1. Introduction
The need for transdisciplinary efforts to bridge the gap between production agriculture and public health nutrition is clear [
1]. A particularly useful channel through which this can occur is among scientists working in horticulture and in human health since horticulture is a science familiar to most consumers and is practiced by many. In particular, it is critical for nutritionists to recognize the many factors that are involved in providing consumers with health-beneficial cultivated varieties (cultivars) of a crop and that potential value-added health benefits may result if cultivar-specific food labeling becomes a reality [
2,
3]. From a horticultural perspective, this creates the opportunity for niche markets and for the direct marketing of specific cultivars to the consumer [
4,
5]. One crop that has received limited attention relative to its health benefits, particularly those related to phenolic compounds, is snap bean (
Phaseolus vulgaris L.), which is the topic of this investigation.
Snap beans (also known as garden, green, string, or French beans) are a form of common bean. As opposed to dry bean, in which the mature seeds are eaten, the immature pods and seeds of snap beans are consumed as a vegetable. Dry beans are considered a good source of protein, complex carbohydrates, soluble fiber, folic acid and vitamin B. In contrast, because of their higher water content (~90% water compared to ~12%), snap beans have lower protein and total carbohydrate levels, but they possess certain vitamins and carotenoids that dry beans do not have or have only in trace amounts. These include vitamin C, β-carotene, lutein and zeaxanthin, α-tocopherol, and phylloquinone [
6]. About 1.5 million Mg of snap beans are produced annually on a global basis [
7]. The US accounts for 71% of world production of snap beans, and Americans consume about 3.0 kg per person per year [
8], and as such, snap beans are a significant source of vitamins, carotenoids and other secondary plant compounds in the human diet.
Among secondary plant compounds are phenolics and related flavonoids. Phenolics constitute a large class of phytochemicals with over 8000 individual compounds having been identified. Structurally, they are characterized by an aromatic ring with one or more hydroxyl groups [
9,
10,
11]. Phenolics and flavonoids are further divided into flavonols, flavones, flavan-3-ols, anthocyanidins, flavanones, isoflavones, and other minor subclasses, and nonflavonoids, which include phenolic acids, hydroxycinnamates, and stilbenes [
9].
In general, phenolics are associated with various human health benefits including acting as anticarcinogens, antimicrobial agents, anti-inflammatories and improving vascular health, possibly by their metabolism via the gut-associated microbiome [
12]. Total phenolic content has a strong antioxidant activity in legumes [
13,
14,
15]. Dry bean consumption is associated with a reduced risk of diabetes, heart disease, colon cancer, and prostate cancer, and the mitigation of obesity [
16,
17,
18,
19]. These benefits may accrue in part from the phenolic content of seeds. We know less about the health benefits of snap beans, and to our knowledge, no human health studies similar to those conducted for dry beans have been reported.
Several research groups have characterized snap beans for various phenolics. Abu-Reidah et al. [
20] found more than 72 compounds including 10 phenolic acids and 59 flavonoids. Predominant among these were the flavonol glucosides quercetin and kaempferol along with the phenolic acids (+) catechin and (−) epicatechin, chlorogenic and protocatechuic acids, as well as procyanidins [
21], [
22]. Flavonoid levels in processed fresh snap beans ranged from 20 to 51 mg kg
−1 [
23]. In a study of six wax and green bean cultivars, flavonols levels ranged from 19 to 184 µg g
−1 fresh weight (FW) for quercetin derivatives and 6 to 15 µg g
−1 for kaempferol derivatives [
24]. Kleintop et al. [
25] examined the total phenolic content (TPC) in fresh and frozen pods of a snap bean diversity panel of 149 accessions. They found more than a four-fold difference among cultivars, which ranged from 0.29 to 1.31 mg g
−1 gallic acid equivalents (GAE). Higher levels were generally associated with cultivars that had colored seeds and a purple or pink flower color.
Snap beans grown for processing vs. the fresh market have different quality characteristics that may affect TPC. Processors require that cultivars be white-seeded to ensure that no water-soluble flavonoids are produced in any plant parts including the pods as they will leach into the liquor in canned beans and present as a colored ring in the testa in cut frozen beans, thereby reducing quality [
26]. The standards for fresh market beans are less stringent, and while colored-seeded cultivars are acceptable, they are generally brown-seeded (produced by flavonols) rather than black or purple (a result of anthocyanins) because the former are less noticeable in the pod. Brown-seeded cultivars will generally have pods that are green in appearance, while purple- or black-seeded cultivars may have solid purple or striped purple and green pods.
In common bean, several gene combinations may give a white seed color, but in the commercial trade, essentially all white-seeded snap beans possess the recessive
p gene. This gene is a major switch in the flavonoid biosynthetic pathway, with the dominant form allowing the expression of other color genes while the recessive allele blocks expression.
P is located on Pv07 (
Table 1) and appears to be a basic helix loop helix (bHLH) MYB transcription factor [
27]. Other effects of
p are a reduced lignin content in seeds and greater sensitivity to imbibitional injury and reduced germination and emergence if the seed is not protected with fungicides [
28,
29]. The types of flavonols and anthocyanins produced and their expression are controlled by more than 10 additional genes [
30]. These can be divided into those that act in the flavonoid biosynthetic pathway and those that affect spatial and temporal expression [
31]. Most but not all of these factors have been mapped with varying degrees of accuracy [
27,
32,
33] (
Table 1). Little is known about the genetic control of phenolic acids in common bean. Kleintop et al. [
25] observed a substantial difference in TPC in colored-seeded vs. white-seeded accessions in a snap bean diversity panel, but within white-seeded types, variation in TPC was also noted, suggesting that there is a genetic variation for colorless phenolic compounds.
Genome-Wide Association Studies (GWAS) are a means of identifying and mapping quantitative trait loci (QTL) associated with phenotypic traits in plant populations. GWAS relies on linkage disequilibrium in individuals in regions in which QTL reside. A number of association-mapping studies have been conducted on common bean. These have analyzed traits associated with phenological variation [
36,
37,
38], agronomic performance [
39,
40], biological nitrogen fixation [
41], plant architecture [
42], seed quality [
43,
44], and abiotic [
45,
46] and biotic [
47,
48,
49], stress variation. None have focused on traits specific to snap bean. Association studies of phenolics in non-legume crops have focused primarily on cereals such as barley [
50,
51], whole rice [
52,
53,
54], dehulled rice [
55], wheat [
56], sorghum [
57], and maize [
58], with single studies of tomato [
59], rapeseed [
60], and olive [
61].
Considering the fairly substantial intake of snap beans in the United States, improving the phenolic content of snap beans could have a meaningful impact on human health. Therefore, the objective of this study was to conduct GWAS on a snap bean diversity panel that had been evaluated for total phenolic content. We wanted to 1) identify genomic regions associated with variation in TPC, 2) determine whether known color genes co-segregate with these regions, and 3) identify potentially novel genes that regulate TPC, especially in white-seeded cultivars, as a basis for further improving snap bean phenolic content.
4. Discussion
Because anthocyanins are well known for their red-to-blue color, and flavonols are associated with yellow-to-brown pigments, we classified snap bean accessions for flower and seed color as well as obtaining a color profile for pods, in addition to measuring TPC. However, other phenolic compounds may be colorless and may account for the two-fold variation in TPC that was observed apart from that attributed to flavonoids. Kleintop et al. [
25] found moderate significant correlations between TPC and a* (0.40) and between TPC and b* (−0.34). There was essentially no correlation between TPC and L*. These results indicate that, of the parameters evaluated here, a* and b* are more likely to be associated with QTN for TPC than L*.
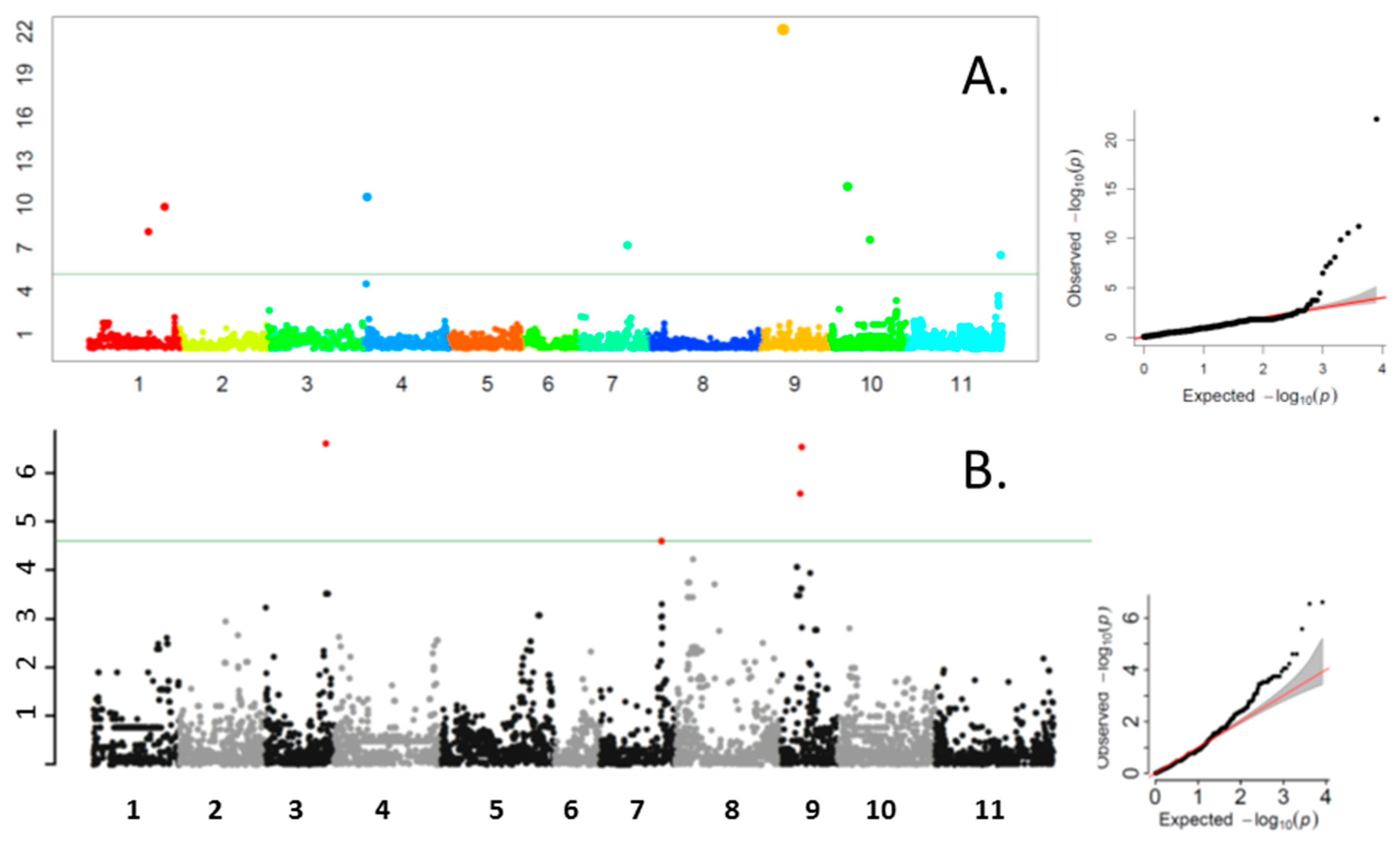
In this association study, we found 11 QTN for TPC located on seven chromosomes of common bean, but only one QTN (TPC7.1) was associated with a known major gene that has a large phenotypic effect. In addition, the QTN was located 324–245kb from the gene models associated with this trait. In a dry x snap bean mapping population, Hagerty et al. [
32] found SNP markers associated with
p that were located ~17.2Mb from the candidate genes identified by [
27]. While the slow rate of LD decay in common bean may influence marker-gene associations,
p is not located in a low recombination region of Pv07, and so there may be other explanations for why distances between this locus and markers are rather large.
Compared to the other phenotypic categories, there were relatively few pod color SNPs, and most were not likely to be associated with pigmented flavonoids. Based on the moderate correlations between pod color and total TPC identified in [
25], we would expect that a* and b* would be most likely to be associated with phenolics; in this case, however, a* (and L*) are associated with the wax bean trait, which is marked by a deficit of chlorophyll and not variation in TPC. Three b* QTN may be related to phenolic-based pod color, which is supported by B*3.2-associated SNP also underlying TPC 3.1. In addition, FC 3.1 was located within the LD decay window at 255 kb from B*3.2 and TPC 3.1. These results all support the idea that a QTN affecting flower and pod color and phenolic content resides in this region of Pv03. This QTN does not appear to be related to any known color genes. All b* QTN may represent novel genes affecting pod color that have not been identified in classical genetic studies.
The findings for flower color QTN were unexpected. We predicted that we would find two or three QTN associated with
P and
V, and possibly
T, because these genes are known to modify flower color. As described above,
P is located on Pv07, whereas
V has previously been placed on Pv06 (
Table 1). Dominant
P is required for observing segregation at
V and
T, which means that the number of genotypes that could be classified for these genes was small, and any associations that might have been found in GWAS would be discarded because they fell below a 5% MAF cut-off. The 13 QTN associated with flower color appeared to be novel and it is difficult to see how these were identified during GWAS, since the classification of genotypes was based on a three-point scale. This may point to the need for the better quantification of flower color. The pink class in particular can sometimes be difficult to classify as it varies from very similar to white on one hand to purple on the other hand. One QTN (FC 9.1) had the same underlying SNP as a TPC QTN (TPC 9.1), which suggests that the pigment difference in that case was real.
The data used in this study represent a single environment. In our previous study [
25], we examined TPC over two years in a subset of pole and bush bean accessions. We found no genotype by the year of interaction for the indeterminate pole bean group, and a weak genotype by the year of interaction for determinant bush accessions. TPC levels were significantly higher in the second year, and the interaction term for the bush bean group was driven by changes in magnitude—particularly by one accession—rather than by cross-over interaction. Based on these results, we concluded that the genotype by environment effect on TPC in this population was relatively small, and that we could proceed with a GWAS analysis based on the single environment. In some cases, the magnitude of the TPC’s effects from GWAS in the present situation was small and would need to be verified in more environments to provide validation. With additional resources, it would be useful to conduct a more extensive analysis of environmental effects as well as other factors, such as pod maturity, on TPC.
While most structural genes in the phenolic biosynthetic pathway have been identified, the regulatory control of the pathway is in the early stages of analysis. Some regulatory factors have been identified, with the majority of these characterized in Arabidopsis [
66,
67,
68,
69,
70,
71] and grape [
80]. One difficulty is that a function in one species is not necessarily the same in other species, especially where particular biosynthetic pathways may be missing (such as the glucosinolate biosynthetic pathway found in Brassicaceae but not in most other plant families). This works both ways, in that regulatory genes identified as being in the phenolics biosynthetic pathway in Arabidopsis may be involved in different pathways in another organism and vice versa. The lack of functional studies for regulatory genes in legumes in general and common bean in particular hampers the association of putative phenolics biosynthetic pathway regulatory genes with gene function. It may be that for several QTN (TPC 1.2, TPC 3.1, TPC 10.1 and TPC 10.2), novel genes may be involved. In the present case, these may take the form of unrecognized regulatory or structural genes, or genes that have yet to receive annotation in either model systems, such as Arabidopsis, or in the species of interest. The same may be true for those QTN for which we did identify potential genes. There may be another unrecognized candidate gene in the vicinity of the QTN. The ultimate proof will be research that demonstrates gene function.
The identification of QTN for health-promoting phenolics will be an asset to plant breeders. High-throughput molecular markers can readily be derived from SNP positions that have been identified as associated to a trait. Marker-assisted selection, along with simple colorimetric assays as described by [
25], will result in a powerful set of tools for selecting for these traits. This will not only benefit fresh market beans but also processing beans because the white-seeded varieties required by processors can be bred for higher TPC. As previous research [
25] has shown, the cultivar McCaslan No. 42 may be highly valuable with regard to developing white-seeded processing beans with increased TPC levels, and molecular markers will greatly assist in moving these traits into other snap bean backgrounds. Considering the fairly substantial intake of snap beans in the United States, improving the phenolic content of snap beans could have a meaningful impact on human health.

 ,
, 

{kind=link}