Effects of Frozen Storage on Phospholipid Content in Atlantic Cod Fillets and the Influence on Diet-Induced Obesity in Mice

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Animal Studies
2.3. Lipid Class Composition
2.4. Fatty Acid Composition in the Polar and Neutral Lipid Fractions
2.5. Experimental Diets
2.6. Body Composition of the Mice
2.7. Apparent Digestibility of Nitrogen and Fat
2.8. Oral Glucose Tolerance Test
2.9. Statistics
3. Results
3.1. Frozen Storage of Cod Fillets Decreased the Polar Lipid:FFA Ratio
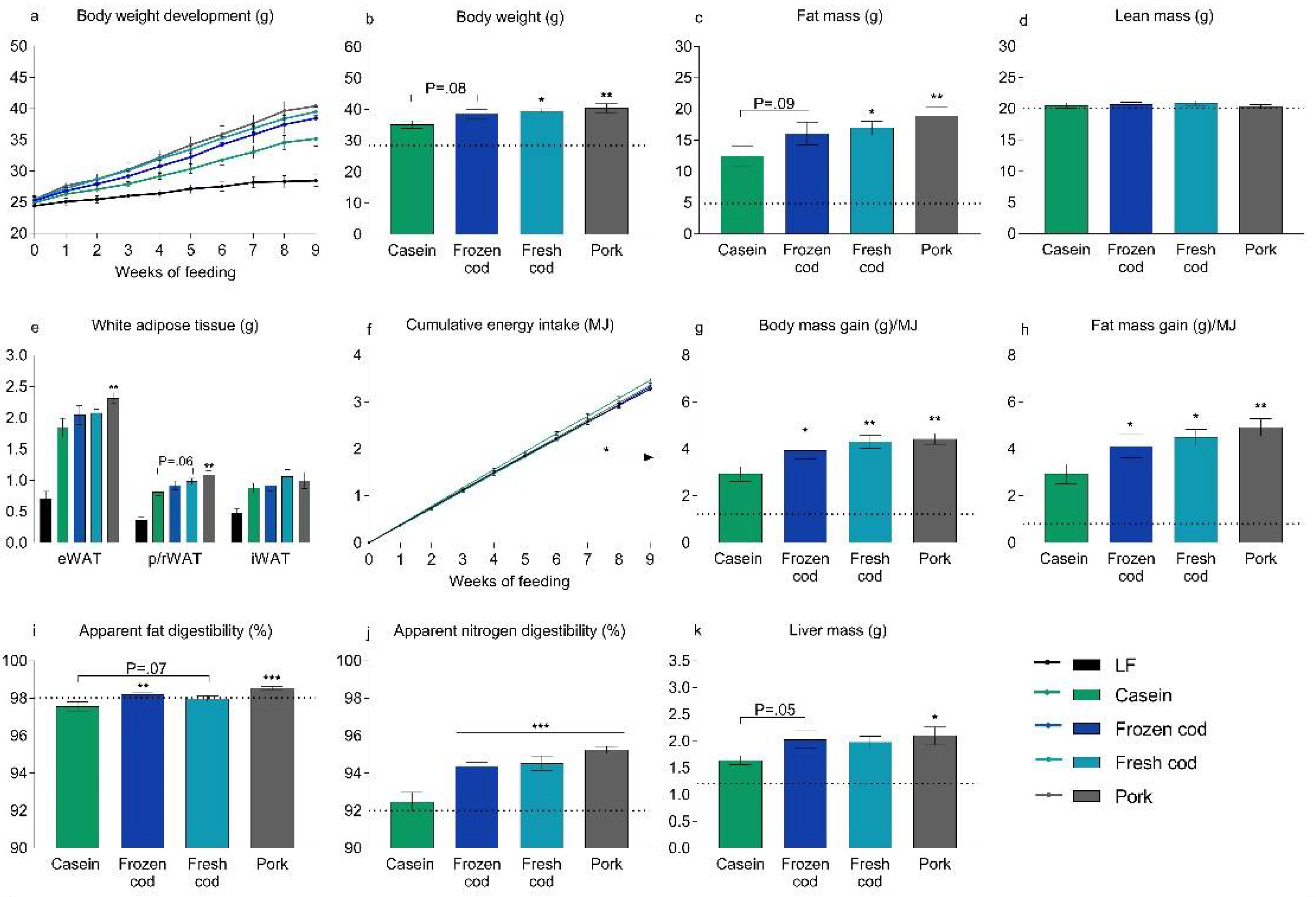
3.2. Fresh Cod, But Not Frozen Cod, Is More Obesogenic than Casein in a Western Diet
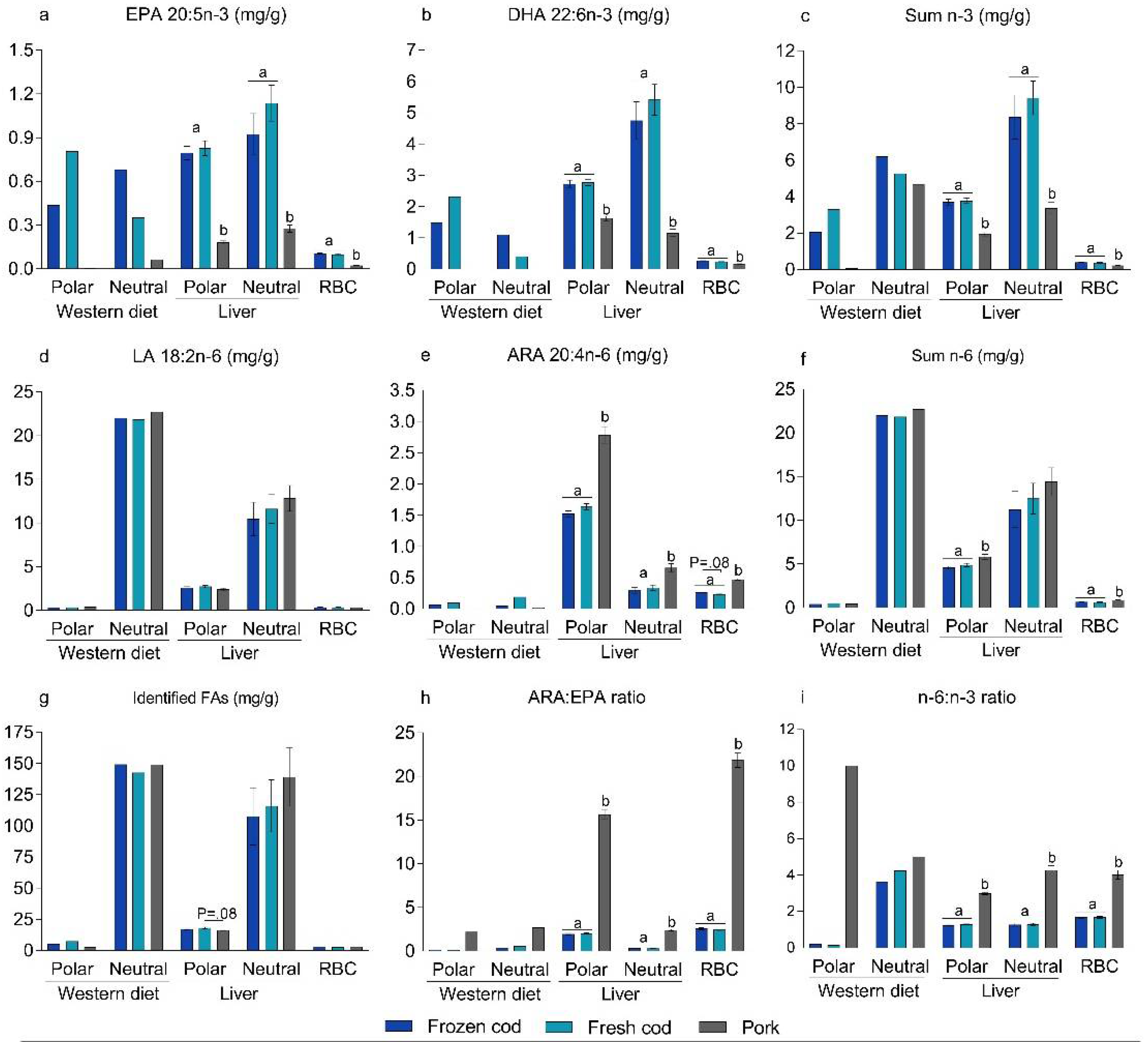
3.3. Bioavailability of n-3 PUFAs in Frozen and Fresh Cod Diets
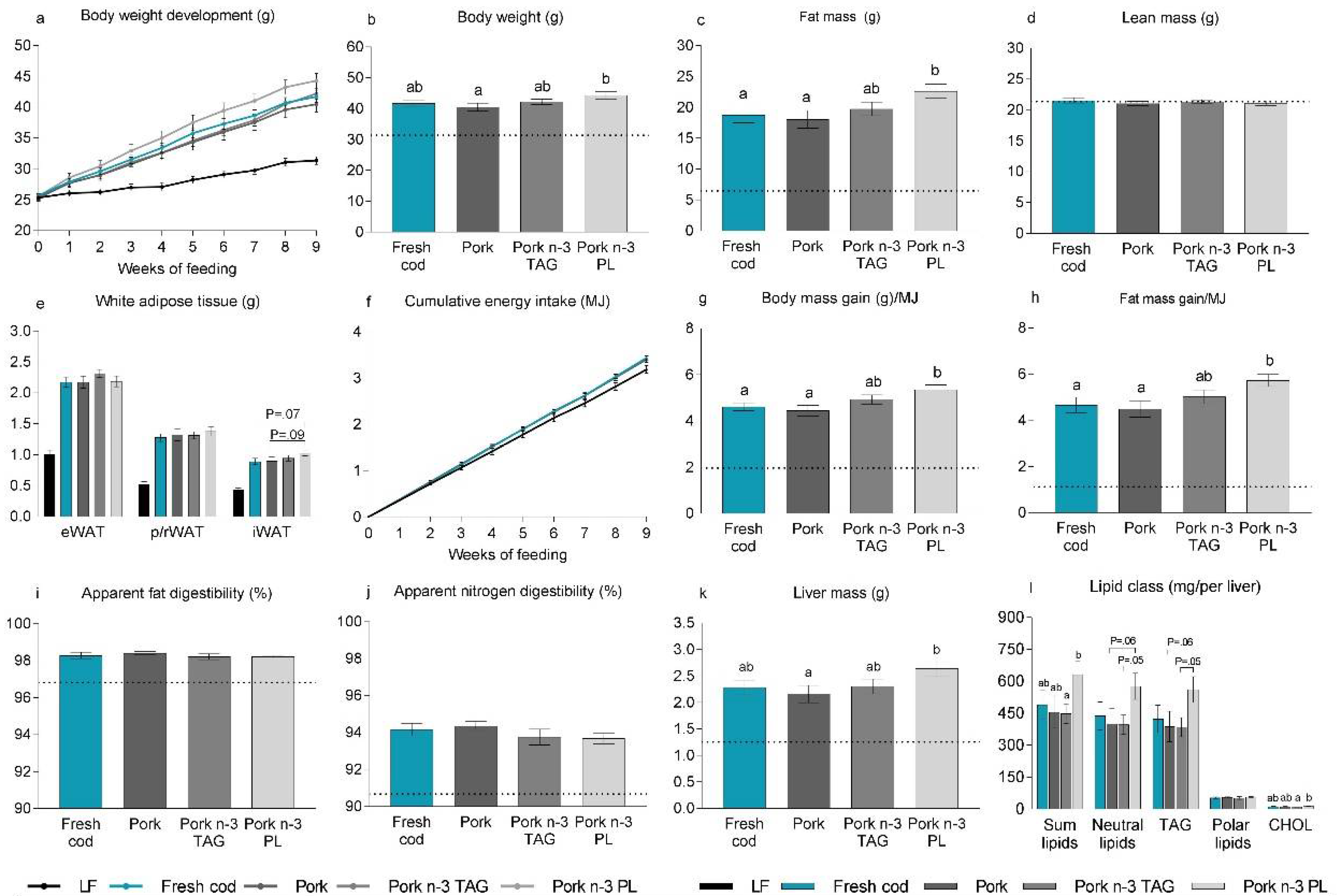
3.4. Supplementation with n-3 PUFAs in Pork-Based Western Diets
3.5. Adding PL-Bound n-3 PUFAs to a Western Diet Containing Pork Promoted Obesity in Mice
3.6. Bioavailability of PL-Bound n-3 PUFAs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mozaffarian, D. Dietary and policy priorities for cardiovascular disease, diabetes, and obesity: A comprehensive review. Circulation 2016, 133, 187–225. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.D.; Hou, T.; Ludwig, D.S.; Rimm, E.B.; Willett, W.; Hu, F.B.; Mozaffarian, D. Changes in intake of protein foods, carbohydrate amount and quality, and long-term weight change: Results from 3 prospective cohorts. Am. J. Clin. Nutr. 2015, 101, 1216–1224. [Google Scholar] [CrossRef] [PubMed]
- Holm, J.B.; Ronnevik, A.; Tastesen, H.S.; Fjaere, E.; Fauske, K.R.; Liisberg, U.; Madsen, L.; Kristiansen, K.; Liaset, B. Diet-induced obesity, energy metabolism and gut microbiota in c57bl/6j mice fed western diets based on lean seafood or lean meat mixtures. J. Nutr. Biochem. 2016, 31, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Liisberg, U.; Fauske, K.R.; Kuda, O.; Fjaere, E.; Myrmel, L.S.; Norberg, N.; Froyland, L.; Graff, I.E.; Liaset, B.; Kristiansen, K.; et al. Intake of a western diet containing cod instead of pork alters fatty acid composition in tissue phospholipids and attenuates obesity and hepatic lipid accumulation in mice. J. Nutr. Biochem. 2016, 33, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Lavigne, C.; Marette, A.; Jacques, H. Cod and soy proteins compared with casein improve glucose tolerance and insulin sensitivity in rats. Am. J. Physiol. Endocrinol. Metab. 2000, 278, E491–E500. [Google Scholar] [CrossRef] [PubMed]
- Lavigne, C.; Tremblay, F.; Asselin, G.; Jacques, H.; Marette, A. Prevention of skeletal muscle insulin resistance by dietary cod protein in high fat-fed rats. Am. J. Physiol. Endocrinol. Metab. 2001, 281, E62–E71. [Google Scholar] [CrossRef] [PubMed]
- Tastesen, H.S.; Keenan, A.H.; Madsen, L.; Kristiansen, K.; Liaset, B. Scallop protein with endogenous high taurine and glycine content prevents high-fat, high-sucrose-induced obesity and improves plasma lipid profile in male c57bl/6j mice. Amino. Acids 2014, 46, 1659–1671. [Google Scholar] [CrossRef] [PubMed]
- Liisberg, U.; Myrmel, L.S.; Fjaere, E.; Ronnevik, A.K.; Bjelland, S.; Fauske, K.R.; Holm, J.B.; Basse, A.L.; Hansen, J.B.; Liaset, B.; et al. The protein source determines the potential of high protein diets to attenuate obesity development in c57bl/6j mice. Adipocyte 2016, 5, 196–211. [Google Scholar] [CrossRef] [PubMed]
- Lie, Ø.; Lambertsen, G. Fatty acid composition of glycerophospholipids in seven tissues of cod (Gadus morhua), determined by combined high-performance liquid chromatography and gas chromatography. J. Chromatogr. B Biomed. Sci. Appl. 1991, 565, 119–129. [Google Scholar] [CrossRef]
- Murru, E.; Banni, S.; Carta, G. Nutritional properties of dietary omega-3-enriched phospholipids. Biomed. Res. Int. 2013, 2013, 965417. [Google Scholar] [CrossRef] [PubMed]
- Rossmeisl, M.; Jilkova, Z.M.; Kuda, O.; Jelenik, T.; Medrikova, D.; Stankova, B.; Kristinsson, B.; Haraldsson, G.G.; Svensen, H.; Stoknes, I. Metabolic effects of n-3 pufa as phospholipids are superior to triglycerides in mice fed a high-fat diet: Possible role of endocannabinoids. PLoS ONE 2012, 7, e38834. [Google Scholar] [CrossRef] [PubMed]
- Roiha, I.S.; Jonsson, A.; Backi, C.J.; Lunestad, B.T.; Karlsdottir, M.G. A comparative study of quality and safety of atlantic cod (Gadus morhua) fillets during cold storage, as affected by different thawing methods of pre-rigor frozen headed and gutted fish. J. Sci. Food Agric. 2018, 98, 400–409. [Google Scholar] [CrossRef] [PubMed]
- Bligh, E.; Scott, M.A. Lipids of cod muscle and the effect of frozen storage. J. Fish. Board Can. 1966, 23, 1025–1036. [Google Scholar] [CrossRef]
- Olley, J.; Lovern, J. Phospholipid hydrolysis in cod flesh stored at various temperatures. J. Sci. Food Agric. 1960, 11, 644–652. [Google Scholar] [CrossRef]
- Anderson, M.L.; Ravesi, E.M. Reaction of free fatty acids with protein in cod muscle frozen and stored at−29 c after aging in ice. J. Fish. Board Can. 1969, 26, 2727–2736. [Google Scholar] [CrossRef]
- Dyer, W.; Fraser, D.I. Proteins in fish muscle. 13. Lipid hydrolysis. J. Fish. Board Can. 1959, 16, 43–52. [Google Scholar] [CrossRef]
- Badii, F.; Howell, N.K. Changes in the texture and structure of cod and haddock fillets during frozen storage. Food Hydrocoll. 2002, 16, 313–319. [Google Scholar] [CrossRef]
- Badii, F.; Howell, N.K. A comparison of biochemical changes in cod (Gadus morhua) and haddock (Melanogrammus aeglefinus) fillets during frozen storage. J. Sci. Food Agric. 2002, 82, 87–97. [Google Scholar] [CrossRef]
- Jordal, A.E.; Lie, Ø.; Torstensen, B. Complete replacement of dietary fish oil with a vegetable oil blend affect liver lipid and plasma lipoprotein levels in atlantic salmon (Salmo salar L.). Aquacult. Nutr. 2007, 13, 114–130. [Google Scholar] [CrossRef]
- Torstensen, B.E.; Frøyland, L.; Ørnsrud, R.; Lie, Ø. Tailoring of a cardioprotective muscle fatty acid composition of atlantic salmon (Salmo salar) fed vegetable oils. Food Chem. 2004, 87, 567–580. [Google Scholar] [CrossRef]
- Mariotti, F.; Tome, D.; Mirand, P.P. Converting nitrogen into protein—Beyond 6.25 and jones’ factors. Crit. Rev. Food Sci. Nutr. 2008, 48, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Halldorsdottir, S.; Carmody, J.; Boozer, C.N.; Leduc, C.A.; Leibel, R.L. Reproducibility and accuracy of body composition assessments in mice by dual energy x-ray absorptiometry and time domain nuclear magnetic resonance. Int. J. Body Compos. Res. 2009, 7, 147. [Google Scholar] [PubMed]
- Andrikopoulos, S.; Blair, A.R.; Deluca, N.; Fam, B.C.; Proietto, J. Evaluating the glucose tolerance test in mice. Am. J. Physiol. Endocrinol. Metab. 2008, 295, E1323–E1332. [Google Scholar] [CrossRef] [PubMed]
- Batetta, B.; Griinari, M.; Carta, G.; Murru, E.; Ligresti, A.; Cordeddu, L.; Giordano, E.; Sanna, F.; Bisogno, T.; Uda, S. Endocannabinoids may mediate the ability of (n-3) fatty acids to reduce ectopic fat and inflammatory mediators in obese zucker rats. J. Nutr. 2009, 139, 1495–1501. [Google Scholar] [CrossRef] [PubMed]
- Vigerust, N.F.; Bjorndal, B.; Bohov, P.; Brattelid, T.; Svardal, A.; Berge, R.K. Krill oil versus fish oil in modulation of inflammation and lipid metabolism in mice transgenic for tnf-alpha. Eur. J. Nutr. 2013, 52, 1315–1325. [Google Scholar] [CrossRef] [PubMed]
- Tillander, V.; Bjorndal, B.; Burri, L.; Bohov, P.; Skorve, J.; Berge, R.K.; Alexson, S.E. Fish oil and krill oil supplementations differentially regulate lipid catabolic and synthetic pathways in mice. Nutr. Metab. 2014, 11, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burri, L.; Berge, K.; Wibrand, K.; Berge, R.K.; Barger, J.L. Differential effects of krill oil and fish oil on the hepatic transcriptome in mice. Front. Genet. 2011, 2, 45. [Google Scholar] [CrossRef] [PubMed]
- Awada, M.; Meynier, A.; Soulage, C.O.; Hadji, L.; Geloen, A.; Viau, M.; Ribourg, L.; Benoit, B.; Debard, C.; Guichardant, M.; et al. N-3 pufa added to high-fat diets affect differently adiposity and inflammation when carried by phospholipids or triacylglycerols in mice. Nutr. Metab. 2013, 10, 23. [Google Scholar] [CrossRef] [PubMed]
- Tou, J.C.; Jaczynski, J.; Chen, Y.C. Krill for human consumption: Nutritional value and potential health benefits. Nutr. Rev. 2007, 65, 63–77. [Google Scholar] [CrossRef] [PubMed]
- Vance, D.E. Phospholipid methylation in mammals: From biochemistry to physiological function. Biochim. Biophys. Acta. 2014, 1838, 1477–1487. [Google Scholar] [CrossRef] [PubMed]
- Niki, E. Role of vitamin e as a lipid-soluble peroxyl radical scavenger: In vitro and in vivo evidence. Free Radic. Biol. Med. 2014, 66, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.; Nagashimada, M.; Zhuge, F.; Zhan, L.; Nagata, N.; Tsutsui, A.; Nakanuma, Y.; Kaneko, S.; Ota, T. Astaxanthin prevents and reverses diet-induced insulin resistance and steatohepatitis in mice: A comparison with vitamin e. Sci. Rep. 2015, 5, 17192. [Google Scholar] [CrossRef] [PubMed]
- Tandy, S.; Chung, R.W.; Wat, E.; Kamili, A.; Berge, K.; Griinari, M.; Cohn, J.S. Dietary krill oil supplementation reduces hepatic steatosis, glycemia, and hypercholesterolemia in high-fat-fed mice. J. Agric. Food Chem. 2009, 57, 9339–9345. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Cui, J.; Li, Z.; Xu, J.; Wang, J.; Xue, C.; Wang, Y. Comparative study of dha-enriched phospholipids and epa-enriched phospholipids on metabolic disorders in diet-induced-obese c57bl/6j mice. Eur. J. Lipid Sci. Technol. 2014, 116, 255–265. [Google Scholar] [CrossRef]
- Madsen, L.; Frøyland, L.; Dyrøy, E.; Helland, K.; Berge, R.K. Docosahexaenoic and eicosapentaenoic acids are differently metabolized in rat liver during mitochondria and peroxisome proliferation. J. Lipid Res. 1998, 39, 583–593. [Google Scholar] [PubMed]
- Lopez-Vicario, C.; Rius, B.; Alcaraz-Quiles, J.; Garcia-Alonso, V.; Lopategi, A.; Titos, E.; Claria, J. Pro-resolving mediators produced from epa and dha: Overview of the pathways involved and their mechanisms in metabolic syndrome and related liver diseases. Eur. J. Pharmacol. 2016, 785, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Frøyland, L.; Madsen, L.; Vaagenes, H.; Totland, G.; Auwerx, J.; Kryvi, H.; Staels, B.; Berge, R. Mitochondrion is the principal target for nutritional and pharmacological control of triglyceride metabolism. J. Lipid Res. 1997, 38, 1851–1858. [Google Scholar] [PubMed]
- Alvheim, A.R.; Torstensen, B.E.; Lin, Y.H.; Lillefosse, H.H.; Lock, E.-J.; Madsen, L.; Hibbeln, J.R.; Malde, M.K. Dietary linoleic acid elevates endogenous 2-arachidonoylglycerol and anandamide in atlantic salmon (Salmo salar L.) and mice, and induces weight gain and inflammation in mice. Br. J. Nutr. 2013, 109, 1508–1517. [Google Scholar] [CrossRef] [PubMed]
- Midtbø, L.K.; Borkowska, A.G.; Bernhard, A.; Rønnevik, A.K.; Lock, E.-J.; Fitzgerald, M.L.; Torstensen, B.E.; Liaset, B.; Brattelid, T.; Pedersen, T.L. Intake of farmed atlantic salmon fed soybean oil increases hepatic levels of arachidonic acid-derived oxylipins and ceramides in mice. J. Nutr. Biochem. 2015, 26, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Midtbø, L.K.; Ibrahim, M.M.; Myrmel, L.S.; Aune, U.L.; Alvheim, A.R.; Liland, N.S.; Torstensen, B.E.; Rosenlund, G.; Liaset, B.; Brattelid, T. Intake of farmed atlantic salmon fed soybean oil increases insulin resistance and hepatic lipid accumulation in mice. PLoS ONE 2013, 8, e53094. [Google Scholar] [CrossRef] [PubMed]
- Salem, N., Jr.; Kuratko, C.N. A reexamination of krill oil bioavailability studies. Lipids Health Dis. 2014, 13, 137. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.W.; Cannon, B.; Nedergaard, J. Optimal housing temperatures for mice to mimic the thermal environment of humans: An experimental study. Mol. Metab. 2018, 7, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Sadurskis, A.; Dicker, A.; Cannon, B.; Nedergaard, J. Polyunsaturated fatty acids recruit brown adipose tissue: Increased ucp content and nst capacity. Am. J. Physiol. 1995, 269, E351–E360. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Goto, T.; Yu, R.; Uchida, K.; Tominaga, M.; Kano, Y.; Takahashi, N.; Kawada, T. Fish oil intake induces ucp1 upregulation in brown and white adipose tissue via the sympathetic nervous system. Sci. Rep. 2015, 5, 18013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannon, B.; Nedergaard, J. Brown adipose tissue: Function and physiological significance. Physiol. Rev. 2004, 84, 277–359. [Google Scholar] [CrossRef] [PubMed]
- Feldmann, H.M.; Golozoubova, V.; Cannon, B.; Nedergaard, J. Ucp1 ablation induces obesity and abolishes diet-induced thermogenesis in mice exempt from thermal stress by living at thermoneutrality. Cell Metab. 2009, 9, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Goldgof, M.; Xiao, C.; Chanturiya, T.; Jou, W.; Gavrilova, O.; Reitman, M.L. The chemical uncoupler 2,4-dinitrophenol (dnp) protects against diet-induced obesity and improves energy homeostasis in mice at thermoneutrality. J. Biol. Chem. 2014, 289, 19341–19350. [Google Scholar] [CrossRef] [PubMed]
- Ziętak, M.; Kovatcheva-Datchary, P.; Markiewicz, L.H.; Ståhlman, M.; Kozak, L.P.; Bäckhed, F. Altered microbiota contributes to reduced diet-induced obesity upon cold exposure. Cell Metab. 2016, 23, 1216–1223. [Google Scholar] [CrossRef] [PubMed]
- Rossmeisl, M.; Medrikova, D.; Van Schothorst, E.M.; Pavlisova, J.; Kuda, O.; Hensler, M.; Bardova, K.; Flachs, P.; Stankova, B.; Vecka, M. Omega-3 phospholipids from fish suppress hepatic steatosis by integrated inhibition of biosynthetic pathways in dietary obese mice. Biochim. Biophys. Acta BBA Mol. Cell Biol. Lipids 2014, 1841, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Dyer, W. Protein denaturation in frozen and stored fish. J. Food Sci. 1951, 16, 522–527. [Google Scholar] [CrossRef]
- Shenouda, S.Y. Theories of protein denaturation during frozen storage of fish flesh. Adv. Food Res. 1980, 26, 275–311. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Freeze-Dried Frozen Cod Fillets | Freeze-Dried Fresh Cod Fillets | |||
|---|---|---|---|---|
| Lipid Class | mg/g | % | mg/g | % |
| PC | 18.80 ± 0.3 | 42.0 | 36.5 ± 0.8 | 57.9 |
| PE | 5.1 ± 2.2 | 11.5 | 14.2 ± 0.3 | 22.6 |
| PI | <0.01 | <0.01 | 0.48 ± 0.05 | 0.76 |
| PS | 0.12 ± 0.02 | 0.27 | 0.86 ± 0.07 | 1.4 |
| LPC | 2.43 ± 0.03 | 5.43 | 2.4 ± 0.1 | 3.9 |
| SM | 0.66 ± 0.02 | 1.48 | 0.81 ± 0.03 | 1.28 |
| CL | 0.145 ± 0.005 | 0.33 | 0.64 ± 0.06 | 1.0 |
| Sum polar lipids | 27.3 ± 0.5 | 61.0 | 56.0 ± 0.7 | 88.8 |
| FFA | 15.1 ± 0.2 | 33.72 | 4.30 ± 0.07 | 6.8 |
| CHOL | 2.36 ± 0.03 | 5.27 | 2.64 ± 0.04 | 4.20 |
| TAG | <0.01 | <0.01 | 0.11 ± 0.03 | 0.18 |
| DAG | <0.01 | <0.01 | <0.01 | <0.01 |
| CE | <0.01 | <0.01 | <0.01 | <0.01 |
| Sum neutral lipids | 17.4 ± 0.2 | 39.0 | 7.1 ± 0.1 | 11.2 |
| Sum lipids | 44.7 ± 0.7 | 63.0 ± 0.7 | ||
| Polar lipid:FFA ratio | 1.809 ± 0.008 | 13.0 ± 0.2 | ||
| Fatty Acid (mg/g) | Fresh Cod | Pork | Pork n-3 TAG | Pork n-3 PL |
|---|---|---|---|---|
| Polar lipid fraction | ||||
| Sum SFA | 1.20 ± 0.02 | 0.76 ± 0.01 | 1.64 ± 0.07 | 2.12 ± 0.08 |
| Sum MUFA | 0.73 ± 0.03 | 0.474 ± 0.007 | 1.20 ± 0.06 | 1.014 ± 0.006 |
| LA 18:2n-6 | 0.100 ± 0.009 | 0.76 ± 0.01 | 1.07 ± 0.04 | 0.91 ± 0.03 |
| ARA 20:4n-6 | 0.097 ± 0.002 | 0.236 ± 0.005 | 0.257 ± 0.009 | 0.312 ± 0.008 |
| Sum n-6 | 0.22 ± 0.01 | 1.06 ± 0.02 | 1.42 ± 0.05 | 1.23 ± 0.04 |
| ALA 18:3n-3 | 0.020 ± 0.001 | 0.0240 ± 0.0001 | 0.067 ± 0.003 | 0.037 ± 0.006 |
| EPA 20:5n-3 | 0.690 ± 0.007 | 0.02763 ± 0.0004 | 0.043 ± 0.003 | 0.74 ± 0.06 |
| DHA 22:6n-3 | 1.78 ± 0.01 | 0.015 ± 0.001 | 0.037 ± 0.003 | 2.0 ± 0.1 |
| Sum EPA+DHA | 2.47 ± 0.02 | 0.043 ± 0.001 | 0.080 ± 0.005 | 2.7 ± 0.2 |
| Sum n-3 | 2.61 ± 0.03 | 0.130 ± 0.002 | 0.22 ± 0.01 | 2.8 ± 0.2 |
| Sum identified FAs | 4.8 ± 0.1 | 2.44 ± 0.04 | 4.5 ± 0.2 | 7.2 ± 0.3 |
| n-6:n-3 ratio | 0.085 ± 0.005 | 8.2 ± 0.2 | 6.4 ± 0.1 | 0.43 ± 0.03 |
| EPA:DHA ratio | 0.388 ± 0.001 | 1.80 ± 0.05 | 2.2 ± 0.3 | 0.376 ± 0.008 |
| ARA:EPA ratio | 0.14 ± 0.001 | 8.53 ± 0.04 | 6.0 ± 0.3 | 0.43 ± 0.04 |
| Neutral lipid fraction | ||||
| Sum SFA | 76 ± 1 | 76 ± 2 | 71 ± 2 | 72 ± 0.5 |
| Sum MUFA | 58 ± 2 | 60 ± 2 | 63 ± 2 | 57.7 ± 0.8 |
| LA 18:2n-6 | 24 ± 1 | 23.1 ± 0.4 | 22.1 ± 0.7 | 22.5 ± 0.5 |
| ARA 20:4n-6 | 0.188 ± 0.007 | 0.302 ± 0.009 | 0.36 ± 0.02 | 0.32 ± 0.02 |
| Sum n-6 | 24 ± 1 | 23.7 ± 0.4 | 23.0 ± 0.8 | 23.0 ± 0.5 |
| ALA 18:3n-3 | 4.0 ± 0.2 | 3.8 ± 0.2 | 3.7 ± 0.1 | 3.86 ± 0.04 |
| EPA 20:5n-3 | 0.15 ± 0.01 | <0.01 | 1.17 ± 0.06 | 0.15 ± 0.01 |
| DHA 22:6n-3 | 0.16 ± 0.01 | <0.01 | 1.64 ± 0.09 | 0.096 ± 0.004 |
| Sum EPA+DHA | 0.31 ± 0.01 | <0.01 | 2.8 ± 0.2 | 0.24 ± 0.01 |
| Sum n-3 | 4.7 ± 0.2 | 4.1 ± 0.2 | 7.3 ± 0.3 | 4.51 ± 0.05 |
| Sum identified FAs | 164 ± 5 | 163 ± 4 | 164 ± 4 | 157 ± 2 |
| n-6:n-3 ratio | 5.2 ± 0.1 | 5.8 ± 0.3 | 3.2 ± 0.2 | 5.00 ± 0.06 |
| EPA:DHA ratio | 0.93 ± 0.15 | * | 0.725 ± 0.007 | 1.6 ± 0.1 |
| ARA:EPA ratio | 1.3 ± 0.1 | * | 0.31 ± 0.03 | 2.1 ± 0.1 |
| Fatty Acid (mg/g) | Fresh Cod | Pork | Pork n-3 TAG | Pork n-3 PL |
|---|---|---|---|---|
| Sum SFA | 1.56 ± 0.06 | 1.51 ± 0.04 | 1.47 ± 0.05 | 1.48 ± 0.04 |
| Sum MUFA | 0.69 ± 0.03 | 0.70 ± 0.02 | 0.67 ± 0.01 | 0.65 ± 0.02 |
| LA 18:2n-6 | 0.48 ± 0.03 | 0.46 ± 0.02 | 0.44 ± 0.03 | 0.43 ± 0.02 |
| ARA 20:4n-6 | 0.41 ± 0.01 a | 0.66 ± 0.01 b | 0.45 ± 0.01 ac | 0.464 ± 0.009 c |
| Sum n-6 | 0.99 ± 0.05 a | 1.27 ± 0.03 b | 0.97 ± 0.03 a | 0.99 ± 0.02 a |
| ALA 18:3n-3 | <0.01 | <0.01 | <0.01 | <0.01 |
| EPA 20:5n-3 | 0.119 ± 0.005 a | 0.031 ± 0.001 b | 0.127 ± 0.004 a | 0.095 ± 0.002 c |
| DHA 22:6n-3 | 0.39 ± 0.02 a | 0.224 ± 0.007 b | 0.354 ± 0.009 c | 0.381 ± 0.007 ac |
| Sum EPA+DHA | 0.51 ± 0.02 a | 0.255 ± 0.008 b | 0.48 ± 0.01 a | 0.477 ± 0.008 a |
| Sum n-3 | 0.57 ± 0.02 a | 0.313 ± 0.008 b | 0.55 ± 0.01 a | 0.535 ± 0.009 a |
| Sum identified FAs | 3.8 ± 0.1 | 3.80 ± 0.09 | 3.7 ± 0.1 | 3.66 ± 0.08 |
| n-6:n-3 ratio | 1.73 ± 0.03 a | 4.08 ± 0.09 b | 1.76 ± 0.03 a | 1.85 ± 0.04 a |
| ARA:EPA ratio | 3.49 ± 0.09 a | 21.7 ± 0.8 b | 3.6 ± 0.1 a | 4.9 ± 0.2 c |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fauske, K.R.; Bernhard, A.; Fjære, E.; Myrmel, L.S.; Frøyland, L.; Kristiansen, K.; Liaset, B.; Madsen, L. Effects of Frozen Storage on Phospholipid Content in Atlantic Cod Fillets and the Influence on Diet-Induced Obesity in Mice. Nutrients 2018, 10, 695. https://0-doi-org.brum.beds.ac.uk/10.3390/nu10060695
Fauske KR, Bernhard A, Fjære E, Myrmel LS, Frøyland L, Kristiansen K, Liaset B, Madsen L. Effects of Frozen Storage on Phospholipid Content in Atlantic Cod Fillets and the Influence on Diet-Induced Obesity in Mice. Nutrients. 2018; 10(6):695. https://0-doi-org.brum.beds.ac.uk/10.3390/nu10060695
Chicago/Turabian StyleFauske, Kristin Røen, Annette Bernhard, Even Fjære, Lene Secher Myrmel, Livar Frøyland, Karsten Kristiansen, Bjørn Liaset, and Lise Madsen. 2018. "Effects of Frozen Storage on Phospholipid Content in Atlantic Cod Fillets and the Influence on Diet-Induced Obesity in Mice" Nutrients 10, no. 6: 695. https://0-doi-org.brum.beds.ac.uk/10.3390/nu10060695