Sicilian Litchi Fruit Extracts Induce Autophagy versus Apoptosis Switch in Human Colon Cancer Cells



 ,
,  , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Preparation of Litchi Fruit Extracts
2.3. Cell Cultures
2.4. Evaluation of Polyphenol Contents
2.5. Evaluation of Cell Viability
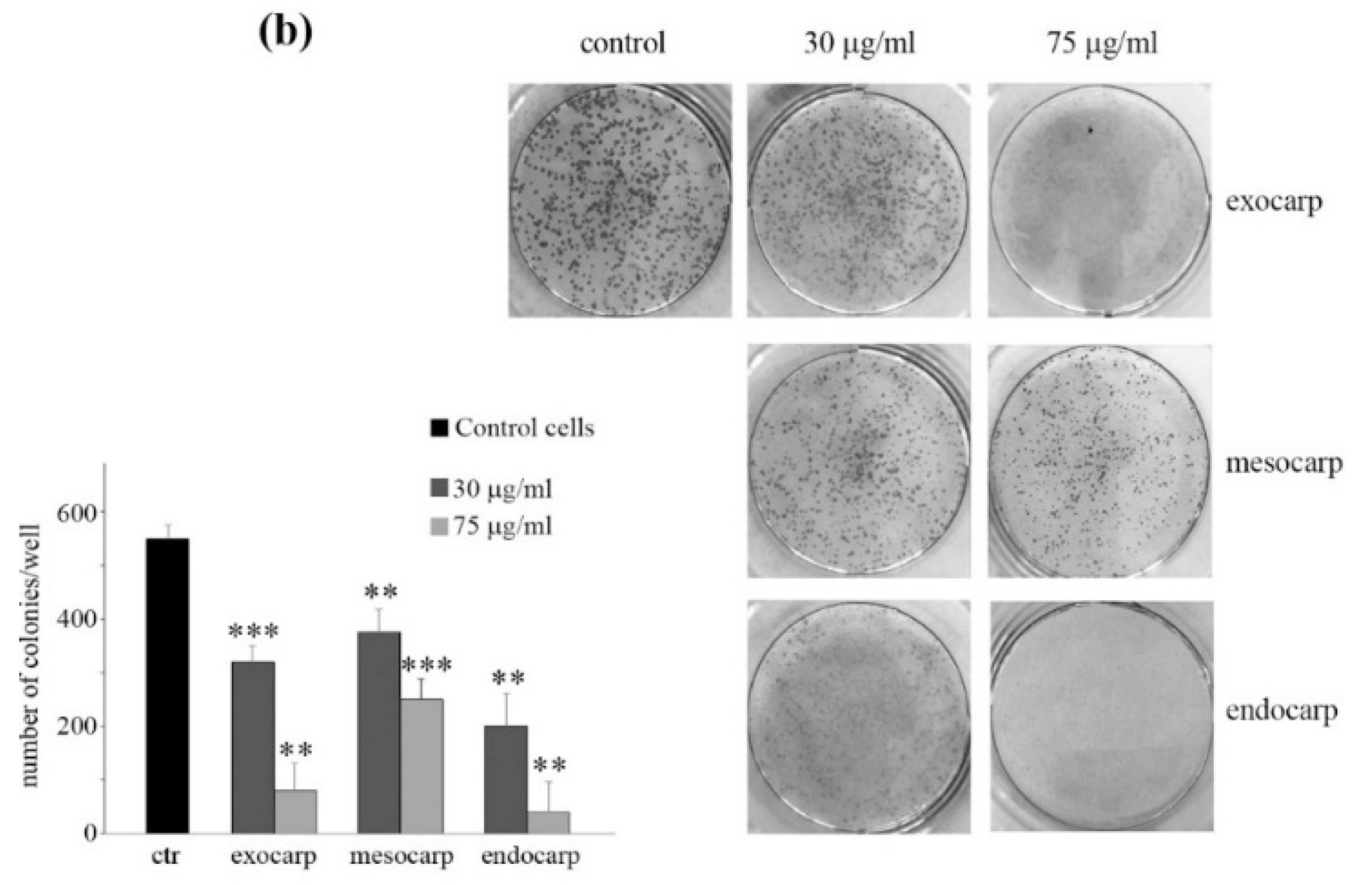
2.6. Clonogenic Assay
2.7. Flow Cytometry Analyses
2.8. Monodansylcadaverine (MDC) Labeling
2.9. Western Blotting Analysis
2.10. Gene Silencing
2.11. Reversed Phase HPLC, MS and MS/MS Experiments
2.12. Statistical Analysis
3. Results
3.1. Quantification of Polyphenol Content in Litchi Extracts
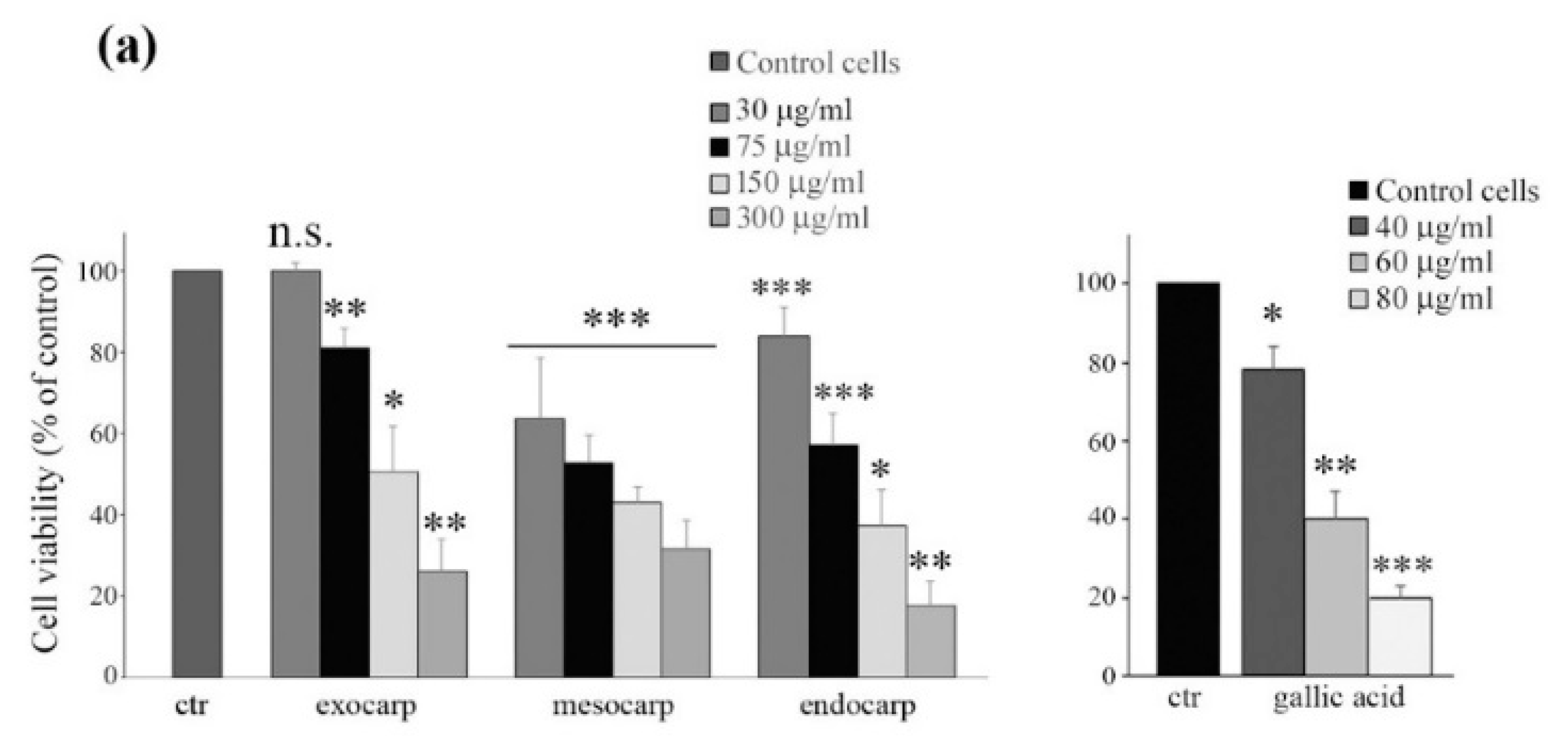
3.2. Litchi Extracts Induce Cell Viability Reduction and Growth Inhibition in Colon Cancer Cells
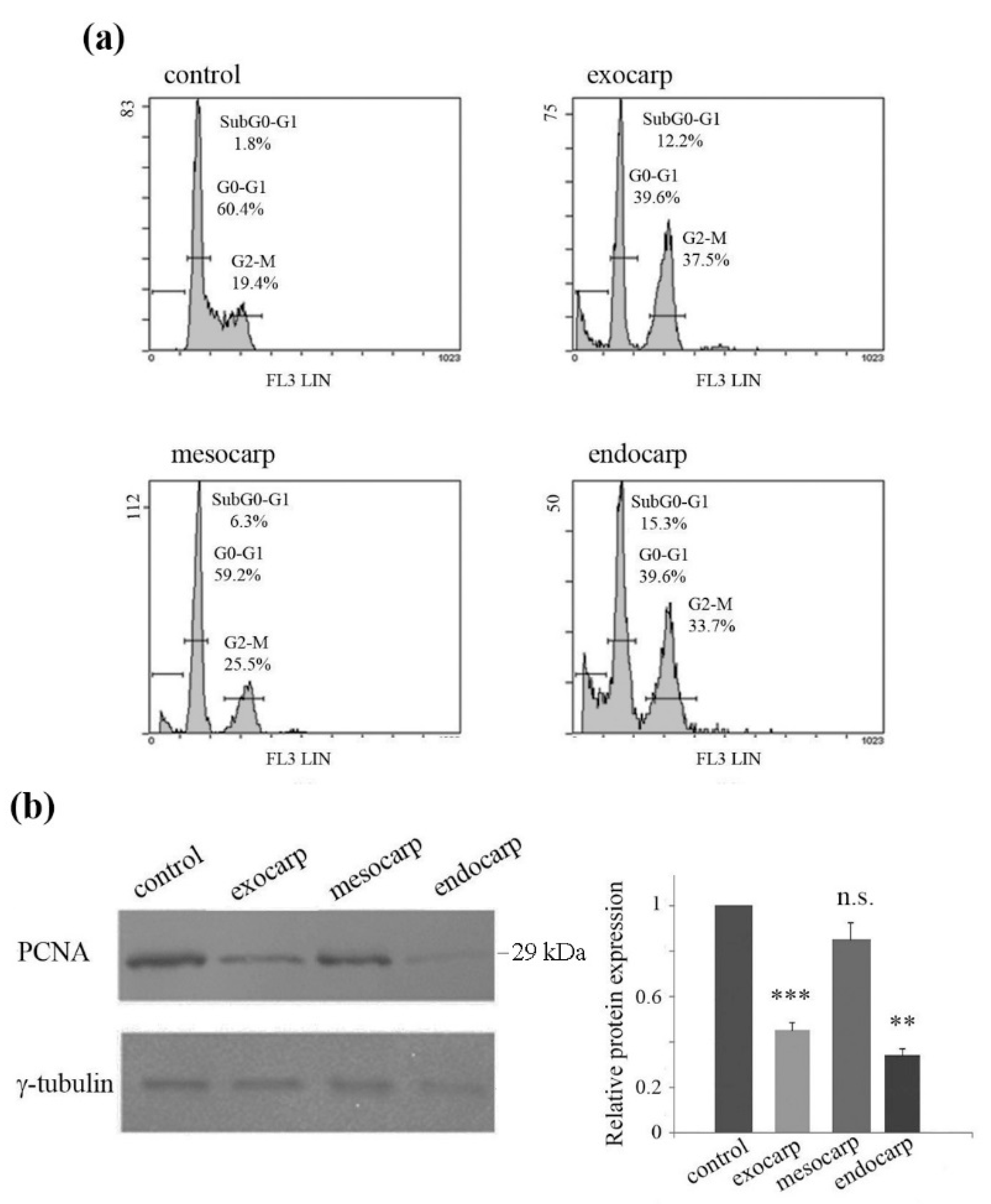
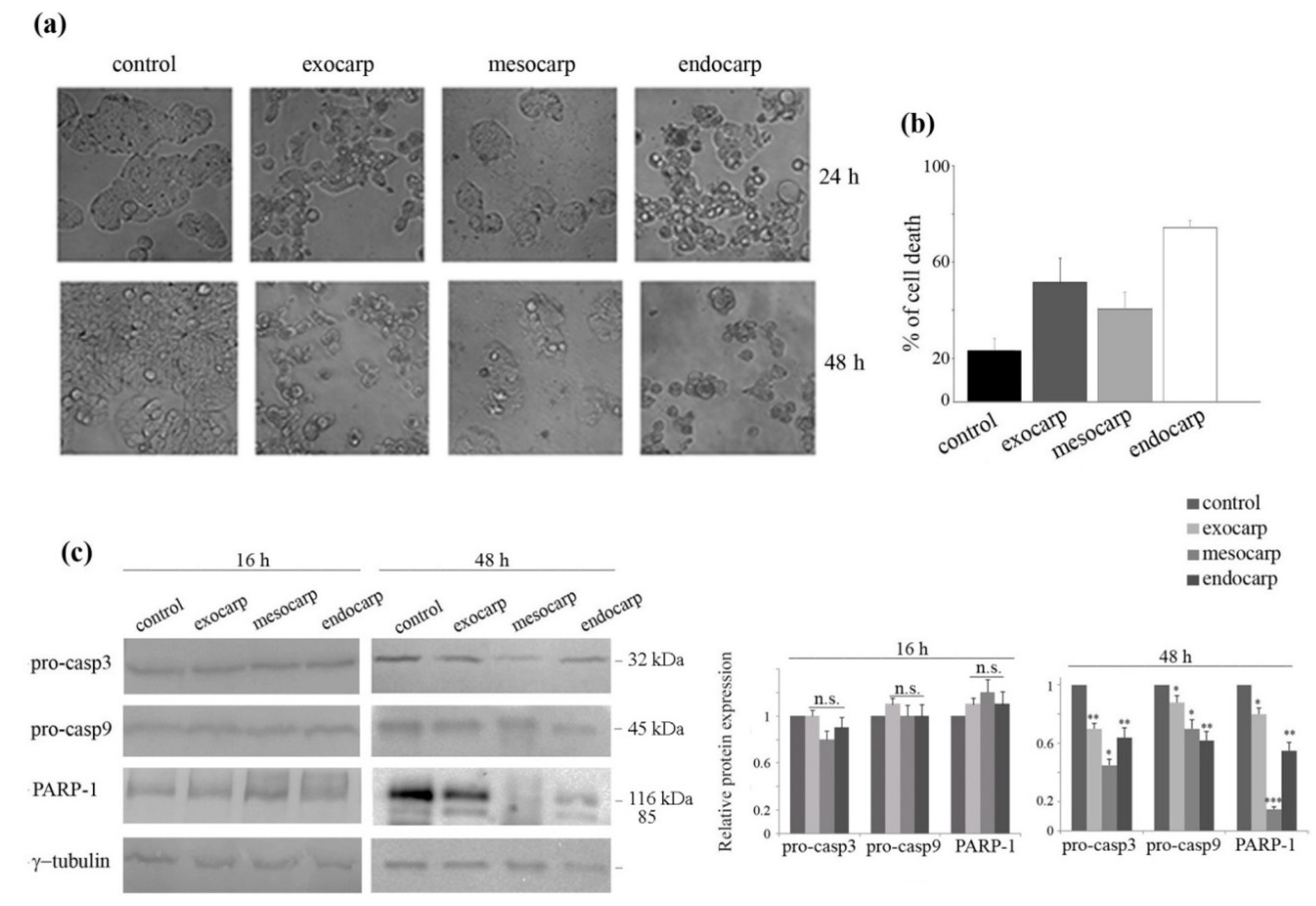
3.3. Growth Inhibition of Colon Cancer Cells is Due to Cell Cycle Arrest and Apoptotic Cell Death
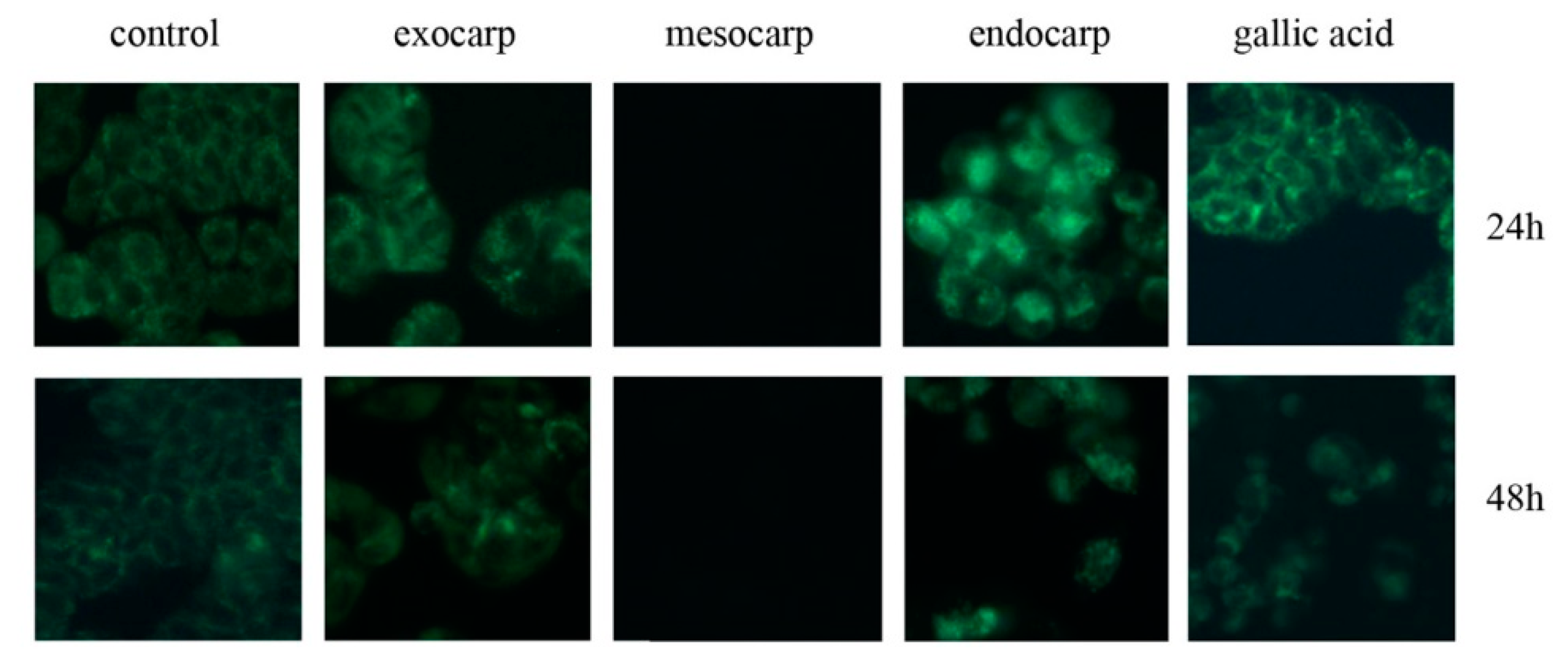
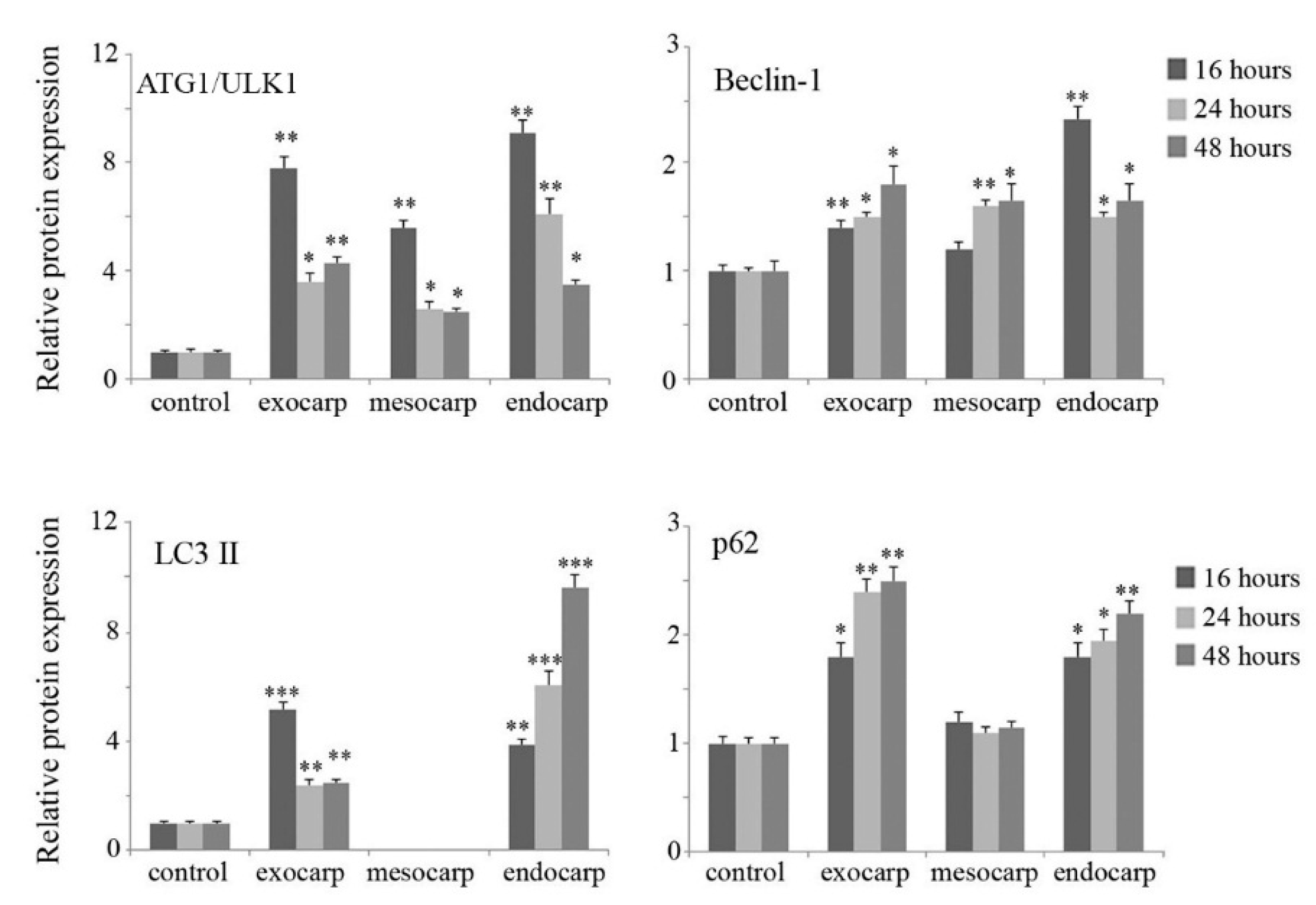
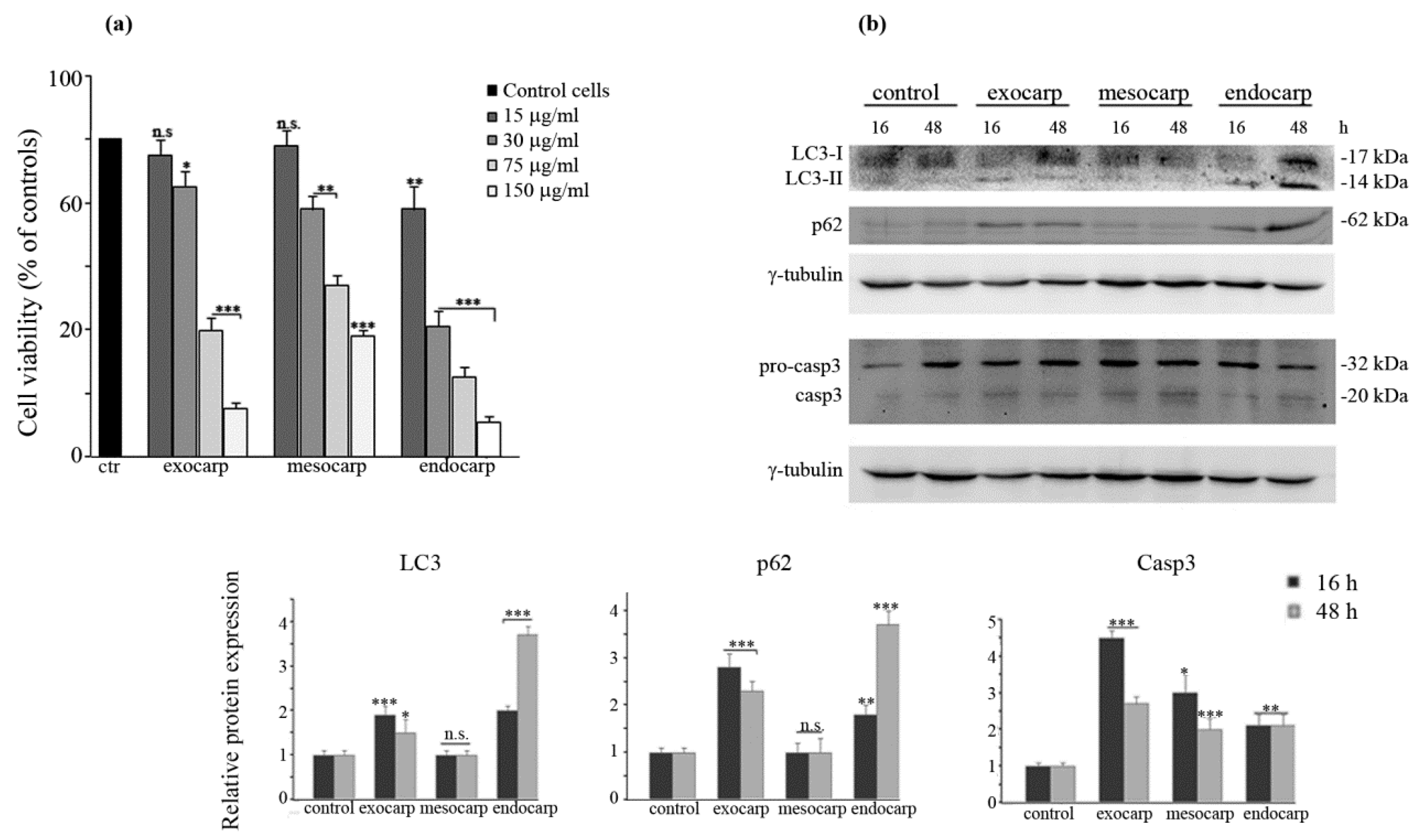
3.4. Induction of Autophagic Flux by Litchi Exocarp and Endocarp
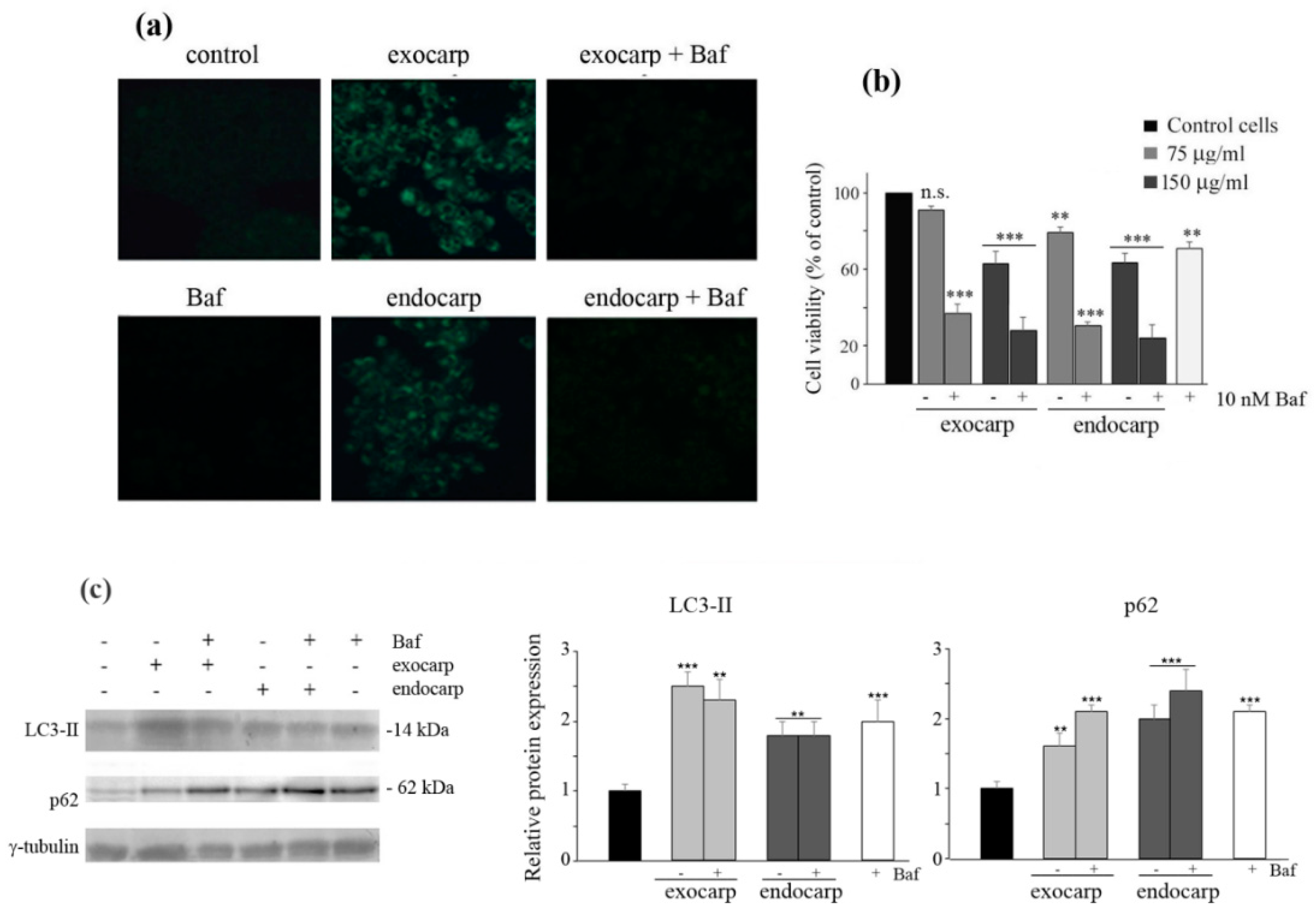
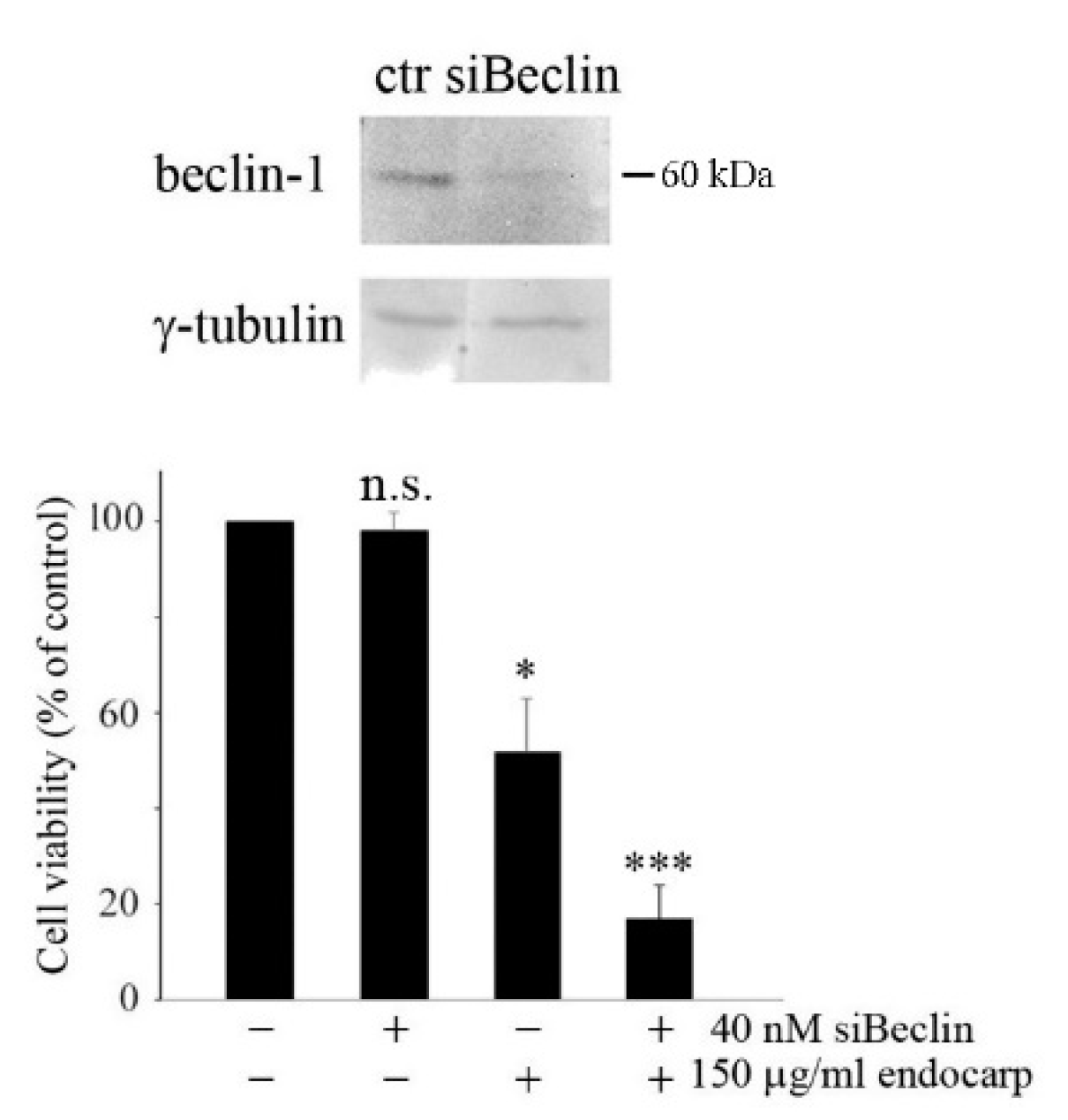
3.5. Inhibiting Autophagy Promotes Cell Death
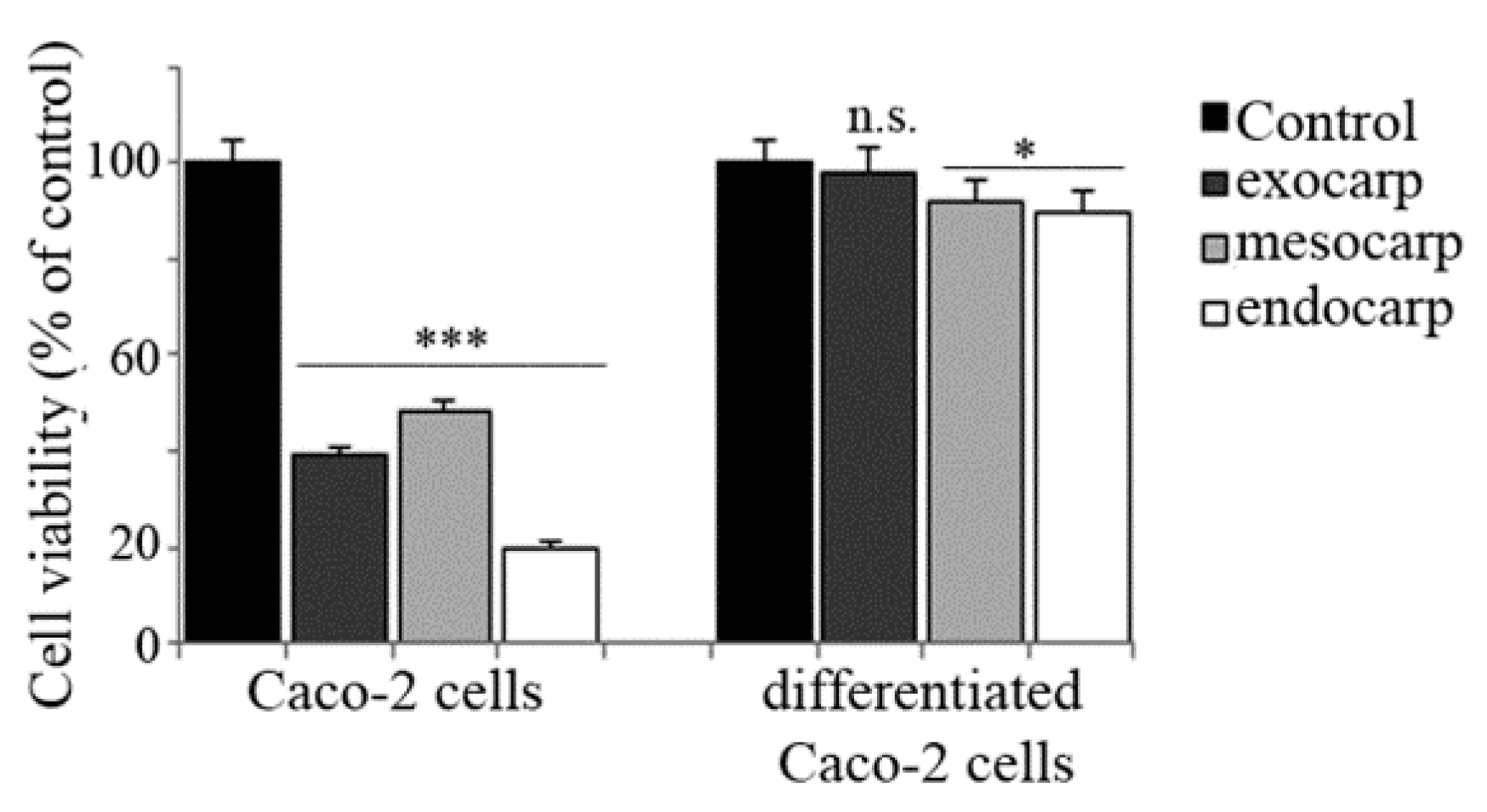
3.6. Effects of Litchi Extracts on Other Colon Cancer Cells and Non Tumor Counterparts
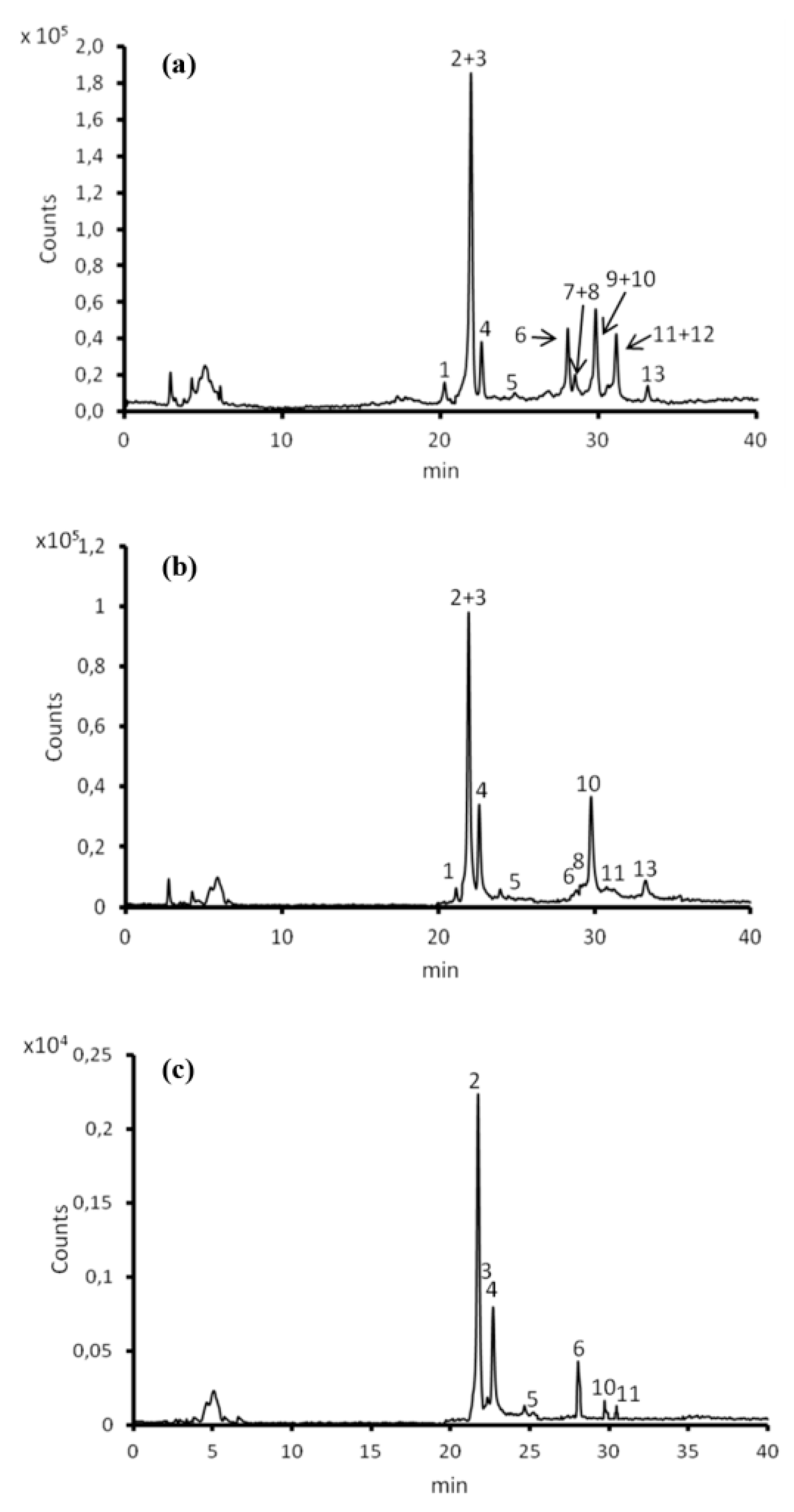
3.7. Contents of Polyphenols in Litchi Exocarp, Mesocarp and Endocarp Extracts and Their Individual Composition
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Menzel, C.M.; Waite, G.K. (Eds.) Litchi and Longan: Botany, Production, and Uses; CABI Pub: Wallingford, UK; Cambridge, MA, USA, 2005; ISBN 978-0-85199-696-7. [Google Scholar]
- Kilari, E.; Putta, S. Biological and phytopharmacological descriptions of Litchi chinensis. Pharmacogn. Rev. 2016, 10, 60. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.R.M.; Mohamed, G.A. Litchi chinensis: Medicinal uses, phytochemistry, and pharmacology. J. Ethnopharmacol. 2015, 174, 492–513. [Google Scholar] [CrossRef] [PubMed]
- Lauricella, M.; Emanuele, S.; Calvaruso, G.; Giuliano, M.; D’Anneo, A. Multifaceted Health Benefits of Mangifera indica L. (Mango): The Inestimable Value of Orchards Recently Planted in Sicilian Rural Areas. Nutrients 2017, 9, 525. [Google Scholar] [CrossRef] [PubMed]
- Testa, R.; Tudisca, S.; Schifani, G.; Di Trapani, A.; Migliore, G. Tropical Fruits as an Opportunity for Sustainable Development in Rural Areas: The Case of Mango in Small-Sized Sicilian Farms. Sustainability 2018, 10, 1436. [Google Scholar] [CrossRef]
- Hu, X.; Wang, J.; Jing, Y.; Song, L.; Zhu, J.; Cui, X.; Yu, R. Structural elucidation and in vitro antioxidant activities of a new heteropolysaccharide from Litchi chinensis. Drug Discov. Ther. 2015, 9, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Zhang, R.; Dong, L.; Guo, J.; Deng, Y.; Yi, Y.; Zhang, M. Antioxidant and antiproliferative activities of polysaccharide fractions from litchi pulp. Food Funct. 2015, 6, 2598–2606. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Zhang, R.; Liu, Y.; Xiao, J.; Liu, L.; Wei, Z.; Yi, Y.; Zhang, M.; Liu, D. Dietary litchi pulp polysaccharides could enhance immunomodulatory and antioxidant effects in mice. Int. J. Biol. Macromol. 2016, 92, 1067–1073. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.-P.; Lin, C.-C.; Huang, C.-C.; Lin, Y.-H.; Chou, J.-C.; Tsia, Y.-T.; Su, J.-R.; Chung, Y.-C. Induction of apoptosis and cell cycle arrest in human colorectal carcinoma by Litchi seed extract. J. Biomed. Biotechnol. 2012, 2012, 341479. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Jiang, Y. Litchi flavonoids: Isolation, identification and biological activity. Mol. Basel Switz. 2007, 12, 745–758. [Google Scholar] [CrossRef]
- Wang, X.; Yuan, S.; Wang, J.; Lin, P.; Liu, G.; Lu, Y.; Zhang, J.; Wang, W.; Wei, Y. Anticancer activity of litchi fruit pericarp extract against human breast cancer in vitro and in vivo. Toxicol. Appl. Pharmacol. 2006, 215, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Swart, C.; Du Toit, A.; Loos, B. Autophagy and the invisible line between life and death. Eur. J. Cell Biol. 2016, 95, 598–610. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, T.; Mizushima, N. The ULK complex initiates autophagosome formation at phosphatidylinositol synthase-enriched ER subdomains. Autophagy 2017, 13, 1795–1796. [Google Scholar] [CrossRef] [PubMed]
- Pellerito, O.; Notaro, A.; Sabella, S.; De Blasio, A.; Vento, R.; Calvaruso, G.; Giuliano, M. WIN induces apoptotic cell death in human colon cancer cells through a block of autophagic flux dependent on PPARγ down-regulation. Apoptosis 2014, 19, 1029–1042. [Google Scholar] [CrossRef] [PubMed]
- Notaro, A.; Sabella, S.; Pellerito, O.; Di Fiore, R.; De Blasio, A.; Vento, R.; Calvaruso, G.; Giuliano, M. Involvement of PAR-4 in cannabinoid-dependent sensitization of osteosarcoma cells to TRAIL-induced apoptosis. Int. J. Biol. Sci. 2014, 10, 466–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amaravadi, R.; Kimmelman, A.C.; White, E. Recent insights into the function of autophagy in cancer: Table 1. Genes Dev. 2016, 30, 1913–1930. [Google Scholar] [CrossRef] [PubMed]
- White, E. The role for autophagy in cancer. J. Clin. Investig. 2015, 125, 42–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Degenhardt, K.; Mathew, R.; Beaudoin, B.; Bray, K.; Anderson, D.; Chen, G.; Mukherjee, C.; Shi, Y.; Gélinas, C.; Fan, Y.; et al. Autophagy promotes tumor cell survival and restricts necrosis, inflammation, and tumorigenesis. Cancer Cell 2006, 10, 51–64. [Google Scholar] [CrossRef] [PubMed]
- Qu, X.; Yu, J.; Bhagat, G.; Furuya, N.; Hibshoosh, H.; Troxel, A.; Rosen, J.; Eskelinen, E.-L.; Mizushima, N.; Ohsumi, Y.; et al. Promotion of tumorigenesis by heterozygous disruption of the beclin 1 autophagy gene. J. Clin. Investig. 2003, 112, 1809–1820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, L.; Wang, Z.; Wang, Y.; Guo, D.; Yang, J.; Chen, L.; Tan, N. Natural Cyclopeptide RA-XII, a New Autophagy Inhibitor, Suppresses Protective Autophagy for Enhancing Apoptosis through AMPK/mTOR/P70S6K Pathways in HepG2 Cells. Molecules 2017, 22, 1934. [Google Scholar] [CrossRef] [PubMed]
- Portanova, P.; Russo, T.; Pellerito, O.; Calvaruso, G.; Giuliano, M.; Vento, R.; Tesoriere, G. The role of oxidative stress in apoptosis induced by the histone deacetylase inhibitor suberoylanilide hydroxamic acid in human colon adenocarcinoma HT-29 cells. Int. J. Oncol. 2008, 33, 325–331. [Google Scholar] [PubMed]
- Natoli, M.; Leoni, B.D.; D’Agnano, I.; Zucco, F.; Felsani, A. Good Caco-2 cell culture practices. Toxicol. In Vitro 2012, 26, 1243–1246. [Google Scholar] [CrossRef] [PubMed]
- Cicco, N.; Lanorte, M.T.; Paraggio, M.; Viggiano, M.; Lattanzio, V. A reproducible, rapid and inexpensive Folin–Ciocalteu micro-method in determining phenolics of plant methanol extracts. Microchem. J. 2009, 91, 107–110. [Google Scholar] [CrossRef]
- Lauricella, M.; Carlisi, D.; Giuliano, M.; Calvaruso, G.; Cernigliaro, C.; Vento, R.; D’Anneo, A. The analysis of estrogen receptor-α positive breast cancer stem-like cells unveils a high expression of the serpin proteinase inhibitor PI-9: Possible regulatory mechanisms. Int. J. Oncol. 2016, 49, 352–360. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Luo, H.; Yuan, H.; Xia, Y.; Shu, P.; Huang, X.; Lu, Y.; Liu, X.; Keller, E.T.; Sun, D.; et al. Litchi seed extracts diminish prostate cancer progression via induction of apoptosis and attenuation of EMT through Akt/GSK-3β signaling. Sci. Rep. 2017, 7, 41656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campanella, C.; D’Anneo, A.; Marino Gammazza, A.; Caruso Bavisotto, C.; Barone, R.; Emanuele, S.; Lo Cascio, F.; Mocciaro, E.; Fais, S.; De Macario, E.C.; et al. The histone deacetylase inhibitor SAHA induces HSP60 nitration and its extracellular release by exosomal vesicles in human lung-derived carcinoma cells. Oncotarget 2016, 7, 28849–28867. [Google Scholar] [CrossRef] [PubMed]
- Kouroku, Y.; Fujita, E.; Tanida, I.; Ueno, T.; Isoai, A.; Kumagai, H.; Ogawa, S.; Kaufman, R.J.; Kominami, E.; Momoi, T. ER stress (PERK/eIF2alpha phosphorylation) mediates the polyglutamine-induced LC3 conversion, an essential step for autophagy formation. Cell Death Differ. 2007, 14, 230–239. [Google Scholar] [CrossRef] [PubMed]
- Bjørkøy, G.; Lamark, T.; Pankiv, S.; Øvervatn, A.; Brech, A.; Johansen, T. Monitoring autophagic degradation of p62/SQSTM1. Methods Enzymol. 2009, 452, 181–197. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Liang, H.; Zhang, M.-W.; Zhang, R.-F.; Deng, Y.-Y.; Wei, Z.-C.; Zhang, Y.; Tang, X.-J. Phenolic profiles and antioxidant activity of litchi (Litchi chinensis Sonn.) fruit pericarp from different commercially available cultivars. Molecules 2012, 17, 14954–14967. [Google Scholar] [CrossRef] [PubMed]
- Emanuele, S.; Lauricella, M.; Calvaruso, G.; D’Anneo, A.; Giuliano, M. Litchi chinensis as a Functional Food and a Source of Antitumor Compounds: An Overview and a Description of Biochemical Pathways. Nutrients 2017, 9, 992. [Google Scholar] [CrossRef] [PubMed]
- Niedzwiecki, A.; Roomi, M.W.; Kalinovsky, T.; Rath, M. Anticancer Efficacy of Polyphenols and Their Combinations. Nutrients 2016, 8, 552. [Google Scholar] [CrossRef] [PubMed]
- Yamanishi, R.; Yoshigai, E.; Okuyama, T.; Mori, M.; Murase, H.; Machida, T.; Okumura, T.; Nishizawa, M. The anti-inflammatory effects of flavanol-rich lychee fruit extract in rat hepatocytes. PLoS ONE 2014, 9, e93818. [Google Scholar] [CrossRef] [PubMed]
- Pietrocola, F.; Mariño, G.; Lissa, D.; Vacchelli, E.; Malik, S.A.; Niso-Santano, M.; Zamzami, N.; Galluzzi, L.; Maiuri, M.C.; Kroemer, G. Pro-autophagic polyphenols reduce the acetylation of cytoplasmic proteins. Cell Cycle 2012, 11, 3851–3860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alegre, F.; Moragrega, Á.B.; Polo, M.; Marti-Rodrigo, A.; Esplugues, J.V.; Blas-Garcia, A.; Apostolova, N. Role of p62/SQSTM1 beyond autophagy: A lesson learned from drug-induced toxicity in vitro: Role of p62 in the hepatic effect of an antiretroviral drug. Br. J. Pharmacol. 2017. [Google Scholar] [CrossRef]
- Cohen-Kaplan, V.; Ciechanover, A.; Livneh, I. p62 at the crossroad of the ubiquitin-proteasome system and autophagy. Oncotarget 2016, 7, 83833–83834. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | RT * (min) | ESI ** (M − H)-(m/z) | Compound | Area % | |||
|---|---|---|---|---|---|---|---|
| Exo | Endo | Meso | |||||
| 1 | 20.24 | 577.1352 (M − H)− | Procyanidin B2 | 4.4 | 1.9 | - | Flavan-3-ol |
| 2 | 21.92 | 289.0716 (M − H)− | Epicatechin | 45.4 | 40.1 | 55.2 | Flavan-3-ol |
| 3 | 22.05 | 863.1824 (M − H)− | Epicatechin trimer | 1.0 | 1.7 | 0.6 | Flavan-3-ol |
| 4 | 22.58 | 863.1828 (M − H)− | Pavetannin B2 | 13.5 | 19.1 | 24.5 | Flavan-3-ol |
| 5 | 24.68 | 863.1832 (M − H)− | Epicatechin trimer | 0.7 | 1.3 | 0.9 | Flavan-3-ol |
| 6 | 28.05 | 609.1463 (M − H)− | Rutin (quercetin-3-O-rutinoside) | 7.2 | 2.4 | 14.7 | Flavonol glycoside |
| 7 | 28.46 | 593.1515 (M − H)− | Nicotiflorin (Kaempherol-3-rutinoside) | 1.9 | - | - | Flavonol glycoside |
| 8 | 28.48 | 577.1350 (M − H)− | Procyanidin B4 | 0.9 | 1.2 | - | Flavan-3-ol |
| 9 | 29.56 | 463.0893 (M − H)− | Quercetin-glucoside | 1.4 | - | - | Flavonol glycoside |
| 10 | 29.83 | 575.1191 (M − H)− | Procyanidin A2 | 16.4 | 25.1 | 2.7 | Flavan-3-ol |
| 11 | 30.53 | 863.1820 (M − H)− | Epicatechin trimer | 1.1 | 1.3 | 1.3 | Flavan-3-ol |
| 12 | 31.13 | 593.1505 (M − H)− 629.1276 (M + Cl − H)- | Antirhinin (Cyanidin-3-rutinoside) | 4.8 | - | - | Anthocyanin |
| 13 | 33.12 | 575.1188 (M − H)- | Procyanidin A isomer | 1.3 | 6.0 | - | Flavan-3-ol |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Emanuele, S.; Notaro, A.; Palumbo Piccionello, A.; Maggio, A.; Lauricella, M.; D’Anneo, A.; Cernigliaro, C.; Calvaruso, G.; Giuliano, M. Sicilian Litchi Fruit Extracts Induce Autophagy versus Apoptosis Switch in Human Colon Cancer Cells. Nutrients 2018, 10, 1490. https://0-doi-org.brum.beds.ac.uk/10.3390/nu10101490
Emanuele S, Notaro A, Palumbo Piccionello A, Maggio A, Lauricella M, D’Anneo A, Cernigliaro C, Calvaruso G, Giuliano M. Sicilian Litchi Fruit Extracts Induce Autophagy versus Apoptosis Switch in Human Colon Cancer Cells. Nutrients. 2018; 10(10):1490. https://0-doi-org.brum.beds.ac.uk/10.3390/nu10101490
Chicago/Turabian StyleEmanuele, Sonia, Antonietta Notaro, Antonio Palumbo Piccionello, Antonella Maggio, Marianna Lauricella, Antonella D’Anneo, Cesare Cernigliaro, Giuseppe Calvaruso, and Michela Giuliano. 2018. "Sicilian Litchi Fruit Extracts Induce Autophagy versus Apoptosis Switch in Human Colon Cancer Cells" Nutrients 10, no. 10: 1490. https://0-doi-org.brum.beds.ac.uk/10.3390/nu10101490