Impact of Fecal Microbiota Transplantation on Obesity and Metabolic Syndrome—A Systematic Review

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Eligibility Criteria
2.2. Information Sources
2.3. Study Selection and Data Extraction
2.4. Data Synthesis
2.5. Assessment of Risk of Bias
3. Results
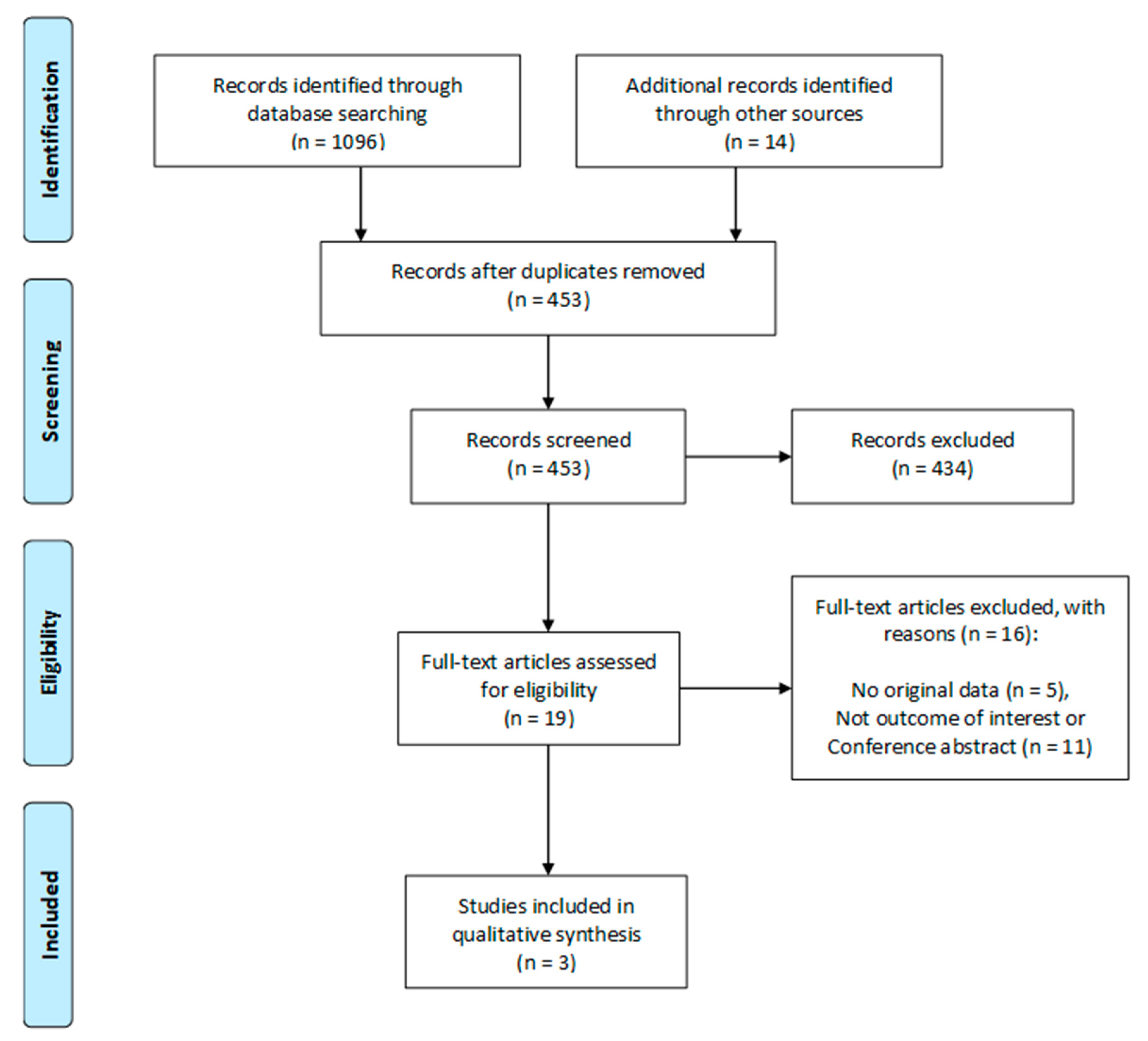
3.1. Search Results
3.2. Study Characteristics and Baseline Demographics
3.3. Fecal Microbiota Transplantation (FMT) Donor and Delivery Method
3.4. Assessment of Risk of Bias
3.5. FMT and Metabolic Outcomes
3.6. FMT Influences Microbiome Composition and Derived Metabolites
4. Discussion
4.1. Effect of FMT on the Metabolic Parameters
4.2. The Ecological Challenges of the FMT on Obesity and its Related Metabolic Disorders
4.3. FMT in Clinical Practice
4.4. Limitations and Strengths
4.5. Future Direction
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- WHO. Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight: (accessed on 16 February 2018).
- O’Neill, S.; O’Driscoll, L. Metabolic syndrome: A closer look at the growing epidemic and its associated pathologies. Obes. Rev. 2015, 16, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Saklayen, M.G. The Global Epidemic of the Metabolic Syndrome. Curr. Hypertens. Rep. 2018, 20, 12. [Google Scholar] [CrossRef] [PubMed]
- Bomberg, E.; Birch, L.; Endenburg, N.; German, A.J.; Neilson, J.; Seligman, H.; Takashima, G.; Day, M.J. The Financial Costs, Behaviour and Psychology of Obesity: A One Health Analysis. J. Comp. Pathol. 2017, 156, 310–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, A.N.; Lachman, M.E. Behavior Change with Fitness Technology in Sedentary Adults: A Review of the Evidence for Increasing Physical Activity. Front Public Health 2016, 4, 289. [Google Scholar] [CrossRef] [PubMed]
- Burgess, E.; Hassmen, P.; Pumpa, K.L. Determinants of adherence to lifestyle intervention in adults with obesity: A systematic review. Clin. Obes. 2017, 7, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Yancy, W.S., Jr.; Olsen, M.K.; Guyton, J.R.; Bakst, R.P.; Westman, E.C. A low-carbohydrate, ketogenic diet versus a low-fat diet to treat obesity and hyperlipidemia: A randomized, controlled trial. Ann. Int. Med. 2004, 140, 769–777. [Google Scholar] [CrossRef] [PubMed]
- Mehta, A.; Marso, S.P.; Neeland, I.J. Liraglutide for weight management: A critical review of the evidence. Obes. Sci. Pract. 2017, 3, 3–14. [Google Scholar] [CrossRef]
- le Roux, C.W.; Astrup, A.; Fujioka, K.; Greenway, F.; Lau, D.C.W.; Van Gaal, L.; Ortiz, R.V.; Wilding, J.P.H.; Skjoth, T.V.; Manning, L.S.; et al. 3 years of liraglutide versus placebo for type 2 diabetes risk reduction and weight management in individuals with prediabetes: A randomised, double-blind trial. Lancet 2017, 389, 1399–1409. [Google Scholar] [CrossRef]
- Tek, C. Naltrexone HCI/bupropion HCI for chronic weight management in obese adults: Patient selection and perspectives. Patient Prefer Adherence 2016, 10, 751–759. [Google Scholar] [CrossRef] [PubMed]
- Pinkney, J.; Kerrigan, D. Current status of bariatric surgery in the treatment of type 2 diabetes. Obes. Rev. 2004, 5, 69–78. [Google Scholar] [CrossRef]
- Peterli, R.; Wolnerhanssen, B.K.; Peters, T.; Vetter, D.; Kroll, D.; Borbely, Y.; Schultes, B.; Beglinger, C.; Drewe, J.; Schiesser, M.; et al. Effect of Laparoscopic Sleeve Gastrectomy vs. Laparoscopic Roux-en-Y Gastric Bypass on Weight Loss in Patients With Morbid Obesity: The SM-BOSS Randomized Clinical Trial. JAMA 2018, 319, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Schauer, P.R.; Bhatt, D.L.; Kirwan, J.P.; Wolski, K.; Aminian, A.; Brethauer, S.A.; Navaneethan, S.D.; Singh, R.P.; Pothier, C.E.; Nissen, S.E.; et al. Bariatric Surgery versus Intensive Medical Therapy for Diabetes—5-Year Outcomes. N. Engl. J. Med. 2017, 376, 641–651. [Google Scholar] [CrossRef] [PubMed]
- Sender, R.; Fuchs, S.; Milo, R. Revised Estimates for the Number of Human and Bacteria Cells in the Body. PLoS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turnbaugh, P.J.; Ley, R.E.; Hamady, M.; Fraser-Liggett, C.M.; Knight, R.; Gordon, J.I. The human microbiome project. Nature 2007, 449, 804–810. [Google Scholar] [CrossRef] [PubMed]
- Baktash, A.; Terveer, E.M.; Zwittink, R.D.; Hornung, B.V.H.; Corver, J.; Kuijper, E.J.; Smits, W.K. Mechanistic Insights in the Success of Fecal Microbiota Transplants for the Treatment of Clostridium difficile Infections. Front. Microbiol. 2018, 9, 1242. [Google Scholar] [CrossRef] [PubMed]
- Brahe, L.K.; Astrup, A.; Larsen, L.H. Can We Prevent Obesity-Related Metabolic Diseases by Dietary Modulation of the Gut Microbiota? Adv. Nutr. 2016, 7, 90–101. [Google Scholar] [CrossRef]
- He, M.; Shi, B. Gut microbiota as a potential target of metabolic syndrome: The role of probiotics and prebiotics. Cell Biosci. 2017, 7, 54. [Google Scholar] [CrossRef]
- Wortelboer, K.; Nieuwdorp, M.; Herrema, H. Fecal microbiota transplantation beyond Clostridioides difficile infections. EBioMedicine 2019. [Google Scholar] [CrossRef]
- Tamboli, C.P.; Neut, C.; Desreumaux, P.; Colombel, J.F. Dysbiosis in inflammatory bowel disease. Gut 2004, 53, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Walker, A.W.; Lawley, T.D. Therapeutic modulation of intestinal dysbiosis. Pharmacol. Res. 2013, 69, 75–86. [Google Scholar] [CrossRef]
- Machiels, K.; Joossens, M.; Sabino, J.; De Preter, V.; Arijs, I.; Eeckhaut, V.; Ballet, V.; Claes, K.; Van Immerseel, F.; Verbeke, K.; et al. A decrease of the butyrate-producing species Roseburia hominis and Faecalibacterium prausnitzii defines dysbiosis in patients with ulcerative colitis. Gut 2014, 63, 1275–1283. [Google Scholar] [CrossRef] [PubMed]
- Scheithauer, T.P.; Dallinga-Thie, G.M.; de Vos, W.M.; Nieuwdorp, M.; van Raalte, D.H. Causality of small and large intestinal microbiota in weight regulation and insulin resistance. Mol. Metab. 2016, 5, 759–770. [Google Scholar] [CrossRef] [PubMed]
- Duranti, S.; Ferrario, C.; van Sinderen, D.; Ventura, M.; Turroni, F. Obesity and microbiota: An example of an intricate relationship. Genes Nutr. 2017, 12, 18. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E.; Backhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef] [PubMed]
- Ridaura, V.K.; Faith, J.J.; Rey, F.E.; Cheng, J.; Duncan, A.E.; Kau, A.L.; Griffin, N.W.; Lombard, V.; Henrissat, B.; Bain, J.R.; et al. Gut microbiota from twins discordant for obesity modulate metabolism in mice. Science 2013, 341, 1241214. [Google Scholar] [CrossRef] [PubMed]
- Vindigni, S.M.; Surawicz, C.M. Fecal Microbiota Transplantation. Gastroenterol. Clin. North Am. 2017, 46, 171–185. [Google Scholar] [CrossRef]
- Wilson, B.C.; Vatanen, T.; Cutfield, W.S.; O’Sullivan, J.M. The Super-Donor Phenomenon in Fecal Microbiota Transplantation. Front. Cell Infect. Microbiol. 2019, 9, 2. [Google Scholar] [CrossRef] [Green Version]
- Kriss, M.; Hazleton, K.Z.; Nusbacher, N.M.; Martin, C.G.; Lozupone, C.A. Low diversity gut microbiota dysbiosis: Drivers, functional implications and recovery. Curr. Opin. Microbiol. 2018, 44, 34–40. [Google Scholar] [CrossRef]
- Li, Y.T.; Cai, H.F.; Wang, Z.H.; Xu, J.; Fang, J.Y. Systematic review with meta-analysis: Long-term outcomes of faecal microbiota transplantation for Clostridium difficile infection. Aliment. Pharmacol. Ther. 2016, 43, 445–457. [Google Scholar] [CrossRef]
- Quraishi, M.N.; Widlak, M.; Bhala, N.; Moore, D.; Price, M.; Sharma, N.; Iqbal, T.H. Systematic review with meta-analysis: The efficacy of faecal microbiota transplantation for the treatment of recurrent and refractory Clostridium difficile infection. Aliment. Pharmacol. Ther. 2017, 46, 479–493. [Google Scholar] [CrossRef] [PubMed]
- Hui, W.; Li, T.; Liu, W.; Zhou, C.; Gao, F. Fecal microbiota transplantation for treatment of recurrent C. difficile infection: An updated randomized controlled trial meta-analysis. PLoS ONE 2019, 14, e0210016. [Google Scholar] [CrossRef] [PubMed]
- Khanna, S.; Vazquez-Baeza, Y.; Gonzalez, A.; Weiss, S.; Schmidt, B.; Muniz-Pedrogo, D.A.; Rainey, J.F., III; Kammer, P.; Nelson, H.; Sadowsky, M.; et al. Changes in microbial ecology after fecal microbiota transplantation for recurrent C. difficile infection affected by underlying inflammatory bowel disease. Microbiome 2017, 5, 55. [Google Scholar] [CrossRef] [PubMed]
- Theriot, C.M.; Young, V.B. Interactions between the Gastrointestinal Microbiome and Clostridium difficile. Annu. Rev. Microbiol. 2015, 69, 445–461. [Google Scholar] [CrossRef] [PubMed]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Group, P. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef]
- Borody, T.J.; Khoruts, A. Fecal microbiota transplantation and emerging applications. Nat. Rev. Gastroenterol. Hepatol. 2011, 9, 88–96. [Google Scholar] [CrossRef]
- Walter, J.; Maldonado-Gomez, M.X.; Martinez, I. To engraft or not to engraft: An ecological framework for gut microbiome modulation with live microbes. Curr. Opin. Biotechnol. 2018, 49, 129–139. [Google Scholar] [CrossRef]
- Higgins, J.P.T.; Green, S. Cochrane Handbook for Systematic Reviews of Interventions Version 5.1.0. Updated March 2011. The Cochrane Collaboration. Available online: www.handbook.cochrane.org (accessed on 26 June 2019).
- Vrieze, A.; Van Nood, E.; Holleman, F.; Salojarvi, J.; Kootte, R.S.; Bartelsman, J.F.; Dallinga-Thie, G.M.; Ackermans, M.T.; Serlie, M.J.; Oozeer, R.; et al. Transfer of intestinal microbiota from lean donors increases insulin sensitivity in individuals with metabolic syndrome. Gastroenterology 2012, 143, 913–916. [Google Scholar] [CrossRef]
- Kootte, R.S.; Levin, E.; Salojarvi, J.; Smits, L.P.; Hartstra, A.V.; Udayappan, S.D.; Hermes, G.; Bouter, K.E.; Koopen, A.M.; Holst, J.J.; et al. Improvement of Insulin Sensitivity after Lean Donor Feces in Metabolic Syndrome Is Driven by Baseline Intestinal Microbiota Composition. Cell Metab. 2017, 26, 611–619. [Google Scholar] [CrossRef]
- Smits, L.P.; Kootte, R.S.; Levin, E.; Prodan, A.; Fuentes, S.; Zoetendal, E.G.; Wang, Z.; Levison, B.S.; Cleophas, M.C.P.; Kemper, E.M.; et al. Effect of Vegan Fecal Microbiota Transplantation on Carnitine- and Choline-Derived Trimethylamine-N-Oxide Production and Vascular Inflammation in Patients with Metabolic Syndrome. J. Am. Heart Assoc. 2018, 7. [Google Scholar] [CrossRef]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut microbiota functions: Metabolism of nutrients and other food components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Baxter, N.T.; Schmidt, A.W.; Venkataraman, A.; Kim, K.S.; Waldron, C.; Schmidt, T.M. Dynamics of Human Gut Microbiota and Short-Chain Fatty Acids in Response to Dietary Interventions with Three Fermentable Fibers. MBio 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Tolhurst, G.; Heffron, H.; Lam, Y.S.; Parker, H.E.; Habib, A.M.; Diakogiannaki, E.; Cameron, J.; Grosse, J.; Reimann, F.; Gribble, F.M. Short-chain fatty acids stimulate glucagon-like peptide-1 secretion via the G-protein-coupled receptor FFAR2. Diabetes 2012, 61, 364–371. [Google Scholar] [CrossRef] [PubMed]
- McNabney, S.M.; Henagan, T.M. Short Chain Fatty Acids in the Colon and Peripheral Tissues: A Focus on Butyrate, Colon Cancer, Obesity and Insulin Resistance. Nutrients 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Holst, J.J. The physiology of glucagon-like peptide 1. Physiol. Rev. 2007, 87, 1409–1439. [Google Scholar] [CrossRef] [PubMed]
- De Vadder, F.; Kovatcheva-Datchary, P.; Goncalves, D.; Vinera, J.; Zitoun, C.; Duchampt, A.; Backhed, F.; Mithieux, G. Microbiota-generated metabolites promote metabolic benefits via gut-brain neural circuits. Cell 2014, 156, 84–96. [Google Scholar] [CrossRef] [PubMed]
- Petersen, C.; Bell, R.; Klag, K.A.; Lee, S.-H.; Soto, R.; Ghazaryan, A.; Buhrke, K.; Ekiz, H.A.; Ost, K.S.; Boudina, S.; et al. T cell–mediated regulation of the microbiota protects against obesity. Science 2019, 365, eaat9351. [Google Scholar] [CrossRef] [PubMed]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef] [PubMed]
- Shin, N.R.; Lee, J.C.; Lee, H.Y.; Kim, M.S.; Whon, T.W.; Lee, M.S.; Bae, J.W. An increase in the Akkermansia spp. population induced by metformin treatment improves glucose homeostasis in diet-induced obese mice. Gut 2014, 63, 727–735. [Google Scholar] [CrossRef] [PubMed]
- Anhe, F.F.; Roy, D.; Pilon, G.; Dudonne, S.; Matamoros, S.; Varin, T.V.; Garofalo, C.; Moine, Q.; Desjardins, Y.; Levy, E.; et al. A polyphenol-rich cranberry extract protects from diet-induced obesity, insulin resistance and intestinal inflammation in association with increased Akkermansia spp. population in the gut microbiota of mice. Gut 2015, 64, 872–883. [Google Scholar] [CrossRef] [PubMed]
- Dao, M.C.; Everard, A.; Aron-Wisnewsky, J.; Sokolovska, N.; Prifti, E.; Verger, E.O.; Kayser, B.D.; Levenez, F.; Chilloux, J.; Hoyles, L.; et al. Akkermansia muciniphila and improved metabolic health during a dietary intervention in obesity: Relationship with gut microbiome richness and ecology. Gut 2016, 65, 426–436. [Google Scholar] [CrossRef] [PubMed]
- Plovier, H.; Everard, A.; Druart, C.; Depommier, C.; Van Hul, M.; Geurts, L.; Chilloux, J.; Ottman, N.; Duparc, T.; Lichtenstein, L.; et al. A purified membrane protein from Akkermansia muciniphila or the pasteurized bacterium improves metabolism in obese and diabetic mice. Nat. Med. 2017, 23, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Adachi, K.; Sugiyama, T.; Shimozato, A.; Ebi, M.; Ogasawara, N.; Funaki, Y.; Goto, C.; Sasaki, M.; Kasugai, K. Association of Intestinal Microbiota with Metabolic Markers and Dietary Habits in Patients with Type 2 Diabetes. Digestion 2016, 94, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Gentile, C.L.; Weir, T.L. The gut microbiota at the intersection of diet and human health. Science 2018, 362, 776–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, K.E.; Parke, E.C.; O’Malley, M.A. How Causal are Microbiomes? A Comparison with the Helicobacter pylori Explanation of Ulcers. Biol. Philos. 2019, pitt philsci, 15777. [Google Scholar]
- Armour, C.R.; Nayfach, S.; Pollard, K.S.; Sharpton, T.J. A Metagenomic Meta-analysis Reveals Functional Signatures of Health and Disease in the Human Gut Microbiome. mSystems 2019, 4. [Google Scholar] [CrossRef]
- Tannock, G.W. Understanding the Gut Microbiota; John Wiley & Sons: Hoboken, NJ, USA, 2017. [Google Scholar]
- Mallon, C.A.; Elsas, J.D.V.; Salles, J.F. Microbial invasions: The process, patterns, and mechanisms. Trends Microbiol. 2015, 23, 719–729. [Google Scholar] [CrossRef]
- Fuentes, S.; van Nood, E.; Tims, S.; Heikamp-de Jong, I.; ter Braak, C.J.; Keller, J.J.; Zoetendal, E.G.; de Vos, W.M. Reset of a critically disturbed microbial ecosystem: Faecal transplant in recurrent Clostridium difficile infection. ISME J. 2014, 8, 1621–1633. [Google Scholar] [CrossRef]
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, stability and resilience of the human gut microbiota. Nature 2012, 489, 220–230. [Google Scholar] [CrossRef] [Green Version]
- Levy, M.; Kolodziejczyk, A.A.; Thaiss, C.A.; Elinav, E. Dysbiosis and the immune system. Nat. Rev. Immunol. 2017, 17, 219–232. [Google Scholar] [CrossRef]
- Lackey, D.E.; Olefsky, J.M. Regulation of metabolism by the innate immune system. Nat. Rev. Endocrinol. 2016, 12, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Belkaid, Y.; Hand, T.W. Role of the microbiota in immunity and inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef] [PubMed]
- Thaiss, C.A.; Zmora, N.; Levy, M.; Elinav, E. The microbiome and innate immunity. Nature 2016, 535, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking long-term dietary patterns with gut microbial enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [PubMed]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Pereira, F.C.; Berry, D. Microbial nutrient niches in the gut. Environ. Microbiol. 2017, 19, 1366–1378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lachnit, T.; Bosch, T.C.G.; Deines, P. Exposure of the Host-Associated Microbiome to Nutrient-Rich Conditions May Lead to Dysbiosis and Disease Development-an Evolutionary Perspective. MBio 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Chu, N.D.; Smith, M.B.; Perrotta, A.R.; Kassam, Z.; Alm, E.J. Profiling Living Bacteria Informs Preparation of Fecal Microbiota Transplantations. PLoS ONE 2017, 12, e0170922. [Google Scholar] [CrossRef] [PubMed]
- Costello, S.P.; Soo, W.; Bryant, R.V.; Jairath, V.; Hart, A.L.; Andrews, J.M. Systematic review with meta-analysis: Faecal microbiota transplantation for the induction of remission for active ulcerative colitis. Aliment. Pharmacol. Ther. 2017, 46, 213–224. [Google Scholar] [CrossRef]
- Miquel, S.; Martin, R.; Rossi, O.; Bermudez-Humaran, L.G.; Chatel, J.M.; Sokol, H.; Thomas, M.; Wells, J.M.; Langella, P. Faecalibacterium prausnitzii and human intestinal health. Curr. Opin. Microbiol. 2013, 16, 255–261. [Google Scholar] [CrossRef]
- Reunanen, J.; Kainulainen, V.; Huuskonen, L.; Ottman, N.; Belzer, C.; Huhtinen, H.; de Vos, W.M.; Satokari, R. Akkermansia muciniphila Adheres to Enterocytes and Strengthens the Integrity of the Epithelial Cell Layer. Appl. Environ. Microbiol. 2015, 81, 3655–3662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costello, S.P.; Hughes, P.A.; Waters, O.; Bryant, R.V.; Vincent, A.D.; Blatchford, P.; Katsikeros, R.; Makanyanga, J.; Campaniello, M.A.; Mavrangelos, C.; et al. Effect of Fecal Microbiota Transplantation on 8-Week Remission in Patients With Ulcerative Colitis: A Randomized Clinical Trial. JAMA 2019, 321, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Cammarota, G.; Ianiro, G.; Gasbarrini, A. Fecal microbiota transplantation for the treatment of Clostridium difficile infection: A systematic review. J. Clin. Gastroenterol. 2014, 48, 693–702. [Google Scholar] [CrossRef] [PubMed]
- Davidovics, Z.H.; Michail, S.; Nicholson, M.R.; Kociolek, L.K.; Pai, N.; Hansen, R.; Schwerd, T.; Maspons, A.; Shamir, R.; Szajewska, H.; et al. Fecal Microbiota Transplantation for Recurrent Clostridium difficile Infection and Other Conditions in Children: A Joint Position Paper From the North American Society for Pediatric Gastroenterology, Hepatology, and Nutrition and the European Society for Pediatric Gastroenterology, Hepatology, and Nutrition. J. Pediatr. Gastroenterol. Nutr. 2019, 68, 130–143. [Google Scholar] [CrossRef] [PubMed]
- Whitacre, C.C. Sex differences in autoimmune disease. Nat. Immunol. 2001, 2, 777–780. [Google Scholar] [CrossRef] [PubMed]
- Danska, J.S. Sex matters for mechanism. Sci. Transl. Med. 2014, 6, 258fs240. [Google Scholar] [CrossRef] [PubMed]
- Arnold, A.P.; Lusis, A.J. Understanding the sexome: Measuring and reporting sex differences in gene systems. Endocrinology 2012, 153, 2551–2555. [Google Scholar] [CrossRef]
- Markle, J.G.; Frank, D.N.; Mortin-Toth, S.; Robertson, C.E.; Feazel, L.M.; Rolle-Kampczyk, U.; von Bergen, M.; McCoy, K.D.; Macpherson, A.J.; Danska, J.S. Sex differences in the gut microbiome drive hormone-dependent regulation of autoimmunity. Science 2013, 339, 1084–1088. [Google Scholar] [CrossRef]
- Bolnick, D.I.; Snowberg, L.K.; Hirsch, P.E.; Lauber, C.L.; Org, E.; Parks, B.; Lusis, A.J.; Knight, R.; Caporaso, J.G.; Svanback, R. Individual diet has sex-dependent effects on vertebrate gut microbiota. Nat. Commun. 2014, 5, 4500. [Google Scholar] [CrossRef]
- Freire, A.C.; Basit, A.W.; Choudhary, R.; Piong, C.W.; Merchant, H.A. Does sex matter? The influence of gender on gastrointestinal physiology and drug delivery. Int. J. Pharm. 2011, 415, 15–28. [Google Scholar] [CrossRef]
- Org, E.; Mehrabian, M.; Parks, B.W.; Shipkova, P.; Liu, X.; Drake, T.A.; Lusis, A.J. Sex differences and hormonal effects on gut microbiota composition in mice. Gut Microbes 2016, 7, 313–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Study | Country | Follow up (Weeks) | Study Arms | Patients (n) | Age (Years) | Sex (%Female) | Obesity Criteria | Metabolic Syndrome Criteria |
|---|---|---|---|---|---|---|---|---|
| Vrieze et al. 2012 [40] | NL | 6 | FMT | 9 | 47 ± 12 | 0 | BMI > 30 | Waist circumference >102 cm and a FPG level >5.6 mmol/L |
| Placebo | 9 | 53 ± 9 | 0 | |||||
| Koote et al. 2017 [41] | NL | 6 and 18 | FMT | 26 | 54 (49–60) | 0 | BMI ≥ 30 | National Cholesterol Education Program (NCEP) |
| Placebo | 12 | 54 (49–58) | 0 | |||||
| Smits et al. 2018 [42] | NL | 2 | FMT | 10 | 52 ± 7.4 | 0 | BMI ≥ 30 | FPG ≥ 5.6 mmol/L; TG ≥ 1.7 mmol/L; HDL-C < 1.0 mmol/L; blood pressure ≥ 130/85 mm Hg; waist circumference ≥ 102 cm |
| Placebo | 10 | 58 ± 8.5 | 0 |
| Study | Vrieze et al. 2012 [40] | Koote et al. 2017 [41] | Smits et al. 2018 [42] | |||||
|---|---|---|---|---|---|---|---|---|
| Follow-up (weeks) | 6 | 6 | 2 | |||||
| Study Arms | FMT | Placebo | FMT | Placebo | FMT | Placebo | ||
| Patients (n) | 9 | 9 | 13 | 12 | 10 | 10 | ||
| BMI | kg/m2 | BL | 35.7 ± 4.5 | 35.6 ± 4.5 | 33.8 (32.5–35.7) | 35.8 (33.1–40.4) | 33.9 ± 3.9 | 33.8 ± 4 |
| EP | 35.6 ± 4.2 | 35.7 ± 4.8 | 33.6 (32.5–35.8) | 36.1 (32.5–41.5) | - | - | ||
| SBP | mmHg | BL | 138 ± 9 | 140 ± 6 | 141 (132–154 | 148 (134–62) | 148 ± 12 | 152 ± 13 |
| EP | 132 ± 18 | 142 ± 24 | - | - | - | - | ||
| DBP | mmHg | BL | 85 ± 6 | 84 ± 6 | 90 (78–97) | 94 (83–105) | 93 ± 10 | 93 ± 8 |
| EP | 83 ± 15 | 86 ± 18 | - | - | - | - | ||
| FPG | mmol/L | BL | 5.7 ± 0.6 | 5.7 ± 0.6 | 5.5 (5.3–6.1) | 5.9 (5.5–6.4) | 5.8 ± 0.5 | 6.2 ± 0.9 |
| EP | 5.7 ± 0.6 | 5.7 ± 0.6 | 5.6 (5.4–6.9) | 5.9 (5.7–6.7) | 5.9 ± 0.6 | 6.0 ± 0.6 | ||
| HbA1c | mmol/mol | BL | 39 ± 3.3 | 40 ± 4.5 | 40 (36–41) | 43 (36–46) | 36.9 ± 5.1 | 38.7 ± 3.6 |
| EP | 38 ± 3.6 | 39 ± 9.0 | 38 (34–41) | 42 (35–46) | 37.4 ± 3.5 | 38.3 ± 3.9 | ||
| Insulin | pmol/L | BL | 74 (40–230) | 135 (26–220) | 121 (93–143) | 107 (80–159) | 146 ± 63.7 | 107 ± 45.5 |
| EP | 77 (18–250) | 140 (30–287) | 103 (81–126) | 126 (97–171) | 140 ± 67.8 | 121 ± 76.5 | ||
| EGP | μmol/kg/min | BL | 3.8 (2.9–9.8) | 4.6 (2.6–12.1) | 4 (3.3–5.1) | 4.6 (3.6–5.5) | - | - |
| EP | 3.8 (1.2–7.8) | 4.8 (3.9–12.5) | 3.8 (3.2–4.5) | 4.7 (2.9–5.5) | - | - | ||
| Rd | μmol/kg/min | BL | 26.2 (12.6–55.1) | 18.9 (10.8–35.9) | 25.8 (19.3–34.7) | 22.5 (19.6–30.2) | - | - |
| EP | 45.3 (10.6–62.0) | 19.5 (13.5–33.2) | 28.8 (21.4–36.9) | 20.8 (17.6–29.5) | - | - | ||
| Total cholesterol | mmol/L | BL | 4.5 ± 1.2 | 4.8 ± 0.9 | 5.5 (4.8–6.6) | 5.5 (4.8–6.6) | 5.3 ± 0.9 | 5.3 ± 0.9 |
| EP | 4.6 ± 1.2 | 4.8 ± 0.6 | 5.4 (4.8–6.3) | 5.4 (5.1–5.7) | 5.3 ± 0.9 | 5.0 ± 0.7 | ||
| HDL-C | mmol/L | BL | 1.0 ± 0.3 | 1.0 ± 0.3 | 1.1 (0.9–1.4) | 1.0 (0.9–1.1) | 1.1 ± 0.2 | 1.2 ± 0.2 |
| EP | 1.0 ± 0.3 | 0.9 ± 0.3 | 1.1 (1–1.3) | 1.0 (0.9–1.2) | 1.2 ± 0.2 | 1.2 ± 0.2 | ||
| LDL-C | mmol/L | BL | 3.1 ± 1.2 | 2.9 ± 0.6 | 3.9 (3.2–4.5) | 3.7 (3.0–4.8) | 3.3 ± 0.7 | 3.1 ± 1.3 |
| EP | 3.0 ± 0.9 | 2.9 ± 0.6 | 3.8 (3.1–4.5) | 3.5 (3.2–4.1) | 3.5 ± 0.8 | 3.2 ± 0.5 | ||
| TG | mmol/L | BL | 1.4 ± 0.9 | 1.6 ± 0.9 | 1.2 (0.9–1.7) | 1.3 (1.1–1.8) | 1.3 (1–1.6) | 1.3 (1.1–1.6) |
| EP | 1.5 ± 1.2 | 1.8 ± 1.2 | 1.3 (0.9–1.6) | 1.7 (1.2–2.0) | 1.3 (1–2.2) | 1.0 (0.7–1.5) | ||
| Study | Vrieze et al. 2012 [40] | Koote et al. 2017 [41] | Smits et al. 2018 [42] |
|---|---|---|---|
| FMT Route | Nasoduodenal | Nasoduodenal | Nasoduodenal |
| Donor stool | Single unpooled FMT from different lean donors | Single unpooled FMT from different lean omnivorous donors | Single unpooled FMT from different vegan donors |
| Stool preparation | Fresh sample was immediately covered with sterile saline (500 mL, 0.9% NaCl), and stirred in blender (10 min) and filtered twice through metal sieve. | Fresh sample was immediately covered with sterile saline (500 mL, 0.9% NaCl), and stirred in blender (10 min) and filtered twice through metal | Fresh sample was immediately covered with sterile saline (500 mL, 0.9% NaCl), and stirred in blender (10 min) and filtered twice through metal sieve. |
| Stool Dose | Not reported | Not reported | Not reported |
| Time to FMT from stool donation | <6 h | <6 h | <6 h |
| FMT replicates | 1 | 2 | 1 |
| FMT infusion time | 30 min | Not reported | 30 min |
| Adverse event | N/A | No serious events | No serious events |
| Study (Sequencing Method) | Fecal Microbiota Changes in Metabolic Syndrome Patients Relative to Donors | Fecal Microbiota Changes within Group after FMT Infusion | Fecal Microbiota Changes in the FMT Group Relative to the Placebo Group | Microbiota Associated Metabolites Changes Post-FMT Infusion |
|---|---|---|---|---|
| Vrieze et al. 2012 [40] (HITChip microarray) | ↑ Bacteroidetes ↓ Clostridium cluster XIVa | ↑ α-diversity (Observed Species) ↑ Dorea formicigenerans, ↑ Clostridium sphenoides, ↑ Clostridium symbiosum, ↑ Clostridium ramosum, ↑ Clostridium nexile, ↑ Coprobacillus catenaformis, ↑ Ruminococcus gnavus, ↑ Ruminococcus lactaris, ↑ Ruminococcus callidus, ↑ Ruminococcus bromii, ↑ Roseburia intestinalis, ↑ Aneurinibacillus, ↑ Anaerotruncus colihominis, ↑ Eubacterium siraeum, ↑ Sporobacter termitidis, ↑ Oxalobacter formigenes | ↑ Dorea formicigenerans, ↑ Clostridium sphenoides, ↑ Clostridium nexile, ↑ Coprobacillus catenaformis, ↑ Ruminococcus lactaris, ↑ Oxalobacter formigenes | Fecal SCFAs ↓ Acetate ↔ Propionate ↓ Butyrate |
| Kootte et al. 2017 [41] (HITChip microarray) | Not reported | ↔ α-diversity (Shannon index). Composition change in Responders a compared to non-responders ↑ Eubacterium ventriosum ↑ Akkermansia muciniphila ↑ Clostridium sporogenes ↓ Roseburia intestinalis ↓ Bacteroides plebeius | Eubacterium siraeum Lactobacillus ruminis | Fecal SCFAs ↑ Acetate ↔ Propionate ↔ Butyrate Fecal Bile Acids ↑ Cholic acid ↔ Chenodeoxycholic acid ↔ Deoxycholic acid ↔ Lithocholic acid |
| Smits et al. 2018 [42] (HITChip microarray) | ↔ α-diversity (Shannon index) The distinction in fecal microbiota composition between patients and donors is driven by Anaerostipes caccae, Lachnobacterium and Clostridium spp. | ↔ α-diversity (Shannon index) | Bryantella formatexigens Megamonas hypermegale Lachnobacterium bovis Clostridium cluster XIVa | TMAO Metabolites ↔ Plasma TMAO ↔ Urinary TMA Excretion ↔ Urinary TMA Excretion |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Mocanu, V.; Cai, C.; Dang, J.; Slater, L.; Deehan, E.C.; Walter, J.; Madsen, K.L. Impact of Fecal Microbiota Transplantation on Obesity and Metabolic Syndrome—A Systematic Review. Nutrients 2019, 11, 2291. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11102291
Zhang Z, Mocanu V, Cai C, Dang J, Slater L, Deehan EC, Walter J, Madsen KL. Impact of Fecal Microbiota Transplantation on Obesity and Metabolic Syndrome—A Systematic Review. Nutrients. 2019; 11(10):2291. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11102291
Chicago/Turabian StyleZhang, Zhengxiao, Valentin Mocanu, Chenxi Cai, Jerry Dang, Linda Slater, Edward C. Deehan, Jens Walter, and Karen L. Madsen. 2019. "Impact of Fecal Microbiota Transplantation on Obesity and Metabolic Syndrome—A Systematic Review" Nutrients 11, no. 10: 2291. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11102291