Cholesterol-Lowering Action of a Novel Nutraceutical Combination in Uremic Rats: Insights into the Molecular Mechanism in a Hepatoma Cell Line

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and In Vivo Experimental Protocol
2.2. Reagents
2.3. Quantification of MK-7 Plasma Levels by LC-DAD-ESI-MS
2.4. Cell Cultures
2.5. In Vitro Calcification Assay
2.6. Cholesterol Biosynthesis Assay
2.7. Reversetranscription and Quantitative PCR (RT-qPCR)
2.8. Western Blot Analysis
2.9. ELISA
2.10. LDL-Isolation and Labeling
2.11. Fluorescent LDL Uptake Cell-Based Assay
2.12. Statistical Analysis
3. Results
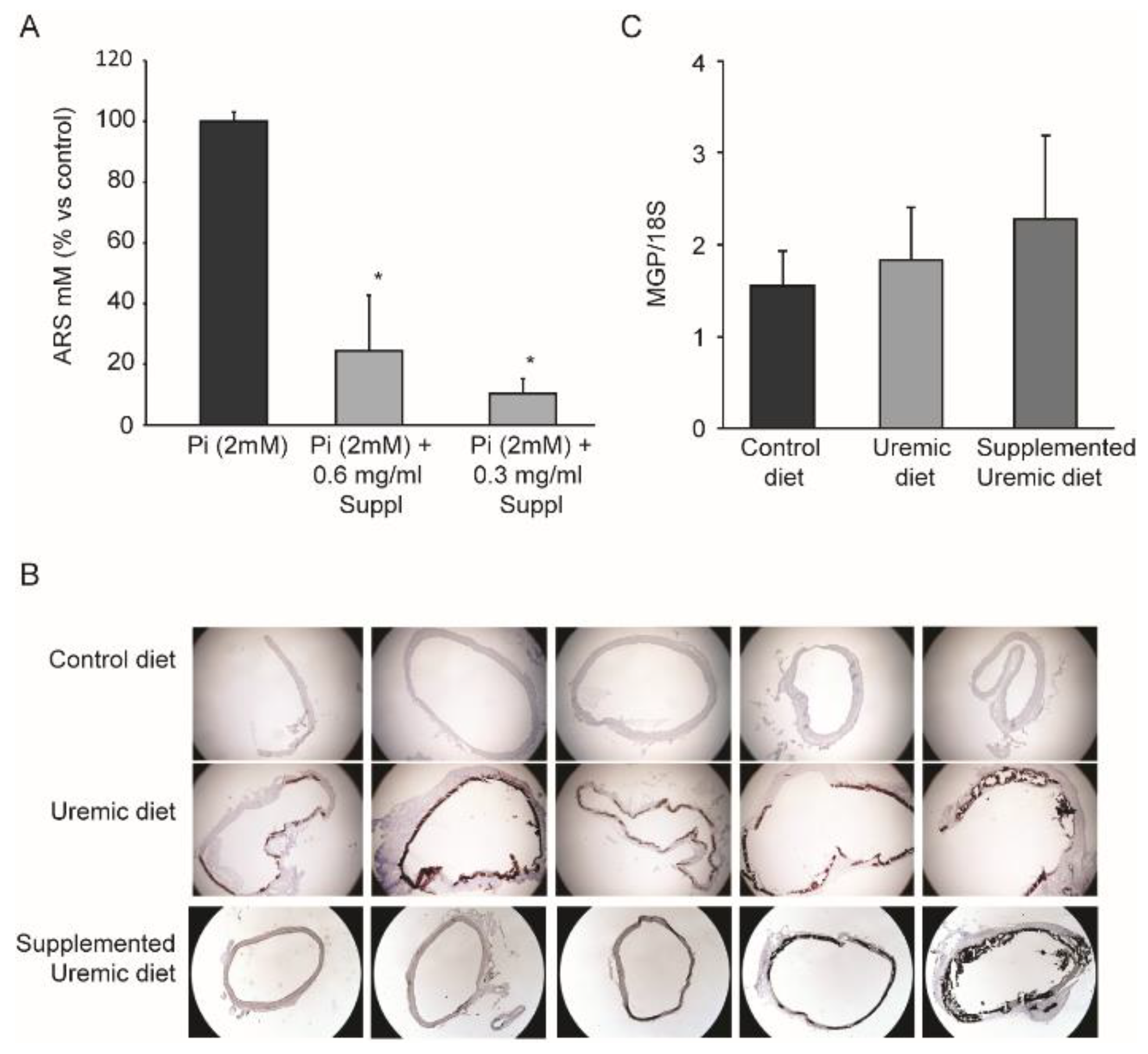
3.1. Effects of MK-7, MgCO3, and Sucrosomial® Iron on Vascular Calcification
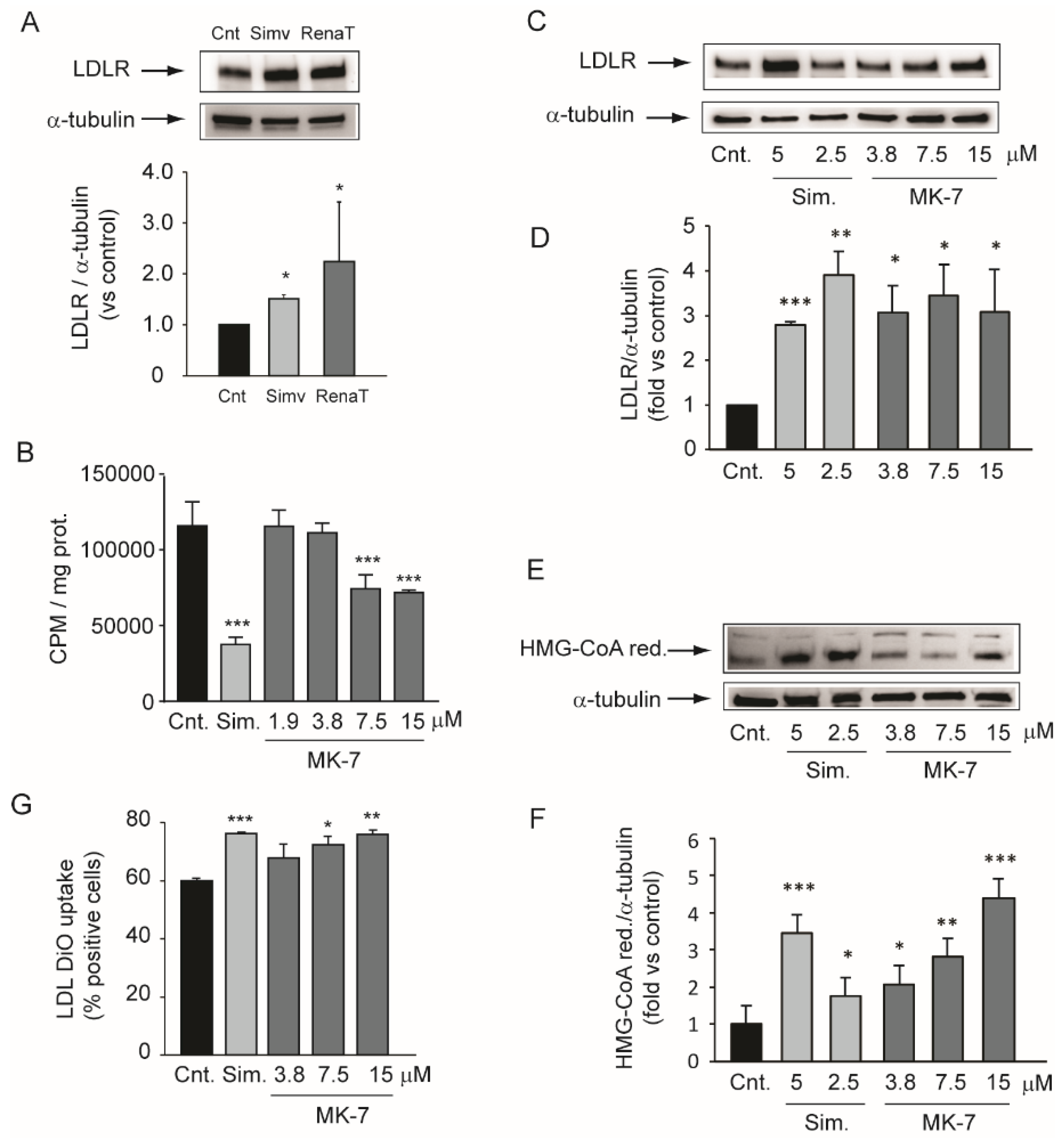
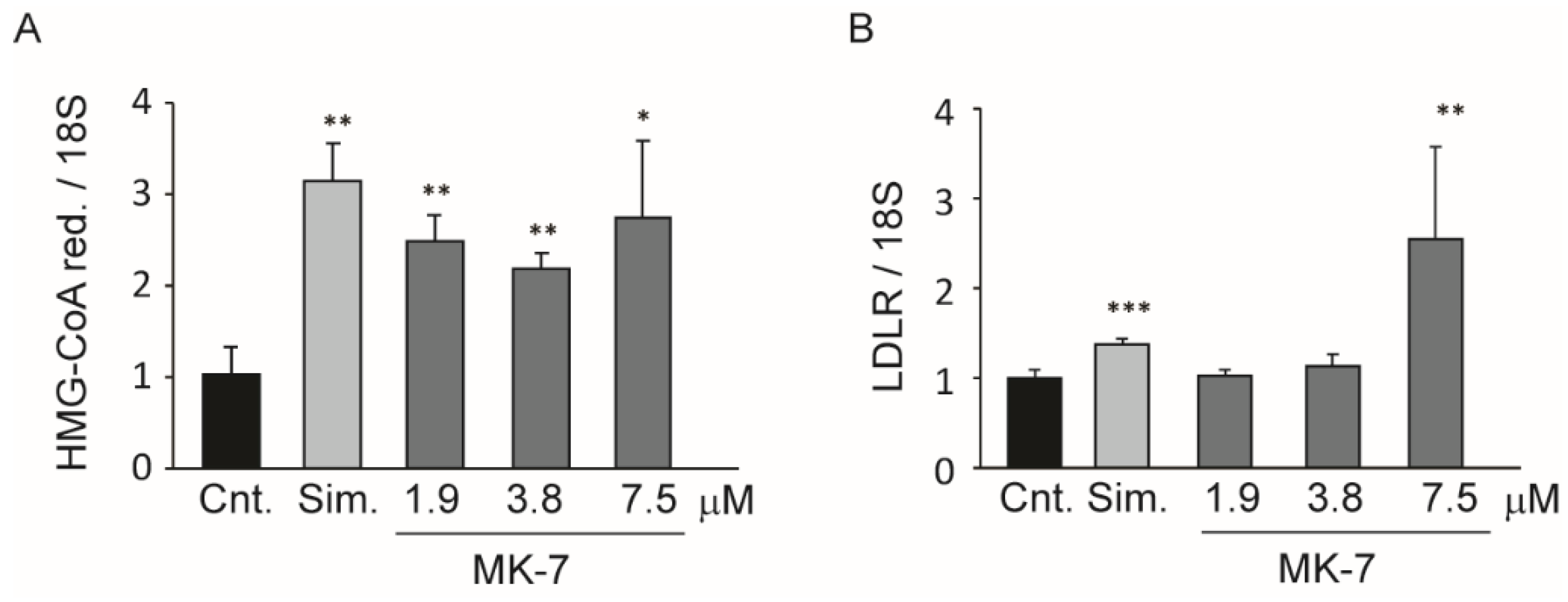
3.2. Inhibition of Cholesterol Biosynthesis in the Hepatic Cell Line
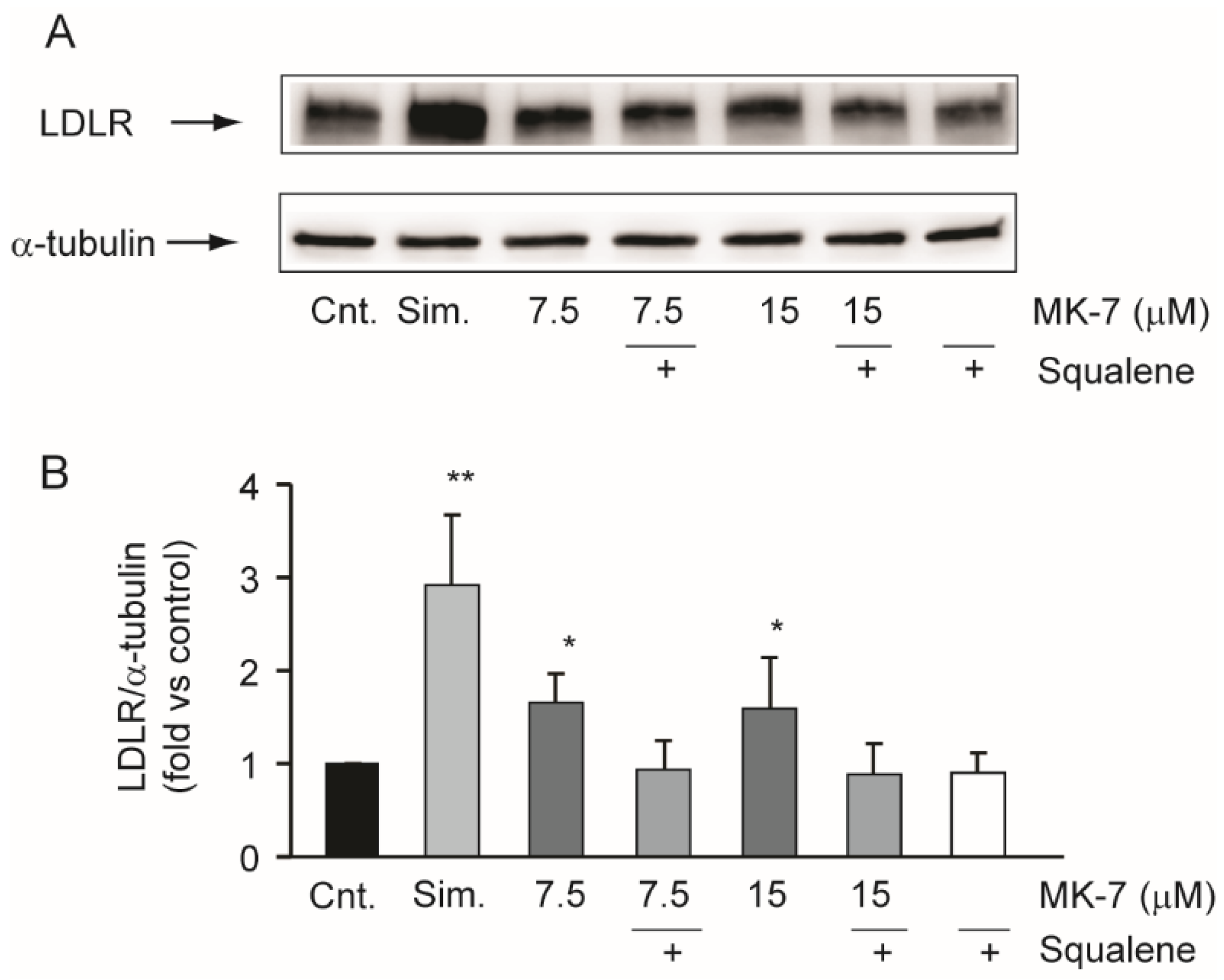
3.3. Squalene Counteracts the Effects of MK-7 on LDLR in the Hepatic Cell Line
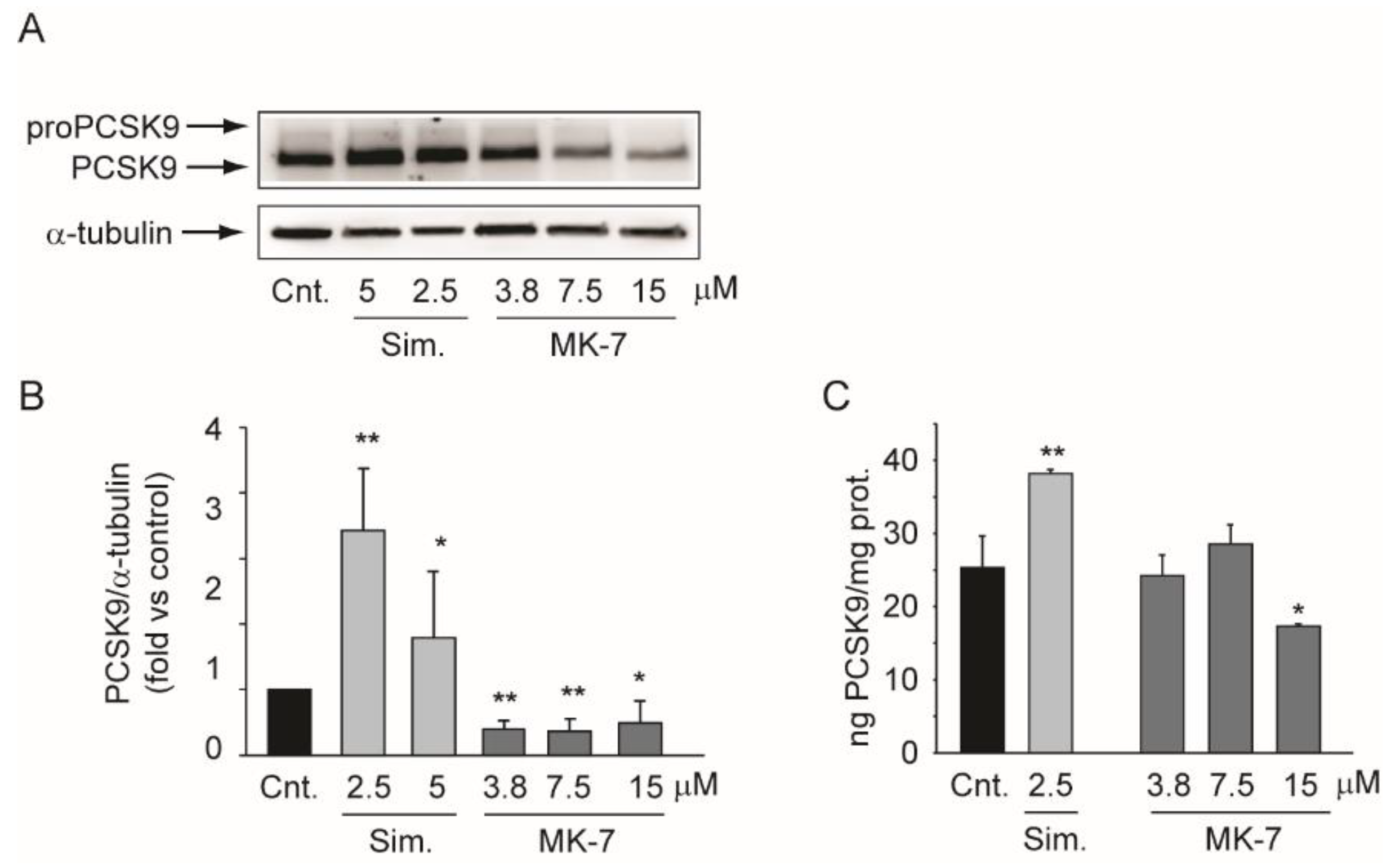
3.4. MK-7 Reduces the Expression of PCSK9 in the Hepatoma Cell Line
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kottgen, A.; Russell, S.D.; Loehr, L.R.; Crainiceanu, C.M.; Rosamond, W.D.; Chang, P.P.; Chambless, L.E.; Coresh, J. Reduced kidney function as a risk factor for incident heart failure: The atherosclerosis risk in communities (ARIC) study. J. Am. Soc. Nephrol. JASN 2007, 18, 1307–1315. [Google Scholar] [CrossRef] [PubMed]
- Abramson, J.L.; Jurkovitz, C.T.; Vaccarino, V.; Weintraub, W.S.; McClellan, W. Chronic kidney disease, anemia, and incident stroke in a middle-aged, community-based population: The ARIC Study. Kidney Int. 2003, 64, 610–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wattanakit, K.; Folsom, A.R.; Selvin, E.; Coresh, J.; Hirsch, A.T.; Weatherley, B.D. Kidney function and risk of peripheral arterial disease: Results from the Atherosclerosis Risk in Communities (ARIC) Study. J. Am. Soc. Nephrol. JASN 2007, 18, 629–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Astor, J. Re: The IAAP and the JAP: A brief history of their management of theoretical diversity. J. Anal. Psychol. 2006, 51, 620. [Google Scholar] [CrossRef] [PubMed]
- Alonso, A.; Lopez, F.L.; Matsushita, K.; Loehr, L.R.; Agarwal, S.K.; Chen, L.Y.; Soliman, E.Z.; Astor, B.C.; Coresh, J. Chronic kidney disease is associated with the incidence of atrial fibrillation: The Atherosclerosis Risk in Communities (ARIC) study. Circulation 2011, 123, 2946–2953. [Google Scholar] [CrossRef] [Green Version]
- Krane, V.; Wanner, C. Statins, inflammation and kidney disease. Nat. Rev. Nephrol. 2011, 7, 385–397. [Google Scholar] [CrossRef]
- Gansevoort, R.T.; Correa-Rotter, R.; Hemmelgarn, B.R.; Jafar, T.H.; Heerspink, H.J.; Mann, J.F.; Matsushita, K.; Wen, C.P. Chronic kidney disease and cardiovascular risk: Epidemiology, mechanisms, and prevention. Lancet 2013, 382, 339–352. [Google Scholar] [CrossRef]
- Duranton, F.; Laget, J.; Gayrard, N.; Saulnier-Blache, J.S.; Lundin, U.; Schanstra, J.P.; Mischak, H.; Weinberger, K.M.; Servel, M.F.; Argiles, A. The CKD plasma lipidome varies with disease severity and outcome. J. Clin. Lipidol. 2019, 13, 176–185. e178. [Google Scholar] [CrossRef] [Green Version]
- Okubo, K.; Ikewaki, K.; Sakai, S.; Tada, N.; Kawaguchi, Y.; Mochizuki, S. Abnormal HDL apolipoprotein A-I and A-II kinetics in hemodialysis patients: A stable isotope study. J. Am. Soc. Nephrol. JASN 2004, 15, 1008–1015. [Google Scholar] [CrossRef] [Green Version]
- Ruscica, M.; Ferri, N.; Fogacci, F.; Rosticci, M.; Botta, M.; Marchiano, S.; Magni, P.; D’Addato, S.; Giovannini, M.; Borghi, C.; et al. Circulating levels of proprotein convertase subtilisin/kexin type 9 and arterial stiffness in a large population sample: Data from The Brisighella Heart Study. J. Am. Heart Assoc. 2017, 6, e005764. [Google Scholar] [CrossRef] [Green Version]
- Kronenberg, F. Causes and consequences of lipoprotein(a) abnormalities in kidney disease. Clin. Exp. Nephrol. 2014, 18, 234–237. [Google Scholar] [CrossRef] [PubMed]
- Ketteler, M.; Schlieper, G.; Floege, J. Calcification and cardiovascular health: New insights into an old phenomenon. Hypertension 2006, 47, 1027–1034. [Google Scholar] [CrossRef] [PubMed]
- Schiffrin, E.L.; Lipman, M.L.; Mann, J.F. Chronic kidney disease: Effects on the cardiovascular system. Circulation 2007, 116, 85–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, G.; Ducy, P.; McKee, M.D.; Pinero, G.J.; Loyer, E.; Behringer, R.R.; Karsenty, G. Spontaneous calcification of arteries and cartilage in mice lacking matrix GLA protein. Nature 1997, 386, 78–81. [Google Scholar] [CrossRef] [PubMed]
- Schurgers, L.J.; Uitto, J.; Reutelingsperger, C.P. Vitamin K-dependent carboxylation of matrix Gla-protein: A crucial switch to control ectopic mineralization. Trends Mol. Med. 2013, 19, 217–226. [Google Scholar] [CrossRef] [PubMed]
- van Ballegooijen, A.J.; Beulens, J.W. The Role of Vitamin K Status in Cardiovascular Health: Evidence from Observational and Clinical Studies. Curr. Nutr. Rep. 2017, 6, 197–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elder, S.J.; Haytowitz, D.B.; Howe, J.; Peterson, J.W.; Booth, S.L. Vitamin k contents of meat, dairy, and fast food in the u.s. Diet. J. Agric. Food Chem. 2006, 54, 463–467. [Google Scholar] [CrossRef]
- Sakano, T.; Nagaoka, T.; Morimoto, A.; Hirauchi, K. Measurement of K vitamins in human and animal feces by high-performance liquid chromatography with fluorometric detection. Chem. Pharm. Bull. 1986, 34, 4322–4326. [Google Scholar] [CrossRef] [Green Version]
- Ji, Y.; Li, X.; Tso, P. Intestinal Fatty acid Absorption. Immunol. Endocr. Metab. Agents Med. Chem. 2009, 9, 60–73. [Google Scholar] [CrossRef]
- Cranenburg, E.C.; Schurgers, L.J.; Uiterwijk, H.H.; Beulens, J.W.; Dalmeijer, G.W.; Westerhuis, R.; Magdeleyns, E.J.; Herfs, M.; Vermeer, C.; Laverman, G.D. Vitamin K intake and status are low in hemodialysis patients. Kidney Int. 2012, 82, 605–610. [Google Scholar] [CrossRef] [Green Version]
- Dalmeijer, G.W.; van der Schouw, Y.T.; Magdeleyns, E.J.; Vermeer, C.; Verschuren, W.M.; Boer, J.M.; Beulens, J.W. Matrix Gla protein species and risk of cardiovascular events in type 2 diabetic patients. Diabetes Care 2013, 36, 3766–3771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schurgers, L.J.; Barreto, D.V.; Barreto, F.C.; Liabeuf, S.; Renard, C.; Magdeleyns, E.J.; Vermeer, C.; Choukroun, G.; Massy, Z.A. The circulating inactive form of matrix gla protein is a surrogate marker for vascular calcification in chronic kidney disease: A preliminary report. Clin. J. Am. Soc. Nephrol. CJASN 2010, 5, 568–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlieper, G.; Westenfeld, R.; Kruger, T.; Cranenburg, E.C.; Magdeleyns, E.J.; Brandenburg, V.M.; Djuric, Z.; Damjanovic, T.; Ketteler, M.; Vermeer, C.; et al. Circulating nonphosphorylated carboxylated matrix gla protein predicts survival in ESRD. J. Am. Soc. Nephrol. JASN 2011, 22, 387–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cozzolino, M.; Mangano, M.; Galassi, A.; Ciceri, P.; Messa, P.; Nigwekar, S. Vitamin K in Chronic Kidney Disease. Nutrients 2019, 11. [Google Scholar] [CrossRef] [Green Version]
- Scheiber, D.; Veulemans, V.; Horn, P.; Chatrou, M.L.; Potthoff, S.A.; Kelm, M.; Schurgers, L.J.; Westenfeld, R. High-Dose Menaquinone-7 Supplementation Reduces Cardiovascular Calcification in a Murine Model of Extraosseous Calcification. Nutrients 2015, 7, 6991–7011. [Google Scholar] [CrossRef] [Green Version]
- Zanotti, I.; Poti, F.; Favari, E.; Steffensen, K.R.; Gustafsson, J.A.; Bernini, F. Pitavastatin effect on ATP binding cassette A1-mediated lipid efflux from macrophages: Evidence for liver X receptor (LXR)-dependent and LXR-independent mechanisms of activation by cAMP. J. Pharmacol. Exp. Ther. 2006, 317, 395–401. [Google Scholar] [CrossRef]
- Adorni, M.P.; Cipollari, E.; Favari, E.; Zanotti, I.; Zimetti, F.; Corsini, A.; Ricci, C.; Bernini, F.; Ferri, N. Inhibitory effect of PCSK9 on Abca1 protein expression and cholesterol efflux in macrophages. Atherosclerosis 2016, 256, 1–6. [Google Scholar] [CrossRef]
- Raines, E.W.; Garton, K.J.; Ferri, N. Beyond the endothelium: NF-kappaB regulation of smooth muscle function. Circ. Res. 2004, 94, 706–708. [Google Scholar] [CrossRef] [Green Version]
- Morton, R.E.; Evans, T.A. Modification of the bicinchoninic acid protein assay to eliminate lipid interference in determining lipoprotein protein content. Anal. Biochem. 1992, 204, 332–334. [Google Scholar] [CrossRef]
- Ferri, N.; Tibolla, G.; Pirillo, A.; Cipollone, F.; Mezzetti, A.; Pacia, S.; Corsini, A.; Catapano, A.L. Proprotein convertase subtilisin kexin type 9 (PCSK9) secreted by cultured smooth muscle cells reduces macrophages LDLR levels. Atherosclerosis 2012, 220, 381–386. [Google Scholar] [CrossRef]
- Pirillo, A.; Reduzzi, A.; Ferri, N.; Kuhn, H.; Corsini, A.; Catapano, A.L. Upregulation of lectin-like oxidized low-density lipoprotein receptor-1 (LOX-1) by 15-lipoxygenase-modified LDL in endothelial cells. Atherosclerosis 2011, 214, 331–337. [Google Scholar] [CrossRef]
- Shuvy, M.; Nyska, A.; Beeri, R.; Abedat, S.; Gal-Moscovici, A.; Rajamannan, N.M.; Lotan, C. Histopathology and apoptosis in an animal model of reversible renal injury. Exp. Toxicol. Pathol.: Off. J. Ges. Fur Toxikol. Pathol. 2011, 63, 303–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokozawa, T.; Zheng, P.D.; Oura, H.; Koizumi, F. Animal model of adenine-induced chronic renal failure in rats. Nephron 1986, 44, 230–234. [Google Scholar] [CrossRef] [PubMed]
- Grunler, J.; Ericsson, J.; Dallner, G. Branch-point reactions in the biosynthesis of cholesterol, dolichol, ubiquinone and prenylated proteins. Biochim. Biophys. Acta 1994, 1212, 259–277. [Google Scholar] [CrossRef]
- Ferri, N.; Corsini, A.; Macchi, C.; Magni, P.; Ruscica, M. Proprotein convertase subtilisin kexin type 9 and high-density lipoprotein metabolism: Experimental animal models and clinical evidence. Transl. Res. J. Lab. Clin. Med. 2016, 173, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Dubuc, G.; Chamberland, A.; Wassef, H.; Davignon, J.; Seidah, N.G.; Bernier, L.; Prat, A. Statins upregulate PCSK9, the gene encoding the proprotein convertase neural apoptosis-regulated convertase-1 implicated in familial hypercholesterolemia. Arter. Thromb. Vasc. Biol. 2004, 24, 1454–1459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferri, N.; Marchiano, S.; Lupo, M.G.; Trenti, A.; Biondo, G.; Castaldello, P.; Corsini, A. Geranylgeraniol prevents the simvastatin-induced PCSK9 expression: Role of the small G protein Rac1. Pharmacol. Res. 2017, 122, 96–104. [Google Scholar] [CrossRef]
- Schlieper, G.; Schurgers, L.; Brandenburg, V.; Reutelingsperger, C.; Floege, J. Vascular calcification in chronic kidney disease: An update. Nephrol. Dial. Transplant. Off. Publ. Eur. Dial. Transpl. Assoc.-Eur. Ren. Assoc. 2016, 31, 31–39. [Google Scholar] [CrossRef] [Green Version]
- Ruderman, I.; Holt, S.G.; Hewitson, T.D.; Smith, E.R.; Toussaint, N.D. Current and potential therapeutic strategies for the management of vascular calcification in patients with chronic kidney disease including those on dialysis. Semin. Dial. 2018, 31, 487–499. [Google Scholar] [CrossRef]
- Giachelli, C.M. Vascular calcification: In vitro evidence for the role of inorganic phosphate. J. Am. Soc. Nephrol. JASN 2003, 14, S300–S304. [Google Scholar] [CrossRef] [Green Version]
- Ministero della Salute. Apporti giornalieri di vitamine e minerali ammessi negli integratori alimentari. In Direzione generale per l’igiene e la sicurezza degli alimenti e la nutrizione – Ufficio 4. Available online: http://www.salute.gov.it/imgs/C_17_pagineAree_1268_5_file.pdf (accessed on 6 January 2020).
- Nagata, Y.; Hidaka, Y.; Ishida, F.; Kamei, T. Effect of simvastatin (MK-733) on the regulation of cholesterol synthesis in Hep G2 cells. Biochem. Pharmacol. 1990, 40, 843–850. [Google Scholar] [CrossRef]
- Cameron, J.; Ranheim, T.; Kulseth, M.A.; Leren, T.P.; Berge, K.E. Berberine decreases PCSK9 expression in HepG2 cells. Atherosclerosis 2008, 201, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Lupo, M.G.; Macchi, C.; Marchiano, S.; Cristofani, R.; Greco, M.F.; Dall’Acqua, S.; Chen, H.; Sirtori, C.R.; Corsini, A.; Ruscica, M.; et al. Differential effects of red yeast rice, Berberis aristata and Morus alba extracts on PCSK9 and LDL uptake. Nutr. Metab. Cardiovasc. Dis. NMCD 2019. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.; Wei, J.; Abidi, P.; Lin, M.; Inaba, S.; Li, C.; Wang, Y.; Wang, Z.; Si, S.; Pan, H.; et al. Berberine is a novel cholesterol-lowering drug working through a unique mechanism distinct from statins. Nat. Med. 2004, 10, 1344–1351. [Google Scholar] [CrossRef] [PubMed]
- Stein, E.A.; Bays, H.; O’Brien, D.; Pedicano, J.; Piper, E.; Spezzi, A. Lapaquistat acetate: Development of a squalene synthase inhibitor for the treatment of hypercholesterolemia. Circulation 2011, 123, 1974–1985. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward (5′–3′) | Reverse (5′–3′) | Efficiency |
|---|---|---|---|
| HMGCR | CTTGTGTGTCCTTGGTATTAGAGCTT | GCTGAGCTGCCAAATTGGA | 125% |
| LDLR | TCTATGGAAGAACTGGCGGC | ACCATCTGTCTCGAGGGGTA | 93% |
| MGP | GCAGCCCTGTGCTATGAATCT | TTTAGCGTGCCATCTCTGCT | 91% |
| 18S | CGGCTACCACATCCACGGAA | CCTGAATTGTTATTTTTCGTCACTACC | 99% |
| Group | Phosphate (mmol/L) | Ca2+ Aorta (mg/g tissue) | Total Cholesterol (mg/dL) | Creatinine (µmol/L) | Iron (µmol/L) |
|---|---|---|---|---|---|
| Control diet | 2.8 ± 0.3 | 0.28 ± 0.12 | 63.0 ± 19.0 | 25.3 ± 2.8 | 27.5 |
| Uremic diet | 5.1 ± 1.5 * | 2.87 ± 1.76 *** | 95.9 ± 8.2 * | 206.4 ± 45.0 ** | 25.8 |
| Supplemented uremic diet | 4.5 ± 1.4 * | 3.28 ± 1.99 *** | 77.7 ± 116 *§ | 241.0 ± 71.7 ** | 32.0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lupo, M.G.; Biancorosso, N.; Brilli, E.; Tarantino, G.; Adorni, M.P.; Vivian, G.; Salvalaio, M.; Dall’Acqua, S.; Sut, S.; Neutel, C.; et al. Cholesterol-Lowering Action of a Novel Nutraceutical Combination in Uremic Rats: Insights into the Molecular Mechanism in a Hepatoma Cell Line. Nutrients 2020, 12, 436. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12020436
Lupo MG, Biancorosso N, Brilli E, Tarantino G, Adorni MP, Vivian G, Salvalaio M, Dall’Acqua S, Sut S, Neutel C, et al. Cholesterol-Lowering Action of a Novel Nutraceutical Combination in Uremic Rats: Insights into the Molecular Mechanism in a Hepatoma Cell Line. Nutrients. 2020; 12(2):436. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12020436
Chicago/Turabian StyleLupo, Maria Giovanna, Noemi Biancorosso, Elisa Brilli, Germano Tarantino, Maria Pia Adorni, Greta Vivian, Marika Salvalaio, Stefano Dall’Acqua, Stefania Sut, Cédric Neutel, and et al. 2020. "Cholesterol-Lowering Action of a Novel Nutraceutical Combination in Uremic Rats: Insights into the Molecular Mechanism in a Hepatoma Cell Line" Nutrients 12, no. 2: 436. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12020436