Anti-Obesity Effects of Grateloupia elliptica, a Red Seaweed, in Mice with High-Fat Diet-Induced Obesity via Suppression of Adipogenic Factors in White Adipose Tissue and Increased Thermogenic Factors in Brown Adipose Tissue


Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. G. elliptica Ethanol Extract (GEE)
2.3. Cell Culture and Differentiation
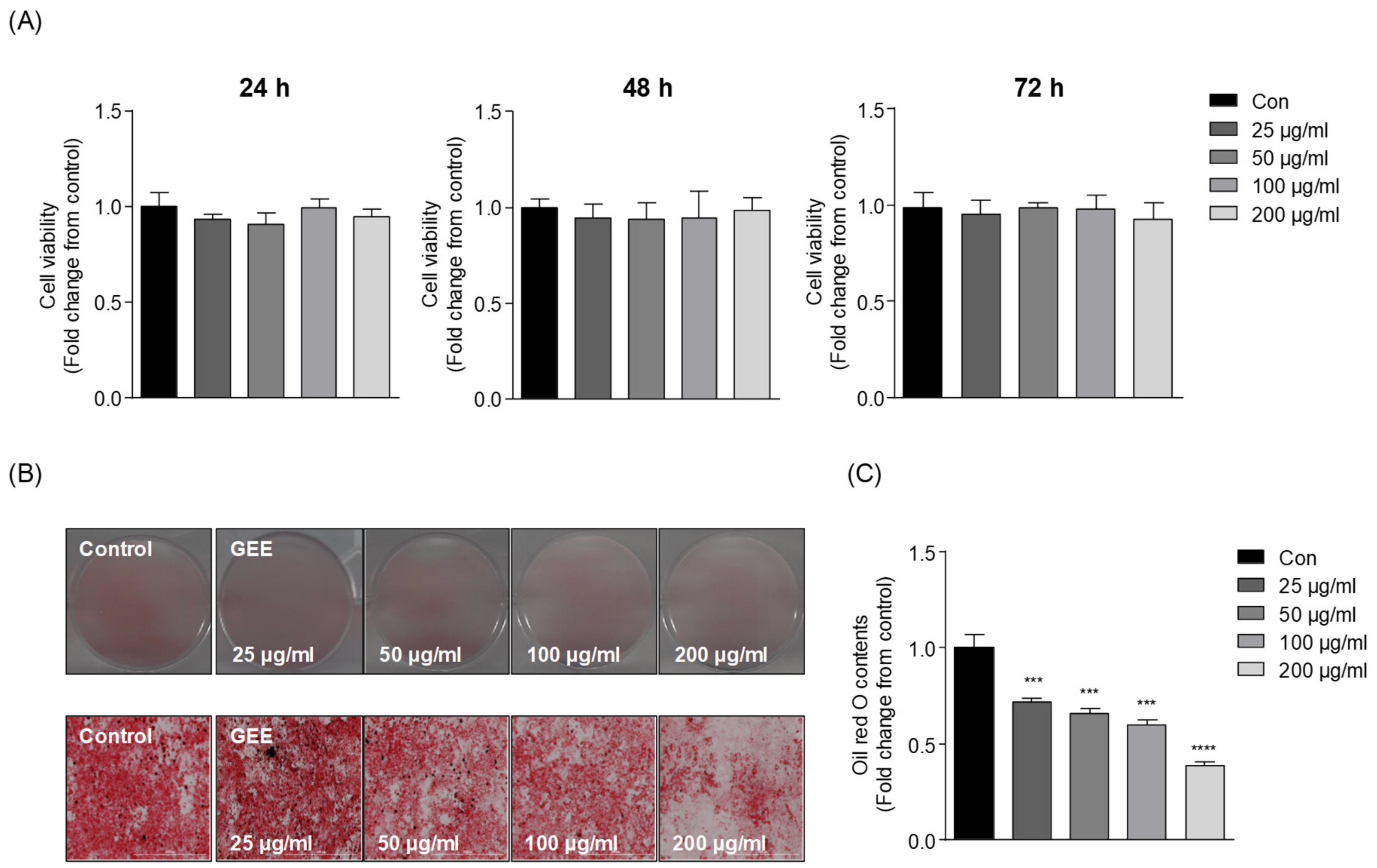
2.4. Cell Viability Assay
2.5. Oil Red O Staining
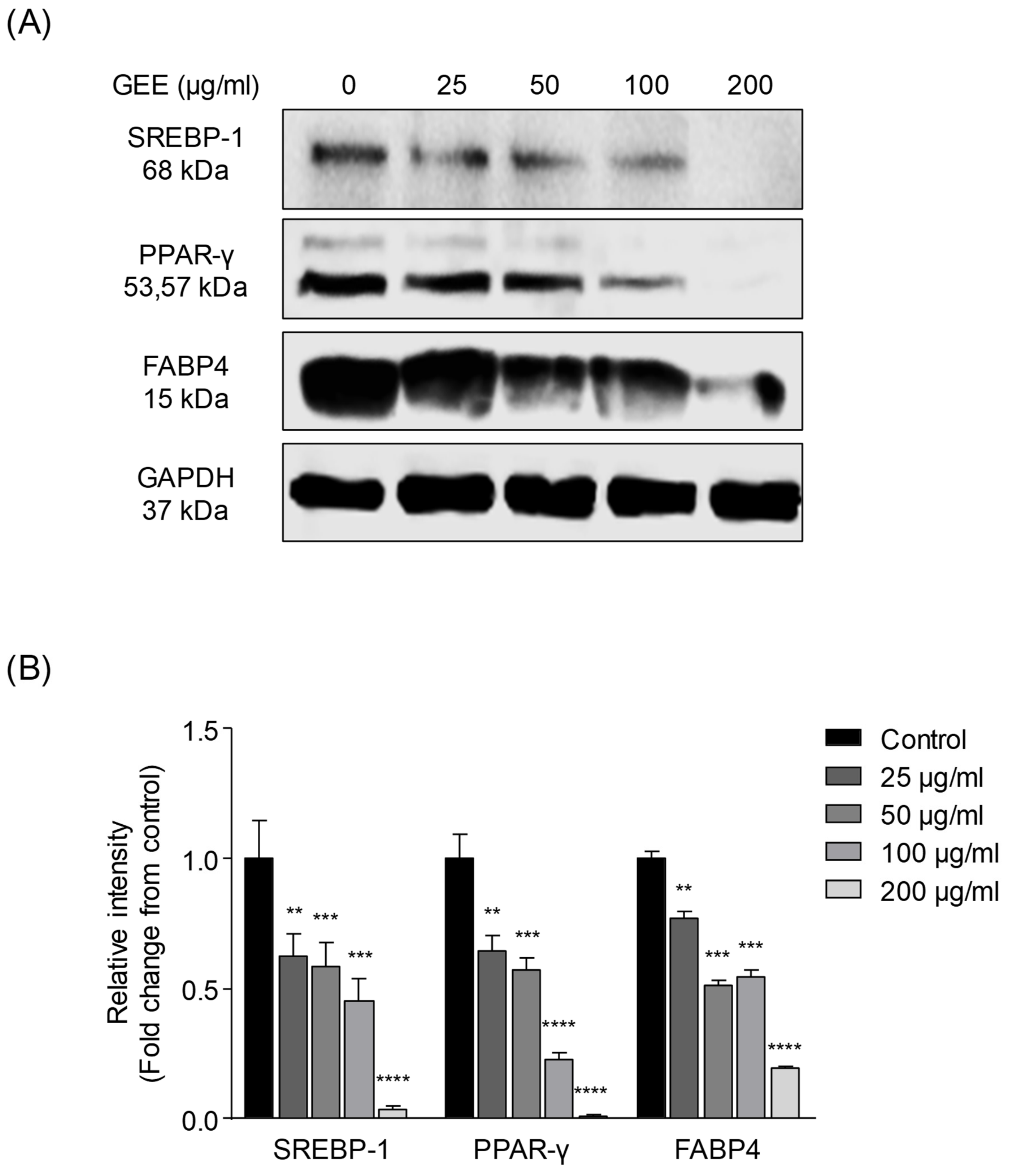
2.6. Western Blotting Analysis
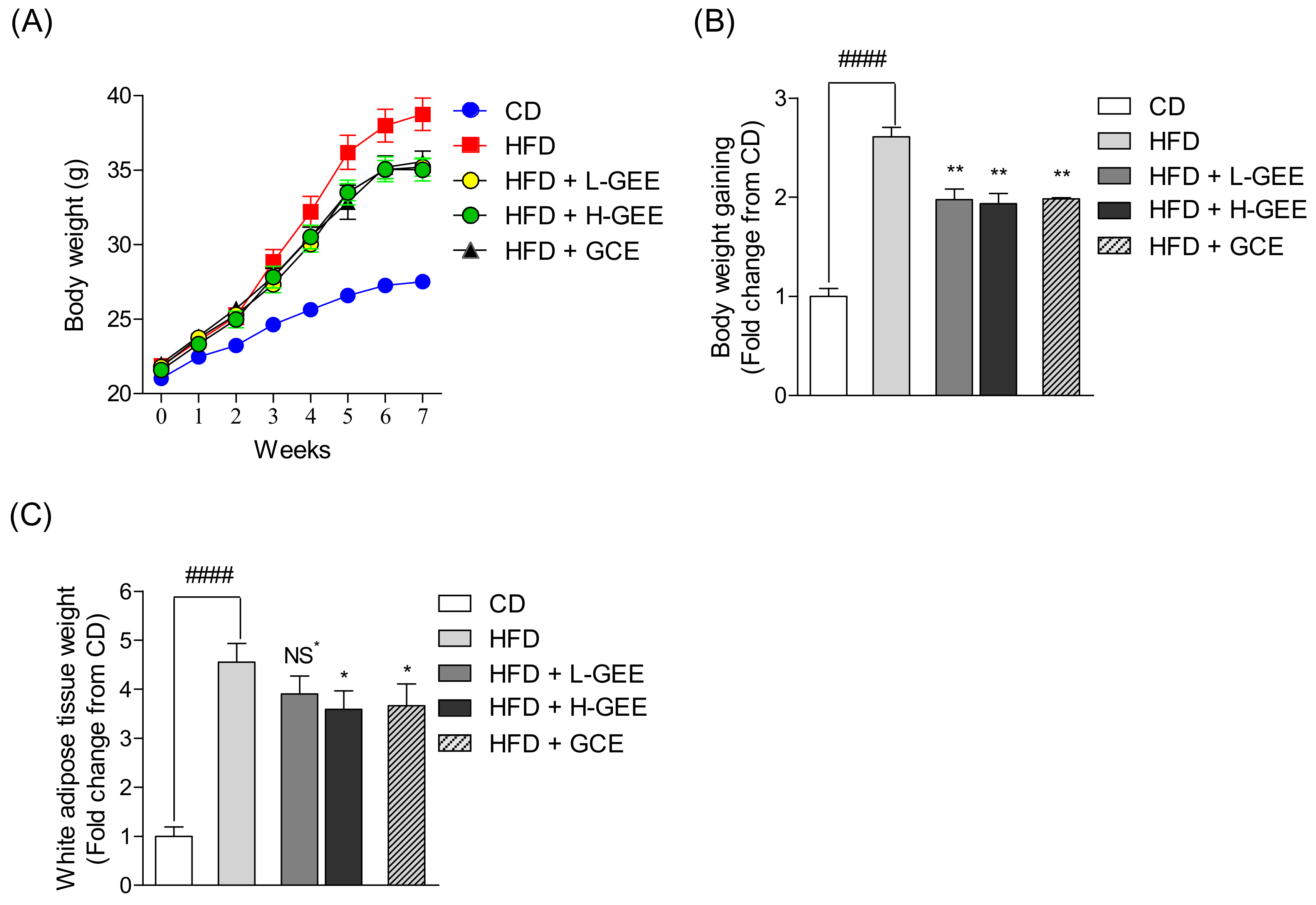
2.7. Animals
2.8. Realtime polymerase reaction (RT-PCR) Analysis of messenger RNA (mRNA) Expression of Adipogenic Genes
2.9. Serum Analysis
2.10. Histological Analysis
2.11. Statistical Analysis
3. Results
3.1. Effect of GEE on Adipocyte Differentiation and Lipid Accumulation in 3T3-L1 Cells
3.2. In Vitro Expression of Adipogenic and Lipogenic Proteins after GEE Treatment
3.3. Effects of GEE on Body and Adipose Tissue Weights in Mice with HFD-Induced Obesity
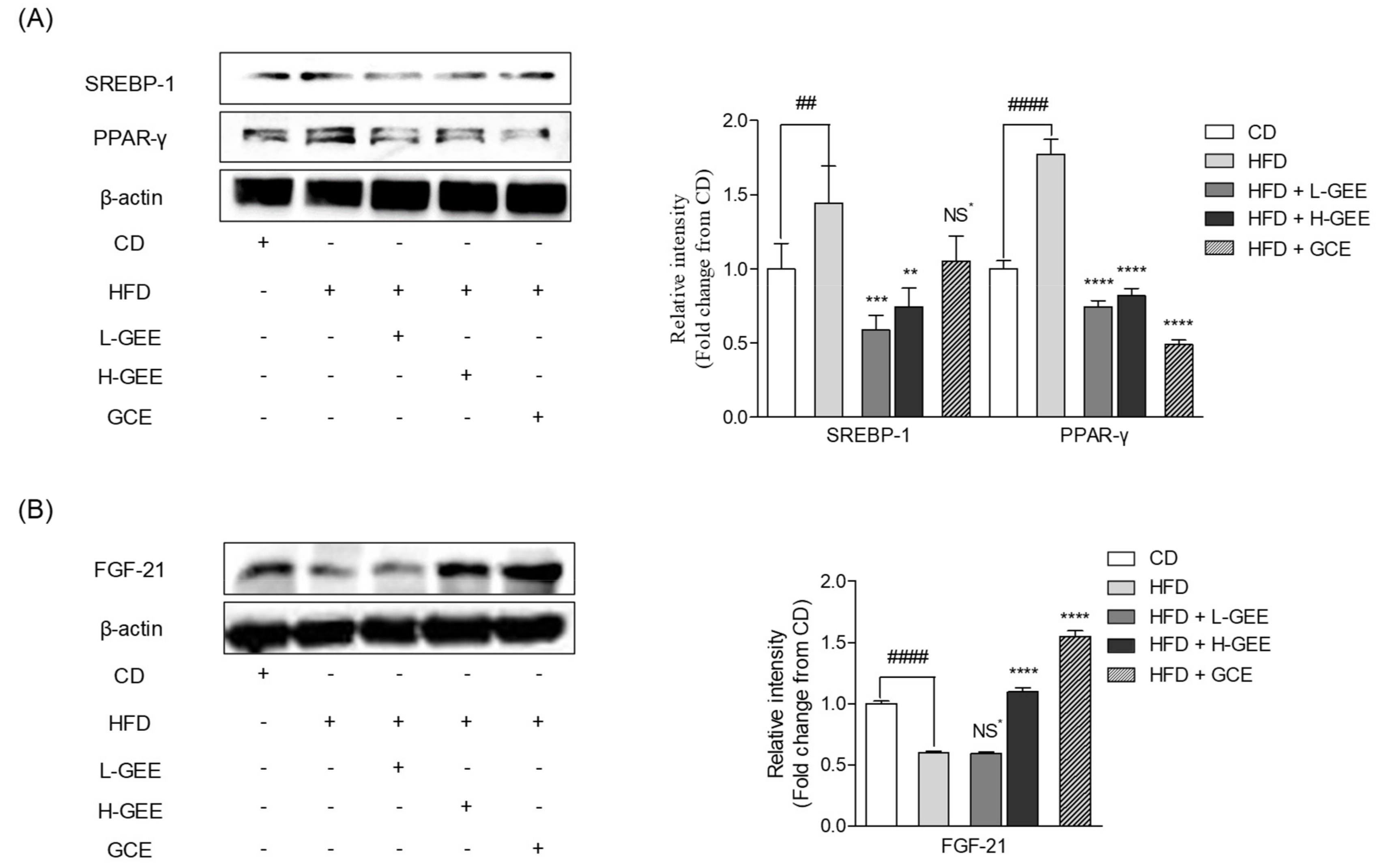
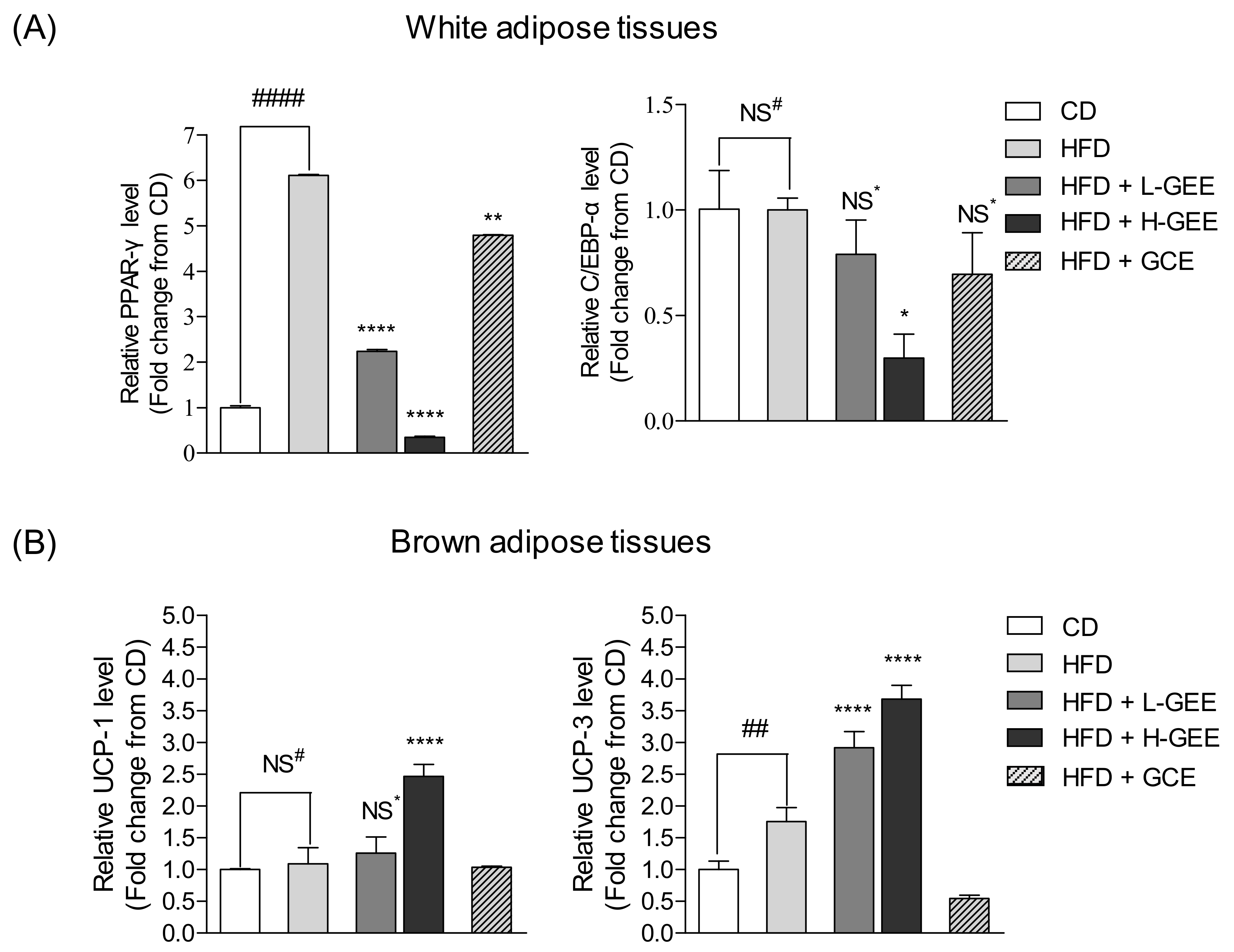
3.4. Potential Action of GEE on White Adipose Tissue and the Adipogenic Proteins
3.5. Effect of GEE on the Expression of the Metabolic Regulator Protein FGF-21 in White Adipose Tissue
3.6. Inhibitory Effect of GEE on Expression of Adipogenic Genes in White Adipose Tissue
3.7. Effect of GEE on Thermogenic Gene Expression in Brown Adipose Tissue
3.8. Effect of GEE on Mouse Blood Serum Biochemical Indices
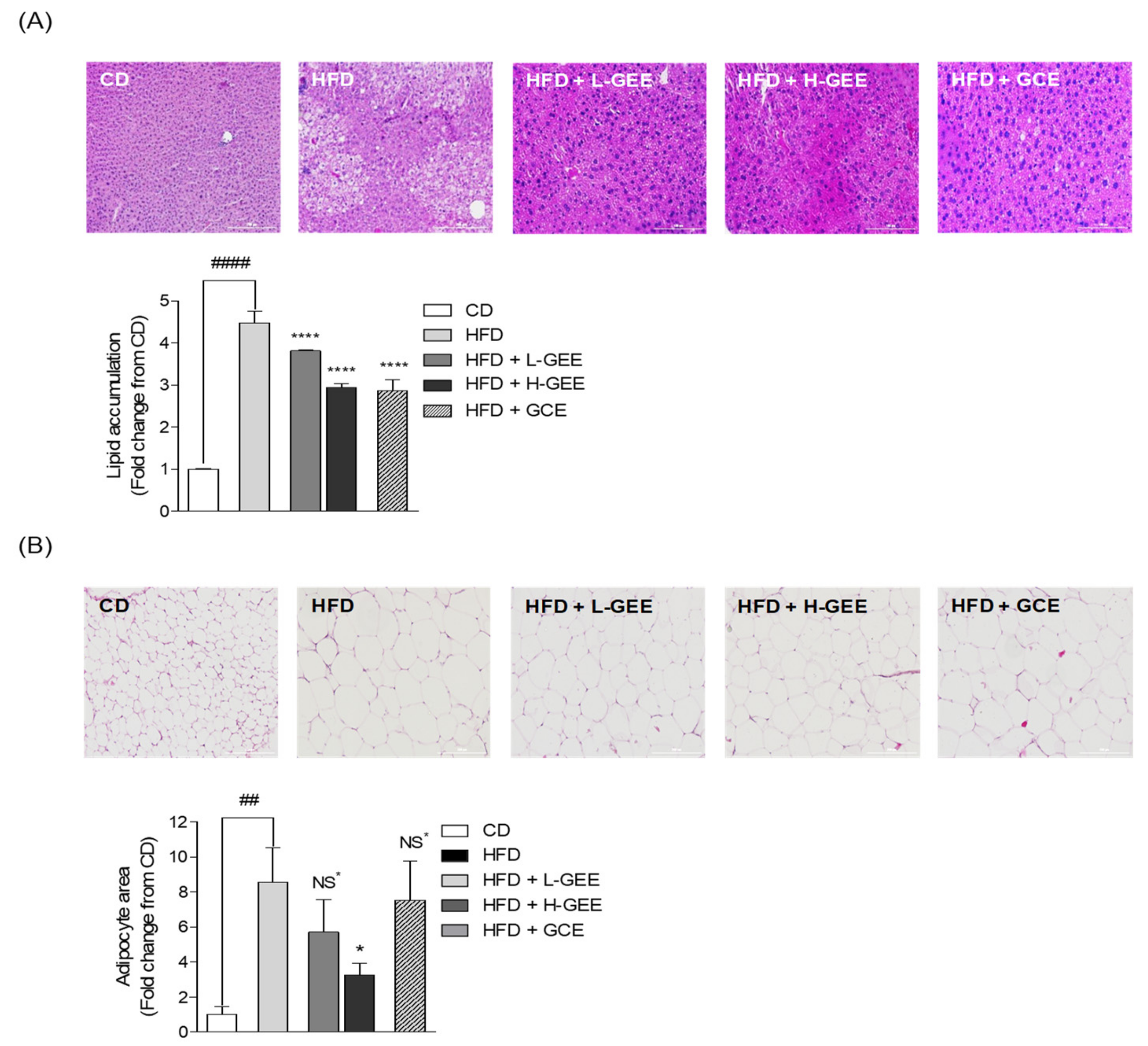
3.9. Histological Assay of Mouse Liver Tissue and White Adipose Tissue
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| BS | bovine serum |
| CD | chow diet |
| DMSO | dimethyl sulfoxide |
| DMEM | Dulbecco’s modified Eagle’s medium |
| FABP4 | fatty acid-binding protein 4 |
| FBS | fetal bovine serum |
| GCE | garcinia cambogia extract |
| GAPDH | glyceraldehyde 3-phosphate dehydrogenase |
| GEE | Grateloupia elliptica ethanol extract |
| H&E | hematoxylin and eosin |
| HFD | high-fat diet |
| ORO | oil red o |
| PPAR-γ | peroxisome proliferator-activated receptor gamma |
| SREBP-1 | sterol regulatory element-binding protein 1 |
| TC | total cholesterol |
| TG | triglyceride |
References
- Agovino, M.; Crociata, A.; Sacco, P.L. Proximity effects in obesity rates in the US: A Spatial Markov Chains approach. Soc. Sci. Med. 2019, 220, 301–311. [Google Scholar] [CrossRef] [PubMed]
- Hardy, L.L.; Jin, K.; Mihrshahi, S.; Ding, D. Trends in overweight, obesity, and waist-to-height ratio among Australian children from linguistically diverse backgrounds, 1997 to 2015. Int. J. Obes. 2019, 43, 116–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, N.; Tao, K.; Wang, G.; Xia, Z. Global obesity research trends during 1999 to 2017: A bibliometric analysis. Medicine 2019, 98, e14132. [Google Scholar] [CrossRef] [PubMed]
- Ng, M.; Fleming, T.; Robinson, M.; Thomson, B.; Graetz, N.; Margono, C.; Mullany, E.C.; Biryukov, S.; Abbafati, C.; Abera, S.F.; et al. Global, regional, and national prevalence of overweight and obesity in children and adults during 1980–2013: A systematic analysis for the Global Burden of Disease Study 2013. Lancet 2014, 384, 766–781. [Google Scholar] [CrossRef] [Green Version]
- Alford, S.; Patel, D.; Perakakis, N.; Mantzoros, C. Obesity as a risk factor for Alzheimer’s disease: Weighing the evidence. Obes. Rev. 2018, 19, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Gui, G.; Ji, W.; Xue, Q.; Wang, C.; Li, H. The relationship between obesity subtypes based on BMI and cardio-cerebrovascular disease. Hypertens. Res. 2019, 1, 912–919. [Google Scholar] [CrossRef]
- Kim, N.H.; Jung, Y.S.; Park, J.H.; Park, D.I.; Sohn, C.I. Abdominal Obesity is More Predictive of Advanced Colorectal Neoplasia Risk Than Overall Obesity in Men: A Cross-sectional Study. J. Clin. Gastroenterol. 2019, 53, e284–e290. [Google Scholar] [CrossRef]
- Leggio, M.; Lombardi, M.; Caldarone, E.; Severi, P.; D’emidio, S.; Armeni, M.; Bravi, V.; Bendini, M.G.; Mazza, A. The relationship between obesity and hypertension: An updated comprehensive overview on vicious twins. Hypertens. Res. 2017, 40, 947–963. [Google Scholar] [CrossRef]
- Liu, P.-H.; Wu, K.; Ng, K.; Zauber, A.G.; Nguyen, L.H.; Song, M.; He, X.; Fuchs, C.S.; Ogino, S.; Willett, W.C.; et al. Association of obesity with risk of early-onset colorectal cancer among women. JAMA Oncol. 2019, 5, 37–44. [Google Scholar] [CrossRef] [Green Version]
- Nehus, E. Obesity and chronic kidney disease. Curr. Opin. Pediatrics 2018, 30, 241–246. [Google Scholar] [CrossRef]
- Bray, G.; Kim, K.; Wilding, J.; Federation, W.O. Obesity: A chronic relapsing progressive disease process. A position statement of the World Obesity Federation. Obes. Rev. 2017, 18, 715–723. [Google Scholar] [PubMed] [Green Version]
- Jung, R.T. Obesity as a disease. Br. Med Bull. 1997, 53, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Kahan, S.; Zvenyach, T. Obesity as a disease: Current policies and implications for the future. Curr. Obes. Rep. 2016, 5, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Donohoe, C.; Pidgeon, G.; Lysaght, J.; Reynolds, J. Obesity and gastrointestinal cancer. Br. J. Surg. 2010, 97, 628–642. [Google Scholar] [CrossRef]
- Frezza, E.E.; Wachtel, M.S.; Chiriva-Internati, M. Influence of obesity on the risk of developing colon cancer. Gut 2006, 55, 285–291. [Google Scholar] [CrossRef]
- Grundy, S.M. Obesity, metabolic syndrome, and cardiovascular disease. J. Clin. Endocrinol. Metab. 2004, 89, 2595–2600. [Google Scholar] [CrossRef]
- Khandekar, M.J.; Cohen, P.; Spiegelman, B.M. Molecular mechanisms of cancer development in obesity. Nat. Rev. Cancer 2011, 11, 886–895. [Google Scholar] [CrossRef]
- Van Gaal, L.F.; Mertens, I.L.; Christophe, E. Mechanisms linking obesity with cardiovascular disease. Nature 2006, 444, 875–880. [Google Scholar] [CrossRef]
- Padwal, R.S.; Majumdar, S.R. Drug treatments for obesity: Orlistat, sibutramine, and rimonabant. Lancet 2007, 369, 71–77. [Google Scholar] [CrossRef]
- Barbier, P.; Hadvary, P.; Lengsfeld, H. Method of Reducing Gastrointestinal Side Effects Associated with Orlistat Treatment. Patent No. 6,756,364, 29 June 2004. [Google Scholar]
- Cavaliere, H.; Floriano, I.; Medeiros-Neto, G. Gastrointestinal side effects of orlistat may be prevented by concomitant prescription of natural fibers (psyllium mucilloid). Int. J. Obes. 2001, 25, 1095–1099. [Google Scholar] [CrossRef] [Green Version]
- Filippatos, T.D.; Derdemezis, C.S.; Gazi, I.F.; Nakou, E.S.; Mikhailidis, D.P.; Elisaf, M.S. Orlistat-associated adverse effects and drug interactions. Drug Saf. 2008, 31, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Hanl, K.; Kimura, Y.; Okuda, H. Anti-obesity effects of natural products. In Studies in Natural Products Chemistry; Elsevier: Amsterdam, The Netherlands, 2005; pp. 79–110. [Google Scholar]
- Kang, M.-C.; Ding, Y.; Kim, H.-S.; Jeon, Y.-J.; Lee, S.-H. Inhibition of Adipogenesis by Diphlorethohydroxycarmalol (DPHC) through AMPK activation in adipocytes. Mar. Drugs 2019, 17, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, M.-C.; Kang, N.; Kim, S.-Y.; Lima, I.S.; Ko, S.-C.; Kim, Y.-T.; Kim, Y.B.; Jeung, H.D.; Choi, K.S.; Jeon, Y.J. Popular edible seaweed, Gelidium amansii prevents against diet-induced obesity. Food Chem. Toxicol. 2016, 90, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.-C.; Kang, N.; Ko, S.-C.; Kim, Y.-B.; Jeon, Y.-J. Anti-obesity effects of seaweeds of Jeju Island on the differentiation of 3T3-L1 preadipocytes and obese mice fed a high-fat diet. Food Chem. Toxicol. 2016, 90, 36–44. [Google Scholar] [CrossRef]
- Mohamed, G.A.; Ibrahim, S.R.; Elkhayat, E.S.; El Dine, R.S. Natural anti-obesity agents. Bull. Fac. Pharm. Cairo Univ. 2014, 52, 269–284. [Google Scholar] [CrossRef] [Green Version]
- Vermaak, I.; Viljoen, A.M.; Hamman, J.H. Natural products in anti-obesity therapy. Nat. Prod. Rep. 2011, 28, 1493–1533. [Google Scholar] [CrossRef]
- Fernando, I.S.; Sanjeewa, K.A.; Ann, Y.-S.; Ko, C.-I.; Lee, S.-H.; Lee, W.W.; Jeon, Y.J. Apoptotic and antiproliferative effects of Stigmast-5-en-3-ol from Dendronephthya gigantea on human leukemia HL-60 and human breast cancer MCF-7 cells. Toxicol. Vitr. 2018, 52, 297–305. [Google Scholar] [CrossRef]
- Fernando, I.S.; Sanjeewa, K.A.; Kim, H.S.; Wang, L.; Lee, W.W.; Jeon, Y.J. Apoptotic and antiproliferative properties of 3β-hydroxy-Δ5-steroidal congeners from a partially purified column fraction of Dendronephthya gigantea against HL-60 and MCF-7 cancer cells. J. Appl. Toxicol. 2018, 38, 527–536. [Google Scholar] [CrossRef]
- Fernando, I.S.; Sanjeewa, K.A.; Samarakoon, K.W.; Lee, W.W.; Kim, H.-S.; Ranasinghe, P.; Gunasekara, U.K.D.S.S.; Jeon, Y.J. Antioxidant and anti-inflammatory functionality of ten Sri Lankan seaweed extracts obtained by carbohydrase assisted extraction. Food Sci. Biotechnol. 2018, 27, 1761–1769. [Google Scholar] [CrossRef]
- Kim, H.-S.; Je, J.-G.; Ryu, B.; Kang, N.; Fernando, I.S.; Jayawardena, T.U.; Sanjeewa, K.K.A.; Oh, J.-Y.; Lee, T.-G.; Jeon, Y.-J. Antioxidant and angiotensin-I converting enzyme inhibitory peptides from Hippocampus abdominalis. Eur. Food Res. Technol. 2019, 245, 479–487. [Google Scholar] [CrossRef]
- Sanjeewa, K.K.A.; Kang, N.; Ahn, G.; Jee, Y.; Kim, Y.-T.; Jeon, Y.-J. Bioactive potentials of sulfated polysaccharides isolated from brown seaweed Sargassum spp in related to human health applications: A review. Food Hydrocoll. 2018, 81, 200–208. [Google Scholar] [CrossRef]
- Wang, L.; Lee, W.; Oh, J.; Cui, Y.; Ryu, B.; Jeon, Y.-J. Protective Effect of Sulfated Polysaccharides from Celluclast-Assisted Extract of Hizikia fusiforme Against Ultraviolet B-Induced Skin Damage by Regulating NF-κB, AP-1, and MAPKs Signaling Pathways In Vitro in Human Dermal Fibroblasts. Mar. Drugs 2018, 16, 239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Lee, W.W.; Yang, H.-W.; Ryu, B.M.; Cui, Y.R.; Lee, S.-C.; Lee, T.G.; Jeon, Y.J. Protective Effect of Water Extract of Citrus Pomace against AAPH-Induced Oxidative Stress In Vitro in Vero Cells and In Vivo in Zebrafish. Prev. Nutr. Food Sci. 2018, 23, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Oh, J.Y.; Kim, H.S.; Lee, W.; Cui, Y.; Lee, H.G.; Kim, Y.T.; Ko, J.Y.; Jeon, Y.J. Protective effect of polysaccharides from Celluclast-assisted extract of Hizikia fusiforme against hydrogen peroxide-induced oxidative stress in vitro in Vero cells and in vivo in zebrafish. Int. J. Biol. Macromol. 2018, 112, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Park, Y.-J.; Jeon, Y.-J.; Ryu, B. Bioactivities of the edible brown seaweed, Undaria pinnatifida: A review. Aquaculture 2018, 495, 873–880. [Google Scholar] [CrossRef]
- Wan-Loy, C.; Siew-Moi, P. Marine algae as a potential source for anti-obesity agents. Mar. Drugs 2016, 14, 222. [Google Scholar] [CrossRef] [Green Version]
- Seo, M.-J.; Lee, O.-H.; Choi, H.-S.; Lee, B.-Y. Extract from edible red seaweed (Gelidium amansii) inhibits lipid accumulation and ROS production during differentiation in 3T3-L1 cells. Prev. Nutr. Food Sci. 2012, 17, 129–135. [Google Scholar] [CrossRef] [Green Version]
- Birari, R.; Javia, V.; Bhutani, K.K. Antiobesity and lipid lowering effects of Murraya koenigii (L.) Spreng leaves extracts and mahanimbine on high fat diet induced obese rats. Fitoterapia 2010, 81, 1129–1133. [Google Scholar]
- Ekanem, A.; Wang, M.; Simon, J.; Moreno, D. Antiobesity properties of two African plants (Afromomum meleguetta and Spilanthes acmella) by pancreatic lipase inhibition. Phytother. Res. Int. J. Devoted Pharmacol. Toxicol. Eval. Nat. Prod. Deriv. 2007, 21, 1253–1255. [Google Scholar]
- Gooda Sahib, N.; Saari, N.; Ismail, A.; Khatib, A.; Mahomoodally, F.; Abdul Hamid, A. Plants’ metabolites as potential antiobesity agents. Sci. World J. 2012, 2012, 436039. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.-S.; Kim, T.-W.; Kim, D.-J.; Hwang, H.-J.; Lee, H.-J.; Choe, M. Effects of natural plants supplementation on adipocyte size of the epididymal fat pads in rats. J. Korean Soc. Food Sci. Nutr. 2007, 36, 419–423. [Google Scholar] [CrossRef]
- Sergent, T.; Vanderstraeten, J.; Winand, J.; Beguin, P.; Schneider, Y.-J. Phenolic compounds and plant extracts as potential natural anti-obesity substances. Food Chem. 2012, 135, 68–73. [Google Scholar] [CrossRef]
- Sun, N.-N.; Wu, T.-Y.; Chau, C.-F. Natural dietary and herbal products in anti-obesity treatment. Molecules 2016, 21, 1351. [Google Scholar] [CrossRef] [PubMed]
- Bae, N.-Y.; Kim, M.-J.; Kim, K.; Ahn, N.-K.; Choi, Y.-U.; Park, J.-H.; Park, S.-H.; Ahn, D.-H. Anti-inflammatory effect of ethanol extract from Grateloupia elliptica Holmes on lipopolysaccharide-induced inflammatory responses in RAW 264.7 cells and mice ears. J. Korean Sci. Food Nutr. 2015, 44, 1128–1136. [Google Scholar] [CrossRef] [Green Version]
- Cho, M.; Park, G.-M.; Kim, S.-N.; Amna, T.; Lee, S.; Shin, W.-S. Glioblastoma-specific anticancer activity of pheophorbide a from the edible red seaweed Grateloupia elliptica. J. Microbiol. Biotechnol. 2014, 24, 346–353. [Google Scholar] [CrossRef] [Green Version]
- Heo, S.-J.; Cha, S.-H.; Lee, K.-W.; Jeon, Y.-J. Antioxidant activities of red algae from Jeju Island. Algae 2006, 21, 149–156. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; Nam, K.; Kurihara, H.; Kim, S. Potent α-glucosidase inhibitors purified from the red alga Grateloupia elliptica. Phytochemistry 2008, 69, 2820–2825. [Google Scholar] [CrossRef]
- Kim, J.B.; Spiegelman, B.M. ADD1/SREBP1 promotes adipocyte differentiation and gene expression linked to fatty acid metabolism. Genes Dev. 1996, 10, 1096–1107. [Google Scholar] [CrossRef] [Green Version]
- Lane, M.; Lin, F.; MacDougald, O.; Vasseur-Cognet, M. Control of adipocyte differentiation by CCAAT/enhancer binding protein alpha (C/EBP alpha). Int. J. Obes. Relat. Metab. Disord. J. Int. Assoc. Study obes. 1996, 20, S91–S96. [Google Scholar]
- Schadinger, S.E.; Bucher, N.L.; Schreiber, B.M.; Farmer, S.R. PPARγ2 regulates lipogenesis and lipid accumulation in steatotic hepatocytes. Am. J. Physiol.-Endocrinol. Metab. 2005, 288, E1195–E1205. [Google Scholar] [CrossRef]
- Spiegelman, B.; Puigserver, P.; Wu, Z. Regulation of adipogenesis and energy balance by PPARγ and PGC-1. Int. J. Obes. 2000, 24, S8–S10. [Google Scholar] [CrossRef] [Green Version]
- Vidal-Puig, A.; Jimenez-Liñan, M.; Lowell, B.B.; Hamann, A.; Hu, E.; Spiegelman, B.; Flier, J.S.; Moller, D.E. Regulation of PPAR gamma gene expression by nutrition and obesity in rodents. J. Clin. Investig. 1996, 97, 2553–2561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, A.; Wang, Y.; Xu, J.Y.; Stejskal, D.; Tam, S.; Zhang, J.; Wat, N.M.; Wong, W.K.; Lam, K.S. Adipocyte fatty acid–binding protein is a plasma biomarker closely associated with obesity and metabolic syndrome. Clin. Chem. 2006, 52, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Loftus, T.M.; Lane, M.D. Development, Modulating the transcriptional control of adipogenesis. Curr. Opin. Genet. Dev. 1997, 7, 603–608. [Google Scholar] [CrossRef]
- Rosen, E.D.; Walkey, C.J.; Puigserver, P.; Spiegelman, B.M. Development, Transcriptional regulation of adipogenesis. Genes Dev. 2000, 14, 1293–1307. [Google Scholar]
- Liang, Y.-C.; Yang, M.-T.; Lin, C.-J.; Chang, C.L.-T.; Yang, W.-C. Bidens pilosa and its active compound inhibit adipogenesis and lipid accumulation via down-modulation of the C/EBP and PPARγ pathways. Sci. Rep. 2016, 6, 24285. [Google Scholar] [CrossRef] [Green Version]
- Tang, Q.-Q.; Zhang, J.-W.; Lane, M.D. Sequential gene promoter interactions by C/EBPβ, C/EBPα, and PPARγ during adipogenesis. Biochem. Biophys. Res. Commun. 2004, 318, 213–218. [Google Scholar] [CrossRef]
- Jemai, R.; Drira, R.; Makni, M.; Fetoui, H.; Sakamoto, K.J.F.B. Colocynth (Citrullus colocynthis) seed extracts attenuate adipogenesis by down-regulating PPARγ/SREBP-1c and C/EBPα in 3T3-L1 cells. Food Biosci. 2020, 33, 100491. [Google Scholar] [CrossRef]
- Naowaboot, J.; Chung, C.H.; Pannangpetch, P.; Choi, R.; Kim, B.H.; Lee, M.Y.; Kukongviriyapan, U. Mulberry leaf extract increases adiponectin in murine 3T3-L1 adipocytes. Nutr Res. 2012, 32, 39–44. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Groups | ||||
|---|---|---|---|---|---|
| CD | HFD | HFD + L-GEE | HFD + H-GEE | HFD + GCE | |
| Triglyceride (mmol/μL) | 65.51 ± 2.03 | 110.78 ± 1.62 #### | 77.41 ± 0.14 ** | 58.22 ± 0.14 *** | 94.43 ± 0.14 ** |
| Total cholesterol (μg/μL) | 38.75 ± 0.00 | 50.17 ± 0.05 ## | 89.53 ± 0.17 ** | 29.33 ± 0.07 *** | 53.66 ± 0.08 NS* |
| Leptin (pg/mL) | 212.50 ± 2.50 | 4308.13 ± 59.37 #### | 3176.88 ± 44.38 *** | 1896.88 ± 1.87 *** | 4561.88 ± 4.38 ** |
| Insulin (ng/mL) | 1.48 ± 0.02 | 4.50 ± 0.18 #### | 4.10 ± 0.05 NS* | 3.97 ± 0.00 NS* | 6.23 ± 0.18 ** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.-G.; Lu, Y.A.; Li, X.; Hyun, J.-M.; Kim, H.-S.; Lee, J.J.; Kim, T.H.; Kim, H.M.; Kang, M.-C.; Jeon, Y.-J. Anti-Obesity Effects of Grateloupia elliptica, a Red Seaweed, in Mice with High-Fat Diet-Induced Obesity via Suppression of Adipogenic Factors in White Adipose Tissue and Increased Thermogenic Factors in Brown Adipose Tissue. Nutrients 2020, 12, 308. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12020308
Lee H-G, Lu YA, Li X, Hyun J-M, Kim H-S, Lee JJ, Kim TH, Kim HM, Kang M-C, Jeon Y-J. Anti-Obesity Effects of Grateloupia elliptica, a Red Seaweed, in Mice with High-Fat Diet-Induced Obesity via Suppression of Adipogenic Factors in White Adipose Tissue and Increased Thermogenic Factors in Brown Adipose Tissue. Nutrients. 2020; 12(2):308. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12020308
Chicago/Turabian StyleLee, Hyo-Geun, Yu An Lu, Xining Li, Ji-Min Hyun, Hyun-Soo Kim, Jeong Jun Lee, Tae Hee Kim, Hye Min Kim, Min-Cheol Kang, and You-Jin Jeon. 2020. "Anti-Obesity Effects of Grateloupia elliptica, a Red Seaweed, in Mice with High-Fat Diet-Induced Obesity via Suppression of Adipogenic Factors in White Adipose Tissue and Increased Thermogenic Factors in Brown Adipose Tissue" Nutrients 12, no. 2: 308. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12020308