Comparative Proteomic Profiling of Tumor-Associated Proteins in Human Gastric Cancer Cells Treated with Pectolinarigenin

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagent
2.2. Cell Viability Assay
2.3. Preparation of the Cellular Extract for 2-DE
2.4. Separation of Proteins by 2-DE and Image Analysis
2.5. Matrix-Assisted Laser Desorption/Ionization-Time of Flight Mass Spectrometry (MALDI-TOF MS) Mass Spectrometry Analysis
2.6. Database Search for Protein Identification
2.7. Western Blot Analysis
2.8. Molecular Docking Studies
2.9. Protein-Protein Interaction & Gene Ontology (GO) Analysis
2.10. Statistical Analysis
3. Results
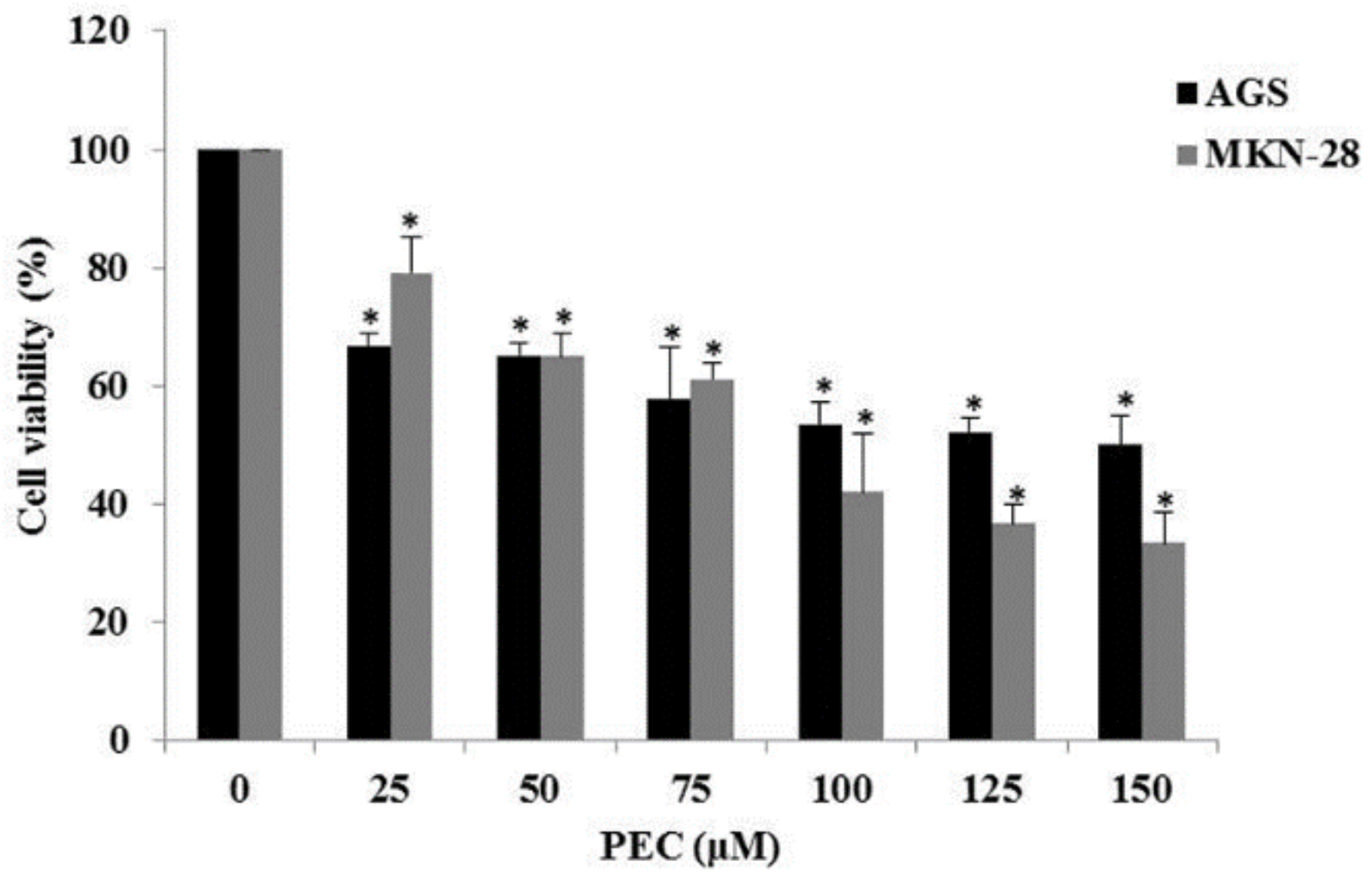
3.1. Pectolinarigenin (PEC) Reduced Cell Viability
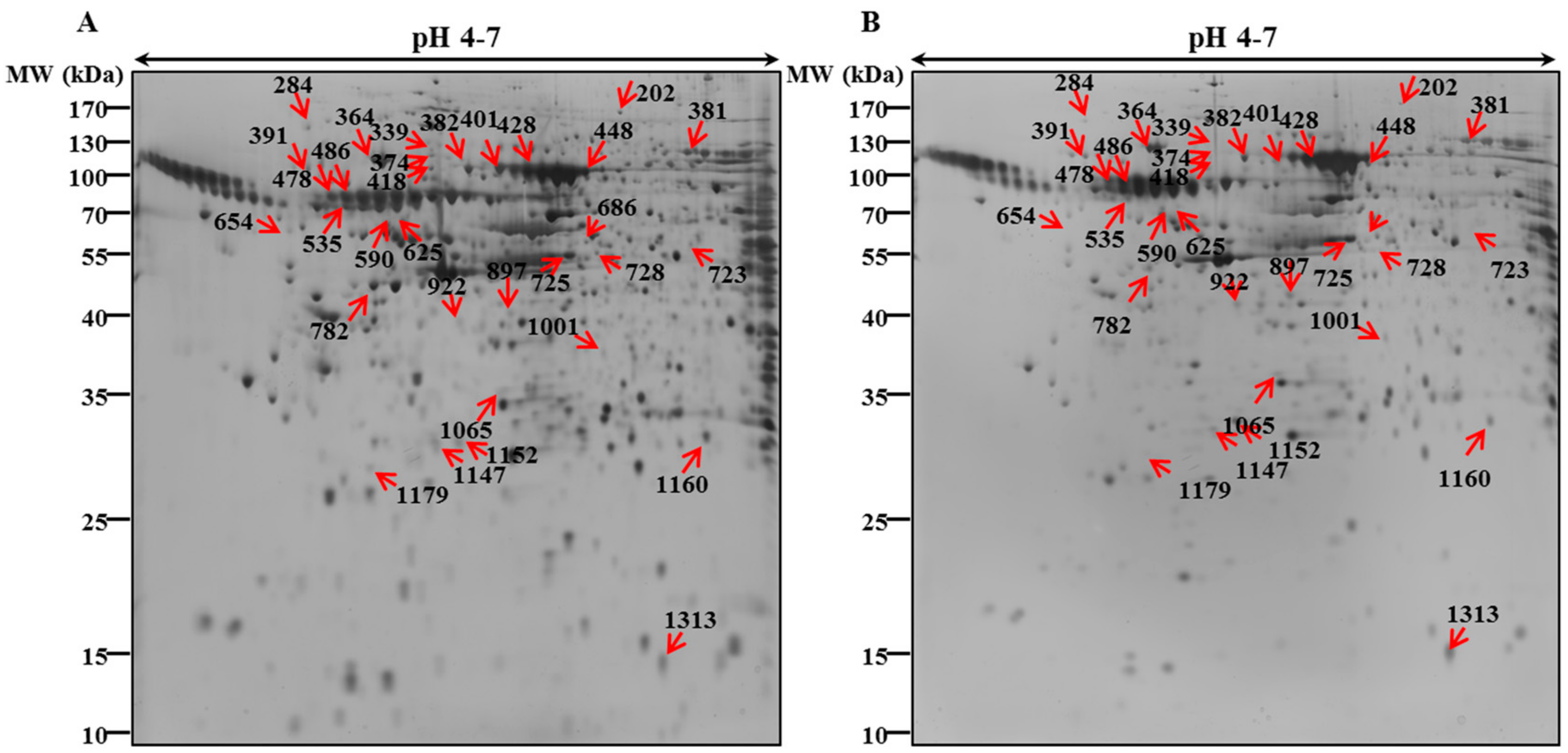
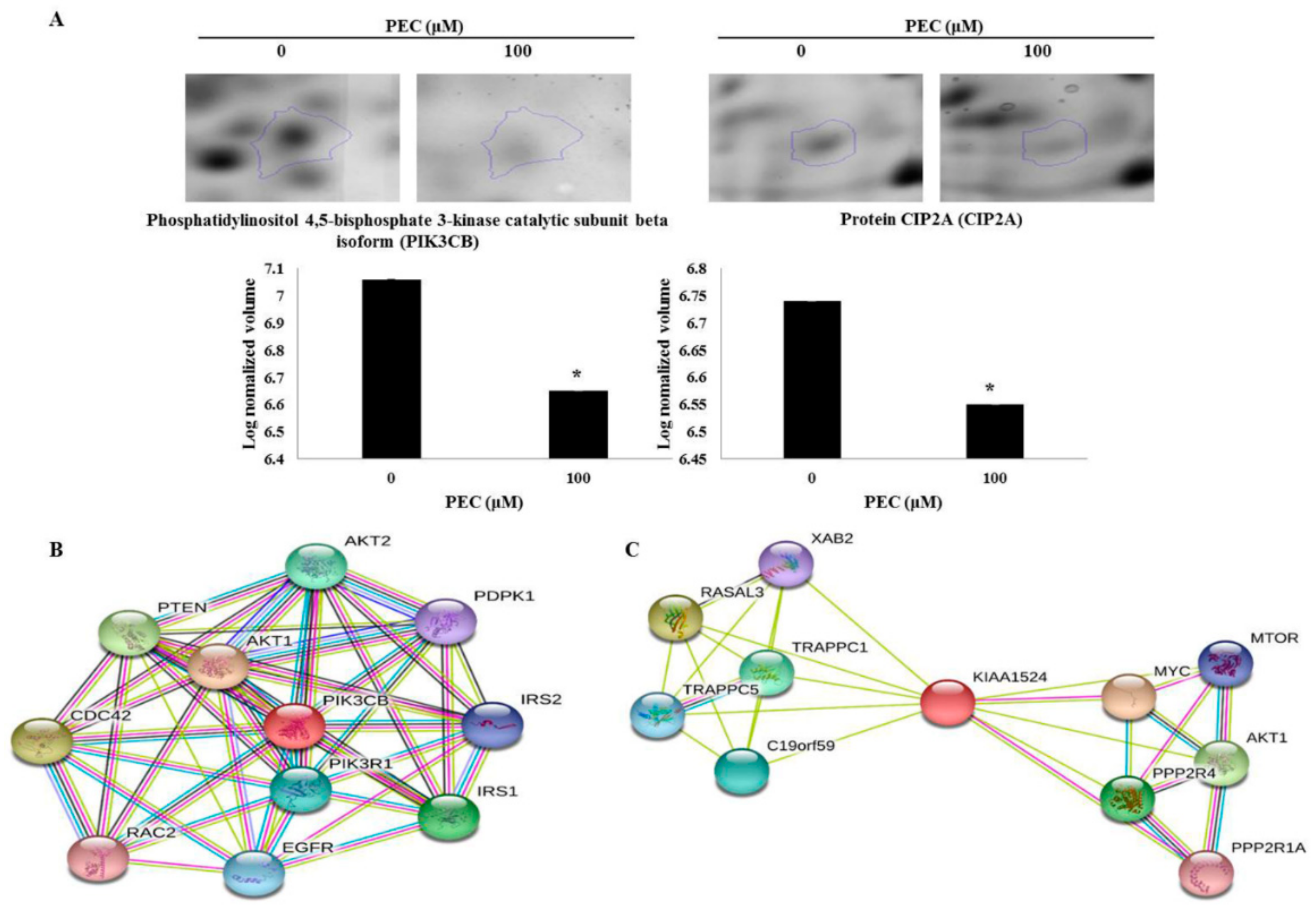
3.2. 2-DE Analysis and Protein Identification by MALDI/TOF-MS
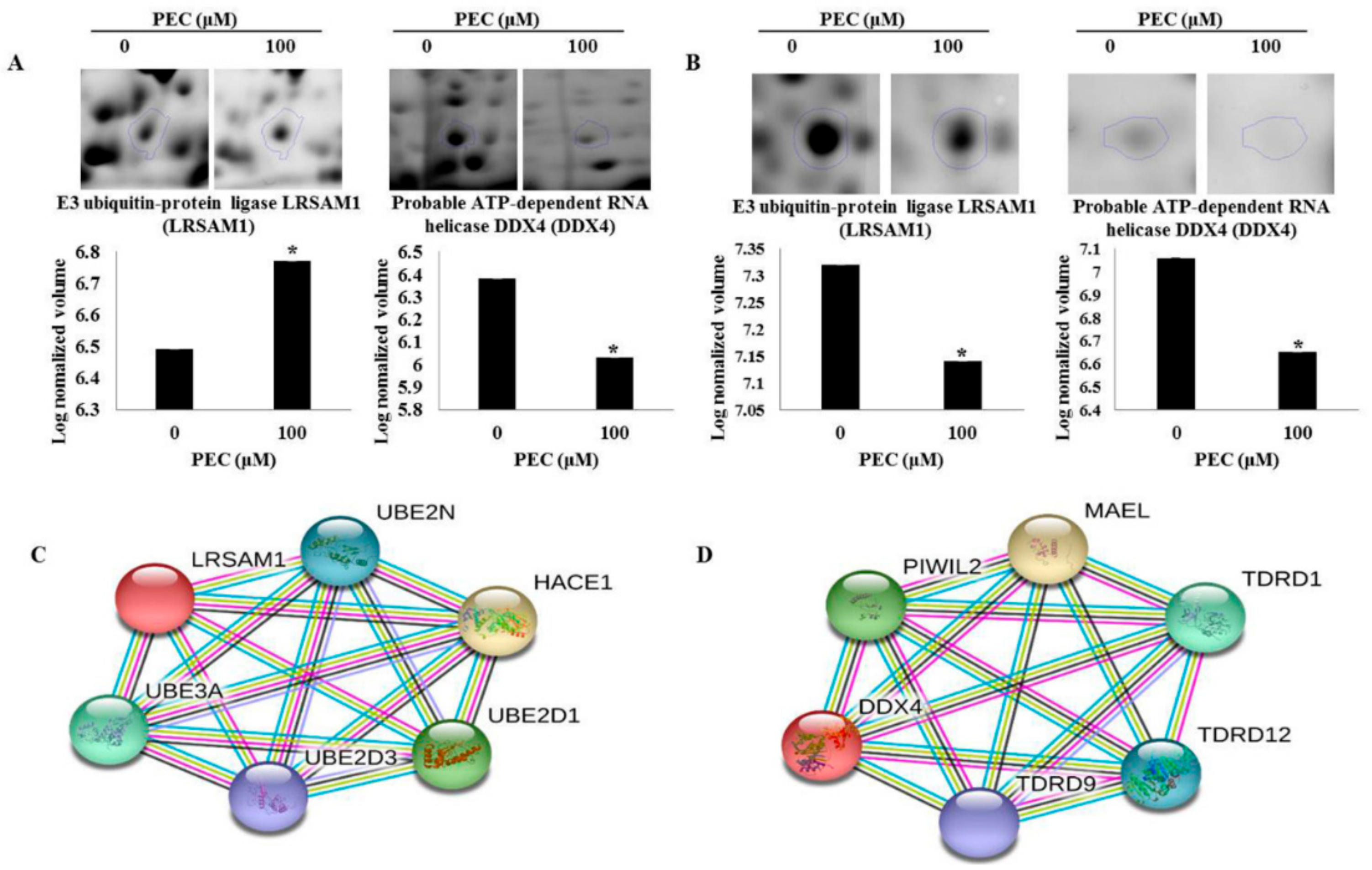
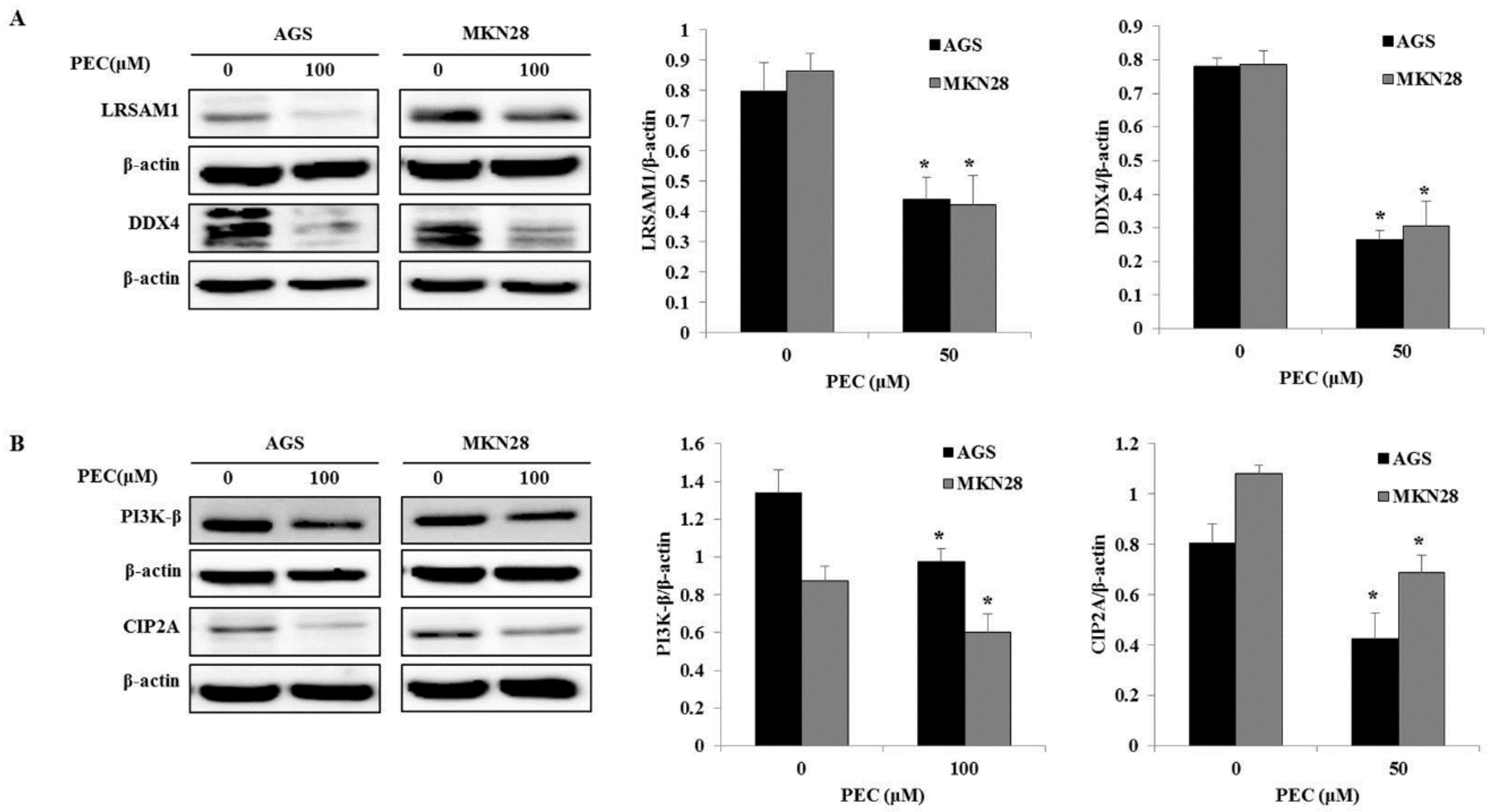
3.3. Validation of Differentially Expressed Proteins by Western Blot Analysis.
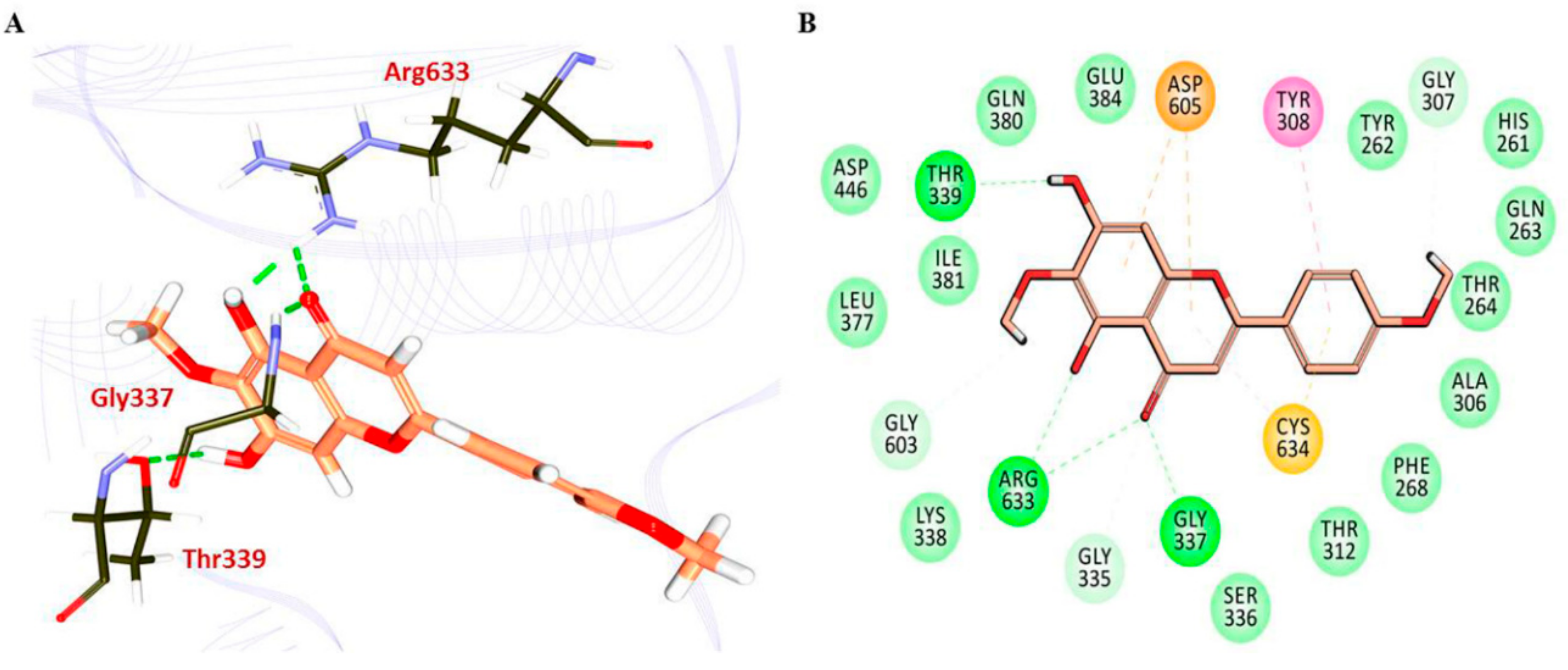
3.4. Molecular Docking Studies
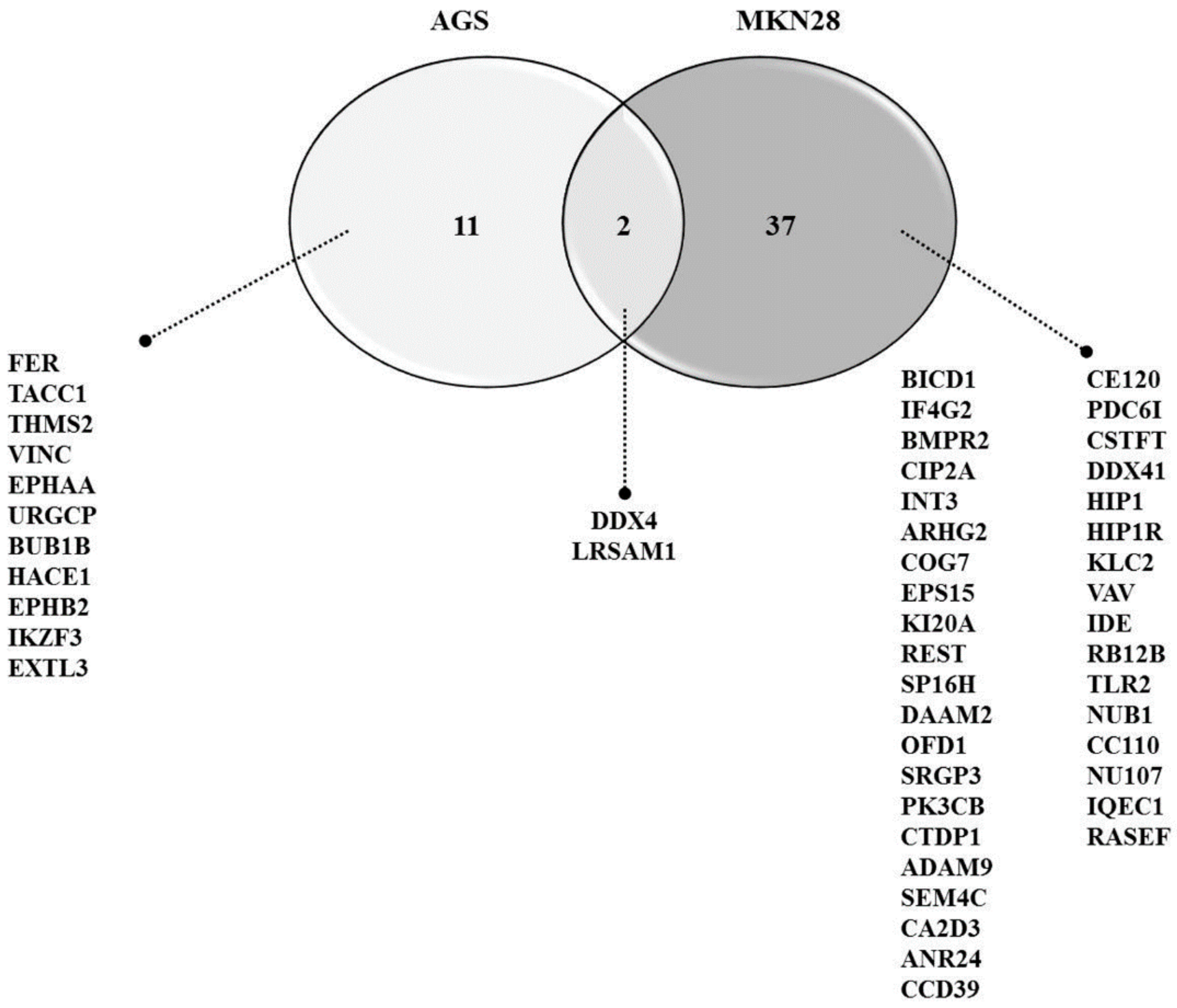
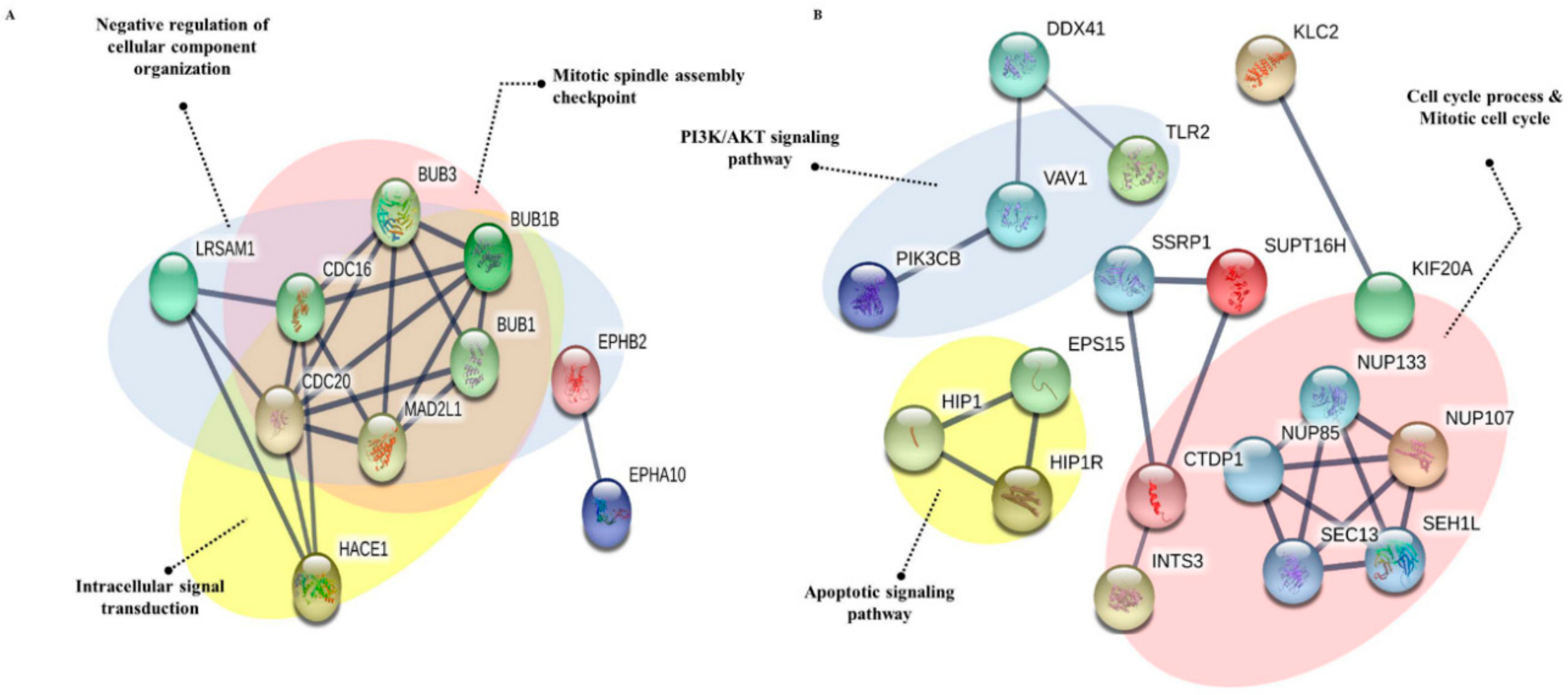
3.5. Protein-Protein Interaction
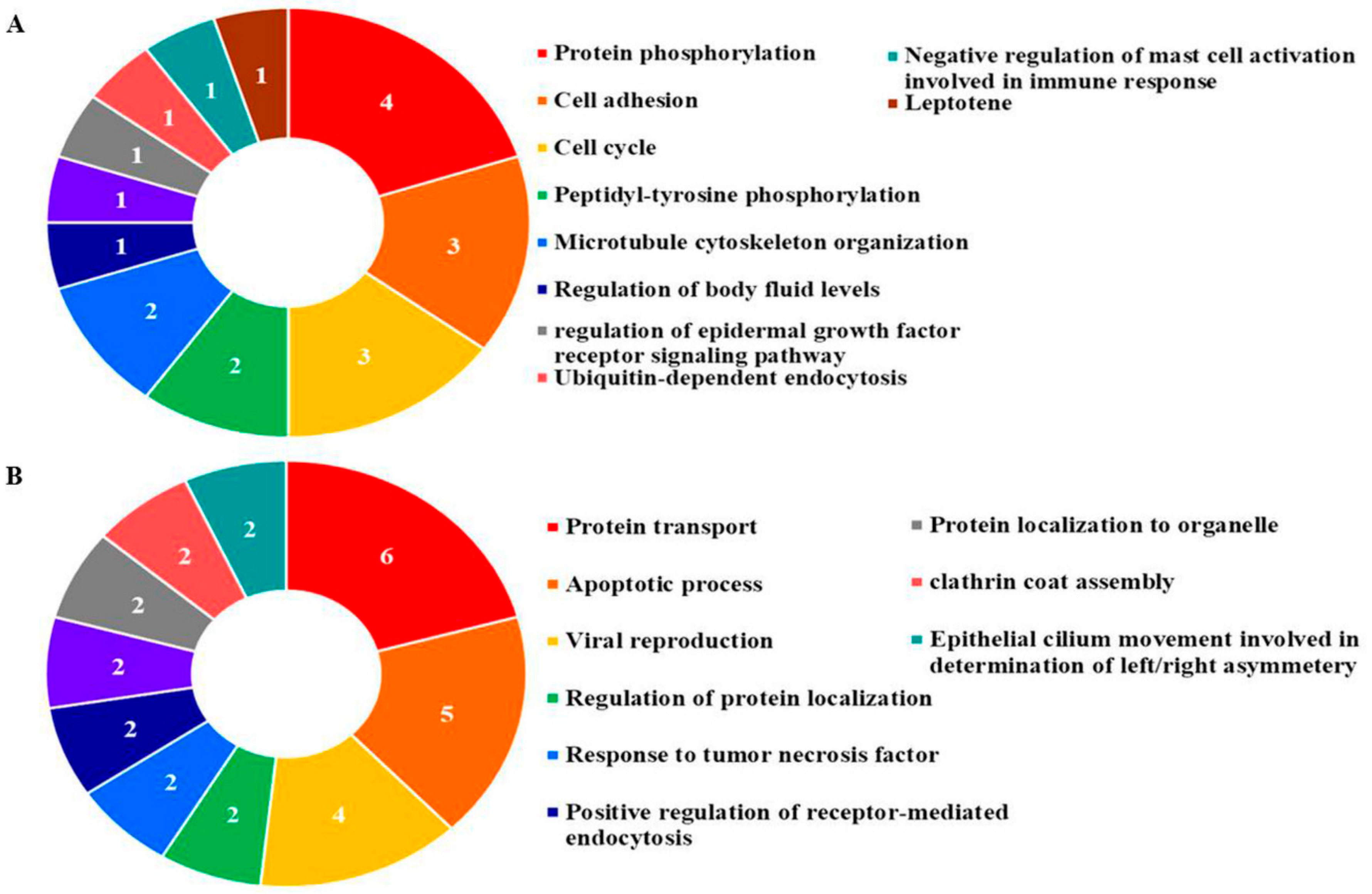
3.6. Gene Ontology Analysis
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Fock, K.M. Review article: The epidemiology and prevention of gastric cancer. Aliment. Pharmacol. Ther. 2014, 40, 250–260. [Google Scholar] [CrossRef] [PubMed]
- Shin, A.; Kim, J.; Park, S. Gastric cancer epidemiology in Korea. J. Gastric Cancer 2011, 11, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Rahman, R.; Asombang, A.W.; Ibdah, J.A. Characteristics of gastric cancer in Asia. World J. Gastroenterol. 2014, 20, 4483–4490. [Google Scholar] [CrossRef] [PubMed]
- Duijvesz, D.; Burnum-Johnson, K.E.; Gritsenko, M.A.; Hoogland, A.M.; Vredenbregt-van den Berg, M.S.; Willemsen, R.; Luider, T.; Pasa-Tolic, L.; Jenster, G. Proteomic profiling of exosomes leads to the identification of novel biomarkers for prostate cancer. PLoS ONE 2013, 8, e82589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, W.C. Contribution of oncoproteomics to cancer biomarker discovery. Mol. Cancer 2007, 6, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludwig, J.A.; Weinstein, J.N. Biomarkers in cancer staging, prognosis and treatment selection. Nat. Rev. Cancer 2005, 5, 845–856. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Cai, H.; Sasazuki, S.; Tsugane, S.; Zheng, W.; Cho, E.R.; Jee, S.H.; Michel, A.; Pawlita, M.; Xiang, Y.B.; et al. Fruit and vegetable consumption, Helicobacter pylori antibodies, and gastric cancer risk: A pooled analysis of prospective studies in China, Japan, and Korea. Int. J. Cancer 2017, 140, 591–599. [Google Scholar] [CrossRef] [PubMed]
- Nijveldt, R.J.; van Nood, E.; van Hoorn, D.E.; Boelens, P.G.; van Norren, K.; van Leeuwen, P.A. Flavonoids: A review of probable mechanisms of action and potential applications. Am. J. Clin. Nutr. 2001, 74, 418–425. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.S.; Fareed, S.; Ansari, S.; Rahman, M.A.; Ahmad, I.Z.; Saeed, M. Current approaches toward production of secondary plant metabolites. J. Pharm. Bioallied Sci. 2012, 4, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Nagappan, A.; Park, H.S.; Hong, G.E.; Yumnam, S.; Raha, S.; Saralamma, V.V.; Lee, W.S.; Kim, E.H.; Kim, G.S. Flavonoids isolated from Citrus platymamma induce mitochondrial-dependent apoptosis in AGS cells by modulation of the PI3K/AKT and MAPK pathways. Oncol. Rep. 2015, 34, 1517–1525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, Y.M.; Nam, J.H.; Kim, M.Y.; Choi, J.; Park, H.J. Pectolinarin and Pectolinarigenin of Cirsium setidens Prevent the Hepatic Injury in Rats Caused by D-Galactosamine via an Antioxidant Mechanism. Biol. Pharm. Bull. 2008, 31, 760–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, M. Pectolinarigenin—A Flavonoid Compound from Cirsium Japonicum with Potential Anti-proliferation Activity in MCF-7 Breast Cancer Cell. Trop. J. Pharm. Res. 2014, 13, 225–228. [Google Scholar] [CrossRef]
- Lee, H.J.; Venkatarame Gowda Saralamma, V.; Kim, S.M.; Ha, S.E.; Raha, S.; Lee, W.S.; Kim, E.H.; Lee, S.J.; Heo, J.D.; Kim, G.S. Pectolinarigenin Induced Cell Cycle Arrest, Autophagy, and Apoptosis in Gastric Cancer Cell via PI3K/AKT/mTOR Signaling Pathway. Nutrients 2018, 10, 1043. [Google Scholar] [CrossRef] [PubMed]
- Swain, M.; Ross, N.W. A silver stain protocol for proteins yielding high resolution and transparent background in sodium dodecyl sulfate-polyacrylamide gels. Electrophoresis 1995, 16, 948–951. [Google Scholar] [CrossRef] [PubMed]
- Shevchenko, A.; Wilm, M.; Vorm, O.; Mann, M. Mass spectrometric sequencing of proteins silver-stained polyacrylamide gels. Anal. Chem. 1996, 68, 850–858. [Google Scholar] [CrossRef] [PubMed]
- Rampogu, S.; Baek, A.; Zeb, A.; Lee, K.W. Exploration for novel inhibitors showing back-to-front approach against VEGFR-2 kinase domain (4AG8) employing molecular docking mechanism and molecular dynamics simulations. BMC Cancer 2018, 18, 264. [Google Scholar] [CrossRef] [PubMed]
- Carmona-Saez, P.; Chagoyen, M.; Tirado, F.; Carazo, J.M.; Pascual-Montano, A. GENECODIS: A web-based tool for finding significant concurrent annotations in gene lists. Genome Biol. 2007, 8, R3. [Google Scholar] [CrossRef] [PubMed]
- Tabas-Madrid, D.; Nogales-Cadenas, R.; Pascual-Montano, A. GeneCodis3: A non-redundant and modular enrichment analysis tool for functional genomics. Nucleic Acids Res. 2012, 40, W478–W483. [Google Scholar] [CrossRef] [PubMed]
- Bian, W.; Guo, Y.; Zhang, Y.; Li, H. The self-association and activity regulation of LRSAM1 E3 ligase. Biochem. Biophys. Res. Commun. 2017, 485, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Celli, J. LRSAM1, an E3 Ubiquitin ligase with a sense for bacteria. Cell. Host Microbe 2012, 12, 735–736. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Hu, J.; Li, G.; Qu, L.; He, Q.; Lou, Y.; Song, Q.; Ma, D.; Chen, Y. PHF23 (plant homeodomain finger protein 23) negatively regulates cell autophagy by promoting ubiquitination and degradation of E3 ligase LRSAM1. Autophagy 2014, 10, 2158–2170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schudrowitz, N.; Takagi, S.; Wessel, G.M.; Yajima, M. Germline factor DDX4 functions in blood-derived cancer cell phenotypes. Cancer Sci. 2017, 108, 1612–1619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Zhu, Y.; Liu, Z.J.; Ouyang, S. The emerging roles of the DDX41 protein in immunity and diseases. Protein Cell. 2017, 8, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Peters, D.; Radine, C.; Reese, A.; Budach, W.; Sohn, D.; Janicke, R.U. The DEAD-box RNA helicase DDX41 is a novel repressor of p21(WAF1/CIP1) mRNA translation. J. Biol. Chem. 2017, 292, 8331–8341. [Google Scholar] [CrossRef] [PubMed]
- Bartholomeusz, C.; Gonzalez-Angulo, A.M. Targeting the PI3K signaling pathway in cancer therapy. Expert Opin. Ther. Targets 2012, 16, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Crowder, R.J.; Phommaly, C.; Tao, Y.; Hoog, J.; Luo, J.; Perou, C.M.; Parker, J.S.; Miller, M.A.; Huntsman, D.G.; Lin, L.; et al. PIK3CA and PIK3CB inhibition produce synthetic lethality when combined with estrogen deprivation in estrogen receptor-positive breast cancer. Cancer Res. 2009, 69, 3955–3962. [Google Scholar] [CrossRef] [PubMed]
- Wee, S.; Wiederschain, D.; Maira, S.M.; Loo, A.; Miller, C.; deBeaumont, R.; Stegmeier, F.; Yao, Y.M.; Lengauer, C. PTEN-deficient cancers depend on PIK3CB. Proc. Natl. Acad. Sci. USA 2008, 105, 13057–13062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Z.; Liu, D.; Su, Z. CIP2A mediates prostate cancer progression via the c-MYC signaling pathway. Tumour Biol. 2015, 36, 3583–3589. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Wen, Z.S.; Liu, Z.; Hu, Z.; Ma, J.; Chen, X.Q.; Liu, Y.Q.; Pu, J.X.; Xiao, W.L.; Sun, H.D.; et al. Overexpression and small molecule-triggered downregulation of CIP2A in lung cancer. PLoS ONE 2011, 6, e20159. [Google Scholar] [CrossRef] [PubMed]
- Niemela, M.; Kauko, O.; Sihto, H.; Mpindi, J.P.; Nicorici, D.; Pernila, P.; Kallioniemi, O.P.; Joensuu, H.; Hautaniemi, S.; Westermarck, J. CIP2A signature reveals the MYC dependency of CIP2A-regulated phenotypes and its clinical association with breast cancer subtypes. Oncogene 2012, 31, 4266–4278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.S.; Kim, E.J.; Oh, J.S.; Park, I.C.; Hwang, S.G. CIP2A modulates cell-cycle progression in human cancer cells by regulating the stability and activity of Plk1. Cancer Res. 2013, 73, 6667–6678. [Google Scholar] [CrossRef] [PubMed]
- Davenport, J.W.; Fernandes, E.R.; Harris, L.D.; Neale, G.A.; Goorha, R. The mouse mitotic checkpoint gene bub1b, a novel bub1 family member, is expressed in a cell cycle-dependent manner. Genomics 1999, 55, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Liu, Y.; Shang, L.; Yu, J.; Qu, Q. The FOXM1/BUB1B signaling pathway is essential for the tumorigenicity and radioresistance of glioblastoma. Oncol. Rep. 2017, 38, 3367–3375. [Google Scholar] [CrossRef] [PubMed]
- Myrie, K.A.; Percy, M.J.; Azim, J.N.; Neeley, C.K.; Petty, E.M. Mutation and expression analysis of human BUB1 and BUB1B in aneuploid breast cancer cell lines. Cancer Lett. 2000, 152, 193–199. [Google Scholar] [CrossRef]
- Song, J.; Xie, H.; Lian, Z.; Yang, G.; Du, R.; Du, Y.; Zou, X.; Jin, H.; Gao, J.; Liu, J.; et al. Enhanced cell survival of gastric cancer cells by a novel gene URG4. Neoplasia 2006, 8, 995–1002. [Google Scholar] [CrossRef] [PubMed]
- Yan, G.R.; Zou, F.Y.; Dang, B.L.; Zhang, Y.; Yu, G.; Liu, X.; He, Q.Y. Genistein-induced mitotic arrest of gastric cancer cells by downregulating KIF20A, a proteomics study. Proteomics 2012, 12, 2391–2399. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Gong, H.; Huang, K. Oncogenic role of kinesin proteins and targeting kinesin therapy. Cancer Sci. 2013, 104, 651–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rath, O.; Kozielski, F. Kinesins and cancer. Nat. Rev. Cancer 2012, 12, 527–539. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, H.N.; Gibbs, J.; Jordan, T.; Blackshear, M. Depletion of a single nucleoporin, Nup107, induces apoptosis in eukaryotic cells. Mol. Cell. Biochem. 2010, 343, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Yoon, S.; Won, M.; Sim, S.H.; Ko, J.J.; Han, S.; Lee, K.A.; Lee, K.; Bae, J. HIP1R interacts with a member of Bcl-2 family, BCL2L10, and induces BAK-dependent cell death. Cell. Physiol. Biochem. 2009, 23, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Gervais, F.G.; Singaraja, R.; Xanthoudakis, S.; Gutekunst, C.A.; Leavitt, B.R.; Metzler, M.; Hackam, A.S.; Tam, J.; Vaillancourt, J.P.; Houtzager, V.; et al. Recruitment and activation of caspase-8 by the Huntingtin-interacting protein Hip-1 and a novel partner Hippi. Nat. Cell Biol. 2002, 4, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Mai, A.; Muharram, G.; Barrow-McGee, R.; Baghirov, H.; Rantala, J.; Kermorgant, S.; Ivaska, J. Distinct c-Met activation mechanisms induce cell rounding or invasion through pathways involving integrins, RhoA and HIP1. J. Cell Sci. 2014, 127 Pt 9, 1938–1952. [Google Scholar] [CrossRef]
- Fernandez-Zapico, M.E.; Gonzalez-Paz, N.C.; Weiss, E.; Savoy, D.N.; Molina, J.R.; Fonseca, R.; Smyrk, T.C.; Chari, S.T.; Urrutia, R.; Billadeau, D.D. Ectopic expression of VAV1 reveals an unexpected role in pancreatic cancer tumorigenesis. Cancer Cell 2005, 7, 39–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazer, G.; Idelchuk, Y.; Schapira, V.; Pikarsky, E.; Katzav, S. The haematopoietic specific signal transducer Vav1 is aberrantly expressed in lung cancer and plays a role in tumourigenesis. J. Pathol. 2009, 219, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Husi, H.; Skipworth, R.J.; Cronshaw, A.; Stephens, N.A.; Wackerhage, H.; Greig, C.; Fearon, K.C.; Ross, J.A. Programmed cell death 6 interacting protein (PDCD6IP) and Rabenosyn-5 (ZFYVE20) are potential urinary biomarkers for upper gastrointestinal cancer. Proteomics Clin. Appl. 2015, 9, 586–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kent, O.A.; Sandi, M.J.; Burston, H.E.; Brown, K.R.; Rottapel, R. An oncogenic KRAS transcription program activates the RHOGEF ARHGEF2 to mediate transformed phenotypes in pancreatic cancer. Oncotarget 2017, 8, 4484–4500. [Google Scholar] [CrossRef] [PubMed]
- Badura, M.; Braunstein, S.; Zavadil, J.; Schneider, R.J. DNA damage and eIF4G1 in breast cancer cells reprogram translation for survival and DNA repair mRNAs. Proc. Natl. Acad. Sci. USA 2012, 109, 18767–18772. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Spot No. | Accession No. | Protein Name | MOWSE Score | Sequence Coverage (%)/Peptides Matched | Theoretical Mr (Da) | Theoretical pI Value | Fold Change | Up/Down | p-Value (Anova) |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 364 | P16591 | Tyrosine-protein kinase Fer | 1.04 × 106 | 22.1/21 | 94,639 | 6.7 | 2.1 | ↑ | 0.001 |
| 2 | 478 | O75410 | Transforming acidic coiled-coil- containing protein 1 | 1,640,000 | 17.1/14 | 87,795 | 4.8 | 1.8 | ↑ | 0.003 |
| 3 | 728 | Q6UWE0 | E3 ubiquitin-protein ligase LRSAM1 | 1,510,000 | 23/18 | 83,595 | 5.7 | 1.9 | ↑ | 0.003 |
| 4 | 723 | Q5TEJ8 | Protein THEMIS2 | 7.09 × 106 | 17.9/8 | 72,049 | 5.8 | 2.3 | ↓ | 0.008 |
| 5 | 725 | P18206 | Vinculin | 1.01 × 106 | 16.3/14 | 123,800 | 5.5 | 1.7 | ↑ | 0.009 |
| 6 | 1065 | Q5JZY3 | Ephrin type-A receptor 10 | 1.19 × 106 | 16/11 | 109,717 | 6.5 | 2.7 | ↓ | 0.01 |
| 7 | 448 | Q8TCY9 | Up-regulator of cell proliferation | 1.77 × 106 | 13.7/16 | 104,988 | 6 | 1.6 | ↑ | 0.01 |
| 8 | 1147 | O60566 | Mitotic checkpoint serine/threonine-protein kinase BUB1 beta | 2.57 × 106 | 12.2/12 | 119,546 | 5.2 | 1.9 | ↓ | 0.014 |
| 9 | 202 | Q8IYU2 | E3 ubiquitin-protein ligase HACE1 | 1.58 × 106 | 12.9/10 | 102,343 | 5.6 | 3.1 | ↓ | 0.027 |
| 10 | 339 | P29323 | Ephrin type-B receptor 2 | 1.45 × 106 | 15/12 | 117,494 | 6.1 | 2.4 | ↓ | 0.033 |
| 11 | 381 | Q9UKT9 | Zinc finger protein Aiolos | 2.12 × 106 | 24.4/8 | 58,024 | 6.1 | 1.5 | ↑ | 0.035 |
| 12 | 418 | Q9NQI0 | Probable ATP-dependent RNA helicase DDX4 | 1.23 × 106 | 15.9/9 | 79,308 | 5.6 | 2.2 | ↓ | 0.036 |
| 13 | 1001 | O43909 | Exostosin-like 3 | 8.19 × 107 | 17/16 | 104,750 | 6.1 | 2.5 | ↑ | 0.047 |
| No. | Spot No. | Accession No. | Protein Name | MOWSE Score | Sequence Coverage (%)/Peptides Matched | Theoretical Mr (Da) | Theoretical pI Value | Fold Change | Up/Down | p-Value (Anova) |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 774 | Q96G01 | Protein bicaudal D homolog 1 | 3.68 × 106 | 15.4/16 | 110,751 | 5.6 | 2.1 | ↑ | 0.002 |
| 2 | 2101 | P78344 | Eukaryotic translation initiation factor 4 gamma 2 | 1.50 × 106 | 12.2/13 | 102,363 | 6.7 | 2.9 | ↓ | 0.002 |
| 3 | 1482 | Q13873 | Bone morphogenetic protein receptor type-2 | 1.77 × 106 | 12.8/11 | 115,202 | 5.8 | 2.5 | ↓ | 0.002 |
| 4 | 1027 | Q8TCG1 | Protein CIP2A | 2.92 × 106 | 15.5/13 | 102,186 | 5.9 | 1.5 | ↓ | 0.002 |
| 5 | 765 | Q68E01 | Integrator complex subunit 3 | 7.80 × 106 | 16/13 | 118,071 | 5.5 | 7.3 | ↑ | 0.003 |
| 6 | 1238 | Q92974 | Rho guanine nucleotide exchange factor 2 | 1.39 × 106 | 15.1/13 | 111,544 | 6.9 | 2.1 | ↓ | 0.003 |
| 7 | 1828 | P83436 | Conserved oligomeric Golgi complex subunit 7 | 1.84 × 106 | 15.7/8 | 86,345 | 5.3 | 3.9 | ↓ | 0.004 |
| 8 | 1881 | P42566 | Epidermal growth factor receptor substrate 15 | 2.84 × 1010 | 23.4/19 | 98,657 | 4.5 | 2.7 | ↓ | 0.004 |
| 9 | 2049 | O95235 | Kinesin-like protein KIF20A | 7.75 × 106 | 16.2/12 | 100,279 | 6.5 | 2.3 | ↓ | 0.005 |
| 10 | 1250 | Q13127 | RE1-silencing transcription factor | 1.88 × 106 | 16.8/19 | 121,873 | 6.3 | 2.3 | ↑ | 0.005 |
| 11 | 866 | Q9Y5B9 | FACT complex subunit SPT16 | 6.78 × 1011 | 30.6/37 | 119,915 | 5.5 | 1.7 | ↑ | 0.007 |
| 12 | 1281 | Q86T65 | Disheveled-associated activator of morphogenesis 2 | 1.35 × 107 | 16.8/21 | 123,500 | 6.4 | 1.5 | ↓ | 0.007 |
| 13 | 1477 | O75665 | Oral-facial-digital syndrome 1 protein | 1.49 × 106 | 15.7/21 | 116,672 | 5.8 | 2.1 | ↓ | 0.008 |
| 14 | 650 | O43295 | SLIT-ROBO Rho GTPase-activating protein 3 | 2.74 × 109 | 22.2/26 | 124,505 | 6.2 | 1.8 | ↑ | 0.008 |
| 15 | 1558 | P42338 | Phosphatidylinositol 4,5-bisphosphate 3-kinase catalytic subunit beta isoform | 3.92 × 106 | 15.7/15 | 122,763 | 6.7 | 2.6 | ↓ | 0.008 |
| 16 | 1319 | Q9Y5B0 | RNA polymerase II subunit A C-terminal domain phosphatase | 4.51 × 106 | 17/13 | 104,400 | 5.2 | 2.4 | ↓ | 0.009 |
| 17 | 1431 | Q13443 | Disintegrin and metalloproteinase domain-containing protein 9 | 3.67 × 106 | 17.6/12 | 90,557 | 7.7 | 2.2 | ↓ | 0.009 |
| 18 | 1648 | Q6UWE0 | E3 ubiquitin-protein ligase LRSAM1 | 1.48 × 106 | 16.3/14 | 83,595 | 5.7 | 1.5 | ↓ | 0.009 |
| 19 | 663 | Q9C0C4 | Semaphorin-4C | 368,935 | 13.2/8 | 92,624 | 6.9 | 2.7 | ↑ | 0.009 |
| 20 | 605 | Q8IZS8 | Voltage-dependent calcium channel subunit alpha-2/delta-3 | 1.19 × 106 | 16.5/15 | 123,012 | 5.5 | 2.6 | ↓ | 0.01 |
| 21 | 500 | Q8TF21 | Ankyrin repeat domain-containing protein 24 | 1.88 × 106 | 14/15 | 124,188 | 5 | 2.5 | ↑ | 0.01 |
| 22 | 1706 | Q9UFE4 | Coiled-coil domain-containing protein 39 | 5.24 × 106 | 19.4/19 | 109,901 | 6.1 | 2.2 | ↓ | 0.011 |
| 23 | 2098 | Q8N960 | Centrosomal protein of 120 kDa | 2.94 × 106 | 19/19 | 112,641 | 5.9 | 2.3 | ↓ | 0.011 |
| 24 | 907 | Q8WUM4 | Programmed cell death 6-interacting protein | 1.07 × 106 | 15/12 | 96,024 | 6.1 | 1.8 | ↑ | 0.012 |
| 25 | 336 | Q9H0L4 | Cleavage stimulation factor subunit 2 tau variant | 7.95 × 106 | 21.1/18 | 64,437 | 6.8 | 1.6 | ↓ | 0.013 |
| 26 | 841 | Q9UJV9 | Probable ATP-dependent RNA helicase DDX41 | 1.14 × 106 | 16.7/15 | 69,838 | 6.4 | 1.9 | ↑ | 0.013 |
| 27 | 858 | O00291 | Huntingtin-interacting protein 1 | 1.99 × 106 | 20.3/25 | 116,222 | 5.2 | 3 | ↑ | 0.015 |
| 28 | 2170 | Q9H0B6 | Kinesin light chain 2 | 6.75 × 106 | 22.3/13 | 68,935 | 6.7 | 2.3 | ↓ | 0.021 |
| 29 | 634 | P15498 | Proto-oncogene vav | 3.27 × 106 | 16/13 | 98,315 | 6.2 | 2.2 | ↑ | 0.023 |
| 30 | 1790 | P14735 | Insulin-degrading enzyme | 8.77 × 1011 | 22.1/28 | 117,970 | 6.2 | 1.6 | ↓ | 0.023 |
| 31 | 751 | Q8IXT5 | RNA-binding protein 12B | 1.05 × 106 | 16.4/15 | 118,104 | 6.3 | 1.7 | ↑ | 0.025 |
| 32 | 1644 | O60603 | Toll-like receptor 2 | 1.03 × 106 | 16.8/13 | 89,838 | 6.2 | 1.8 | ↓ | 0.027 |
| 33 | 2044 | Q9NQI0 | Probable ATP-dependent RNA helicase DDX4 | 1.06 × 107 | 18.2/13 | 79,308 | 5.6 | 2.6 | ↓ | 0.032 |
| 34 | 831 | Q9Y5A7 | NEDD8 ultimate buster 1 | 5.37 × 106 | 20.7/15 | 70,539 | 5.7 | 2.4 | ↑ | 0.035 |
| 35 | 1377 | Q8TBZ0 | Coiled-coil domain-containing protein 110 | 1.97 × 106 | 19.9/21 | 96,726 | 5.9 | 1.6 | ↓ | 0.035 |
| 36 | 1562 | P57740 | Nuclear pore complex protein Nup107 | 3.63 × 106 | 18.9/15 | 106,375 | 5.3 | 1.5 | ↓ | 0.036 |
| 37 | 1686 | O75146 | Huntingtin-interacting protein 1-related protein | 1.39 × 106 | 11.1/13 | 119,389 | 6.2 | 2.5 | ↓ | 0.038 |
| 38 | 1340 | Q6DN90 | IQ motif and SEC7 domain-containing protein 1 | 3.48 × 106 | 15.7/16 | 108,315 | 6.5 | 2.3 | ↓ | 0.042 |
| 39 | 664 | Q8IZ41 | Ras and EF-hand domain-containing protein | 1.35 × 106 | 18/12 | 82,880 | 5 | 2.3 | ↑ | 0.043 |
| Cell Line | GO Accession | Biological Process | Number of Genes | Gene Symbol | Gene Name | p-Value |
|---|---|---|---|---|---|---|
| AGS Cells | GO: 0042058 | Regulation of epidermal growth factor receptor signaling pathway | 1 | FER | Tyrosine-protein kinase Fer | 1.14 × 10−3 |
| GO: 0007049 | Cell Cycle | 3 | TACC1 | Transforming acidic coiled-coil-containing protein 1 | 5.31 × 10−4 | |
| BUB1B | Mitotic checkpoint serine/threonine-protein kinase BUB1 beta | |||||
| URGCP | Up-regulator of cell proliferation | |||||
| GO: 0045806 | Negative regulation of endocytosis | 1 | LRSAM1 | E3-ubiquitin-protein ligase | 3.42 × 10−3 | |
| MKN28 Cells | GO: 0006915 | Apoptotic process | 5 | HIP1 | Huntingtin-interacting protein 1 | 4.85 × 10−4 |
| VAV | Proto-oncogene vav | |||||
| PDC6I | Programmed cell death 6-interacting protein | |||||
| DDX41 | Probabale ATP-dependnet RNA helicase DDX41 | |||||
| ARHG2 | Rho guanine nucleotide exchange factor 2 | |||||
| GO: 0000087 | M phase of mitotic cycle | 2 | KI20A | Kinesin-like protein KIFA20A | 5.03 × 10−3 | |
| NU107 | Nuclear pore complex protein Nup107 | |||||
| GO: 0008219 | Cell death | 2 | HIP1 | Huntingtin-interacting protein 1 | 1.30 × 10−3 | |
| IF4G2 | Eukaryotic translation initiation factor 4 gamma 2 | |||||
| GO: 0048260 | Positive regulation of receptor-mediated endocytosis | 2 | HIP1 | Huntingtin-interacting protein 1 | 1.08 × 10−4 | |
| BICD1 | Protein bicaudal D homolog 1 | |||||
| GO: 0034241 | Positive regulation of macrophage fusion | 1 | ADAM9 | Disintegrin and metalloproteinase domain-containing protein 9 | 1.11 × 10−3 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.J.; Venkatarame Gowda Saralamma, V.; Kim, S.M.; Ha, S.E.; Vetrivel, P.; Kim, E.H.; Lee, S.J.; Heo, J.D.; Rampogu, S.; Lee, K.W.; et al. Comparative Proteomic Profiling of Tumor-Associated Proteins in Human Gastric Cancer Cells Treated with Pectolinarigenin. Nutrients 2018, 10, 1596. https://0-doi-org.brum.beds.ac.uk/10.3390/nu10111596
Lee HJ, Venkatarame Gowda Saralamma V, Kim SM, Ha SE, Vetrivel P, Kim EH, Lee SJ, Heo JD, Rampogu S, Lee KW, et al. Comparative Proteomic Profiling of Tumor-Associated Proteins in Human Gastric Cancer Cells Treated with Pectolinarigenin. Nutrients. 2018; 10(11):1596. https://0-doi-org.brum.beds.ac.uk/10.3390/nu10111596
Chicago/Turabian StyleLee, Ho Jeong, Venu Venkatarame Gowda Saralamma, Seong Min Kim, Sang Eun Ha, Preethi Vetrivel, Eun Hee Kim, Snag Joon Lee, Jeong Doo Heo, Shailima Rampogu, Keun Woo Lee, and et al. 2018. "Comparative Proteomic Profiling of Tumor-Associated Proteins in Human Gastric Cancer Cells Treated with Pectolinarigenin" Nutrients 10, no. 11: 1596. https://0-doi-org.brum.beds.ac.uk/10.3390/nu10111596