Effects of Coffee Intake on Dyslipidemia Risk According to Genetic Variants in the ADORA Gene Family among Korean Adults


Abstract
:1. Introduction
2. Materials and Methods
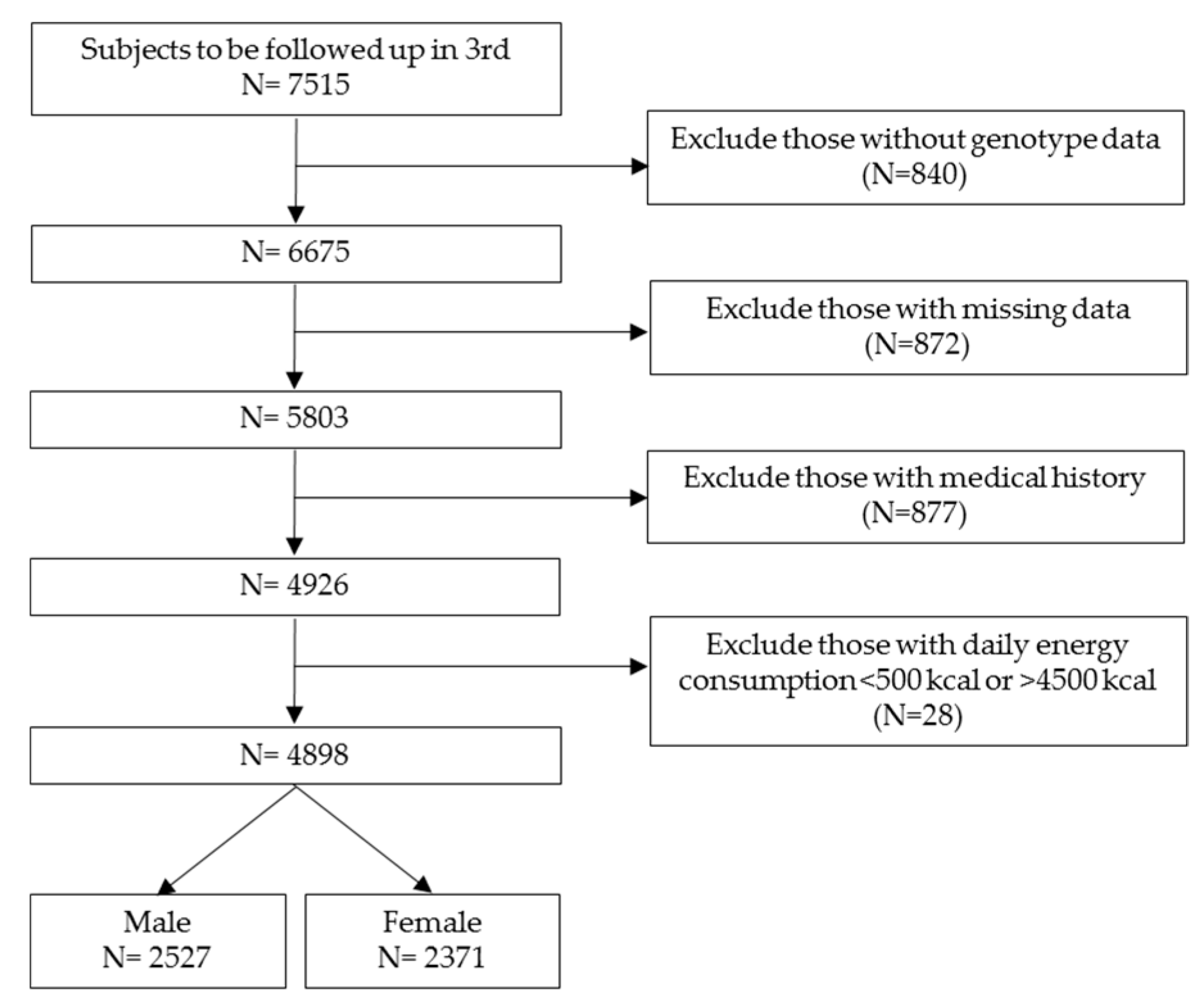
2.1. Study Population
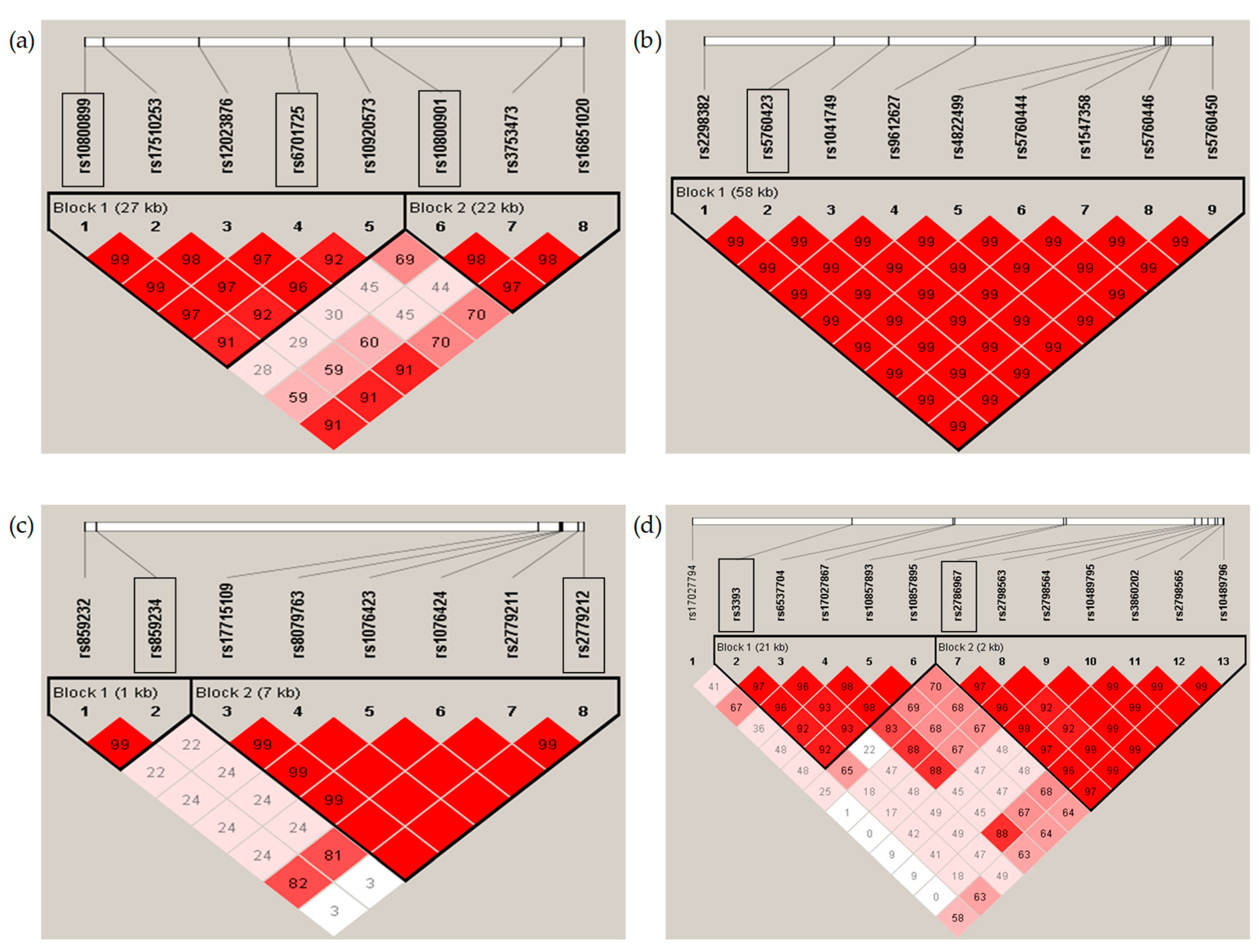
2.2. Genotyping and Analysis of Single Nucleotide Polymorphisms
2.3. Statistical Analysis
3. Results
3.1. Basic Characteristics Depending on Coffee Consumption
3.2. Association of Coffee Intake with the Risk of Dyslipidemia
3.3. Effects of Coffee Intake on the Risk of Dyslipidemia Depending on ADORA Gene Family
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cano-Marquina, A.; Tarin, J.J.; Cano, A. The impact of coffee on health. Maturitas 2013, 75, 7–21. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Ma, D.; Zhang, Y.; Liu, Z.; Wang, P. The effect of coffee consumption on serum lipids: A meta-analysis of randomized controlled trials. Eur. J. Clin. Nutr. 2012, 66, 872–877. [Google Scholar] [CrossRef] [PubMed]
- Poole, R.; Kennedy, O.J.; Roderick, P.; Fallowfield, J.A.; Hayes, P.C.; Parkes, J. Coffee consumption and health: Umbrella review of meta-analyses of multiple health outcomes. BMJ 2017, 359, j5024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, M.; Bhupathiraju, S.N.; Satija, A.; Van Dam, R.M.; Hu, F.B. Long-term coffee consumption and risk of cardiovascular disease: A systematic review and a dose–response meta-analysis of prospective cohort studies. Circulation 2014, 129, 643–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.N.; Ho, S.C.; Zhou, C.; Ling, W.H.; Chen, W.Q.; Wang, C.L.; Chen, Y.M. Coffee consumption and risk of coronary heart diseases: A meta-analysis of 21 prospective cohort studies. Int. J. Cardiol. 2009, 137, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Renda, G.; Zimarino, M.; Antonucci, I.; Tatasciore, A.; Ruggieri, B.; Bucciarelli, T.; Prontera, T.; Stuppia, L.; De Caterina, R. Genetic determinants of blood pressure responses to caffeine drinking. Am. J. Clin. Nutr. 2012, 95, 241–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fredholm, B.B.; AP, I.J.; Jacobson, K.A.; Linden, J.; Muller, C.E. International Union of Basic and Clinical Pharmacology. LXXXI. Nomenclature and classification of adenosine receptors—An update. Pharmacol. Rev. 2011, 63, 1–34. [Google Scholar] [CrossRef]
- Cornelis, M.C.; El-Sohemy, A.; Campos, H. Genetic polymorphism of the adenosine A2A receptor is associated with habitual caffeine consumption. Am. J. Clin. Nutr. 2007, 86, 240–244. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.L.; Qu, W.M.; Eguchi, N.; Chen, J.F.; Schwarzschild, M.A.; Fredholm, B.B.; Urade, Y.; Hayaishi, O. Adenosine A2A, but not A1, receptors mediate the arousal effect of caffeine. Nat. Neurosci. 2005, 8, 858–859. [Google Scholar] [CrossRef]
- Retey, J.V.; Adam, M.; Khatami, R.; Luhmann, U.F.; Jung, H.H.; Berger, W.; Landolt, H.P. A genetic variation in the adenosine A2A receptor gene (ADORA2A) contributes to individual sensitivity to caffeine effects on sleep. Clin. Pharmacol. Ther. 2007, 81, 692–698. [Google Scholar] [CrossRef]
- Alsene, K.; Deckert, J.; Sand, P.; de Wit, H. Association between A2a receptor gene polymorphisms and caffeine-induced anxiety. Neuropsychopharmacology 2003, 28, 1694–1702. [Google Scholar] [CrossRef] [PubMed]
- Leiva, A.; Guzman-Gutierrez, E.; Contreras-Duarte, S.; Fuenzalida, B.; Cantin, C.; Carvajal, L.; Salsoso, R.; Gutierrez, J.; Pardo, F.; Sobrevia, L. Adenosine receptors: Modulators of lipid availability that are controlled by lipid levels. Mol. Asp. Med. 2017, 55, 26–44. [Google Scholar] [CrossRef] [Green Version]
- Eisenstein, A.; Patterson, S.; Ravid, K. The Many Faces of the A2b Adenosine Receptor in Cardiovascular and Metabolic Diseases. J. Cell. Physiol. 2015, 230, 2891–2897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, X.; Mokelke, E.A.; Neeb, Z.P.; Alloosh, M.; Edwards, J.M.; Sturek, M. Adenosine receptor regulation of coronary blood flow in Ossabaw miniature swine. J. Pharmacol. Exp. Ther. 2010, 335, 781–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhai, Y.J.; Liu, P.; He, H.R.; Zheng, X.W.; Wang, Y.; Yang, Q.T.; Dong, Y.L.; Lu, J. The association of ADORA2A and ADORA2B polymorphisms with the risk and severity of chronic heart failure: A case-control study of a northern Chinese population. Int. J. Mol. Sci. 2015, 16, 2732–2746. [Google Scholar] [CrossRef] [Green Version]
- Koupenova, M.; Johnston-Cox, H.; Vezeridis, A.; Gavras, H.; Yang, D.; Zannis, V.; Ravid, K. A2b adenosine receptor regulates hyperlipidemia and atherosclerosis. Circulation 2012, 125, 354–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Han, B.G.; KoGES Group. Cohort Profile: The Korean Genome and Epidemiology Study (KoGES) Consortium. Int. J. Epidemiol. 2017, 46, e20. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.S.; Go, M.J.; Kim, Y.J.; Heo, J.Y.; Oh, J.H.; Ban, H.J.; Yoon, D.; Lee, M.H.; Kim, D.J.; Park, M.; et al. A large-scale genome-wide association study of Asian populations uncovers genetic factors influencing eight quantitative traits. Nat. Genet. 2009, 41, 527–534. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.; Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [Green Version]
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. The Asia-Pacific Perspective: Redefining Obesity and Its Treatment; Health Communications Australia: Sydney, Australia, 2000. [Google Scholar]
- Teng, B.; Smith, J.D.; Rosenfeld, M.E.; Robinet, P.; Davis, M.E.; Morrison, R.R.; Mustafa, S.J. A(1) adenosine receptor deficiency or inhibition reduces atherosclerotic lesions in apolipoprotein E deficient mice. Cardiovasc. Res. 2014, 102, 157–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csoka, B.; Koscso, B.; Toro, G.; Kokai, E.; Virag, L.; Nemeth, Z.H.; Pacher, P.; Bai, P.; Hasko, G. A2B adenosine receptors prevent insulin resistance by inhibiting adipose tissue inflammation via maintaining alternative macrophage activation. Diabetes 2014, 63, 850–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Consortium, G. The Genotype-Tissue Expression (GTEx) project. Nat. Genet. 2013, 45, 580–585. [Google Scholar] [CrossRef]
- Gupta, R.M.; Hadaya, J.; Trehan, A.; Zekavat, S.M.; Roselli, C.; Klarin, D.; Emdin, C.A.; Hilvering, C.R.E.; Bianchi, V.; Mueller, C.; et al. A Genetic Variant Associated with Five Vascular Diseases Is a Distal Regulator of Endothelin-1 Gene Expression. Cell 2017, 170, 522–533. [Google Scholar] [CrossRef] [Green Version]
- Bidel, S.; Hu, G.; Qiao, Q.; Jousilahti, P.; Antikainen, R.; Tuomilehto, J. Coffee consumption and risk of total and cardiovascular mortality among patients with type 2 diabetes. Diabetologia 2006, 49, 2618–2626. [Google Scholar] [CrossRef]
- Grosso, G.; Micek, A.; Godos, J.; Sciacca, S.; Pajak, A.; Martinez-Gonzalez, M.A.; Giovannucci, E.L.; Galvano, F. Coffee consumption and risk of all-cause, cardiovascular, and cancer mortality in smokers and non-smokers: A dose-response meta-analysis. Eur. J. Epidemiol. 2016, 31, 1191–1205. [Google Scholar] [CrossRef]
- Shin, S.; Lim, J.; Lee, H.W.; Kim, C.E.; Kim, S.A.; Lee, J.K.; Kang, D. Association between the prevalence of metabolic syndrome and coffee consumption among Korean adults: Results from the Health Examinees study. Appl. Physiol. Nutr. Metab. 2019, 44, 1371–1378. [Google Scholar] [CrossRef]
- Schliep, K.C.; Schisterman, E.F.; Mumford, S.L.; Pollack, A.Z.; Zhang, C.; Ye, A.; Stanford, J.B.; Hammoud, A.O.; Porucznik, C.A.; Wactawski-Wende, J. Caffeinated beverage intake and reproductive hormones among premenopausal women in the BioCycle Study. Am. J. Clin. Nutr. 2012, 95, 488–497. [Google Scholar] [CrossRef] [Green Version]
- Kolovou, G.D.; Kolovou, V.; Kostakou, P.M.; Mavrogeni, S. Body mass index, lipid metabolism and estrogens: Their impact on coronary heart disease. Curr. Med. Chem. 2014, 21, 3455–3465. [Google Scholar] [CrossRef]
- Doyle, S.E.; Breslin, F.J.; Rieger, J.M.; Beauglehole, A.; Lynch, W.J. Time and sex-dependent effects of an adenosine A2A/A1 receptor antagonist on motivation to self-administer cocaine in rats. Pharmacol. Biochem. Behav. 2012, 102, 257–263. [Google Scholar] [CrossRef] [Green Version]
- Gupta, V.; Goyal, R.; Sharma, P.L. Preconditioning offers cardioprotection in hyperlipidemic rat hearts: Possible role of Dopamine (D2) signaling. BMC Cardiovasc. Disord. 2015, 15, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stonehouse, A.H.; Adachi, M.; Walcott, E.C.; Jones, F.S. Caffeine regulates neuronal expression of the dopamine 2 receptor gene. Mol. Pharmacol. 2003, 64, 1463–1473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richelle, M.; Tavazzi, I.; Offord, E. Comparison of the antioxidant activity of commonly consumed polyphenolic beverages (coffee, cocoa, and tea) prepared per cup serving. J. Agric. Food Chem. 2001, 49, 3438–3442. [Google Scholar] [CrossRef] [PubMed]
- Ohta, T.; Nagashima, J.; Sasai, H.; Ishii, N. Relationship of Cardiorespiratory Fitness and Body Mass Index with the Incidence of Dyslipidemia among Japanese Women: A Cohort Study. Int. J. Environ. Res. Public Health 2019, 16, 4647. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, N.; Sawada, S.S.; Shimada, K.; Lee, I.M.; Gando, Y.; Momma, H.; Kawakami, R.; Miyachi, M.; Hagi, Y.; Kinugawa, C.; et al. Relationship between Cardiorespiratory Fitness and Non-High-Density Lipoprotein Cholesterol: A Cohort Study. J. Atheroscler. Thromb. 2018, 25, 1196–1205. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Gene | SNP ID | Chr | Physical Position | Location | Regulatory Element | Alleles 1 | MAF | HWE |
|---|---|---|---|---|---|---|---|---|
| ADORA1 | rs10800899 | 1 | 203081125 | intron | A/G | 0.1607 | 0.2697 | |
| rs6701725 | 203102728 | intron | A/G | 0.1735 | 0.1381 | |||
| rs10800901 | 203111304 | intron | G/A | 0.4486 | 0.7471 | |||
| ADORA2A | rs5760423 | 22 | 24840118 | intron | T/G | 0.4439 | 0.2536 | |
| ADORA2B | rs17715109 | 17 | 15869557 | intron | H3K4me1, Dnase1 | T/G | 0.0517 | 0.5145 |
| rs2779212 | 15876655 | intron | H3K4me1, eQTL | C/T | 0.2287 | 0.4510 | ||
| ADORA3 | rs3393 | 1 | 112042149 | UTR-3 | Dnase1 | T/C | 0.4457 | 0.7306 |
| rs2786967 | 112075948 | intron | G/A | 0.8891 | 0.8767 |
| Male | Female | |||||
|---|---|---|---|---|---|---|
| Coffee | <1 cup/d | ≥1 cup/d | p2 | <1 cup/d | ≥1 cup/d | p2 |
| (n = 837) | (n = 1690) | (n = 1112) | (n = 1259) | |||
| Age (year) | 56.62 ± 8.90 | 53.95 ± 8.13 | <0.0001 | 57.28 ± 8.75 | 54.55 ± 8.73 | <0.0001 |
| Marriage 3 | ||||||
| Married | 798 (95.34) | 1639 (96.98) | 0.0361 | 939 (84.44) | 1071 (85.07) | 0.6725 |
| Monthly income (×104 KRW) | ||||||
| Low (<100) | 251 (29.99) | 351 (20.77) | <0.0001 | 524 (47.12) | 468 (37.17) | <0.0001 |
| Medium (100–199) | 199 (23.78) | 356 (21.07) | 268 (24.10) | 254 (20.17) | ||
| High (≥200) | 387 (46.24) | 983 (58.17) | 320 (28.78) | 537 (42.65) | ||
| Education (year) | ||||||
| Low (0–6) | 217 (25.93) | 313 (18.52) | <0.0001 | 619 (55.67) | 508 (40.35) | <0.0001 |
| Medium ((7–9) | 171 (20.43) | 348 (20.59) | 222 (19.96) | 264 (20.97) | ||
| High (≥10) | 449 (53.64) | 1029 (60.89) | 271 (24.37) | 487 (36.68) | ||
| Alcohol drinking behavior | ||||||
| Never | 154 (18.40) | 335 (19.82) | 0.6875 | 869 (78.15) | 817 (64.89) | <0.0001 |
| Former | 70 (8.36) | 136 (8.05) | 22 (1.98) | 19 (1.51) | ||
| Current | 613 (73.24) | 1219 (72.13) | 221 (19.87) | 423 (33.60) | ||
| Alcohol intake (g/day) 4 | 25.77 ± 36.07 | 25.00 ± 31.27 | 0.5934 | 3.99 ± 8.89 | 4.88 ± 11.01 | <0.0001 |
| Smoking behavior | ||||||
| Never | 287 (34.29) | 346 (20.47) | <0.0001 | 1094 (98.38) | 1219 (96.82) | 0.0489 |
| Former | 304 (36.32) | 642 (37.99) | 5 (0.45) | 12 (0.95) | ||
| Current | 246 (29.39) | 702 (41.54) | 13 (1.17) | 28 (2.22) | ||
| Tobacco consumption (pack/years) 5 | 16.61 ± 14.57 | 20.25 ± 18.42 | <0.0001 | 7.38 ± 6.04 | 7.42 ± 8.31 | 0.0289 |
| Nutrient intakes | ||||||
| Energy (Kcal) | 1,772.21 ± 510.35 | 1984.88 ± 527.16 | <0.0001 | 1594.52 ± 497.34 | 1753.07 ± 532.77 | <0.0001 |
| Sugar (g/per 1000 Kcal) | 180.35 ± 16.89 | 177.75 ± 14.54 | <0.0001 | 186.28 ± 16.97 | 182.06 ± 15.48 | <0.0001 |
| Fat (g/per 1000 Kcal) | 14.72 ± 5.89 | 16.44 ± 5.13 | <0.0001 | 12.68 ± 5.89 | 14.85 ± 5.44 | <0.0001 |
| Protein (g/per 1000 Kcal) | 32.52 ± 5.93 | 32.48 ± 5.13 | 0.8755 | 31.88 ± 5.30 | 32.44 ± 5.97 | 0.0269 |
| Energy distribution (%) | ||||||
| Carbohydrate | 73.35 ± 7.20 | 71.93 ± 6.13 | <0.0001 | 75.56 ± 7.32 | 73.47 ± 6.63 | <0.0001 |
| Fat | 13.44 ± 5.31 | 14.94 ± 4.61 | <0.0001 | 11.53 ± 5.29 | 13.45 ± 4.85 | <0.0001 |
| Protein | 13.21 ± 2.35 | 13.13 ± 2.03 | 0.4204 | 12.91 ± 2.45 | 13.07 ± 2.33 | 0.0946 |
| SBP (mmHg) | 117.73 ± 15.80 | 116.12 ± 14.87 | 0.0147 | 116.00 ± 17.26 | 113.95 ± 16.84 | 0.0030 |
| DBP (mmHg) | 79.74 ± 10.11 | 79.18 ± 10.10 | 0.1838 | 76.61 ± 10.37 | 75.58 ± 10.53 | 0.0131 |
| Waist circumference (cm) | 84.22 ± 7.76 | 84.77 ± 7.46 | 0.0692 | 83.98 ± 9.68 | 82.76 ± 9.45 | 0.0022 |
| Hip circumference (cm) | 90.90 ± 5.23 | 92.32 ± 5.21 | <0.0001 | 91.13 ± 5.35 | 92.18 ± 5.25 | <0.0001 |
| Height (cm) | 166.32 ± 5.95 | 167.15 ± 5.83 | 0.0008 | 153.23 ± 5.84 | 153.94 ± 5.63 | 0.0026 |
| Weight (kg) | 65.71 ± 9.27 | 68.04 ± 9.57 | <0.0001 | 57.55 ± 8.45 | 58.76 ± 8.03 | 0.0004 |
| BMI (kg/m2) 6 | ||||||
| Underweight (<18.5) | 26 (3.11) | 29 (1.72) | 0.0003 | 18 (1.62) | 15 (1.19) | 0.0739 |
| Normal (18.5–22.9) | 290 (34.65) | 502 (29.70) | 364 (32.73) | 356 (28.28) | ||
| Overweight (23–24.9) | 253 (30.23) | 485 (28.70) | 284 (25.54) | 356 (28.28) | ||
| Obese (≥25) | 268 (32.02) | 674 (39.88) | 446 (40.11) | 532 (42.26) | ||
| HbA1C (%) | 5.41 ± 0.41 | 5.41 ± 0.39 | 0.9575 | 5.47 ± 0.40 | 5.44 ± 0.40 | 0.0628 |
| Total cholesterol (mg/dL) | 183.12 ± 34.42 | 191.06 ± 32.64 | <0.0001 | 193.01 ± 34.56 | 196.65 ± 33.53 | 0.0060 |
| HDL-Cholesterol (mg/dL) | 43.90 ± 10.86 | 43.04 ± 10.24 | 0.0614 | 44.83 ± 9.77 | 46.75 ± 10.16 | <0.0001 |
| Triglyceride (mg/dL) | 144.81 ± 107.99 | 150.9 ± 117.12 | 0.0385 | 124.89 ± 69.71 | 116.24 ± 66.91 | 0.0001 |
| Menopause | 789 (70.95) | 712 (56.55) | <0.0001 | |||
| Female hormone treatment | 25 (2.25) | 34 (2.70) | 0.4804 | |||
| Hypertension 7 | 259 (30.94) | 482 (28.52) | 0.2079 | 367 (33.00) | 333 (26.45) | 0.0005 |
| Coffee | Male | Female | ||||||
|---|---|---|---|---|---|---|---|---|
| Healthy | DLP | Adjusted Model 1 | Healthy | DLP | Adjusted Model 2 | |||
| (n = 1215) | (n = 1312) | OR (95% CI) | p3 | (n = 1330) | (n = 1041) | OR (95% CI) | p3 | |
| <1 cup/d | 427 (35.14) | 410 (31.25) | 1 | 571 (42.93) | 541 (51.97) | 1 | ||
| ≥1 cup/d | 788 (64.86) | 902 (68.75) | 1.107 (0.926–1.323) | 0.2635 | 759 (57.07) | 500 (48.03) | 0.768 (0.645–0.914) | 0.0030 |
| Genes | Alleles | Coffee Intake | Healthy | Dyslipidemia | Adjusted Model 1 | ||
|---|---|---|---|---|---|---|---|
| SNPs | (n = 1215) | (n = 1312) | Odds Ratios (95% CI) | p | p2 | ||
| ADORA1 | |||||||
| rs10800899 | GG | <1 cup/d | 299 (24.61) | 274 (20.88) | 1 | 0.8839 | |
| ≥1 cup/d | 541 (44.53) | 641 (48.86) | 1.176 (0.952–1.453) | 0.1327 | |||
| AG/AA | <1 cup/d | 128 (10.53) | 136 (10.37) | 1.103 (0.816–1.493) | 0.5231 | ||
| ≥1 cup/d | 247 (20.33) | 261 (19.89) | 1.064 (0.828–1.368) | 0.6267 | |||
| rs6701725 | GG | <1 cup/d | 294 (24.20) | 277 (21.11) | 1 | 0.3714 | |
| ≥1 cup/d | 534 (43.95) | 625 (47.64) | 1.127 (0.911–1.394) | 0.2723 | |||
| AG/AA | <1 cup/d | 133 (10.95) | 133 (10.14) | 0.991 (0.733–1.341) | 0.9542 | ||
| ≥1 cup/d | 254 (20.91) | 277 (21.11) | 1.056 (0.823–1.355) | 0.6691 | |||
| rs10800901 | AA | <1 cup/d | 131 (10.78) | 118 (8.99) | 1 | 0.6906 | |
| ≥1 cup/d | 246 (20.25) | 265 (20.20) | 1.115 (0.812–1.532) | 0.5003 | |||
| GA/GG | <1 cup/d | 296 (24.36) | 292 (22.26) | 1.132 (0.833–1.540) | 0.4277 | ||
| ≥1 cup/d | 542 (44.61) | 637 (48.55) | 1.250 (0.938–1.665) | 0.1276 | |||
| ADORA2A | |||||||
| rs5760423 | GG | <1 cup/d | 138 (11.36) | 111 (8.46) | 1 | 0.8317 | |
| ≥1 cup/d | 264 (21.73) | 283 (21.57) | 1.246 (0.910–1.706) | 0.1699 | |||
| TG/TT | <1 cup/d | 289 (23.79) | 299 (22.79) | 1.282 (0.942–1.744) | 0.1139 | ||
| ≥1 cup/d | 524 (43.13) | 619 (47.18) | 1.352 (1.014–1.802) | 0.0402 | |||
| ADORA2B | |||||||
| rs17715109 | GG | <1 cup/d | 381 (31.36) | 360 (27.44) | 1 | 0.8732 | |
| ≥1 cup/d | 712 (58.60) | 817 (62.27) | 1.130 (0.936–1.365) | 0.2020 | |||
| TG/TT | <1 cup/d | 46 (3.79) | 50 (3.81) | 1.149 (0.738–1.786) | 0.5389 | ||
| ≥1 cup/d | 76 (6.26) | 85 (6.48) | 1.076 (0.753–1.536) | 0.6878 | |||
| rs2779212 | TT | <1 cup/d | 252 (20.74) | 226 (17.23) | 1 | 0.0336 | |
| ≥1 cup/d | 494 (40.66) | 555 (42.30) | 1.187 (0.943–1.494) | 0.1433 | |||
| CT/CC | <1 cup/d | 175 (14.40) | 184 (14.02) | 1.155 (0.870–1.534) | 0.3182 | ||
| ≥1 cup/d | 294 (24.20) | 347 (26.45) | 1.165 (0.906–1.497) | 0.2345 | |||
| ADORA3 | |||||||
| rs3393 | CC | <1 cup/d | 122 (10.04) | 147 (11.20) | 1 | 0.5917 | |
| ≥1 cup/d | 228 (18.77) | 284 (21.65) | 0.990 (0.725–1.350) | 0.9477 | |||
| TC/TT | <1 cup/d | 305 (25.10) | 263 (20.05) | 0.731 (0.541– 0.989) | 0.0423 | ||
| ≥1 cup/d | 560 (46.09) | 618 (47.10) | 0.856 (0.647–1.133) | 0.2767 | |||
| rs2786967 | AA | <1 cup/d | 361 (29.71) | 357 (27.21) | 1 | 0.2144 | |
| ≥1 cup/d | 676 (55.64) | 752 (57.32) | 1.036 (0.856– 1.255) | 0.7152 | |||
| GA/GG | <1 cup/d | 66 (5.43) | 53 (4.04) | 0.771 (0.514– 1.155) | 0.2066 | ||
| ≥1 cup/d | 112 (9.22) | 150 (11.43) | 1.257 (0.933–1.694) | 0.1324 | |||
| Genes | Alleles | Coffee Intake | Healthy | Dyslipidemia | Adjusted Model 1 | ||
|---|---|---|---|---|---|---|---|
| SNPs | (n = 1215) | (n = 1312) | Odds ratios (95% CI) | p | p2 | ||
| ADORA1 | |||||||
| rs10800899 | GG | <1 cup/d | 392 (29.47) | 374 (35.93) | 1 | 0.2012 | |
| ≥1 cup/d | 537 (40.38) | 366 (35.16) | 0.789 (0.643–0.968) | 0.0233 | |||
| AG/AA | <1 cup/d | 179 (13.46) | 167 (16.04) | 0.989 (0.761–1.286) | 0.9356 | ||
| ≥1 cup/d | 222 (16.69) | 134 (12.87) | 0.705 (0.537–0.924) | 0.0115 | |||
| rs6701725 | GG | <1 cup/d | 393 (29.55) | 367 (35.25) | 1 | 0.3393 | |
| ≥1 cup/d | 520 (37.10) | 346 (33.24) | 0.784 (0.636–0.966) | 0.0225 | |||
| AG/AA | <1 cup/d | 178 (13.38) | 174 (16.71) | 1.031 (0.794–1.339) | 0.8196 | ||
| ≥1 cup/d | 239 (17.97) | 154 (14.79) | 0.758 (0.585–0.982) | 0.0356 | |||
| rs10800901 | AA | <1 cup/d | 180 (13.53) | 172 (16.52) | 1 | 0.3401 | |
| ≥1 cup/d | 210 (15.79) | 160 (15.37) | 0.891 (0.656–1.210) | 0.4956 | |||
| GA/GG | <1 cup/d | 391 (29.40) | 369 (35.45) | 1.008 (0.777–1.309) | 0.9504 | ||
| ≥1 cup/d | 549 (41.28) | 340 (32.66) | 0.727 (0.560–0.944) | 0.0168 | |||
| ADORA2A | |||||||
| rs5760423 | GG | <1 cup/d | 194 (14.59) | 173 (16.62) | 1 | 0.3188 | |
| ≥1 cup/d | 224 (16.84) | 149 (14.31) | 0.836 (0.617–1.134) | 0.2497 | |||
| TG/TT | <1 cup/d | 377 (28.35) | 368 (35.35) | 1.124 (0.869–1.456) | 0.3736 | ||
| ≥1 cup/d | 535 (40.23) | 351 (33.72) | 0.829 (0.641–1.073) | 0.1539 | |||
| ADORA2B | |||||||
| rs17715109 | GG | <1 cup/d | 508 (38.20) | 486 (46.69) | 1 | 0.7766 | |
| ≥1 cup/d | 671 (50.45) | 461 (44.28) | 0.795 (0.662–0.956) | 0.0146 | |||
| TG/TT | <1 cup/d | 63 (4.74) | 55 (5.28) | 0.903 (0.608–1.339) | 0.6107 | ||
| ≥1 cup/d | 88 (6.62) | 39 (3.75) | 0.500 (0.332–0.754) | 0.0009 | |||
| rs2779212 | TT | <1 cup/d | 343 (25.79) | 335 (32.18) | 1 | 0.8210 | |
| ≥1 cup/d | 421 (31.65) | 301 (28.91) | 0.839 (0.671–1.049) | 0.1226 | |||
| CT/CC | <1 cup/d | 228 (17.14) | 206 (19.79) | 0.943 (0.735–1.210) | 0.6469 | ||
| ≥1 cup/d | 338 (25.41) | 199 (19.12) | 0.645 (0.506–0.823) | 0.0004 | |||
| ADORA3 | |||||||
| rs3393 | CC | <1 cup/d | 179 (13.46) | 163 (15.66) | 1 | 0.9277 | |
| ≥1 cup/d | 247 (18.57) | 157 (15.08) | 0.776 (0.572–1.053) | 0.1035 | |||
| TC/TT | <1 cup/d | 392 (29.47) | 378 (36.31) | 1.053 (0.809–1.370) | 0.7011 | ||
| ≥1 cup/d | 512 (38.50) | 343 (32.95) | 0.806 (0.618–1.051) | 0.1109 | |||
| rs2786967 | AA | <1 cup/d | 486 (36.54) | 444 (42.65) | 1 | 0.0308 | |
| ≥1 cup/d | 617 (46.39) | 411 (39.48) | 0.818 (0.676–0.989) | 0.0384 | |||
| GA/GG | <1 cup/d | 85 (6.39) | 97 (9.32) | 1.306 (0.939–1.816) | 0.1125 | ||
| ≥1 cup/d | 142 (10.68) | 89 (8.55) | 0.737 (0.542–1.002) | 0.0515 | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, J.; Shon, J.; Hwang, J.-Y.; Park, Y.J. Effects of Coffee Intake on Dyslipidemia Risk According to Genetic Variants in the ADORA Gene Family among Korean Adults. Nutrients 2020, 12, 493. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12020493
Han J, Shon J, Hwang J-Y, Park YJ. Effects of Coffee Intake on Dyslipidemia Risk According to Genetic Variants in the ADORA Gene Family among Korean Adults. Nutrients. 2020; 12(2):493. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12020493
Chicago/Turabian StyleHan, Jihee, Jinyoung Shon, Ji-Yun Hwang, and Yoon Jung Park. 2020. "Effects of Coffee Intake on Dyslipidemia Risk According to Genetic Variants in the ADORA Gene Family among Korean Adults" Nutrients 12, no. 2: 493. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12020493