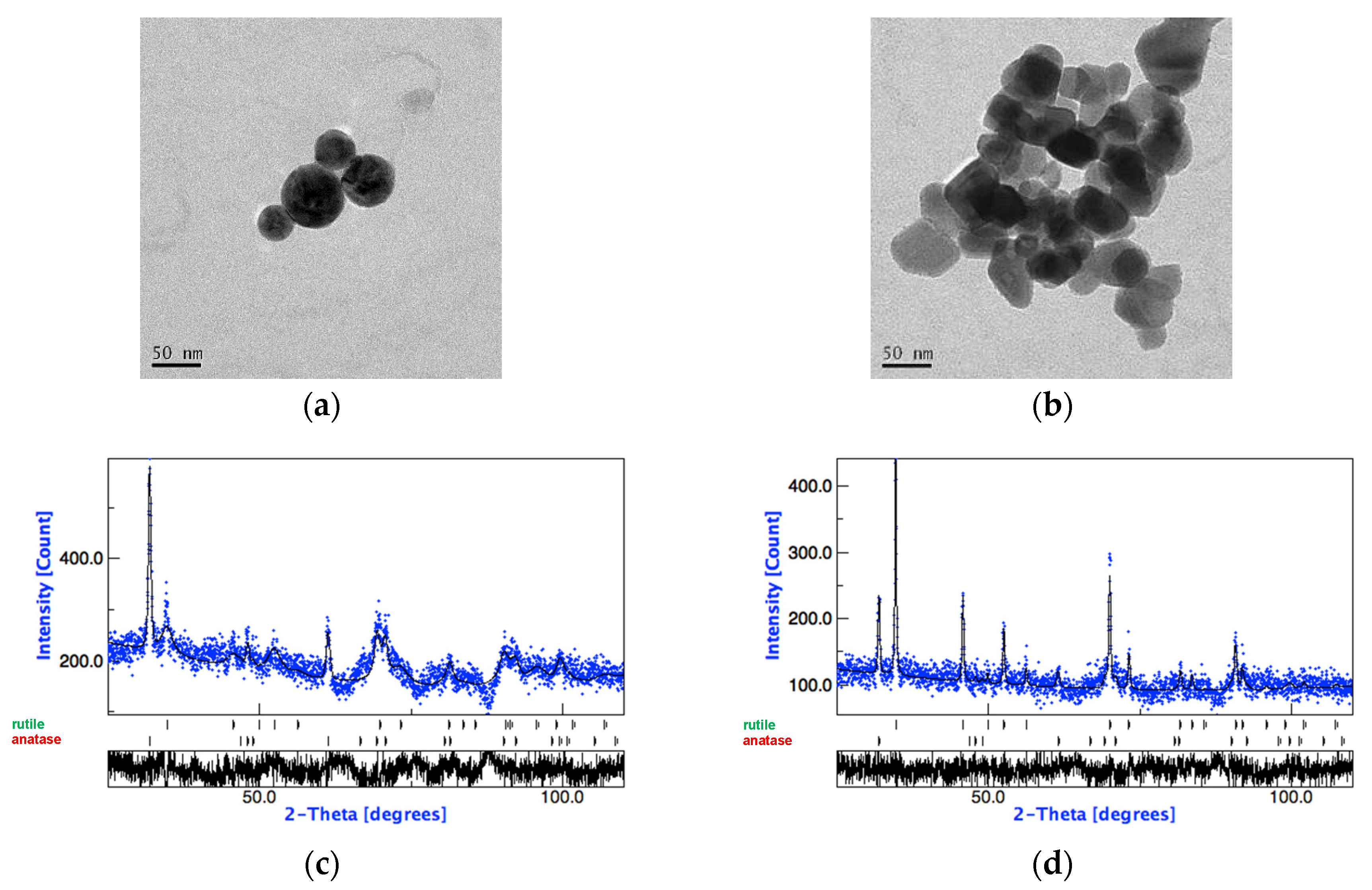
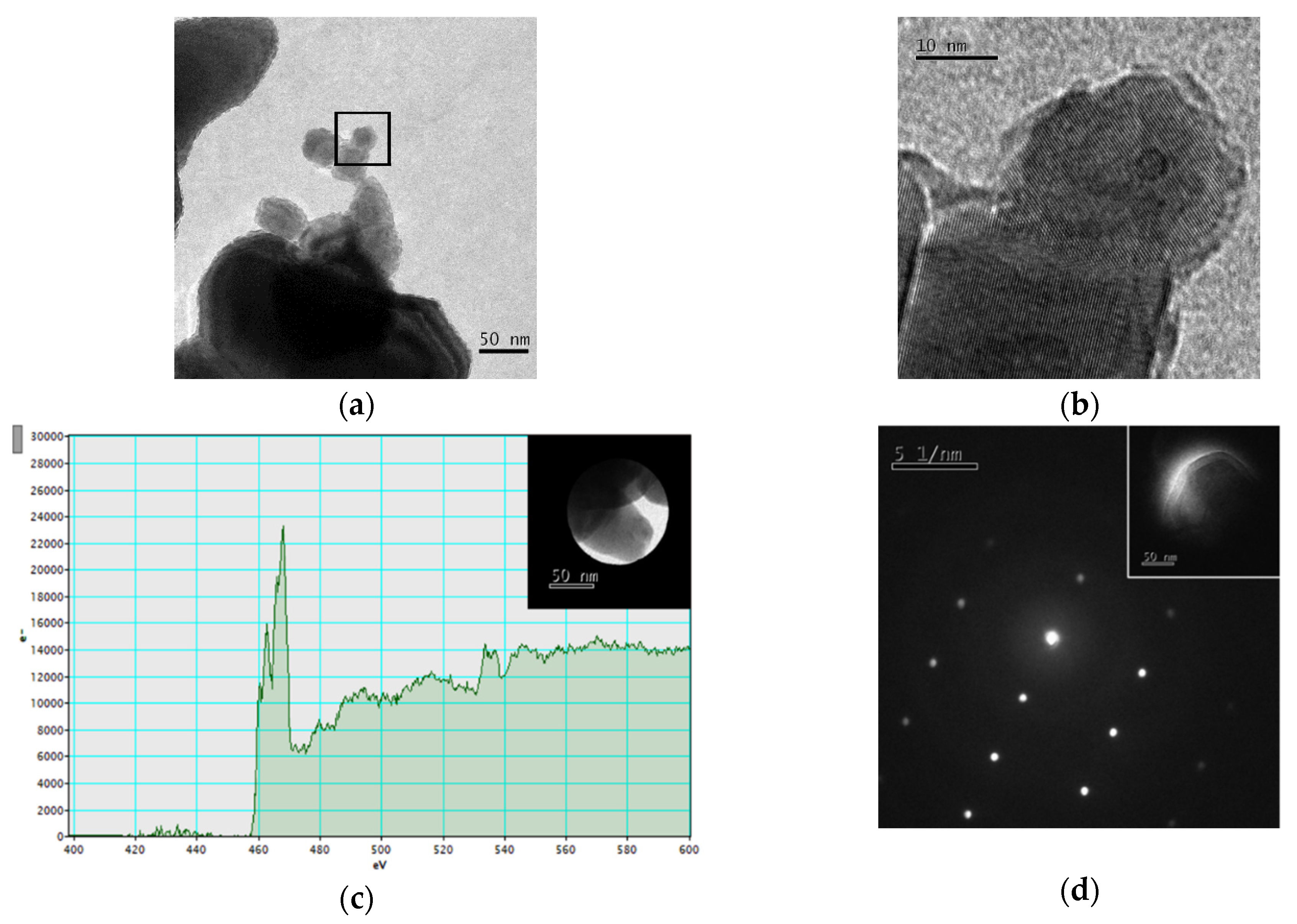
2.2. Nanoparticles Characterization
2.3. Cell Culture
2.4. Exposure
2.5. Cell Viability
2.6. Membrane Damage
2.7. DNA Damage
2.8. Detection of Cytokines
2.9. Statistical Analysis
3. Results
3.1. Nanoparticle Characterization
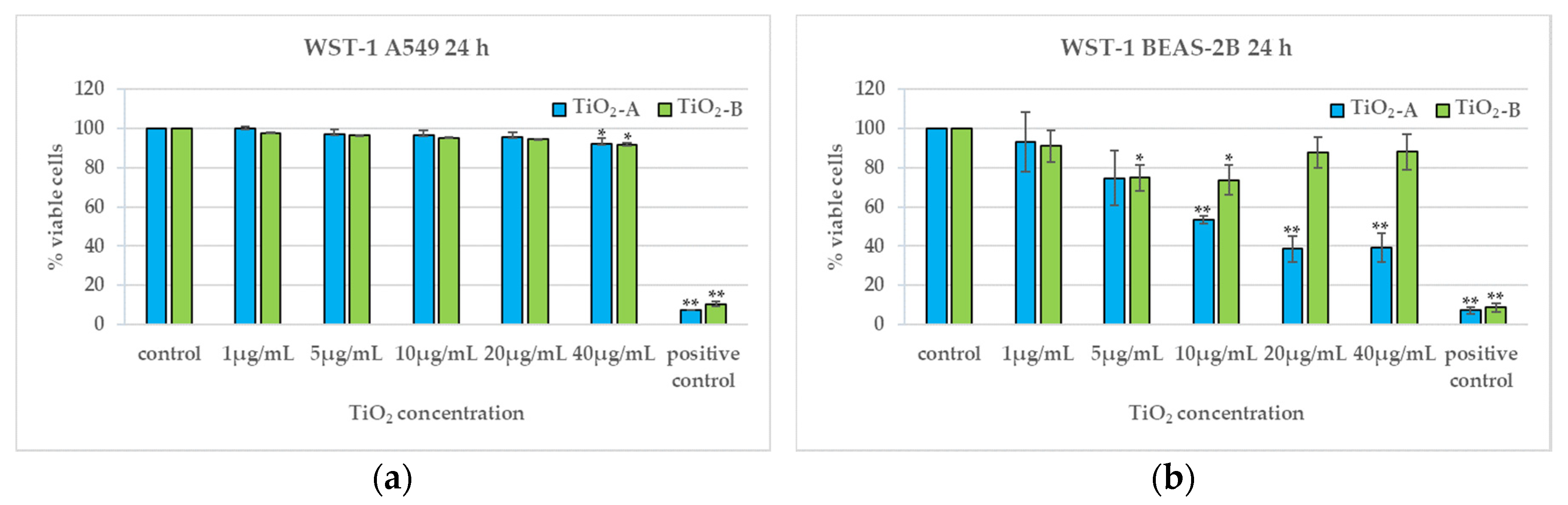
3.2. Cell Viability (24 h)
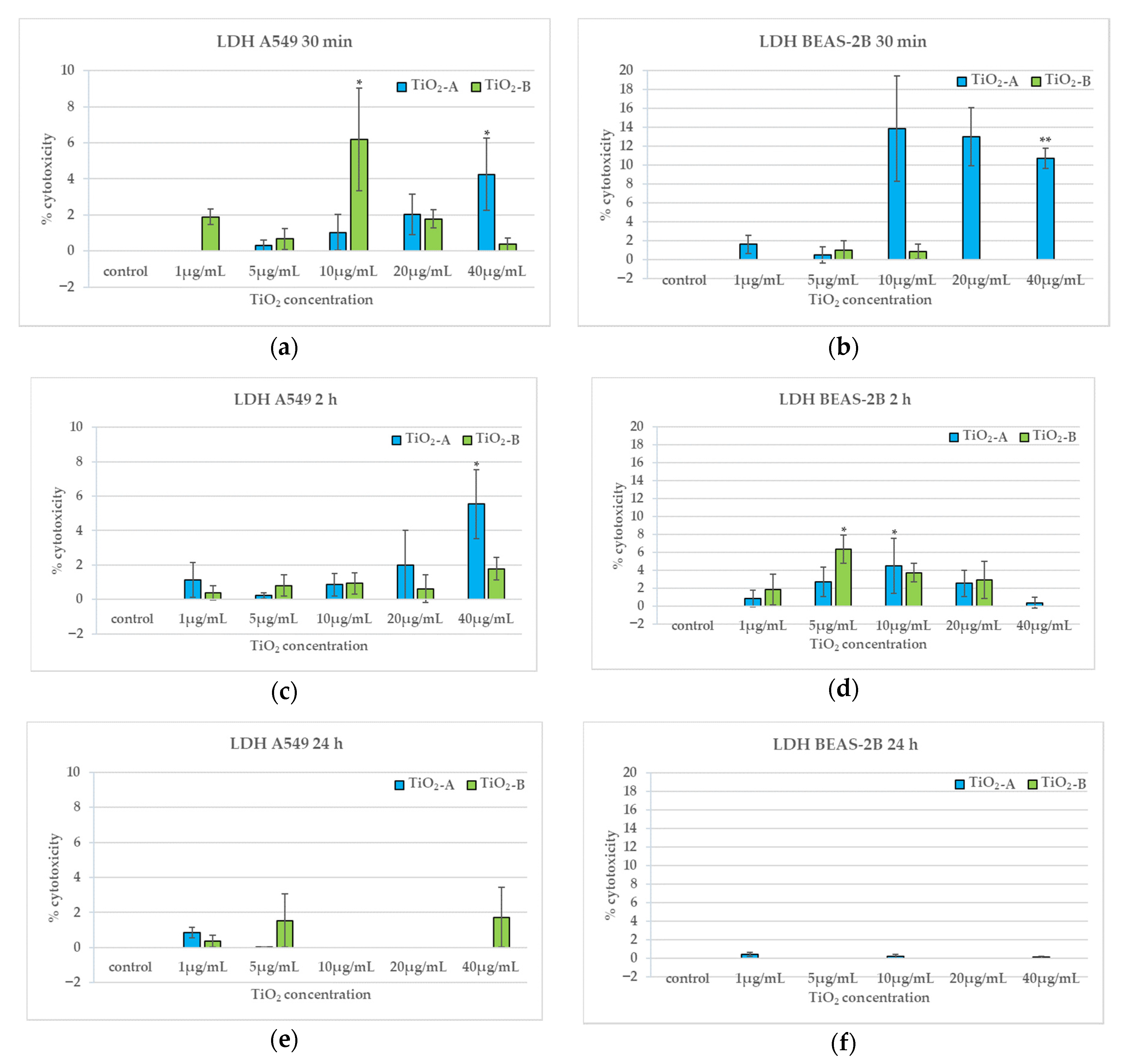
3.3. Membrane Integrity (30 min, 2 h and 24 h)
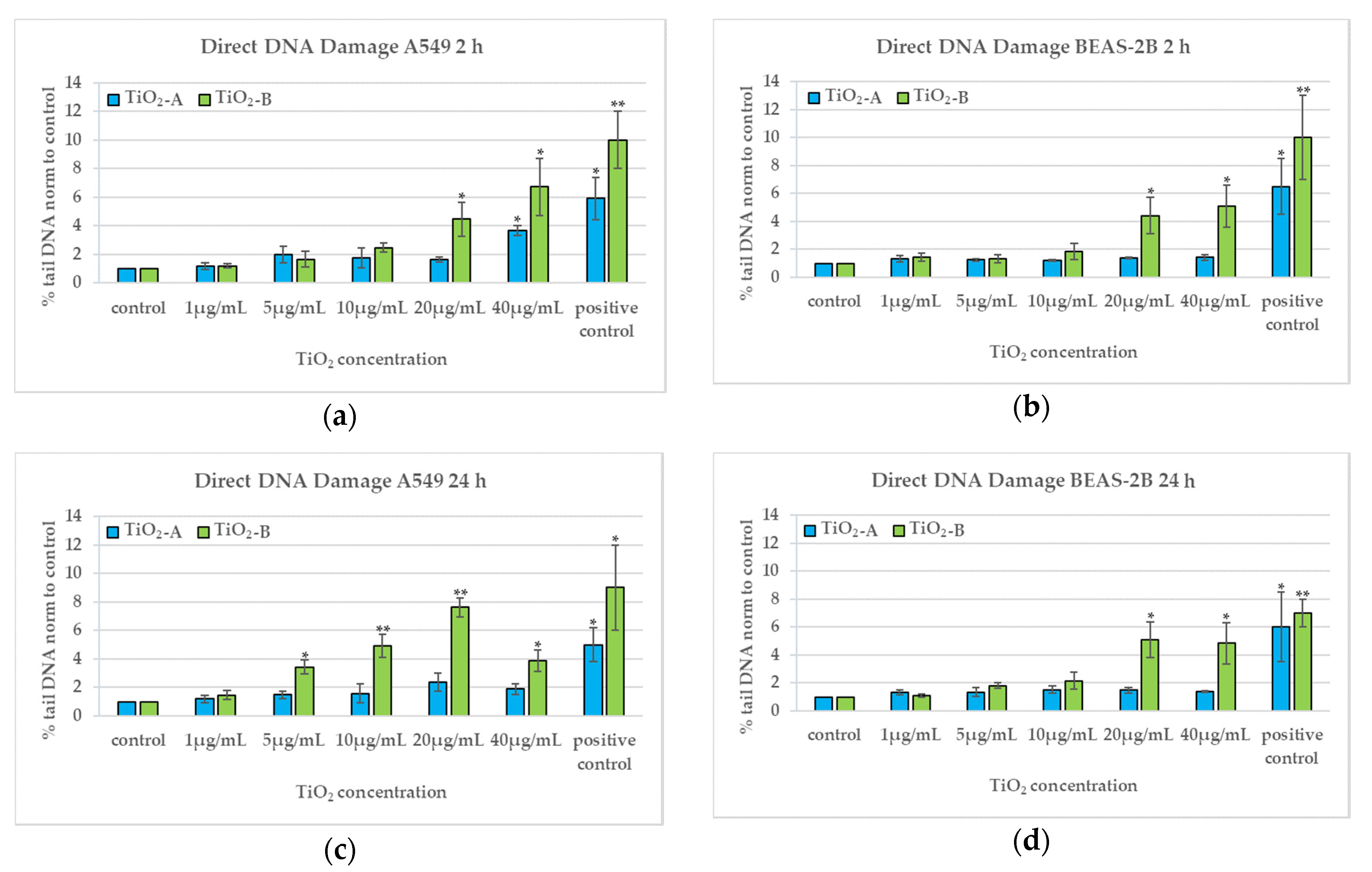
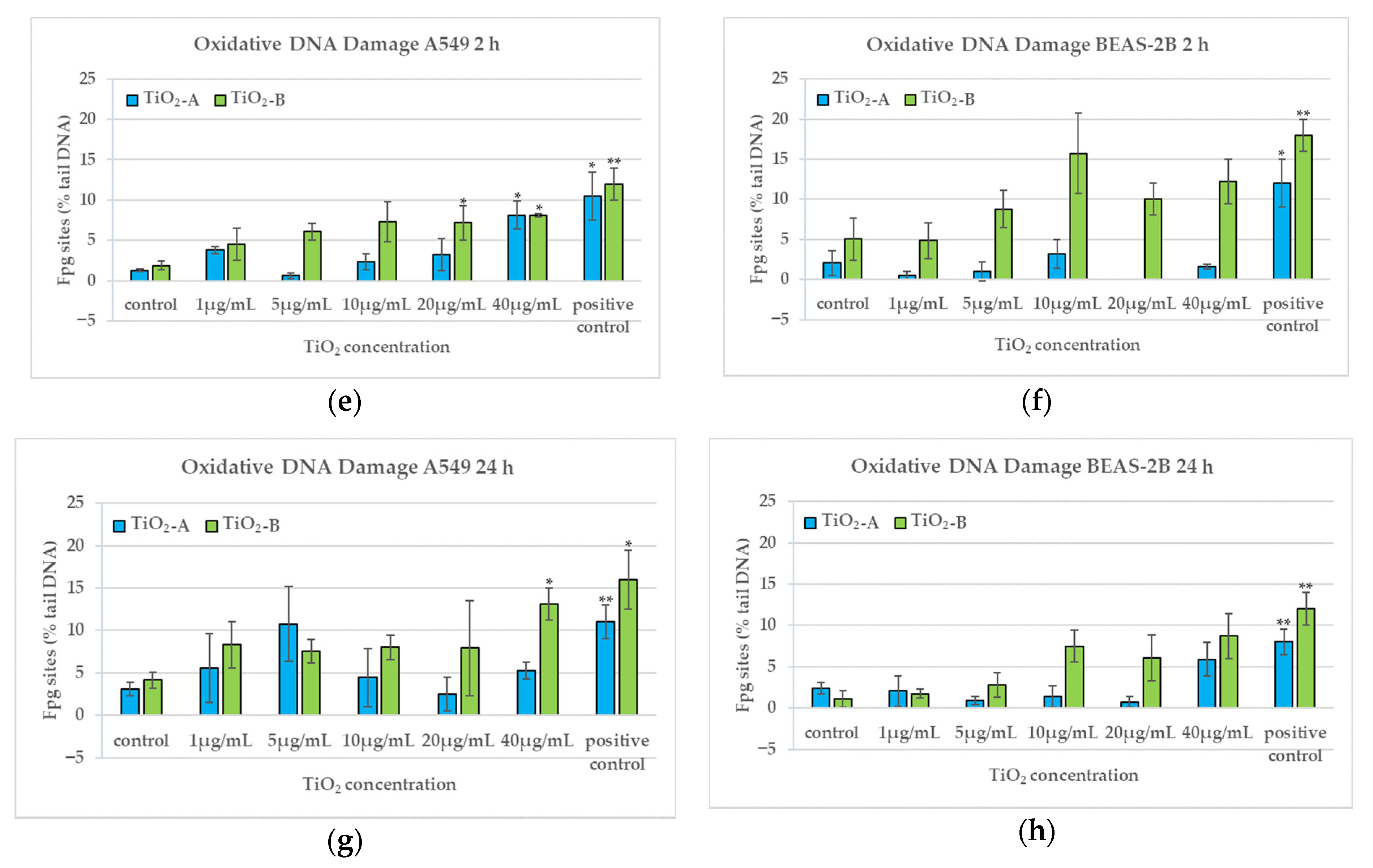
3.4. DNA Damage (2 h and 24 h)
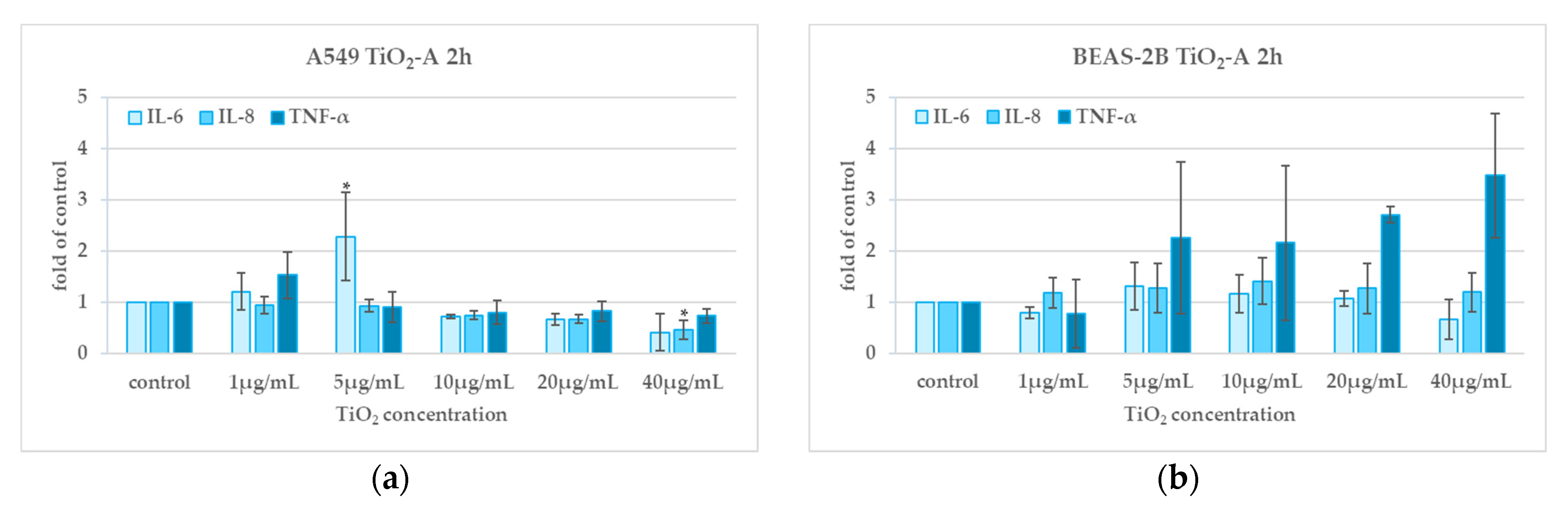
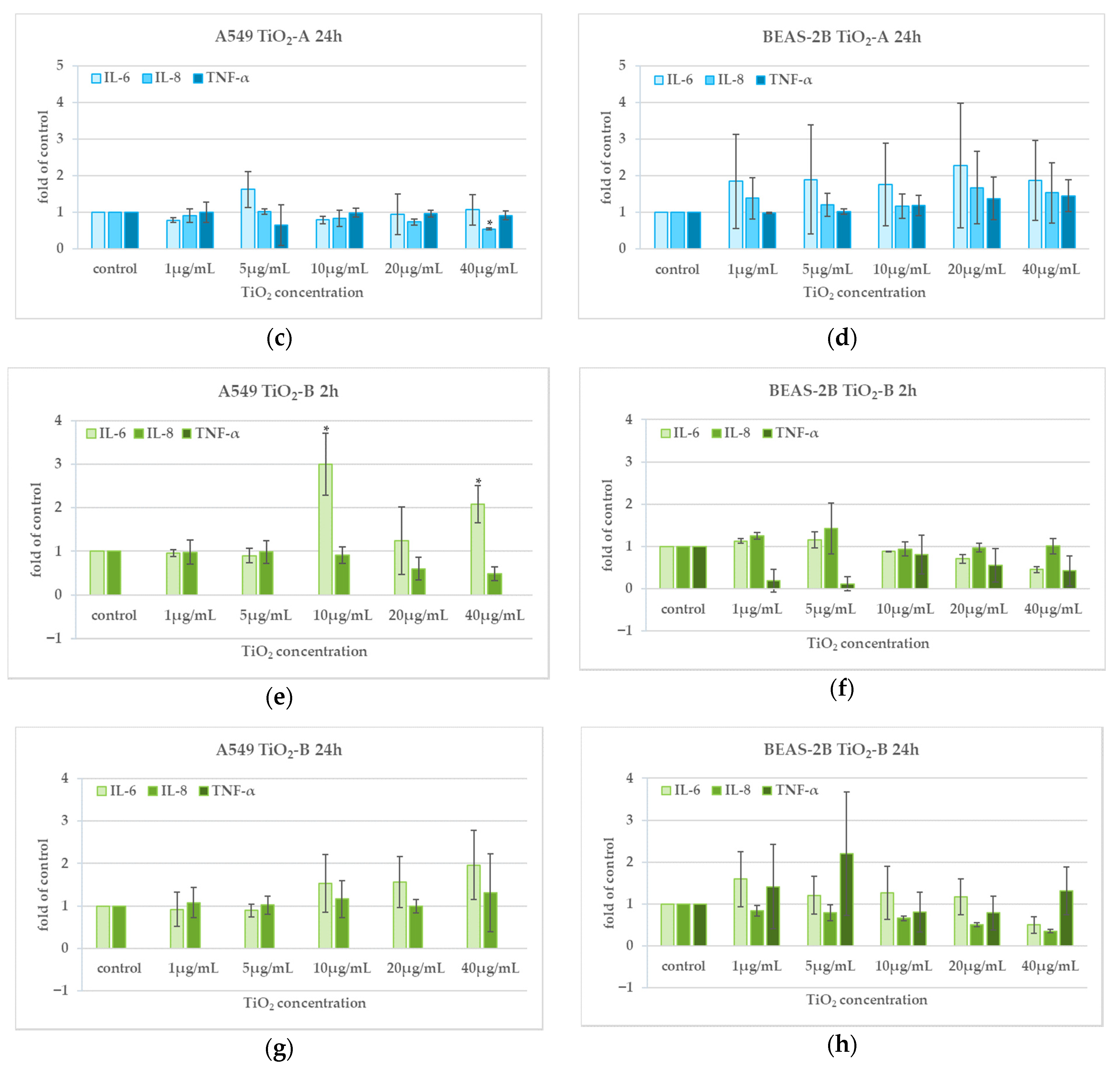
3.5. Cytokine Release (2 h and 24 h)
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, X.; Li, W.; Yang, Z. Toxicology of nanosized titanium dioxide: An update. Arch. Toxicol. 2015, 89, 2207–2217. [Google Scholar] [CrossRef] [PubMed]
- Doak, S.H.; Dusinska, M. NanoGenotoxicology: Present and the future. Mutagenesis 2017, 32, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weir, A.; Westerhoff, P.; Fabricius, L.; Hristovski, K.; von Goetz, N. Titanium dioxide nanoparticles in food and personal care products. Environ. Sci. Technol. 2012, 46, 2242–2250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharya, K.; Davoren, M.; Boertz, J.; Schins, R.P.; Hoffmann, E.; Dopp, E. Titanium dioxide nanoparticles induce oxidative stress and DNA-adduct formation but not DNA-breakage in human lung cells. Part. Fibre Toxicol. 2009, 6, 17. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Zhu, Y.; Chen, Z.; Xu, H.; Zhou, J.; Tang, S.; Xu, Z.; Kong, F.; Li, X.; Zhang, Y.; et al. Cardiopulmonary effects induced by occupational exposure to titanium dioxide nanoparticles. Nanotoxicology 2018, 12, 169–184. [Google Scholar] [CrossRef]
- Song, B.; Zhou, T.; Yang, W.; Liu, J.; Shao, L. Contribution of oxidative stress to TiO2 nanoparticle-induced toxicity. Environ. Toxicol. Pharmacol. 2016, 48, 130–140. [Google Scholar] [CrossRef]
- Chen, T.; Yan, J.; Li, Y. Genotoxicity of titanium dioxide nanoparticles. J. Food Drug Anal. 2014, 22, 95–104. [Google Scholar] [CrossRef]
- Carbon black, titanium dioxide, and talc. IARC Monogr Eval Carcinog Risks Hum. 2010, 93, 1–413.
- Ziental, D.; Czarczynska-Goslinska, B.; Mlynarczyk, D.T.; Glowacka-Sobotta, A.; Stanisz, B.; Goslinski, T.; Sobotta, L. Titanium Dioxide Nanoparticles: Prospects and Applications in Medicine. Nanomaterials 2020, 10, 387. [Google Scholar] [CrossRef] [Green Version]
- Ursini, C.L.; Cavallo, D.; Fresegna, A.M.; Ciervo, A.; Maiello, R.; Tassone, P.; Buresti, G.; Casciardi, S.; Iavicoli, S. Evaluation of cytotoxic, genotoxic and inflammatory response in human alveolar and bronchial epithelial cells exposed to titanium dioxide nanoparticles. J. Appl. Toxicol. 2014, 34, 1209–1219. [Google Scholar] [CrossRef] [PubMed]
- Brandao, F.; Fernandez-Bertolez, N.; Rosario, F.; Bessa, M.J.; Fraga, S.; Pasaro, E.; Teixeira, J.P.; Laffon, B.; Valdiglesias, V.; Costa, C. Genotoxicity of TiO2 Nanoparticles in Four Different Human Cell Lines (A549, HEPG2, A172 and SH-SY5Y). Nanomaterials 2020, 10, 412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tolliver, L.M.; Holl, N.J.; Hou, F.Y.S.; Lee, H.J.; Cambre, M.H.; Huang, Y.W. Differential Cytotoxicity Induced by Transition Metal Oxide Nanoparticles is a Function of Cell Killing and Suppression of Cell Proliferation. Int. J. Mol. Sci. 2020, 21, 1731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baranowska-Wojcik, E.; Szwajgier, D.; Oleszczuk, P.; Winiarska-Mieczan, A. Effects of Titanium Dioxide Nanoparticles Exposure on Human Health-a Review. Biol. Trace Elem. Res. 2020, 193, 118–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salou, S.; Cirtiu, C.M.; Lariviere, D.; Fleury, N. Assessment of strategies for the formation of stable suspensions of titanium dioxide nanoparticles in aqueous media suitable for the analysis of biological fluids. Anal. Bioanal Chem. 2020, 412, 1469–1481. [Google Scholar] [CrossRef]
- Rosario, F.; Bessa, M.J.; Brandao, F.; Costa, C.; Lopes, C.B.; Estrada, A.C.; Tavares, D.S.; Teixeira, J.P.; Reis, A.T. Unravelling the Potential Cytotoxic Effects of Metal Oxide Nanoparticles and Metal(Loid) Mixtures on A549 Human Cell Line. Nanomaterials 2020, 10, 447. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Miura, N.; Hojo, R.; Yanagiba, Y.; Suda, M.; Hasegawa, T.; Miyagawa, M.; Wang, R.S. Genotoxicity assessment of titanium dioxide nanoparticle accumulation of 90 days in the liver of gpt delta transgenic mice. Genes Environ. 2020, 42, 7. [Google Scholar] [CrossRef] [Green Version]
- Kazimirova, A.; El Yamani, N.; Rubio, L.; Garcia-Rodriguez, A.; Barancokova, M.; Marcos, R.; Dusinska, M. Effects of Titanium Dioxide Nanoparticles on the Hprt Gene Mutations in V79 Hamster Cells. Nanomaterials 2020, 10, 465. [Google Scholar] [CrossRef] [Green Version]
- Oh, N.; Park, J.H. Endocytosis and exocytosis of nanoparticles in mammalian cells. Int. J. Nanomed. 2014, 9 (Suppl. 1), 51–63. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Zhou, D.; Han, S.; Zhou, S.; Jia, G. Hepatotoxicity and the role of the gut-liver axis in rats after oral administration of titanium dioxide nanoparticles. Part. Fibre Toxicol. 2019, 16, 48. [Google Scholar] [CrossRef]
- Biola-Clier, M.; Gaillard, J.C.; Rabilloud, T.; Armengaud, J.; Carriere, M. Titanium Dioxide Nanoparticles Alter the Cellular Phosphoproteome in A549 Cells. Nanomaterials 2020, 10, 185. [Google Scholar] [CrossRef] [Green Version]
- Liou, S.H.; Wu, W.T.; Liao, H.Y.; Chen, C.Y.; Tsai, C.Y.; Jung, W.T.; Lee, H.L. Global DNA methylation and oxidative stress biomarkers in workers exposed to metal oxide nanoparticles. J. Hazard. Mater. 2017, 331, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Danielsen, P.H.; Knudsen, K.B.; Strancar, J.; Umek, P.; Koklic, T.; Garvas, M.; Vanhala, E.; Savukoski, S.; Ding, Y.; Madsen, A.M.; et al. Effects of physicochemical properties of TiO2 nanomaterials for pulmonary inflammation, acute phase response and alveolar proteinosis in intratracheally exposed mice. Toxicol. Appl. Pharmacol. 2020, 386, 114830. [Google Scholar] [CrossRef] [PubMed]
- Pelclova, D.; Navratil, T.; Kacerova, T.; Zamostna, B.; Fenclova, Z.; Vlckova, S.; Kacer, P. NanoTiO2 Sunscreen Does Not Prevent Systemic Oxidative Stress Caused by UV Radiation and a Minor Amount of NanoTiO2 is Absorbed in Humans. Nanomaterials 2019, 9, 888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uboldi, C.; Urban, P.; Gilliland, D.; Bajak, E.; Valsami-Jones, E.; Ponti, J.; Rossi, F. Role of the crystalline form of titanium dioxide nanoparticles: Rutile, and not anatase, induces toxic effects in Balb/3T3 mouse fibroblasts. Toxicol. In Vitro 2016, 31, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Vandebriel, R.J.; Vermeulen, J.P.; van Engelen, L.B.; de Jong, B.; Verhagen, L.M.; de la Fonteyne-Blankestijn, L.J.; Hoonakker, M.E.; de Jong, W.H. The crystal structure of titanium dioxide nanoparticles influences immune activity in vitro and in vivo. Part. Fibre Toxicol. 2018, 15, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Q.; Wang, H.; Peng, Q.; Li, Y.; Liu, Z.; Li, M. Different toxicity of anatase and rutile TiO2 nanoparticles on macrophages: Involvement of difference in affinity to proteins and phospholipids. J. Hazard. Mater. 2017, 335, 125–134. [Google Scholar] [CrossRef]
- Ghosh, M.; Oner, D.; Duca, R.C.; Cokic, S.M.; Seys, S.; Kerkhofs, S.; Van Landuyt, K.; Hoet, P.; Godderis, L. Cyto-genotoxic and DNA methylation changes induced by different crystal phases of TiO2-np in bronchial epithelial (16-HBE) cells. Mutat Res. 2017, 796, 1–12. [Google Scholar] [CrossRef]
- Ursini, C.L.; Cavallo, D.; Fresegna, A.M.; Ciervo, A.; Maiello, R.; Buresti, G.; Casciardi, S.; Bellucci, S.; Iavicoli, S. Differences in cytotoxic, genotoxic, and inflammatory response of bronchial and alveolar human lung epithelial cells to pristine and COOH– functionalized multiwalled carbon nanotubes. Biomed. Res. Int. 2014, 2014, 359506. [Google Scholar] [CrossRef]
- Ursini, C.L.; Maiello, R.; Ciervo, A.; Fresegna, A.M.; Buresti, G.; Superti, F.; Marchetti, M.; Iavicoli, S.; Cavallo, D. Evaluation of uptake, cytotoxicity and inflammatory effects in respiratory cells exposed to pristine and –OH and –COOH functionalized multi-wall carbon nanotubes. J. Appl. Toxicol. 2016, 36, 394–403. [Google Scholar] [CrossRef] [Green Version]
- Cavallo, D.; Ciervo, A.; Fresegna, A.M.; Maiello, R.; Tassone, P.; Buresti, G.; Casciardi, S.; Iavicoli, S.; Ursini, C.L. Investigation on cobalt-oxide nanoparticles cyto-genotoxicity and inflammatory response in two types of respiratory cells. J. Appl. Toxicol. 2015, 35, 1102–1113. [Google Scholar] [CrossRef]
- Rietveld, H.M. Line profiles of neutron powder-diffraction peaks for structure refinement. Acta Crystallogr. 1967, 22, 151–152. [Google Scholar] [CrossRef]
- Rietveld, H.M. A profile refinement method for nuclear and magnetic structures. J. Appl. Crystallogr. 1969, 2, 65–71. [Google Scholar] [CrossRef]
- Wilhelmi, V.; Fischer, U.; van Berlo, D.; Schulze-Osthoff, K.; Schins, R.P.; Albrecht, C. Evaluation of apoptosis induced by nanoparticles and fine particles in RAW 264.7 macrophages: Facts and artefacts. Toxicol. In Vitro 2012, 26, 323–334. [Google Scholar] [CrossRef]
- Guadagnini, R.; Halamoda Kenzaoui, B.; Walker, L.; Pojana, G.; Magdolenova, Z.; Bilanicova, D.; Saunders, M.; Juillerat-Jeanneret, L.; Marcomini, A.; Huk, A.; et al. Toxicity screenings of nanomaterials: Challenges due to interference with assay processes and components of classic in vitro tests. Nanotoxicology 2015, 9 (Suppl. 1), 13–24. [Google Scholar] [CrossRef]
- Collins, A.; El Yamani, N.; Dusinska, M. Sensitive detection of DNA oxidation damage induced by nanomaterials. Free Radic Biol. Med. 2017, 107, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Ursini, C.L.; Cavallo, D.; Fresegna, A.M.; Ciervo, A.; Maiello, R.; Buresti, G.; Casciardi, S.; Tombolini, F.; Bellucci, S.; Iavicoli, S. Comparative cyto-genotoxicity assessment of functionalized and pristine multiwalled carbon nanotubes on human lung epithelial cells. Toxicol. In Vitro 2012, 26, 831–840. [Google Scholar] [CrossRef] [PubMed]
- Petersen, E.J.; Reipa, V.; Watson, S.S.; Stanley, D.L.; Rabb, S.A.; Nelson, B.C. DNA damaging potential of photoactivated p25 titanium dioxide nanoparticles. Chem. Res. Toxicol. 2014, 27, 1877–1884. [Google Scholar] [CrossRef]
- Veranth, J.M.; Cutler, N.S.; Kaser, E.G.; Reilly, C.A.; Yost, G.S. Effects of cell type and culture media on Interleukin-6 secretion in response to environmental particles. Toxicol. In Vitro 2008, 22, 498–509. [Google Scholar] [CrossRef] [Green Version]
- Teeguarden, J.G.; Hinderliter, P.M.; Orr, G.; Thrall, B.D.; Pounds, J.G. Particokinetics in vitro: Dosimetry considerations for in vitro nanoparticle toxicity assessments. Toxicol. Sci. 2007, 95, 300–312. [Google Scholar] [CrossRef]
- Skuland, T.; Ovrevik, J.; Lag, M.; Refsnes, M. Role of size and surface area for pro-inflammatory responses to silica nanoparticles in epithelial lung cells: Importance of exposure conditions. Toxicol. In Vitro 2014, 28, 146–155. [Google Scholar] [CrossRef]
- Fenoglio, I.; Greco, G.; Livraghi, S.; Fubini, B. Non-UV-induced radical reactions at the surface of TiO2 nanoparticles that may trigger toxic responses. Chemistry 2009, 15, 4614–4621. [Google Scholar] [CrossRef] [PubMed]
- Magdolenova, Z.; Collins, A.; Kumar, A.; Dhawan, A.; Stone, V.; Dusinska, M. Mechanisms of genotoxicity. A review of in vitro and in vivo studies with engineered nanoparticles. Nanotoxicology 2014, 8, 233–278. [Google Scholar] [CrossRef] [PubMed]
- Sayes, C.M.; Wahi, R.; Kurian, P.A.; Liu, Y.; West, J.L.; Ausman, K.D.; Warheit, D.B.; Colvin, V.L. Correlating nanoscale titania structure with toxicity: A cytotoxicity and inflammatory response study with human dermal fibroblasts and human lung epithelial cells. Toxicol. Sci. 2006, 92, 174–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Matteis, V.; Cascione, M.; Brunetti, V.; Toma, C.C.; Rinaldi, R. Toxicity assessment of anatase and rutile titanium dioxide nanoparticles: The role of degradation in different pH conditions and light exposure. Toxicol. In Vitro 2016, 37, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Falck, G.C.; Lindberg, H.K.; Suhonen, S.; Vippola, M.; Vanhala, E.; Catalan, J.; Savolainen, K.; Norppa, H. Genotoxic effects of nanosized and fine TiO2. Hum. Exp. Toxicol. 2009, 28, 339–352. [Google Scholar] [CrossRef]
- Ursini, C.L.; Campopiano, A.; Fresegna, A.M.; Ciervo, A.; Cannizzaro, A.; Angelosanto, F.; Maiello, R.; Iavicoli, S.; Cavallo, D. Alkaline earth silicate (AES) wools: Evaluation of potential cyto-genotoxic and inflammatory effects on human respiratory cells. Toxicol. In Vitro 2019, 59, 228–237. [Google Scholar] [CrossRef]
- Jugan, M.L.; Barillet, S.; Simon-Deckers, A.; Herlin-Boime, N.; Sauvaigo, S.; Douki, T.; Carriere, M. Titanium dioxide nanoparticles exhibit genotoxicity and impair DNA repair activity in A549 cells. Nanotoxicology 2012, 6, 501–513. [Google Scholar] [CrossRef]
- Gualtieri, M.; Ovrevik, J.; Holme, J.A.; Perrone, M.G.; Bolzacchini, E.; Schwarze, P.E.; Camatini, M. Differences in cytotoxicity versus pro-inflammatory potency of different PM fractions in human epithelial lung cells. Toxicol. In Vitro 2010, 24, 29–39. [Google Scholar] [CrossRef]
- Park, E.J.; Yi, J.; Chung, K.H.; Ryu, D.Y.; Choi, J.; Park, K. Oxidative stress and apoptosis induced by titanium dioxide nanoparticles in cultured BEAS-2B cells. Toxicol. Lett. 2008, 180, 222–229. [Google Scholar] [CrossRef]
- Shi, Y.; Wang, F.; He, J.; Yadav, S.; Wang, H. Titanium dioxide nanoparticles cause apoptosis in BEAS-2B cells through the caspase 8/t-Bid-independent mitochondrial pathway. Toxicol. Lett. 2010, 196, 21–27. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Physicochemical Characteristics | TiO2-A Code 677469 | TiO2-B Code 634662 | Testing Method 1 |
|---|---|---|---|
| diameter (nm) | mean 43.8 ± 17.0 range 13.5–90.6 | mean 76.6 ± 52.7 range 22.3–252.9 | TEM |
| crystal structure | anatase (79%) rutile (21%) | rutile (81%) anatase (19%) | XRD |
| SSA (m2/g) | 14.9 | 13.7 | BET |
| shape | spherical | irregular–tetragonal | TEM |
| Physicochemical characteristics of dispersed NPs in water (20 µg/mL) | TiO2-A Code 677469 | TiO2-B Code 634662 | Testing Method 1 |
| Z Potential (mV) | −32.2 | −30.1 | ELS |
| agglomerate/aggregate size (diam Zav nm) | 139.8 (t0) 130.5 (t24) | 366.0 (t0) 195.0 (t24) | DLS |
| Physicochemical characteristics of dispersed NPs in RPMI with 10% FBS | TiO2-A Code 677469 | TiO2-B Code 634662 | Testing Method 1 |
| Z Potential (mV) (20 µg/mL) | −9.13 | −10.3 | ELS |
| agglomerate/aggregate size (diam Zav nm) (20 µg/mL) | 196.5 (t0) 197.6 (t24) | 368.0 (t0) | DLS |
| agglomerate/aggregate size (diam Zav nm) (100 µg/mL) | 151.0 (t0) | 447.0 (t0) | TEM |
| Physicochemical characteristics of dispersed NPs in BEGM | TiO2-A Code 677469 | TiO2-B Code 634662 | Testing Method 1 |
| Z Potential (mV) (20 µg/mL) | −11.7 | −11.0 | ELS |
| agglomerate/aggregate size (diam Zav nm) (20 µg/mL) | 220.2 (t0) 224.6 (t24) | 428.0 (t0) 282.0 (t24) | DLS |
| agglomerate/aggregate size (diam Zav nm) (100 µg/mL) | 186.0 (t0) | 456.0 (t0) | TEM |
| Biological Endpoint | TiO2-A | TiO2-B | ||
|---|---|---|---|---|
| A549 | BEAS-2B | A549 | BEAS-2B | |
| Cell viability reduction | + | +++ | + | + |
| Membrane damage | ||||
| 30 min | ++ | + | + | − |
| 2 h | + | + | − | + |
| 24 h | − | − | − | − |
| Direct DNA damage | ||||
| 2 h | + | − | +++ | +++ |
| 24 h | − | − | +++ | +++ |
| Oxidative DNA damage | ||||
| 2 h | + | − | +++ | − |
| 24 h | − | − | + | − |
| IL-6 release | ||||
| 2 h | + | − | + | − |
| 24 h | − | − | − | − |
| IL-8 release | ||||
| 2 h | + | − | − | − |
| 24 h | + | − | − | − |
| TNF-α release | ||||
| 2 h | − | − | − | − |
| 24 h | − | − | − | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fresegna, A.M.; Ursini, C.L.; Ciervo, A.; Maiello, R.; Casciardi, S.; Iavicoli, S.; Cavallo, D. Assessment of the Influence of Crystalline Form on Cyto-Genotoxic and Inflammatory Effects Induced by TiO2 Nanoparticles on Human Bronchial and Alveolar Cells. Nanomaterials 2021, 11, 253. https://0-doi-org.brum.beds.ac.uk/10.3390/nano11010253
Fresegna AM, Ursini CL, Ciervo A, Maiello R, Casciardi S, Iavicoli S, Cavallo D. Assessment of the Influence of Crystalline Form on Cyto-Genotoxic and Inflammatory Effects Induced by TiO2 Nanoparticles on Human Bronchial and Alveolar Cells. Nanomaterials. 2021; 11(1):253. https://0-doi-org.brum.beds.ac.uk/10.3390/nano11010253
Chicago/Turabian StyleFresegna, Anna Maria, Cinzia Lucia Ursini, Aureliano Ciervo, Raffaele Maiello, Stefano Casciardi, Sergio Iavicoli, and Delia Cavallo. 2021. "Assessment of the Influence of Crystalline Form on Cyto-Genotoxic and Inflammatory Effects Induced by TiO2 Nanoparticles on Human Bronchial and Alveolar Cells" Nanomaterials 11, no. 1: 253. https://0-doi-org.brum.beds.ac.uk/10.3390/nano11010253