Effect of Maternal Obesity in Mice on IL-6 Levels and Placental Endothelial Cell Homeostasis

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Procedures and Tissue Preparation
2.2. qPCR Analysis
2.3. Protein Isolation and Western Blot Analysis
2.4. Histological Stainings (Immunohistochemistry, Immunofluorescence, Stereology)
2.5. Analysis of IF Sections
2.6. Stereological Analysis of Sections
2.7. IL-6 ELISA
2.8. Cell Culture Assays
2.9. Statistical Analysis
3. Results
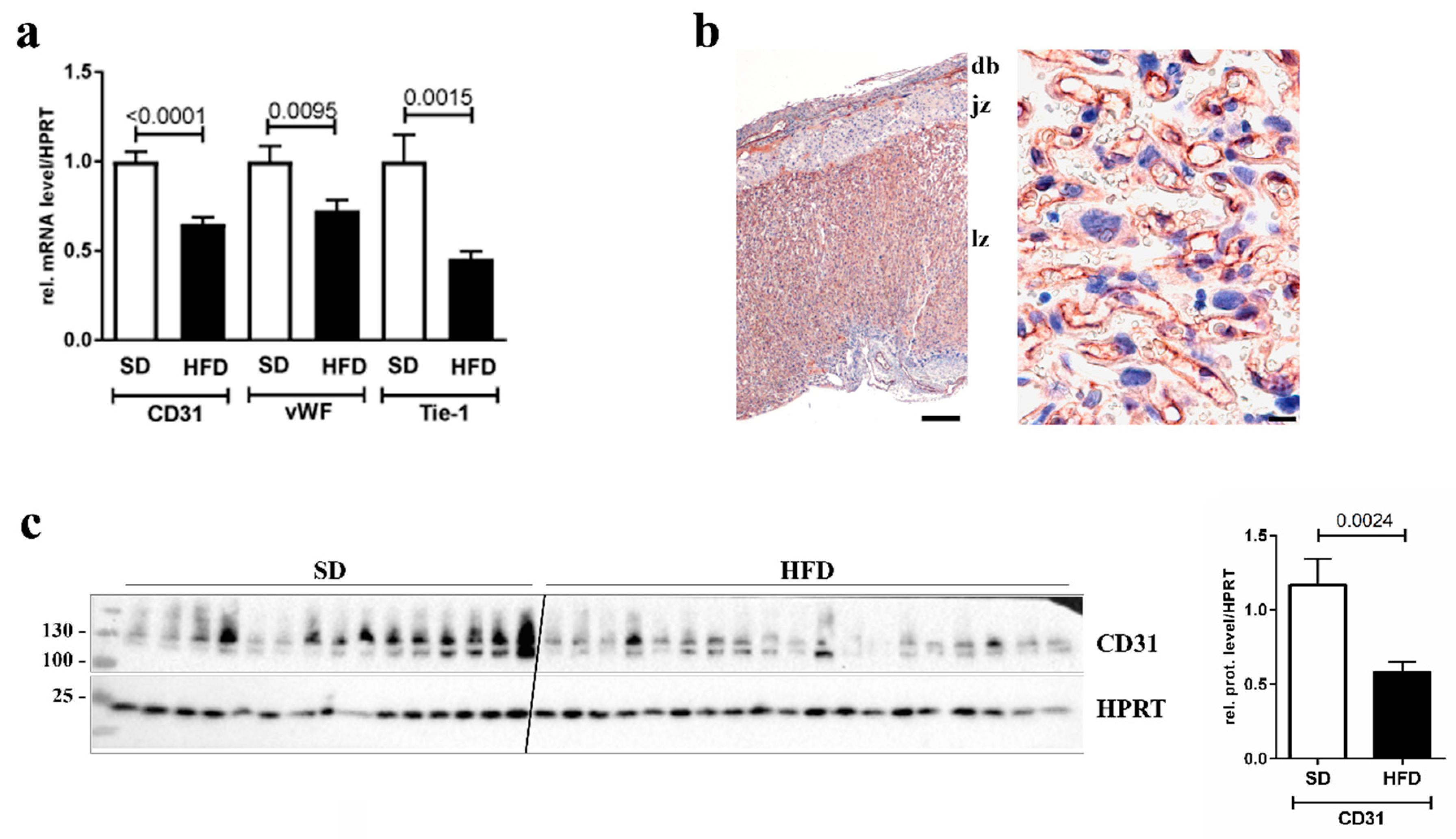
3.1. Downregulation of Endothelial Cell Markers in Placentas of Obese Dams
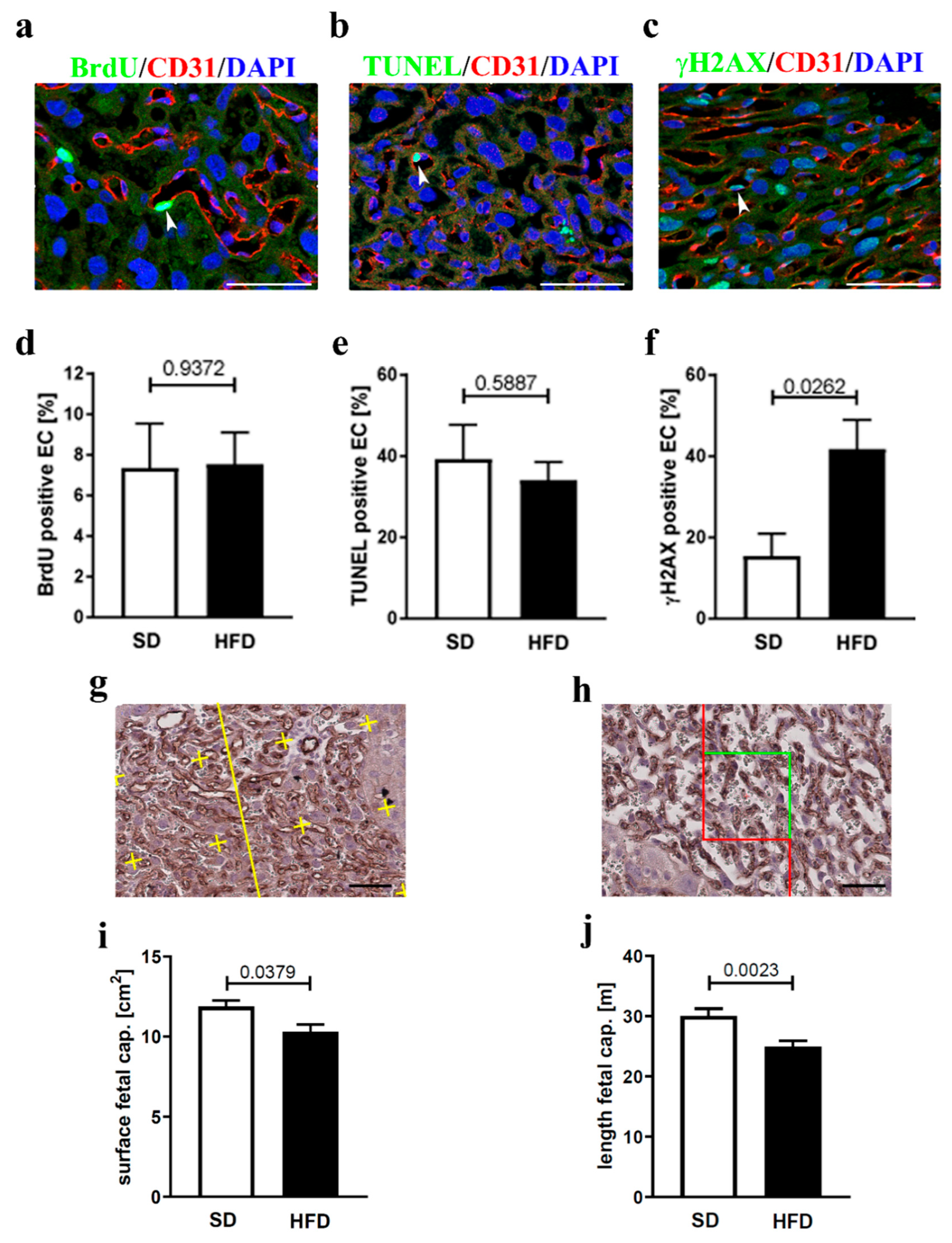
3.2. Endothelial Cell Homeostasis and Vessel Structure
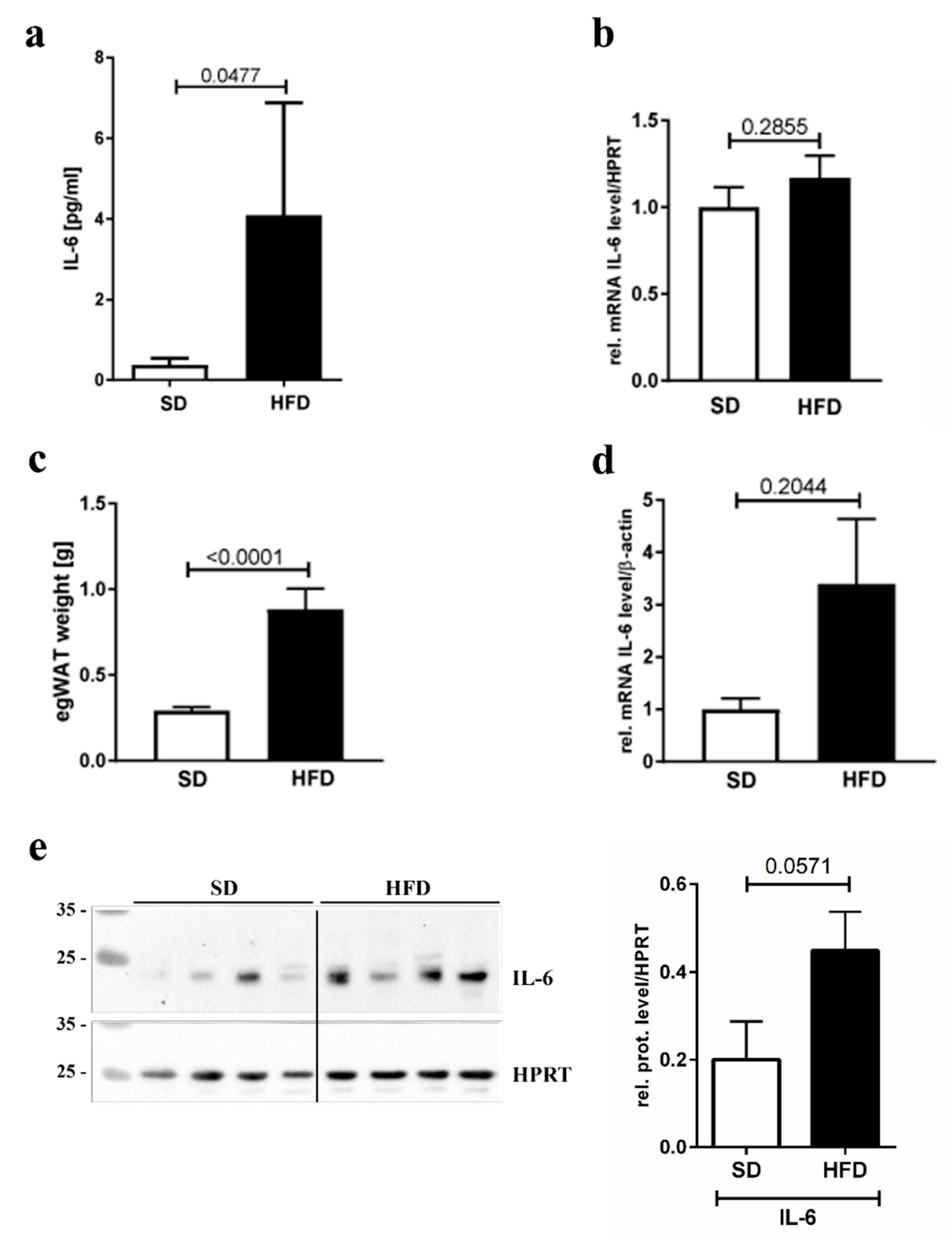
3.3. IL-6 Inflammation Marker Levels
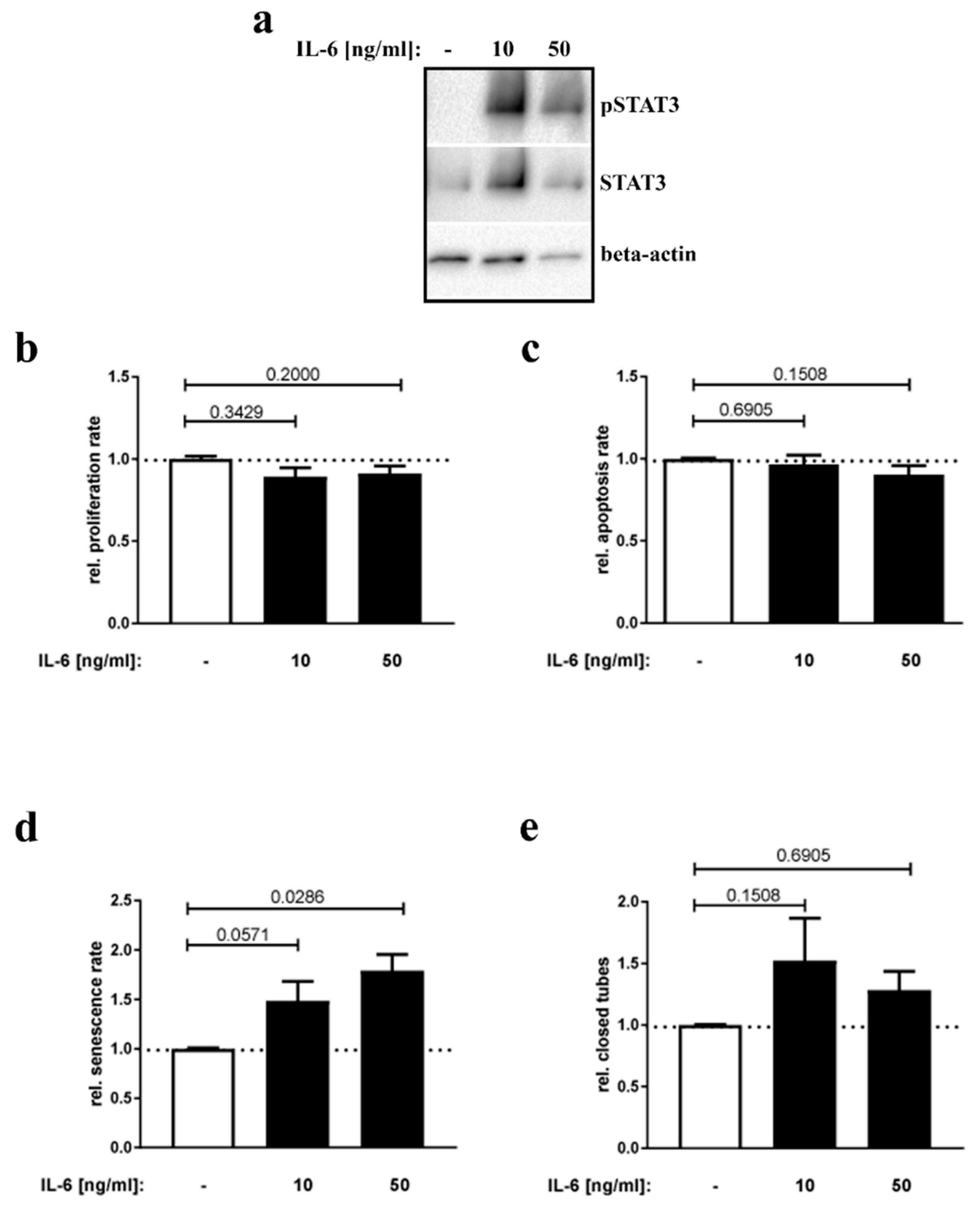
3.4. IL-6 Stimulation of Placental Endothelial Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Linne, Y. Effects of obesity on women’s reproduction and complications during pregnancy. Obes. Rev. 2004, 5, 137–143. [Google Scholar] [CrossRef]
- Perlow, J.H.; Morgan, M.A.; Montgomery, D.; Towers, C.V.; Porto, M. Perinatal outcome in pregnancy complicated by massive obesity. Am. J. Obs. Gynecol. 1992, 167, 958–962. [Google Scholar] [CrossRef]
- Rode, L.; Nilas, L.; Wøjdemann, K.; Tabor, A. Obesity-related complications in Danish single cephalic term pregnancies. Obs. Gynecol. 2005, 105, 537–542. [Google Scholar] [CrossRef] [PubMed]
- Nohr, E.A.; Bech, B.H.; Davies, M.J.; Frydenberg, M.; Henriksen, T.B.; Olsen, J. Prepregnancy obesity and fetal death: A study within the Danish National Birth Cohort. Obs. Gynecol. 2005, 106, 250–259. [Google Scholar] [CrossRef] [PubMed]
- Boney, C.M.; Verma, A.; Tucker, R.; Vohr, B.R. Metabolic syndrome in childhood: Association with birth weight, maternal obesity, and gestational diabetes mellitus. Pediatrics 2005, 115, e290–e296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, L.; Liu, J.; Feng, L.; Chen, Y.; Zhang, J.; Wang, W. Maternal prepregnancy obesity is associated with higher risk of placental pathological lesions. Placenta 2014, 35, 563–569. [Google Scholar] [CrossRef] [PubMed]
- Kovo, M.; Zion-Saukhanov, E.; Schreiber, L.; Mevorach, N.; Divon, M.; Ben-Haroush, A.; Bar, J. The Effect of Maternal Obesity on Pregnancy Outcome in Correlation with Placental Pathology. Reprod. Sci. 2015, 22, 1643–1648. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; DeCourcy, K.; Prater, M.R. High-saturated-fat diet induces gestational diabetes and placental vasculopathy in C57BL/6 mice. Metabolism 2010, 59, 943–950. [Google Scholar] [CrossRef]
- Ma, Y.; Zhu, M.J.; Zhang, L.; Hein, S.M.; Nathanielsz, P.W.; Ford, S.P. Maternal obesity and overnutrition alter fetal growth rate and cotyledonary vascularity and angiogenic factor expression in the ewe. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 299, R249–R258. [Google Scholar] [CrossRef]
- Hayes, E.K.; Lechowicz, A.; Petrik, J.J.; Storozhuk, Y.; Paez-Parent, S.; Dai, Q.; Samjoo, I.A.; Mansell, M.; Gruslin, A.; Holloway, A.C.; et al. Adverse fetal and neonatal outcomes associated with a life-long high fat diet: Role of altered development of the placental vasculature. PLoS ONE 2012, 7, e33370. [Google Scholar] [CrossRef] [Green Version]
- Hayes, E.K.; Tessier, D.R.; Percival, M.E.; Holloway, A.C.; Petrik, J.J.; Gruslin, A.; Raha, S. Trophoblast invasion and blood vessel remodeling are altered in a rat model of lifelong maternal obesity. Reprod. Sci. 2014, 21, 648–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denison, F.C.; Roberts, K.A.; Barr, S.M.; Norman, J.E. Obesity, pregnancy, inflammation, and vascular function. Reproduction 2010, 140, 373–385. [Google Scholar] [CrossRef] [PubMed]
- Christian, L.M.; Porter, K. Longitudinal changes in serum proinflammatory markers across pregnancy and postpartum: Effects of maternal body mass index. Cytokine 2014, 70, 134–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, F.M.; Freeman, D.J.; Ramsay, J.E.; Greer, I.A.; Caslake, M.; Ferrell, W.R. Longitudinal assessment of maternal endothelial function and markers of inflammation and placental function throughout pregnancy in lean and obese mothers. J. Clin. Endocrinol. Metab. 2007, 92, 969–975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pantham, P.; Aye, I.L.M.H.; Powell, T.L. Inflammation in maternal obesity and gestational diabetes mellitus. Placenta 2015, 36, 709–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheller, J.; Chalaris, A.; Schmidt-Arras, D.; Rose-John, S. The pro- and anti-inflammatory properties of the cytokine interleukin-6. Biochim. Et Biophys. Acta 2011, 1813, 878–888. [Google Scholar] [CrossRef] [Green Version]
- Barnes, T.C.; Spiller, D.G.; Anderson, M.E.; Edwards, S.W.; Moots, R.J. Endothelial activation and apoptosis mediated by neutrophil-dependent interleukin 6 trans-signalling: A novel target for systemic sclerosis? Ann. Rheum. Dis. 2011, 70, 366–372. [Google Scholar] [CrossRef] [Green Version]
- Obrenović-Kirćanski, B.; Mikić, A.; Velinović, M.; Bozić, V.; Kovacević-Kostić, N.; Karan, R.; Parapid, B.; Djukić, P.; Savić, D.; Vranes, M. Right ventricular myxoma—A case report. Vojnosanit Pregled 2013, 70, 609–611. [Google Scholar] [CrossRef]
- Appel, S.; Grothe, J.; Storck, S.; Janoschek, R.; Bae-Gartz, I.; Wohlfarth, M.; Handwerk, M.; Hucklenbruch-Rother, E.; Gellhaus, A.; Dötsch, J. Potential Role for GSK3beta in Glucose-Driven Intrauterine Catch-Up Growth in Maternal Obesity. Endocrinology 2019, 160, 377–386. [Google Scholar] [CrossRef] [Green Version]
- Appel, S.; Schulze-Edinghausen, M.; Kretschmer, T.; Storck, S.; Janoschek, R.; Bae-Gartz, I.; Handwerk, M.; Wohlfarth, M.; Nüsken, K.-D.; Hucklenbruch-Rother, E.; et al. Maternal obesity attenuates predelivery inflammatory reaction in C57BL/6N mice. J. Reprod. Immunol. 2017, 122, 10–13. [Google Scholar] [CrossRef]
- Bae-Gartz, I.; Janoschek, R.; Kloppe, C.-S.; Vohlen, C.; Roels, F.; Oberthür, A.; Alcázar, M.A.; Lippach, G.; Muether, P.S.; Dinger, K.; et al. Running Exercise in Obese Pregnancies Prevents IL-6 Trans-signaling in Male Offspring. Med. Sci. Sports Exerc. 2016, 48, 829–838. [Google Scholar] [CrossRef] [PubMed]
- Appel, S.; Turnwald, E.-M.; Alejandre-Alcazar, M.; Ankerne, J.; Rother, E.; Janoschek, R.; Wohlfarth, M.; Vohlen, C.; Schnare, M.; Meißner, U.; et al. Leptin does not induce an inflammatory response in the murine placenta. Horm. Metab. Res. 2014, 46, 384–389. [Google Scholar] [CrossRef] [PubMed]
- Appel, S.; Turnwald, E.M.; Ankerne, J.; Wohlfarth, M.; Appel, J.; Rother, E.; Janoschek, R.; Alcazar, M.A.A.; Schnare, M.; Meißner, U.; et al. Hypoxia-Mediated Soluble Fms-Like Tyrosine Kinase 1 Increase is Not Attenuated in Interleukin 6-Deficient Mice. Reprod. Sci. 2015, 22, 735–742. [Google Scholar] [CrossRef] [PubMed]
- Howard, V.; Reed, M.G. Unbiased Stereology: Three-Dimensional Measurements in Microscopy; Springer: New York, NY, USA, 1998. [Google Scholar]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Baddeley, A.J.; Gundersen, H.J.; Cruz-Orive, L.M. Estimation of surface area from vertical sections. J. Microsc. 1986, 142, 259–276. [Google Scholar] [CrossRef]
- Gundersen, H.J.G.; Bendtsen, T.F.; Korbo, L.; Marcussen, N.; Møller, A.; Nielsen, K.; Nyengaard, J.R.; Pakkenberg, B.; Sørensen, F.B.; Vesterby, A.; et al. Some new, simple and efficient stereological methods and their use in pathological research and diagnosis. APMIS 1988, 96, 379–394. [Google Scholar] [CrossRef]
- Karimu, A.L.; Burton, G.J. The effects of maternal vascular pressure on the dimensions of the placental capillaries. Br. J. Obs. Gynaecol. 1994, 101, 57–63. [Google Scholar] [CrossRef]
- Coan, P.M.; Ferguson-Smith, A.C.; Burton, G.J. Developmental dynamics of the definitive mouse placenta assessed by stereology. Biol. Reprod. 2004, 70, 1806–1813. [Google Scholar] [CrossRef]
- Kipkeew, F.; Kirsch, M.; Klein, D.; Wuelling, M.; Winterhager, E.; Gellhaus, A. CCN1 (CYR61) and CCN3 (NOV) signaling drives human trophoblast cells into senescence and stimulates migration properties. Cell Adhes. Migr. 2016, 10, 163–178. [Google Scholar] [CrossRef]
- Dahlgren, J.; Samuelsson, A.-M.; Jansson, T.; Holmäng, A.; Holm, A. Interleukin-6 in the maternal circulation reaches the rat fetus in mid-gestation. Pediatr. Res. 2006, 60, 147–151. [Google Scholar] [CrossRef] [Green Version]
- Zaretsky, M.V.; Alexander, J.M.; Byrd, W.; Bawdon, R.E. Transfer of inflammatory cytokines across the placenta. Obs. Gynecol. 2004, 103, 546–550. [Google Scholar] [CrossRef] [PubMed]
- Fischer, M.; Goldschmitt, J.; Peschel, C.; Brakenhoff, J.P.; Kallen, K.J.; Wollmer, A.; Grötzinger, J.; Rose-John, S. A bioactive designer cytokine for human hematopoietic progenitor cell expansion. Nat. Biotechnol. 1997, 15, 142–145. [Google Scholar] [CrossRef] [PubMed]
- Nüsken, K.-D.; Warnecke, C.; Hilgers, K.F.; Schneider, H. Intrauterine growth after uterine artery ligation in rats: Dependence on the fetal position in the uterine horn and need for prenatal marking of the animals. J. Hypertens. 2007, 25, 247–248. [Google Scholar] [CrossRef] [PubMed]
- De Clercq, K.; Persoons, E.; Napso, T.; Luyten, C.; Parac-Vogt, T.N.; Sferruzzi-Perri, A.N.; Kerckhofs, G.; Vriens, J. High-resolution contrast-enhanced microCT reveals the true three-dimensional morphology of the murine placenta. Proc. Natl. Acad. Sci. USA 2019, 116, 13927–13936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalisch-Smith, J.; Simmons, D.; Dickinson, H.; Moritz, K. Review: Sexual dimorphism in the formation, function and adaptation of the placenta. Placenta 2017, 54, 10–16. [Google Scholar] [CrossRef]
- Basak, K.; Deán-Ben, X.L.; Gottschalk, S.; Reiss, M.; Razansky, D. Non-invasive determination of murine placental and foetal functional parameters with multispectral optoacoustic tomography. Light. Sci. Appl. 2019, 8, 71. [Google Scholar] [CrossRef] [Green Version]
- Boing, M.; Brand-Saberi, B.; Napirei, M. Murine transcription factor Math6 is a regulator of placenta development. Sci. Rep. 2018, 8, 14997. [Google Scholar] [CrossRef]
- Arthuis, C.J.; Novell, A.; Raes, F.; Escoffre, J.-M.; Lerondel, S.; Le Pape, A.; Bouakaz, A.; Perrotin, F. Real-Time Monitoring of Placental Oxygenation during Maternal Hypoxia and Hyperoxygenation Using Photoacoustic Imaging. PLoS ONE 2017, 12, e0169850. [Google Scholar] [CrossRef] [Green Version]
- Janoschek, R.; Bae-Gartz, I.; Vohlen, C.; Alcázar, M.A.A.; Dinger, K.; Appel, S.; Dötsch, J.; Hucklenbruch-Rother, E. Dietary intervention in obese dams protects male offspring from WAT induction of TRPV4, adiposity, and hyperinsulinemia. Obesity 2016, 24, 1266–1273. [Google Scholar] [CrossRef] [Green Version]
- Odermatt, A. The Western-style diet: A major risk factor for impaired kidney function and chronic kidney disease. American journal of physiology. Ren. Physiol. 2011, 301, F919–F931. [Google Scholar] [CrossRef] [Green Version]
- Sferruzzi-Perri, A.N.; Vaughan, O.R.; Haro, M.; Cooper, W.N.; Musial, B.; Charalambous, M.; Pestana, D.; Ayyar, S.; Ferguson-Smith, A.C.; Burton, G.J.; et al. An obesogenic diet during mouse pregnancy modifies maternal nutrient partitioning and the fetal growth trajectory. FASEB J. 2013, 27, 3928–3937. [Google Scholar] [CrossRef] [PubMed]
- Hariri, N.; Thibault, L. High-fat diet-induced obesity in animal models. Nutr. Res. Rev. 2010, 23, 270–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyank, H. Endothelial cell damage in pre-eclampsia. Zentralblatt fur Gynakologie 1991, 113, 487–492. [Google Scholar] [PubMed]
- Iantorno, M.; Campia, U.; Di Daniele, N.; Nistico, S.; Forleo, G.B.; Cardillo, C.; Tesauro, M. Obesity, inflammation and endothelial dysfunction. J. Boil. Regul. Homeost. Agents 2014, 28, 169–176. [Google Scholar]
- Hohos, N.M.; Skaznik-Wikiel, M.E. High-Fat Diet and Female Fertility. Endocrinology 2017, 158, 2407–2419. [Google Scholar] [CrossRef]
- Song, L.; Sun, B.; Boersma, G.J.; Cordner, Z.A.; Yan, J.; Moran, T.H.; Tamashiro, K.L. Prenatal high-fat diet alters placental morphology, nutrient transporter expression, and mtorc1 signaling in rat. Obesity 2017, 25, 909–919. [Google Scholar] [CrossRef] [Green Version]
- Mahany, E.B.; Han, X.; Borges, B.C.; Cruz-Machado, S.D.S.; Allen, S.J.; Garcia-Galiano, D.; Hoenerhoff, M.J.; Bellefontaine, N.H.; Elias, C.F. Obesity and High-Fat Diet Induce Distinct Changes in Placental Gene Expression and Pregnancy Outcome. Endocrinology 2018, 159, 1718–1733. [Google Scholar] [CrossRef] [Green Version]
- Frias, A.E.; Morgan, T.K.; Evans, A.E.; Rasanen, J.; Oh, K.Y.; Thornburg, K.L.; Grove, K.L. Maternal high-fat diet disturbs uteroplacental hemodynamics and increases the frequency of stillbirth in a nonhuman primate model of excess nutrition. Endocrinology 2011, 152, 2456–2464. [Google Scholar] [CrossRef] [Green Version]
- Barut, F.; Barut, A.; Gun, B.D.; Kandemir, N.O.; Harma, M.I.; Harma, M.; Aktunc, E.; Ozdamar, S.O. Intrauterine growth restriction and placental angiogenesis. Diagn. Pathol. 2010, 5, 24. [Google Scholar] [CrossRef] [Green Version]
- Arroyo, J.A.; Winn, V.D. Vasculogenesis and angiogenesis in the IUGR placenta. Semin. Perinatol. 2008, 32, 172–177. [Google Scholar] [CrossRef]
- Savant, S.; La Porta, S.; Budnik, A.; Busch, K.; Hu, J.; Tisch, N.; Korn, C.; Valls, A.F.; Benest, A.V.; Terhardt, D.; et al. The Orphan Receptor Tie1 Controls Angiogenesis and Vascular Remodeling by Differentially Regulating Tie2 in Tip and Stalk Cells. Cell Rep. 2015, 12, 1761–1773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, B.; Sukhatme, V.P. Suppression of Tie-1 in endothelial cells in vitro induces a change in the genome-wide expression profile reflecting an inflammatory function. FEBS Lett. 2009, 583, 1023–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, P.; Chen, N.; Jia, J.-H.; Gao, X.-J.; Li, S.-H.; Cai, J.; Wang, Z. Tie-1: A potential target for anti-angiogenesis therapy. Acta Acad. Med. Wuhan 2015, 35, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Kayisli, U.; Çayli, S.; Seval, Y.; Tertemiz, F.; Huppertz, B.; Demir, R. Spatial and Temporal Distribution of Tie-1 and Tie-2 During Very Early Development of the Human Placenta. Placenta 2006, 27, 648–659. [Google Scholar] [CrossRef] [PubMed]
- Rogakou, E.P.; Boon, C.; Redon, C.; Bonner, W.M. Megabase Chromatin Domains Involved in DNA Double-Strand Breaks in Vivo. J. Cell Boil. 1999, 146, 905–916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ichijima, Y.; Sakasai, R.; Okita, N.; Asahina, K.; Mizutani, S.; Teraoka, H. Phosphorylation of histone H2AX at M phase in human cells without DNA damage response. Biochem. Biophys. Res. Commun. 2005, 336, 807–812. [Google Scholar] [CrossRef] [PubMed]
- Lawless, C.; Wang, C.; Jurk, D.; Merz, A.; Von Zglinicki, T.; Passos, J.F. Quantitative assessment of markers for cell senescence. Exp. Gerontol. 2010, 45, 772–778. [Google Scholar] [CrossRef]
- Kojima, H.; Kunimoto, H.; Inoue, T.; Nakajima, K. The STAT3-IGFBP5 axis is critical for IL-6/gp130-induced premature senescence in human fibroblasts. Cell Cycle 2012, 11, 730–739. [Google Scholar] [CrossRef]
- Romano, M.; Sironi, M.; Toniatti, C.; Polentarutti, N.; Fruscella, P.; Ghezzi, P.; Faggioni, R.; Luini, W.; Van Hinsbergh, V.; Sozzani, S.; et al. Role of IL-6 and Its Soluble Receptor in Induction of Chemokines and Leukocyte Recruitment. Immunity 1997, 6, 315–325. [Google Scholar] [CrossRef] [Green Version]
- Rose-John, S. IL-6 trans-signaling via the soluble IL-6 receptor: Importance for the pro-inflammatory activities of IL-6. Int. J. Biol. Sci. 2012, 8, 1237–1247. [Google Scholar] [CrossRef]
- Mohamed-Ali, V.; Pinkney, J.H.; Coppack, S.W. Adipose tissue as an endocrine and paracrine organ. Int. J. Obes. Relat. Metab. Disord. 1998, 22, 1145–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coppack, S.W. Pro-inflammatory cytokines and adipose tissue. Proc. Nutr. Soc. 2001, 60, 349–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kretschmer, T.; Schulze-Edinghausen, M.; Turnwald, E.-M.; Janoschek, R.; Bae-Gartz, I.; Zentis, P.; Handwerk, M.; Wohlfarth, M.; Schauss, A.; Hucklenbruch-Rother, E.; et al. Effect of Maternal Obesity in Mice on IL-6 Levels and Placental Endothelial Cell Homeostasis. Nutrients 2020, 12, 296. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12020296
Kretschmer T, Schulze-Edinghausen M, Turnwald E-M, Janoschek R, Bae-Gartz I, Zentis P, Handwerk M, Wohlfarth M, Schauss A, Hucklenbruch-Rother E, et al. Effect of Maternal Obesity in Mice on IL-6 Levels and Placental Endothelial Cell Homeostasis. Nutrients. 2020; 12(2):296. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12020296
Chicago/Turabian StyleKretschmer, Tobias, Merle Schulze-Edinghausen, Eva-Maria Turnwald, Ruth Janoschek, Inga Bae-Gartz, Peter Zentis, Marion Handwerk, Maria Wohlfarth, Astrid Schauss, Eva Hucklenbruch-Rother, and et al. 2020. "Effect of Maternal Obesity in Mice on IL-6 Levels and Placental Endothelial Cell Homeostasis" Nutrients 12, no. 2: 296. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12020296