
Long Non-Coding RNAs: The New Frontier into Understanding the Etiology of Alcohol Use Disorder

 ,
,
Abstract
:1. The RNA World
2. lncRNA Functions
3. Epidemiology of AUD
4. Metabolism of AUD
5. Neuropathology of AUD
6. Genetics of AUD
7. lncRNAs Functions in AUD
8. Epigenetics of AUD
9. Complexity of Comorbidities Associated with AUD
10. Therapeutic Methods Utilizing lncRNAs: A New Frontier
11. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Naidoo, N.; Pawitan, Y.; Soong, R.; Cooper, D.N.; Ku, C.-S. Human genetics and genomics a decade after the release of the draft sequence of the human genome. Hum. Genet. 2011, 5, 577. [Google Scholar] [CrossRef]
- The ENCODE Project Consortium. An integrated encyclopedia of DNA elements in the human genome. Nature 2012, 489, 57–74. [Google Scholar] [CrossRef]
- Pennisi, E. Human Genome Is Much More than Just Genes | Science | AAAS. Science. 2012. Available online: https://www.science.org/content/article/human-genome-much-more-just-genes (accessed on 24 June 2022).
- Evans, J.R.; Feng, F.Y.; Chinnaiyan, A.M. The Bright Side of Dark Matter: LncRNAs in Cancer. J. Clin. Investig. 2016, 126, 2775–2782. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Sood, A.K.; Dang, C.V.; Zhang, L. The Role of Long Noncoding RNAs in Cancer: The Dark Matter Matters. Curr. Opin. Genet. Dev. 2018, 48, 8–15. [Google Scholar] [CrossRef]
- Dahariya, S.; Paddibhatla, I.; Kumar, S.; Raghuwanshi, S.; Pallepati, A.; Gutti, R.K. Long Non-Coding RNA: Classification, Biogenesis and Functions in Blood Cells. Mol. Immunol. 2019, 112, 82–92. [Google Scholar] [CrossRef]
- Ponting, C.P.; Oliver, P.L.; Reik, W. Evolution and Functions of Long Noncoding RNAs. Cell 2009, 136, 629–641. [Google Scholar] [CrossRef] [Green Version]
- Nie, J.-H.; Li, T.-X.; Zhang, X.-Q.; Liu, J. Roles of Non-Coding RNAs in Normal Human Brain Development, Brain Tumor, and Neuropsychiatric Disorders. Non-Coding RNA 2019, 5, 36. [Google Scholar] [CrossRef] [Green Version]
- Spadaro, P.A.; Bredy, T.W. Emerging Role of Non-Coding RNA in Neural Plasticity, Cognitive Function, and Neuropsychiatric Disorders. Front. Gene. 2012, 3, 132. [Google Scholar] [CrossRef] [Green Version]
- Derrien, T.; Johnson, R.; Bussotti, G.; Tanzer, A.; Djebali, S.; Tilgner, H.; Guernec, G.; Martin, D.; Merkel, A.; Knowles, D.G.; et al. The GENCODE v7 Catalog of Human Long Noncoding RNAs: Analysis of Their Gene Structure, Evolution, and Expression. Genome Res. 2012, 22, 1775–1789. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.C.; Chang, H.Y. Molecular Mechanisms of Long Noncoding RNAs. Mol. Cell 2011, 43, 904–914. [Google Scholar] [CrossRef] [Green Version]
- Jarroux, J.; Morillon, A.; Pinskaya, M. History, Discovery, and Classification of LncRNAs. In Long Non Coding RNA Biology; Rao, M.R.S., Ed.; Advances in Experimental Medicine and Biology; Springer: Singapore, 2017; Volume 1008, pp. 1–46. [Google Scholar] [CrossRef]
- Bartolomei, M.S.; Zemel, S.; Tilghman, S.M. Parental Imprinting of the Mouse H19 Gene. Nature 1991, 351, 153–155. [Google Scholar] [CrossRef] [PubMed]
- Frankish, A.; Diekhans, M.; Jungreis, I.; Lagarde, J.; Loveland, J.E.; Mudge, J.M.; Sisu, C.; Wright, J.C.; Armstrong, J.; Barnes, I.; et al. GENCODE 2021. Nucleic Acids Res. 2021, 49, D916–D923. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, J.; Acuña, S.; Aoki, J.; Floeter-Winter, L.; Muxel, S. Long Non-Coding RNAs in the Regulation of Gene Expression: Physiology and Disease. Non-Coding RNA 2019, 5, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, H.; Fang, X. A Brief Review on the Human Encyclopedia of DNA Elements (ENCODE) Project. Genom. Proteom. Bioinform. 2013, 11, 135–141. [Google Scholar] [CrossRef] [Green Version]
- Aliperti, V.; Skonieczna, J.; Cerase, A. Long Non-Coding RNA (LncRNA) Roles in Cell Biology, Neurodevelopment and Neurological Disorders. Non-Coding RNA 2021, 7, 36. [Google Scholar] [CrossRef]
- Balas, M.M.; Johnson, A.M. Exploring the Mechanisms behind Long Noncoding RNAs and Cancer. Non-Coding RNA Res. 2018, 3, 108–117. [Google Scholar] [CrossRef]
- Han, P.; Chang, C.P. Long Non-Coding RNA and Chromatin Remodeling. RNA Biol. 2015, 12, 1094–1098. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Zhang, Y.; Hu, H.; Ren, G.; Zhang, W.; Liu, H. IRWNRLPI: Integrating Random Walk and Neighborhood Regularized Logistic Matrix Factorization for LncRNA-Protein Interaction Prediction. Front. Genet. 2018, 9, 239. [Google Scholar] [CrossRef] [Green Version]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A CeRNA Hypothesis: The Rosetta Stone of a Hidden RNA Language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef] [Green Version]
- Carlevaro-Fita, J.; Rahim, A.; Guigó, R.; Vardy, L.A.; Johnson, R. Cytoplasmic Long Noncoding RNAs Are Frequently Bound to and Degraded at Ribosomes in Human Cells. RNA 2016, 22, 867–882. [Google Scholar] [CrossRef] [Green Version]
- Du, Z.; Sun, T.; Hacisuleyman, E.; Fei, T.; Wang, X.; Brown, M.; Rinn, J.L.; Lee, M.G.S.; Chen, Y.; Kantoff, P.W.; et al. Integrative Analyses Reveal a Long Noncoding RNA-Mediated Sponge Regulatory Network in Prostate Cancer. Nat. Commun. 2016, 7, 10982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.; Ahn, C.; Chun, C.H.; Jin, E.J. A Long Non-Coding RNA, GAS5, Plays a Critical Role in the Regulation of MiR-21 during Osteoarthritis: THE INTER-REGULATION OF MiR-21 AND GAS5 IN OA. J. Orthop. Res. 2014, 32, 1628–1635. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Lim, K.H.; Kim, S.H.; Joo, J.Y. Molecular Landscape of Long Noncoding RNAs in Brain Disorders. Mol. Psychiatry 2021, 26, 1060–1074. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Sun, B.K.; Erwin, J.A.; Song, J.J.; Lee, J.T. Polycomb Proteins Targeted by a Short Repeat RNA to the Mouse X Chromosome. Science 2008, 322, 750–756. [Google Scholar] [CrossRef] [Green Version]
- Schertzer, M.D.; Braceros, K.C.A.; Starmer, J.; Cherney, R.E.; Lee, D.M.; Salazar, G.; Justice, M.; Bischoff, S.R.; Cowley, D.O.; Ariel, P.; et al. LncRNA-Induced Spread of Polycomb Controlled by Genome Architecture, RNA Abundance, and CpG Island DNA. Mol. Cell 2019, 75, 523–537. [Google Scholar] [CrossRef]
- Tsai, M.C.; Manor, O.; Wan, Y.; Mosammaparast, N.; Wang, J.K.; Lan, F.; Shi, Y.; Segal, E.; Chang, H.Y. Long Noncoding RNA as Modular Scaffold of Histone Modification Complexes. Science 2010, 329, 689–693. [Google Scholar] [CrossRef] [Green Version]
- Jovčevska, I.; Videtič Paska, A. Neuroepigenetics of Psychiatric Disorders: Focus on LncRNA. Neurochem. Int. 2021, 149, 105140. [Google Scholar] [CrossRef]
- Özeş, A.R.; Wang, Y.; Zong, X.; Fang, F.; Pilrose, J.; Nephew, K.P. Therapeutic Targeting Using Tumor Specific Peptides Inhibits Long Non-Coding RNA HOTAIR Activity in Ovarian and Breast Cancer. Sci. Rep. 2017, 7, 894. [Google Scholar] [CrossRef] [Green Version]
- Mayfield, R.D. Emerging Roles for NcRNAs in Alcohol Use Disorders. Alcohol 2017, 60, 31–39. [Google Scholar] [CrossRef] [Green Version]
- DiStefano, J.K. The Emerging Role of Long Noncoding RNAs in Human Disease. In Disease Gene Identification; DiStefano, J.K., Ed.; Methods in Molecular Biology; Springer: New York, NY, USA, 2018; Volume 1706, pp. 91–110. [Google Scholar] [CrossRef]
- Sun, X.; Wong, D. Long Non-Coding RNA-Mediated Regulation of Glucose Homeostasis and Diabetes. Am. J. Cardiovasc. Dis. 2016, 6, 17–25. [Google Scholar]
- Shabgah, A.G.; Norouzi, F.; Hedayati-Moghadam, M.; Soleimani, D.; Pahlavani, N.; Navashenaq, J.G. A Comprehensive Review of Long Non-Coding RNAs in the Pathogenesis and Development of Non-Alcoholic Fatty Liver Disease. Nutr. Metab. 2021, 18, 22. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.C.; Ni, J.J.; Cui, W.Y.; Wang, B.Y.; Zhuo, W. Emerging Roles of LncRNA in Cancer and Therapeutic Opportunities. Am. J. Cancer Res. 2019, 9, 1354–1366. [Google Scholar]
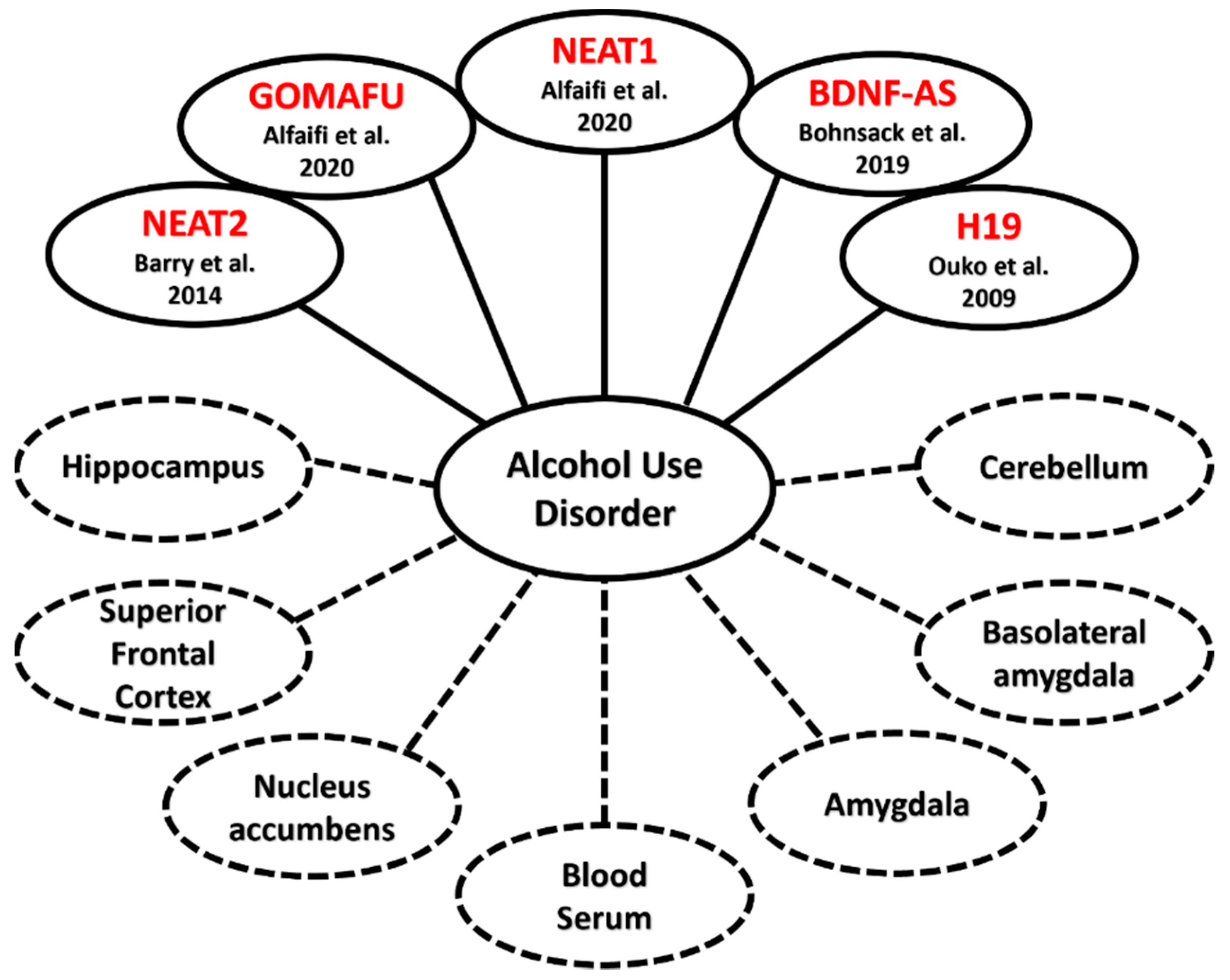
- Barry, G.; Briggs, J.A.; Vanichkina, D.P.; Poth, E.M.; Beveridge, N.J.; Ratnu, V.S.; Nayler, S.P.; Nones, K.; Hu, J.; Bredy, T.W.; et al. The Long Non-Coding RNA Gomafu Is Acutely Regulated in Response to Neuronal Activation and Involved in Schizophrenia-Associated Alternative Splicing. Mol. Psychiatry 2014, 19, 486–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuiji, H.; Yoshimoto, R.; Hasegawa, Y.; Furuno, M.; Yoshida, M.; Nakagawa, S. Competition between a Noncoding Exon and Introns: Gomafu Contains Tandem UACUAAC Repeats and Associates with Splicing Factor-1: Competition between Exons and Introns. Genes Cells 2011, 16, 479–490. [Google Scholar] [CrossRef] [Green Version]
- Sone, M.; Hayashi, T.; Tarui, H.; Agata, K.; Takeichi, M.; Nakagawa, S. The MRNA-like Noncoding RNA Gomafu Constitutes a Novel Nuclear Domain in a Subset of Neurons. J. Cell Sci. 2007, 120, 2498–2506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfaifi, M.; Ali Beg, M.M.; Alshahrani, M.Y.; Ahmad, I.; Alkhathami, A.G.; Joshi, P.C.; Alshehri, O.M.; Alamri, A.M.; Verma, A.K. Circulating Long Non-Coding RNAs NKILA, NEAT1, MALAT1, and MIAT Expression and Their Association in Type 2 Diabetes Mellitus. BMJ. Open Diab. Res. Care 2021, 9, e001821. [Google Scholar] [CrossRef]
- Seki, T.; Yamagata, H.; Uchida, S.; Chen, C.; Kobayashi, A.; Kobayashi, M.; Harada, K.; Matsuo, K.; Watanabe, Y.; Nakagawa, S. Altered Expression of Long Noncoding RNAs in Patients with Major Depressive Disorder. J. Psychiatr. Res. 2019, 117, 92–99. [Google Scholar] [CrossRef]
- Kingwell, K. Double Setback for ASO Trials in Huntington Disease. Nat. Rev. Drug Discov. 2021, 20, 412–413. [Google Scholar] [CrossRef]
- Chanda, K.; Das, S.; Chakraborty, J.; Bucha, S.; Maitra, A.; Chatterjee, R.; Mukhopadhyay, D.; Bhattacharyya, N.P. Altered Levels of Long NcRNAs Meg3 and Neat1 in Cell And Animal Models Of Huntington’s Disease. RNA Biol. 2018, 15, 1348–1363. [Google Scholar] [CrossRef] [Green Version]
- Grant, B.F.; Goldstein, R.B.; Saha, T.D.; Chou, S.P.; Jung, J.; Zhang, H.; Pickering, R.P.; Ruan, W.J.; Smith, S.M.; Huang, B.; et al. Epidemiology of DSM-5 Alcohol Use Disorder: Results From the National Epidemiologic Survey on Alcohol and Related Conditions III. JAMA Psychiatry 2015, 72, 757. [Google Scholar] [CrossRef] [Green Version]
- Stahre, M.; Roeber, J.; Kanny, D.; Brewer, R.D.; Zhang, X. Contribution of Excessive Alcohol Consumption to Deaths and Years of Potential Life Lost in the United States. Prev. Chronic Dis. 2014, 11, 130293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Institute on Alcohol Abuse and Alcoholism. Alcohol Use Disorder: A Comparison between DSM–IV and DSM–5. U.S. Department of Health and Human Services. 2021. Available online: https://www.niaaa.nih.gov/publications/brochures-and-fact-sheets/alcohol-use-disorder-comparison-between-dsm (accessed on 8 June 2022).
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Association: Washington, DC, USA, 2013. [Google Scholar] [CrossRef]
- Van Booven, D.; Li, M.; Rao, J.S.; Blokhin, I.O.; Dayne Mayfield, R.; Barbier, E.; Heilig, M.; Wahlestedt, C. Alcohol Use Disorder Causes Global Changes in Splicing in the Human Brain. Transl. Psychiatry 2021, 11, 2. [Google Scholar] [CrossRef] [PubMed]
- Tawa, E.A.; Hall, S.D.; Lohoff, F.W. Overview of the Genetics of Alcohol Use Disorder. Alcohol Alcohol. 2016, 51, 507–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, E.V.; Harris, R.A.; Pfefferbaum, A. Alcohol’s effects on brain and behavior. Alcohol. Res. Health 2010, 33, 127–143. [Google Scholar] [PubMed]
- Guo, R.; Ren, J. Alcohol and Acetaldehyde in Public Health: From Marvel to Menace. IJERPH 2010, 7, 1285–1301. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, P.L.; Miles, M.; Edenberg, H.J.; Sommer, W.; Tabakoff, B.; Wehner, J.M.; Lewohl, J. Gene Expression in Brain: A Window on Ethanol Dependence, Neuroadaptation, and Preference. Alcohol. Clin. Exp. Res. 2003, 27, 155–168. [Google Scholar] [CrossRef]
- Zhu, S.; Wu, J.; Hu, J. Non-Coding RNA in Alcohol Use Disorder by Affecting Synaptic Plasticity. Exp. Brain. Res. 2022, 240, 365–379. [Google Scholar] [CrossRef]
- Ma, H.; Zhu, G. The Dopamine System and Alcohol Dependence. Shanghai Arch. Psychiatry 2014, 26, 61–68. [Google Scholar] [CrossRef]
- Wu, H.M.; Wang, X.L.; Chang, C.W.; Li, N.; Gao, L.; Geng, N.; Ma, J.H.; Zhao, W.; Gao, G.D. Preliminary Findings in Ablating the Nucleus Accumbens Using Stereotactic Surgery for Alleviating Psychological Dependence on Alcohol. Neurosci. Lett. 2010, 473, 77–81. [Google Scholar] [CrossRef]
- Huang, Y.; Mohan, A.; De Ridder, D.; Sunaert, S.; Vanneste, S. The Neural Correlates of the Unified Percept of Alcohol-Related Craving: A FMRI and EEG Study. Sci. Rep. 2018, 8, 923. [Google Scholar] [CrossRef]
- Gorini, G.; Bell, R.L.; Mayfield, R.D. Molecular Targets of Alcohol Action. Prog. Mol. Biol. Transl. Sci. 2011, 98, 293–347. [Google Scholar] [CrossRef] [Green Version]
- Erdozain, A.M.; Morentin, B.; Bedford, L.; King, E.; Tooth, D.; Brewer, C.; Wayne, D.; Johnson, L.; Gerdes, H.K.; Wigmore, P.; et al. Alcohol-Related Brain Damage in Humans. PLoS ONE 2014, 9, e93586. [Google Scholar] [CrossRef] [PubMed]
- Verhulst, B.; Neale, M.C.; Kendler, K.S. The Heritability of Alcohol Use Disorders: A Meta-Analysis of Twin and Adoption Studies. Psychol. Med. 2015, 45, 1061–1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salvatore, J.E.; Gottesman, I.I.; Dick, D.M. Endophenotypes for Alcohol Use Disorder: An Update on the Field. Curr. Addict. Rep. 2015, 2, 76–90. [Google Scholar] [CrossRef] [PubMed]
- Gottesman, I.I.; Gould, T.D. The Endophenotype Concept in Psychiatry: Etymology and Strategic Intentions. Am. J. Psychiatry 2003, 160, 636–645. [Google Scholar] [CrossRef] [PubMed]
- Deak, J.D.; Miller, A.P.; Gizer, I.R. Genetics of Alcohol Use Disorder: A Review. Curr. Opin. Psychol. 2019, 27, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Clarke, A.J.; Cooper, D.N. GWAS: Heritability Missing in Action? Eur. J. Hum. Genet. 2010, 18, 859–861. [Google Scholar] [CrossRef] [Green Version]
- Gelernter, J.; Kranzler, H.R.; Sherva, R.; Almasy, L.; Koesterer, R.; Smith, A.H.; Anton, R.; Preuss, U.W.; Ridinger, M.; Rujescu, D.; et al. Genome-wide association study of alcohol dependence: Significant findings in African- and European-Americans including novel risk loci. Mol. Psychiatry 2014, 19, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Edwards, S.L.; Beesley, J.; French, J.D.; Dunning, A.M. Beyond GWASs: Illuminating the Dark Road from Association to Function. Am. J. Hum. Genet. 2013, 93, 779–797. [Google Scholar] [CrossRef] [Green Version]
- Tam, V.; Patel, N.; Turcotte, M.; Bossé, Y.; Paré, G.; Meyre, D. Benefits and Limitations of Genome-Wide Association Studies. Nat. Rev. Genet. 2019, 20, 467–484. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, F.; Xu, H.; Liu, Y.; Liu, J.; Zhao, H.; Gelernter, J. Differentially co-expressed genes in postmortem prefrontal cortex of individuals with alcohol use disorders: Influence on alcohol metabolism-related pathways. Hum. Genet. 2014, 133, 1383–1394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapoor, M.; Chao, M.J.; Johnson, E.C. Multi-omics integration analysis identifies novel genes for alcoholism with potential overlap with neurodegenerative diseases. Nat. Commun. 2021, 12, 5071. [Google Scholar] [CrossRef] [PubMed]
- Vornholt, E.; Luo, D.; Qiu, W.; McMichael, G.O.; Liu, Y.; Gillespie, N.; Ma, C.; Vladimirov, V.I. Postmortem Brain Tissue as an Underutilized Resource to Study the Molecular Pathology of Neuropsychiatric Disorders across Different Ethnic Populations. Neurosci. Biobehav. Rev. 2019, 102, 195–207. [Google Scholar] [CrossRef] [PubMed]
- Wall, T.L.; Ehlers, C.L. Genetic Influences Affecting Alcohol Use Among Asians. Alcohol Health Res. World 1995, 19, 184–189. [Google Scholar]
- Edenberg, H.J. The genetics of alcohol metabolism: Role of alcohol dehydrogenase and aldehyde dehydrogenase variants. Alcohol. Res. Health 2007, 30, 5–13. [Google Scholar]
- Adkins, A.E.; Hack, L.M.; Bigdeli, T.B.; Williamson, V.S.; McMichael, G.O.; Mamdani, M.; Edwards, A.C.; Aliev, F.; Chan, R.F.; Bhandari, P.; et al. Genomewide Association Study of Alcohol Dependence Identifies Risk Loci Altering Ethanol-Response Behaviors in Model Organisms. Alcohol. Clin. Exp. Res. 2017, 41, 911–928. [Google Scholar] [CrossRef] [Green Version]
- Xu, K.; Kranzler, H.R.; Sherva, R.; Sartor, C.E.; Almasy, L.; Koesterer, R.; Zhao, H.; Farrer, L.A.; Gelernter, J. Genomewide Association Study for Maximum Number of Alcoholic Drinks in European Americans and African Americans. Alcohol Clin. Exp. Res. 2015, 39, 1137–1147. [Google Scholar] [CrossRef]
- Sharma, U.; Barwal, T.S.; Khandelwal, A.; Rana, M.K.; Rana, A.P.S.; Singh, K.; Jain, A. Circulating Long Non-Coding RNAs LINC00324 and LOC100507053 as Potential Liquid Biopsy Markers for Esophageal Squamous Cell Carcinoma: A Pilot Study. Front. Oncol. 2022, 12, 823953. [Google Scholar] [CrossRef]
- Kryger, R.; Fan, L.; Wilce, P.A.; Jaquet, V. MALAT-1, a Non Protein-Coding RNA Is Upregulated in the Cerebellum, Hippocampus and Brain Stem of Human Alcoholics. Alcohol 2012, 46, 629–634. [Google Scholar] [CrossRef]
- Drake, J.; McMichael, G.O.; Vornholt, E.S.; Cresswell, K.; Williamson, V.; Chatzinakos, C.; Mamdani, M.; Hariharan, S.; Kendler, K.S.; Kalsi, G.; et al. Assessing the Role of Long Noncoding RNA in Nucleus Accumbens in Subjects With Alcohol Dependence. Alcohol. Clin. Exp. Res. 2020, 44, 2468–2480. [Google Scholar] [CrossRef]
- Ouko, L.A.; Shantikumar, K.; Knezovich, J.; Haycock, P.; Schnugh, D.J.; Ramsay, M. Effect of Alcohol Consumption on CpG Methylation in the Differentially Methylated Regions of H19 and IG-DMR in Male Gametes-Implications for Fetal Alcohol Spectrum Disorders. Alcohol. Clin. Exp. Res. 2009, 33, 1615–1627. [Google Scholar] [CrossRef] [PubMed]
- Bohnsack, J.P.; Teppen, T.; Kyzar, E.J.; Dzitoyeva, S.; Pandey, S.C. The LncRNA BDNF-AS Is an Epigenetic Regulator in the Human Amygdala in Early Onset Alcohol Use Disorders. Transl. Psychiatry 2019, 9, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bu, Q.; Hu, Z.; Chen, F.; Zhu, R.; Deng, Y.; Shao, X.; Li, Y.; Zhao, J.; Li, H.; Zhang, B.; et al. Transcriptome analysis of long non-coding RNAs of the nucleus accumbens in cocaine-conditioned mice. J. Neurochem. 2012, 123, 790–799. [Google Scholar] [CrossRef] [PubMed]
- Albertson, D.N.; Pruetz, B.; Schmidt, C.J.; Kuhn, D.M.; Kapatos, G.; Bannon, M.J. Gene expression profile of the nucleus accumbens of human cocaine abusers: Evidence for dysregulation of myelin: Gene expression profile of human cocaine abuse. J. Neurochem. 2004, 88, 1211–1219. [Google Scholar] [CrossRef]
- Michelhaugh, S.K.; Lipovich, L.; Blythe, J.; Jia, H.; Kapatos, G.; Bannon, M.J. Mining Affymetrix microarray data for long non-coding RNAs: Altered expression in the nucleus accumbens of heroin abusers: Long non-coding RNAs in heroin abusers. J. Neurochem. 2011, 116, 459–466. [Google Scholar] [CrossRef] [Green Version]
- Yoon, J.-H.; Abdelmohsen, K.; Srikantan, S.; Yang, X.; Martindale, J.L.; De, S.; Huarte, M.; Zhan, M.; Becker, K.G.; Gorospe, M. LincRNA-p21 Suppresses Target mRNA Translation. Mol. Cell 2012, 47, 648–655. [Google Scholar] [CrossRef] [Green Version]
- Starkman, B.G.; Sakharkar, A.J.; Pandey, S.C. Epigenetics-beyond the Genome in Alcoholism. Alcohol Res. 2012, 34, 293–305. [Google Scholar]
- Holliday, R. Epigenetics: A Historical Overview. Epigenetics 2006, 1, 76–80. [Google Scholar] [CrossRef] [Green Version]
- Zillich, L.; Frank, J.; Streit, F.; Friske, M.M.; Foo, J.C.; Sirignano, L.; Heilmann-Heimbach, S.; Dukal, H.; Degenhardt, F.; Hoffmann, P.; et al. Epigenome-Wide Association Study of Alcohol Use Disorder in Five Brain Regions. Neuropsychopharmacol. 2022, 47, 832–839. [Google Scholar] [CrossRef]
- Brière, F.N.; Rohde, P.; Seeley, J.R.; Klein, D.; Lewinsohn, P.M. Comorbidity between Major Depression and Alcohol Use Disorder from Adolescence to Adulthood. Compr. Psychiatry 2014, 55, 526–533. [Google Scholar] [CrossRef] [Green Version]
- Hunt, G.E.; Malhi, G.S.; Lai, H.M.X.; Cleary, M. Prevalence of Comorbid Substance Use in Major Depressive Disorder in Community and Clinical Settings, 1990–2019: Systematic Review and Meta-Analysis. J. Affect. Disord. 2020, 266, 288–304. [Google Scholar] [CrossRef] [PubMed]
- Zuo, L.; Tan, Y.; Wang, Z.; Wang, K.S.; Zhang, X.; Chen, X.; Li, C.S.R.; Wang, T.; Luo, X. Long Noncoding RNAs in Psychiatric Disorders. Psychiatr. Genet. 2016, 26, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Chen, B.; Dong, P.; Zheng, J. HOTAIR Epigenetically Modulates PTEN Expression via MicroRNA-29b: A Novel Mechanism in Regulation of Liver Fibrosis. Mol. Ther. 2017, 25, 205–217. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Han, X.; Zhang, Z.; Zheng, L.; Hu, Z.; Yao, Q.; Cui, H.; Shu, G.; Si, M.; Li, C.; et al. The liver-enriched lnc-LFAR1 promotes liver fibrosis by activating TGFβ and Notch pathways. Nat. Commun. 2017, 8, 144. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Tan, X.-R.; Li, S.-J.; Zhang, X.-X. LncRNA NEAT1 promotes hepatic lipid accumulation via regulating miR-146a-5p/ROCK1 in nonalcoholic fatty liver disease. Life Sci. 2019, 235, 116829. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, Z.; Chen, X.; Zhang, S. Long Non-Coding RNAs: From Disease Code to Drug Role. Acta Pharm. Sin. B 2021, 11, 340–354. [Google Scholar] [CrossRef] [PubMed]
- Wurster, C.D.; Ludolph, A.C. Antisense Oligonucleotides in Neurological Disorders. Ther. Adv. Neurol. Disord. 2018, 11, 175628641877693. [Google Scholar] [CrossRef] [Green Version]
- Robillard, K.N.; de Vrieze, E.; van Wijk, E.; Lentz, J.J. Altering Gene Expression Using Antisense Oligonucleotide Therapy for Hearing Loss. Hear. Res. 2022, 108523. [Google Scholar] [CrossRef]
- Winkle, M.; El-Daly, S.M.; Fabbri, M.; Calin, G.A. Noncoding RNA Therapeutics—Challenges and Potential Solutions. Nat. Rev. Drug. Discov. 2021, 20, 629–651. [Google Scholar] [CrossRef]
- Dindot, S.V.; Christian, S.; Murphy, W.J.; Berent, A.; Panagoulias, J.; Schlafer, A.; Ballard, J.; Radeva, K.; Robinson, R.; Myers, L.; et al. Development of an ASO Therapy for Angelman Syndrome by Targeting an Evolutionarily Conserved Region at the Start of the UBE3A-AS Transcript. Genetics 2021, Preprint. [Google Scholar] [CrossRef]
- Gu, C.; Chen, C.; Wu, R.; Dong, T.; Hu, X.; Yao, Y.; Zhang, Y. Long Noncoding RNA EBF3-AS Promotes Neuron Apoptosis in Alzheimer’s Disease. DNA Cell Biol. 2018, 37, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Statello, L.; Guo, C.J.; Chen, L.L.; Huarte, M. Gene Regulation by Long Non-Coding RNAs and Its Biological Functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DSM-5 Alcohol Use Disorder Symptoms | |
|---|---|
| 1 | Drinking longer or more than intended |
| 2 | Tried to quit or decrease levels of drinking, but failed |
| 3 | Sick from the aftereffects |
| 4 | Incapability to not think about drinking |
| 5 | Drinking interferes with your daily life (job, family, school, etc…) |
| 6 | Continued drinking habits regardless of daily life struggles |
| 7 | Loss of pleasure in things you once loved |
| 8 | Reckless behavior (driving, fighting, unsafe sex, etc…) |
| 9 | Continued drinking even if depressed or anxious or experiencing memory problems |
| 10 | Increased tolerance to alcohol |
| 11 | Withdrawal symptoms (shakiness, nausea, sweating, etc…) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Denham, A.N.; Drake, J.; Gavrilov, M.; Taylor, Z.N.; Bacanu, S.-A.; Vladimirov, V.I. Long Non-Coding RNAs: The New Frontier into Understanding the Etiology of Alcohol Use Disorder. Non-Coding RNA 2022, 8, 59. https://0-doi-org.brum.beds.ac.uk/10.3390/ncrna8040059
Denham AN, Drake J, Gavrilov M, Taylor ZN, Bacanu S-A, Vladimirov VI. Long Non-Coding RNAs: The New Frontier into Understanding the Etiology of Alcohol Use Disorder. Non-Coding RNA. 2022; 8(4):59. https://0-doi-org.brum.beds.ac.uk/10.3390/ncrna8040059
Chicago/Turabian StyleDenham, Allie N., John Drake, Matthew Gavrilov, Zachary N. Taylor, Silviu-Alin Bacanu, and Vladimir I. Vladimirov. 2022. "Long Non-Coding RNAs: The New Frontier into Understanding the Etiology of Alcohol Use Disorder" Non-Coding RNA 8, no. 4: 59. https://0-doi-org.brum.beds.ac.uk/10.3390/ncrna8040059