Breast Milk Lipids and Fatty Acids in Regulating Neonatal Intestinal Development and Protecting against Intestinal Injury

 ,
,  , ,
, ,  ,
,
Abstract
:1. Introduction
2. Lipids and Fatty Acid Composition in Human Breast Milk
2.1. Concentrations of Breast Milk Lipids and Fatty Acids
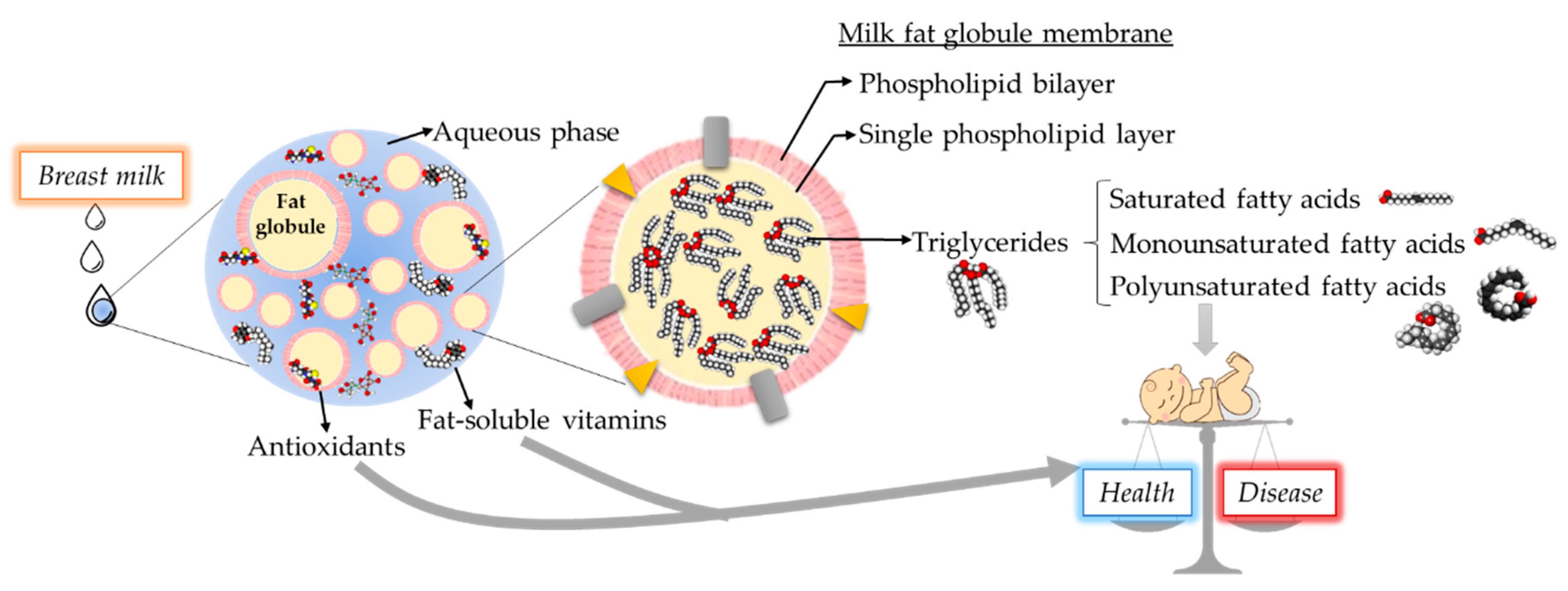
2.2. Breast Milk Fat Globules
2.3. Complex Lipids in Breast Milk
3. Breast Milk Lipids Enhance Neonatal Intestinal Development and Protect against Injury
3.1. Saturated and Monounsaturated Fatty Acids
3.2. Polyunsaturated Fatty Acids
3.3. Milk Fat Globule Membranes
3.4. Complex Lipids
4. Scientific Gaps and Future Directions
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Gila-Diaz, A.; Arribas, S.M.; Algara, A.; Martin-Cabrejas, M.A.; Lopez de Pablo, A.L.; Saenz de Pipaon, M.; Ramiro-Cortijo, D. A review of bioactive factors in human breastmilk: A focus on prematurity. Nutrients 2019, 11, 1307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grote, V.; Verduci, E.; Scaglioni, S.; Vecchi, F.; Contarini, G.; Giovannini, M.; Koletzko, B.; Agostoni, C.; European Childhood Obesity, P. Breast milk composition and infant nutrient intakes during the first 12 months of life. Eur. J. Clin. Nutr. 2016, 70, 250–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munblit, D.; Verhasselt, V.; Warner, J.O. Editorial: Human milk composition and health outcomes in children. Front. Pediatr. 2019, 7, 319. [Google Scholar] [CrossRef] [Green Version]
- Miliku, K.; Duan, Q.L.; Moraes, T.J.; Becker, A.B.; Mandhane, P.J.; Turvey, S.E.; Lefebvre, D.L.; Sears, M.R.; Subbarao, P.; Field, C.J.; et al. Human milk fatty acid composition is associated with dietary, genetic, sociodemographic, and environmental factors in the CHILD Cohort Study. Am. J. Clin. Nutr. 2019, 110, 1370–1383. [Google Scholar] [CrossRef] [PubMed]
- Gidrewicz, D.A.; Fenton, T.R. A systematic review and meta-analysis of the nutrient content of preterm and term breast milk. BMC Pediatr. 2014, 14, 216. [Google Scholar] [CrossRef] [Green Version]
- Floris, L.M.; Stahl, B.; Abrahamse-Berkeveld, M.; Teller, I.C. Human milk fatty acid profile across lactational stages after term and preterm delivery: A pooled data analysis. Prostaglandins Leukot. Essent. Fat. Acids 2019, 102023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyce, C.; Watson, M.; Lazidis, G.; Reeve, S.; Dods, K.; Simmer, K.; McLeod, G. Preterm human milk composition: A systematic literature review. Br. J. Nutr. 2016, 116, 1033–1045. [Google Scholar] [CrossRef]
- Lopez-Lopez, A.; Lopez-Sabater, M.C.; Campoy-Folgoso, C.; Rivero-Urgell, M.; Castellote-Bargallo, A.I. Fatty acid and sn-2 fatty acid composition in human milk from Granada (Spain) and in infant formulas. Eur. J. Clin. Nutr. 2002, 56, 1242–1254. [Google Scholar] [CrossRef] [Green Version]
- Koletzko, B. Human Milk Lipids. Ann. Nutr. Metab. 2016, 69 (Suppl. 2), 28–40. [Google Scholar] [CrossRef] [Green Version]
- Jensen, R.G. Lipids in human milk. Lipids 1999, 34, 1243–1271. [Google Scholar] [CrossRef]
- Martin, C.R.; Ling, P.R.; Blackburn, G.L. Review of infant feeding: Key features of breast milk and infant formula. Nutrients 2016, 8, 279. [Google Scholar] [CrossRef] [Green Version]
- Mozaffarian, D.; Ascherio, A.; Hu, F.B.; Stampfer, M.J.; Willett, W.C.; Siscovick, D.S.; Rimm, E.B. Interplay between different polyunsaturated fatty acids and risk of coronary heart disease in men. Circulation 2005, 111, 157–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aglago, E.K.; Huybrechts, I.; Murphy, N.; Casagrande, C.; Nicolas, G.; Pischon, T.; Fedirko, V.; Severi, G.; Boutron-Ruault, M.C.; Fournier, A.; et al. Consumption of fish and long-chain n-3 polyunsaturated fatty acids is associated with reduced risk of colorectal cancer in a large European cohort. Clin. Gastroenterol. Hepatol. 2019, 18, 654–666. [Google Scholar] [CrossRef] [PubMed]
- Michalski, M.C.; Briard, V.; Michel, F.; Tasson, F.; Poulain, P. Size distribution of fat globules in human colostrum, breast milk, and infant formula. J. Dairy Sci. 2005, 88, 1927–1940. [Google Scholar] [CrossRef] [Green Version]
- Zou, X.Q.; Guo, Z.; Huang, J.H.; Jin, Q.Z.; Cheong, L.Z.; Wang, X.G.; Xu, X.B. Human milk fat globules from different stages of lactation: A lipid composition analysis and microstructure characterization. J. Agric. Food Chem. 2012, 60, 7158–7167. [Google Scholar] [CrossRef]
- Bionaz, M.; Loor, J.J. Gene networks driving bovine milk fat synthesis during the lactation cycle. BMC Genom. 2008, 9, 366. [Google Scholar] [CrossRef] [Green Version]
- Truchet, S.; Honvo-Houeto, E. Physiology of milk secretion. Best Pract. Res. Clin. Endocrinol. Metab. 2017, 31, 367–384. [Google Scholar] [CrossRef]
- Mather, I.H. A review and proposed nomenclature for major proteins of the milk-fat globule membrane. J. Dairy Sci. 2000, 83, 203–247. [Google Scholar] [CrossRef]
- Dewettinck, K.R.; Thienpont, N.; Thien Le, T.; Messens, K.; Van Camp, J. Nutritional and technological aspects of milk fat globule membrane material. Int. Dairy J. 2008, 18, 436–457. [Google Scholar] [CrossRef]
- Chatterton, D.E.; Nguyen, D.N.; Bering, S.B.; Sangild, P.T. Anti-inflammatory mechanisms of bioactive milk proteins in the intestine of newborns. Int. J. Biochem. Cell. Biol. 2013, 45, 1730–1747. [Google Scholar] [CrossRef]
- Bourlieu, C.D.; Deglaire, A.; De Oliveira, S.C.; Ménard, O.; Le Gouar, Y.; Carrière, F.; Dupont, D. Towards infant formula biomimetic of human milk structure and digestive behavior. Proceedings 2017, 24, 17. [Google Scholar] [CrossRef] [Green Version]
- Bourlieu, C.; Menard, O.; De La Chevasnerie, A.; Sams, L.; Rousseau, F.; Madec, M.N.; Robert, B.; Deglaire, A.; Pezennec, S.; Bouhallab, S.; et al. The structure of infant formulas impacts their lipolysis, proteolysis and disintegration during in vitro gastric digestion. Food Chem. 2015, 182, 224–235. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Yang, J.; Yang, D.; Wang, X.; Yang, Z.; Jin, Q.; Wang, M.; Lai, J.; Wang, X. Phospholipid composition and fat globule structure i: Comparison of human milk fat from different gestational ages, lactation stages, and infant formulas. J. Agric. Food Chem. 2019, 67, 13922–13928. [Google Scholar] [CrossRef]
- Gallier, S.; Vocking, K.; Post, J.A.; Van De Heijning, B.; Acton, D.; Van Der Beek, E.M.; Van Baalen, T. A novel infant milk formula concept: Mimicking the human milk fat globule structure. Colloids Surf. B Biointerfaces 2015, 136, 329–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moukarzel, S.; Dyer, R.A.; Garcia, C.; Wiedeman, A.M.; Boyce, G.; Weinberg, J.; Keller, B.O.; Elango, R.; Innis, S.M. Milk fat globule membrane supplementation in formula-fed rat pups improves reflex development and may alter brain lipid composition. Sci. Rep. 2018, 8, 15277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, X.; Xu, W.; Cui, J.; Ma, Y.; Zhou, S. Goat and buffalo milk fat globule membranes exhibit better effects at inducing apoptosis and reduction the viability of HT-29 cells. Sci. Rep. 2019, 9, 2577. [Google Scholar] [CrossRef] [Green Version]
- Hernell, O.; Domellof, M.; Grip, T.; Lonnerdal, B.; Timby, N. Physiological effects of feeding infants and young children formula supplemented with milk fat globule membranes. Nestle Nutr. Inst. Workshop Ser. 2019, 90, 35–42. [Google Scholar] [CrossRef]
- Li, F.; Wu, S.S.; Berseth, C.L.; Harris, C.L.; Richards, J.D.; Wampler, J.L.; Zhuang, W.; Cleghorn, G.; Rudolph, C.D.; Liu, B.; et al. Improved Neurodevelopmental outcomes associated with bovine milk fat globule membrane and lactoferrin in infant formula: A randomized, controlled trial. J. Pediatr. 2019, 215, 24–31. [Google Scholar] [CrossRef]
- Oosting, A.; van Vlies, N.; Kegler, D.; Schipper, L.; Abrahamse-Berkeveld, M.; Ringler, S.; Verkade, H.J.; van der Beek, E.M. Effect of dietary lipid structure in early postnatal life on mouse adipose tissue development and function in adulthood. Br. J. Nutr. 2014, 111, 215–226. [Google Scholar] [CrossRef] [Green Version]
- Armand, M.; Hamosh, M.; Mehta, N.R.; Angelus, P.A.; Philpott, J.R.; Henderson, T.R.; Dwyer, N.K.; Lairon, D.; Hamosh, P. Effect of human milk or formula on gastric function and fat digestion in the premature infant. Pediatr. Res. 1996, 40, 429–437. [Google Scholar] [CrossRef] [Green Version]
- Delplanque, B.; Gibson, R.; Koletzko, B.; Lapillonne, A.; Strandvik, B. Lipid quality in infant nutrition: Current knowledge and future opportunities. J. Pediatr. Gastroenterol. Nutr. 2015, 61, 8–17. [Google Scholar] [CrossRef] [Green Version]
- Giuffrida, F.; Cruz-Hernandez, C.; Fluck, B.; Tavazzi, I.; Thakkar, S.K.; Destaillats, F.; Braun, M. Quantification of phospholipids classes in human milk. Lipids 2013, 48, 1051–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tavazzi, I.; Fontannaz, P.; Lee, L.Y.; Giuffrida, F. Quantification of glycerophospholipids and sphingomyelin in human milk and infant formula by high performance liquid chromatography coupled with mass spectrometer detector. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2018, 1072, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Dinkins, M.B.; Wang, G.; Bieberich, E. Sphingolipid-enriched extracellular vesicles and alzheimer’s disease: A decade of research. J. Alzheimers Dis. 2017, 60, 757–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez, C.; Briard-Bion, V.; Menard, O.; Rousseau, F.; Pradel, P.; Besle, J.M. Phospholipid, sphingolipid, and fatty acid compositions of the milk fat globule membrane are modified by diet. J. Agric. Food Chem. 2008, 56, 5226–5236. [Google Scholar] [CrossRef]
- Moossavi, S.; Atakora, F.; Miliku, K.; Sepehri, S.; Robertson, B.; Duan, Q.L.; Becker, A.B.; Mandhane, P.J.; Turvey, S.E.; Moraes, T.J.; et al. Integrated analysis of human milk microbiota with oligosaccharides and fatty acids in the CHILD cohort. Front. Nutr. 2019, 6, 58. [Google Scholar] [CrossRef]
- Walker, W.A.; Iyengar, R.S. Breast milk, microbiota, and intestinal immune homeostasis. Pediatr. Res. 2015, 77, 220–228. [Google Scholar] [CrossRef] [Green Version]
- Leite, M.E.; Lasekan, J.; Baggs, G.; Ribeiro, T.; Menezes-Filho, J.; Pontes, M.; Druzian, J.; Barreto, D.L.; de Souza, C.O.; Mattos, A.; et al. Calcium and fat metabolic balance, and gastrointestinal tolerance in term infants fed milk-based formulas with and without palm olein and palm kernel oils: A randomized blinded crossover study. BMC Pediatr. 2013, 13, 215. [Google Scholar] [CrossRef] [Green Version]
- Carnielli, V.P.; Luijendijk, I.H.; van Beek, R.H.; Boerma, G.J.; Degenhart, H.J.; Sauer, P.J. Effect of dietary triacylglycerol fatty acid positional distribution on plasma lipid classes and their fatty acid composition in preterm infants. Am. J. Clin. Nutr. 1995, 62, 776–781. [Google Scholar] [CrossRef] [Green Version]
- Petit, V.; Sandoz, L.; Garcia-Rodenas, C.L. Importance of the regiospecific distribution of long-chain saturated fatty acids on gut comfort, fat and calcium absorption in infants. Prostaglandins Leukot. Essent. Fat. Acids 2017, 121, 40–51. [Google Scholar] [CrossRef]
- Beghin, L.; Marchandise, X.; Lien, E.; Bricout, M.; Bernet, J.P.; Lienhardt, J.F.; Jeannerot, F.; Menet, V.; Requillart, J.C.; Marx, J.; et al. Growth, stool consistency and bone mineral content in healthy term infants fed sn-2-palmitate-enriched starter infant formula: A randomized, double-blind, multicentre clinical trial. Clin. Nutr. 2019, 38, 1023–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miles, E.A.; Calder, P.C. The influence of the position of palmitate in infant formula triacylglycerols on health outcomes. Nutr. Res. 2017, 44, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.; Bar-Yoseph, F.; Levi, L.; Lifshitz, Y.; Witte-Bouma, J.; de Bruijn, A.C.; Korteland-van Male, A.M.; van Goudoever, J.B.; Renes, I.B. High beta-palmitate fat controls the intestinal inflammatory response and limits intestinal damage in mucin Muc2 deficient mice. PLoS ONE 2013, 8, e65878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, T.; Liu, B.; Li, J.; Dong, X.; Lin, M.; Zhang, M.; Zhao, J.; Dai, Y.; Chen, L. Association between sn-2 fatty acid profiles of breast milk and development of the infant intestinal microbiome. Food Funct. 2018, 9, 1028–1037. [Google Scholar] [CrossRef]
- Bronsky, J.; Campoy, C.; Embleton, N.; Fewtrell, M.; Mis, N.F.; Gerasimidis, K.; Hojsak, I.; Hulst, J.; Indrio, F.; Lapillonne, A.; et al. Palm oil and beta-palmitate in infant formula: A position paper by the european society for paediatric gastroenterology, hepatology, and nutrition (ESPGHAN) committee on nutrition. J. Pediatr. Gastroenterol. Nutr. 2019, 68, 742–760. [Google Scholar] [CrossRef]
- Zou, L.; Pande, G.; Akoh, C.C. Infant formula fat analogs and human milk fat: New focus on infant developmental needs. Annu. Rev. Food Sci. Technol. 2016, 7, 139–165. [Google Scholar] [CrossRef]
- Sales-Campos, H.; Souza, P.R.; Peghini, B.C.; da Silva, J.S.; Cardoso, C.R. An overview of the modulatory effects of oleic acid in health and disease. Mini Rev. Med. Chem. 2013, 13, 201–210. [Google Scholar]
- Cruz-Chamorro, L.; Puertollano, E.; de Cienfuegos, G.A.; Puertollano, M.A.; de Pablo, M.A. Acquired resistance to Listeria monocytogenes during a secondary infection in a murine model fed dietary lipids. Nutrition 2011, 27, 1053–1060. [Google Scholar] [CrossRef]
- Cury-Boaventura, M.F.; Gorjao, R.; de Lima, T.M.; Newsholme, P.; Curi, R. Comparative toxicity of oleic and linoleic acid on human lymphocytes. Life Sci. 2006, 78, 1448–1456. [Google Scholar] [CrossRef]
- Martin, C.R.; Dasilva, D.A.; Cluette-Brown, J.E.; Dimonda, C.; Hamill, A.; Bhutta, A.Q.; Coronel, E.; Wilschanski, M.; Stephens, A.J.; Driscoll, D.F.; et al. Decreased postnatal docosahexaenoic and arachidonic acid blood levels in premature infants are associated with neonatal morbidities. J. Pediatr. 2011, 159, 743–749.e2. [Google Scholar] [CrossRef] [Green Version]
- Lofqvist, C.A.; Najm, S.; Hellgren, G.; Engstrom, E.; Savman, K.; Nilsson, A.K.; Andersson, M.X.; Hard, A.L.; Smith, L.E.H.; Hellstrom, A. Association of retinopathy of prematurity with low levels of arachidonic acid: A secondary analysis of a randomized clinical trial. JAMA Ophthalmol. 2018, 136, 271–277. [Google Scholar] [CrossRef] [Green Version]
- Singh, P.; Ochoa-Allemant, P.; Brown, J.; Perides, G.; Freedman, S.D.; Martin, C.R. Effect of polyunsaturated fatty acids on postnatal ileum development using the fat-1 transgenic mouse model. Pediatr. Res. 2019, 85, 556–565. [Google Scholar] [CrossRef] [PubMed]
- Najm, S.; Lofqvist, C.; Hellgren, G.; Engstrom, E.; Lundgren, P.; Hard, A.L.; Lapillonne, A.; Savman, K.; Nilsson, A.K.; Andersson, M.X.; et al. Effects of a lipid emulsion containing fish oil on polyunsaturated fatty acid profiles, growth and morbidities in extremely premature infants: A randomized controlled trial. Clin. Nutr. ESPEN 2017, 20, 17–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akinsulire, O.; Perides, G.; Anez-Bustillos, L.; Cluette-Brown, J.; Nedder, A.; Pollack, E.; Singh, P.; Liu, Y.; Sanchez-Fernandez, L.L.; Obregon, E.; et al. Early enteral administration of a complex lipid emulsion supplement prevents postnatal deficits in docosahexaenoic and arachidonic acids and increases tissue accretion of lipophilic nutrients in preterm piglets. JPEN J. Parenter. Enteral. Nutr. 2020, 44, 69–79. [Google Scholar] [CrossRef] [Green Version]
- Vegge, A.; Thymann, T.; Lauritzen, L.; Bering, S.B.; Wiinberg, B.; Sangild, P.T. Parenteral lipids and partial enteral nutrition affect hepatic lipid composition but have limited short term effects on formula-induced necrotizing enterocolitis in preterm piglets. Clin. Nutr. 2015, 34, 219–228. [Google Scholar] [CrossRef]
- Wang, X.; Liu, F.; Wang, Y.M.; Xue, C.H.; Tang, Q.J. The modulation effect of triglyceride type and phospholipids type ω-3 LCPUFA on mice gut microbiota. J. Biosci. Med. 2017, 5, 11. [Google Scholar] [CrossRef] [Green Version]
- Calder, P.C. Marine omega-3 fatty acids and inflammatory processes: Effects, mechanisms and clinical relevance. Biochim. Biophys. Acta 2015, 1851, 469–484. [Google Scholar] [CrossRef]
- Marion-Letellier, R.; Savoye, G.; Beck, P.L.; Panaccione, R.; Ghosh, S. Polyunsaturated fatty acids in inflammatory bowel diseases: A reappraisal of effects and therapeutic approaches. Inflamm. Bowel Dis. 2013, 19, 650–661. [Google Scholar] [CrossRef]
- John, S.; Luben, R.; Shrestha, S.S.; Welch, A.; Khaw, K.T.; Hart, A.R. Dietary n-3 polyunsaturated fatty acids and the aetiology of ulcerative colitis: A UK prospective cohort study. Eur. J. Gastroenterol. Hepatol. 2010, 22, 602–606. [Google Scholar] [CrossRef]
- Barbosa, D.S.; Cecchini, R.; El Kadri, M.Z.; Rodriguez, M.A.; Burini, R.C.; Dichi, I. Decreased oxidative stress in patients with ulcerative colitis supplemented with fish oil omega-3 fatty acids. Nutrition 2003, 19, 837–842. [Google Scholar] [CrossRef]
- Chan, S.S.; Luben, R.; Olsen, A.; Tjonneland, A.; Kaaks, R.; Lindgren, S.; Grip, O.; Bergmann, M.M.; Boeing, H.; Hallmans, G.; et al. Association between high dietary intake of the n-3 polyunsaturated fatty acid docosahexaenoic acid and reduced risk of Crohn’s disease. Aliment. Pharmacol. Ther. 2014, 39, 834–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, S.S.; Hart, A.R. Commentary: The association between high dietary intake of docosahexaenoic acid and reduced risk of Crohn’s disease--authors’ reply. Aliment. Pharmacol. Ther. 2014, 39, 1332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dichi, I.; Frenhane, P.; Dichi, J.B.; Correa, C.R.; Angeleli, A.Y.; Bicudo, M.H.; Rodrigues, M.A.; Victoria, C.R.; Burini, R.C. Comparison of omega-3 fatty acids and sulfasalazine in ulcerative colitis. Nutrition 2000, 16, 87–90. [Google Scholar] [CrossRef]
- Lev-Tzion, R.; Griffiths, A.M.; Leder, O.; Turner, D. Omega 3 fatty acids (fish oil) for maintenance of remission in Crohn’s disease. Cochrane Database Syst. Rev. 2014. [Google Scholar] [CrossRef]
- Feagan, B.G.; Sandborn, W.J.; Mittmann, U.; Bar-Meir, S.; D’Haens, G.; Bradette, M.; Cohen, A.; Dallaire, C.; Ponich, T.P.; McDonald, J.W.; et al. Omega-3 free fatty acids for the maintenance of remission in Crohn disease: The EPIC randomized controlled trials. JAMA 2008, 299, 1690–1697. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Jilling, T.; Li, D.; Caplan, M.S. Polyunsaturated fatty acid supplementation alters proinflammatory gene expression and reduces the incidence of necrotizing enterocolitis in a neonatal rat model. Pediatr. Res. 2007, 61, 427–432. [Google Scholar] [CrossRef] [Green Version]
- Wijendran, V.; Brenna, J.T.; Wang, D.H.; Zhu, W.; Meng, D.; Ganguli, K.; Kothapalli, K.S.; Requena, P.; Innis, S.; Walker, W.A. Long-chain polyunsaturated fatty acids attenuate the IL-1beta-induced proinflammatory response in human fetal intestinal epithelial cells. Pediatr. Res. 2015, 78, 626–633. [Google Scholar] [CrossRef] [Green Version]
- Whiting, C.V.; Bland, P.W.; Tarlton, J.F. Dietary n-3 polyunsaturated fatty acids reduce disease and colonic proinflammatory cytokines in a mouse model of colitis. Inflamm. Bowel Dis. 2005, 11, 340–349. [Google Scholar] [CrossRef]
- Usami, M.; Muraki, K.; Iwamoto, M.; Ohata, A.; Matsushita, E.; Miki, A. Effect of eicosapentaenoic acid (EPA) on tight junction permeability in intestinal monolayer cells. Clin. Nutr. 2001, 20, 351–359. [Google Scholar] [CrossRef]
- Ferrer, R.; Moreno, J.J. Role of eicosanoids on intestinal epithelial homeostasis. Biochem. Pharmacol. 2010, 80, 431–438. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Zhang, Q.; Wang, M.; Zhao, S.; Xu, G.; Li, J. n-3 polyunsaturated fatty acids prevent disruption of epithelial barrier function induced by proinflammatory cytokines. Mol. Immunol. 2008, 45, 1356–1365. [Google Scholar] [CrossRef]
- Willemsen, L.E.; Koetsier, M.A.; Balvers, M.; Beermann, C.; Stahl, B.; van Tol, E.A. Polyunsaturated fatty acids support epithelial barrier integrity and reduce IL-4 mediated permeability in vitro. Eur. J. Nutr. 2008, 47, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Charpentier, C.; Chan, R.; Salameh, E.; Mbodji, K.; Ueno, A.; Coeffier, M.; Guerin, C.; Ghosh, S.; Savoye, G.; Marion-Letellier, R. Dietary n-3 PUFA may attenuate experimental colitis. Mediat. Inflamm. 2018, 2018, 8430614. [Google Scholar] [CrossRef] [PubMed]
- Bento, A.F.; Claudino, R.F.; Dutra, R.C.; Marcon, R.; Calixto, J.B. Omega-3 fatty acid-derived mediators 17(R)-hydroxy docosahexaenoic acid, aspirin-triggered resolvin D1 and resolvin D2 prevent experimental colitis in mice. J. Immunol. 2011, 187, 1957–1969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcon, R.; Bento, A.F.; Dutra, R.C.; Bicca, M.A.; Leite, D.F.; Calixto, J.B. Maresin 1, a proresolving lipid mediator derived from omega-3 polyunsaturated fatty acids, exerts protective actions in murine models of colitis. J. Immunol. 2013, 191, 4288–4298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, J.Y.; Chi, S.G.; Chun, H.S. Oral administration of docosahexaenoic acid attenuates colitis induced by dextran sulfate sodium in mice. Mol. Nutr. Food Res. 2011, 55, 239–246. [Google Scholar] [CrossRef]
- Camuesco, D.; Comalada, M.; Concha, A.; Nieto, A.; Sierra, S.; Xaus, J.; Zarzuelo, A.; Galvez, J. Intestinal anti-inflammatory activity of combined quercitrin and dietary olive oil supplemented with fish oil, rich in EPA and DHA (n-3) polyunsaturated fatty acids, in rats with DSS-induced colitis. Clin. Nutr. 2006, 25, 466–476. [Google Scholar] [CrossRef] [PubMed]
- Camuesco, D.; Galvez, J.; Nieto, A.; Comalada, M.; Rodriguez-Cabezas, M.E.; Concha, A.; Xaus, J.; Zarzuelo, A. Dietary olive oil supplemented with fish oil, rich in EPA and DHA (n-3) polyunsaturated fatty acids, attenuates colonic inflammation in rats with DSS-induced colitis. J. Nutr. 2005, 135, 687–694. [Google Scholar] [CrossRef] [Green Version]
- Gobbetti, T.; Ducheix, S.; le Faouder, P.; Perez, T.; Riols, F.; Boue, J.; Bertrand-Michel, J.; Dubourdeau, M.; Guillou, H.; Perretti, M.; et al. Protective effects of n-6 fatty acids-enriched diet on intestinal ischaemia/reperfusion injury involve lipoxin A4 and its receptor. Br. J. Pharmacol. 2015, 172, 910–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dufton, N.; Perretti, M. Therapeutic anti-inflammatory potential of formyl-peptide receptor agonists. Pharmacol. Ther. 2010, 127, 175–188. [Google Scholar] [CrossRef]
- Caplan, M.S.; Russell, T.; Xiao, Y.; Amer, M.; Kaup, S.; Jilling, T. Effect of polyunsaturated fatty acid (PUFA) supplementation on intestinal inflammation and necrotizing enterocolitis (NEC) in a neonatal rat model. Pediatr. Res. 2001, 49, 647–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Plaen, I.G.; Liu, S.X.; Tian, R.; Neequaye, I.; May, M.J.; Han, X.B.; Hsueh, W.; Jilling, T.; Lu, J.; Caplan, M.S. Inhibition of nuclear factor-kappaB ameliorates bowel injury and prolongs survival in a neonatal rat model of necrotizing enterocolitis. Pediatr. Res. 2007, 61, 716–721. [Google Scholar] [CrossRef] [Green Version]
- Caplan, M.S.; Jilling, T. The role of polyunsaturated fatty acid supplementation in intestinal inflammation and neonatal necrotizing enterocolitis. Lipids 2001, 36, 1053–1057. [Google Scholar] [CrossRef] [PubMed]
- Ohtsuka, Y.; Okada, K.; Yamakawa, Y.; Ikuse, T.; Baba, Y.; Inage, E.; Fujii, T.; Izumi, H.; Oshida, K.; Nagata, S.; et al. omega-3 fatty acids attenuate mucosal inflammation in premature rat pups. J. Pediatr. Surg. 2011, 46, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Akisu, M.; Baka, M.; Coker, I.; Kultursay, N.; Huseyinov, A. Effect of dietary n-3 fatty acids on hypoxia-induced necrotizing enterocolitis in young mice. n-3 fatty acids alter platelet-activating factor and leukotriene B4 production in the intestine. Biol. Neonate 1998, 74, 31–38. [Google Scholar] [CrossRef]
- Smithers, L.G.; Gibson, R.A.; McPhee, A.; Makrides, M. Effect of long-chain polyunsaturated fatty acid supplementation of preterm infants on disease risk and neurodevelopment: A systematic review of randomized controlled trials. Am. J. Clin. Nutr. 2008, 87, 912–920. [Google Scholar] [CrossRef] [Green Version]
- Fewtrell, M.S.; Abbott, R.A.; Kennedy, K.; Singhal, A.; Morley, R.; Caine, E.; Jamieson, C.; Cockburn, F.; Lucas, A. Randomized, double-blind trial of long-chain polyunsaturated fatty acid supplementation with fish oil and borage oil in preterm infants. J. Pediatr. 2004, 144, 471–479. [Google Scholar] [CrossRef]
- Carlson, S.E.; Montalto, M.B.; Ponder, D.L.; Werkman, S.H.; Korones, S.B. Lower incidence of necrotizing enterocolitis in infants fed a preterm formula with egg phospholipids. Pediatr. Res. 1998, 44, 491–498. [Google Scholar] [CrossRef]
- Zhang, P.; Lavoie, P.M.; Lacaze-Masmonteil, T.; Rhainds, M.; Marc, I. Omega-3 long-chain polyunsaturated fatty acids for extremely preterm infants: A systematic review. Pediatrics 2014, 134, 120–134. [Google Scholar] [CrossRef] [Green Version]
- Fewtrell, M.S.; Morley, R.; Abbott, R.A.; Singhal, A.; Isaacs, E.B.; Stephenson, T.; MacFadyen, U.; Lucas, A. Double-blind, randomized trial of long-chain polyunsaturated fatty acid supplementation in formula fed to preterm infants. Pediatrics 2002, 110, 73–82. [Google Scholar] [CrossRef]
- Innis, S.M.; Adamkin, D.H.; Hall, R.T.; Kalhan, S.C.; Lair, C.; Lim, M.; Stevens, D.C.; Twist, P.F.; Diersen-Schade, D.A.; Harris, C.L.; et al. Docosahexaenoic acid and arachidonic acid enhance growth with no adverse effects in preterm infants fed formula. J. Pediatr. 2002, 140, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Clandinin, M.T.; Van Aerde, J.E.; Merkel, K.L.; Harris, C.L.; Springer, M.A.; Hansen, J.W.; Diersen-Schade, D.A. Growth and development of preterm infants fed infant formulas containing docosahexaenoic acid and arachidonic acid. J. Pediatr. 2005, 146, 461–468. [Google Scholar] [CrossRef] [PubMed]
- Henriksen, C.; Haugholt, K.; Lindgren, M.; Aurvag, A.K.; Ronnestad, A.; Gronn, M.; Solberg, R.; Moen, A.; Nakstad, B.; Berge, R.K.; et al. Improved cognitive development among preterm infants attributable to early supplementation of human milk with docosahexaenoic acid and arachidonic acid. Pediatrics 2008, 121, 1137–1145. [Google Scholar] [CrossRef] [PubMed]
- Makrides, M.; Gibson, R.A.; McPhee, A.J.; Collins, C.T.; Davis, P.G.; Doyle, L.W.; Simmer, K.; Colditz, P.B.; Morris, S.; Smithers, L.G.; et al. Neurodevelopmental outcomes of preterm infants fed high-dose docosahexaenoic acid: A randomized controlled trial. JAMA 2009, 301, 175–182. [Google Scholar] [CrossRef] [Green Version]
- Collins, C.T.; Gibson, R.A.; Makrides, M.; McPhee, A.J.; Sullivan, T.R.; Davis, P.G.; Thio, M.; Simmer, K.; Rajadurai, V.S.; Team, N.R.I. The N3RO trial: A randomised controlled trial of docosahexaenoic acid to reduce bronchopulmonary dysplasia in preterm infants < 29 weeks’ gestation. BMC Pediatr. 2016, 16, 72. [Google Scholar] [CrossRef] [Green Version]
- Collins, C.T.; Makrides, M.; McPhee, A.J.; Sullivan, T.R.; Davis, P.G.; Thio, M.; Simmer, K.; Rajadurai, V.S.; Travadi, J.; Berry, M.J.; et al. Docosahexaenoic acid and bronchopulmonary dysplasia in preterm infants. N. Engl. J. Med. 2017, 376, 1245–1255. [Google Scholar] [CrossRef]
- Koning, N.; Kessen, S.F.; Van Der Voorn, J.P.; Appelmelk, B.J.; Jeurink, P.V.; Knippels, L.M.; Garssen, J.; Van Kooyk, Y. Human milk blocks DC-SIGN-pathogen interaction via MUC1. Front. Immunol. 2015, 6, 112. [Google Scholar] [CrossRef] [Green Version]
- Yolken, R.H.; Peterson, J.A.; Vonderfecht, S.L.; Fouts, E.T.; Midthun, K.; Newburg, D.S. Human milk mucin inhibits rotavirus replication and prevents experimental gastroenteritis. J. Clin. Investig. 1992, 90, 1984–1991. [Google Scholar] [CrossRef] [Green Version]
- Stevens, C.R.; Millar, T.M.; Clinch, J.G.; Kanczler, J.M.; Bodamyali, T.; Blake, D.R. Antibacterial properties of xanthine oxidase in human milk. Lancet 2000, 356, 829–830. [Google Scholar] [CrossRef]
- Hanayama, R.; Tanaka, M.; Miwa, K.; Shinohara, A.; Iwamatsu, A.; Nagata, S. Identification of a factor that links apoptotic cells to phagocytes. Nature 2002, 417, 182–187. [Google Scholar] [CrossRef]
- Miksa, M.; Wu, R.; Dong, W.; Das, P.; Yang, D.; Wang, P. Dendritic cell-derived exosomes containing milk fat globule epidermal growth factor-factor VIII attenuate proinflammatory responses in sepsis. Shock 2006, 25, 586–593. [Google Scholar] [CrossRef] [PubMed]
- Cui, T.; Miksa, M.; Wu, R.; Komura, H.; Zhou, M.; Dong, W.; Wang, Z.; Higuchi, S.; Chaung, W.; Blau, S.A.; et al. Milk fat globule epidermal growth factor 8 attenuates acute lung injury in mice after intestinal ischemia and reperfusion. Am. J. Respir. Crit. Care Med. 2010, 181, 238–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, A.; Jacob, A.; Wu, R.; Zhou, M.; Nicastro, J.M.; Coppa, G.F.; Wang, P. Milk fat globule-EGF factor VIII in sepsis and ischemia-reperfusion injury. Mol. Med. 2011, 17, 126–133. [Google Scholar] [CrossRef]
- Chogle, A.; Bu, H.F.; Wang, X.; Brown, J.B.; Chou, P.M.; Tan, X.D. Milk fat globule-EGF factor 8 is a critical protein for healing of dextran sodium sulfate-induced acute colitis in mice. Mol. Med. 2011, 17, 502–507. [Google Scholar] [CrossRef]
- Bu, H.F.; Zuo, X.L.; Wang, X.; Ensslin, M.A.; Koti, V.; Hsueh, W.; Raymond, A.S.; Shur, B.D.; Tan, X.D. Milk fat globule-EGF factor 8/lactadherin plays a crucial role in maintenance and repair of murine intestinal epithelium. J. Clin. Investig. 2007, 117, 3673–3683. [Google Scholar] [CrossRef] [Green Version]
- Ogg, S.L.; Weldon, A.K.; Dobbie, L.; Smith, A.J.; Mather, I.H. Expression of butyrophilin (Btn1a1) in lactating mammary gland is essential for the regulated secretion of milk-lipid droplets. Proc. Natl. Acad. Sci. USA 2004, 101, 10084–10089. [Google Scholar] [CrossRef] [Green Version]
- Rhodes, D.A.; Reith, W.; Trowsdale, J. Regulation of Immunity by Butyrophilins. Annu. Rev. Immunol. 2016, 34, 151–172. [Google Scholar] [CrossRef] [Green Version]
- Bhinder, G.; Allaire, J.M.; Garcia, C.; Lau, J.T.; Chan, J.M.; Ryz, N.R.; Bosman, E.S.; Graef, F.A.; Crowley, S.M.; Celiberto, L.S.; et al. Milk Fat globule membrane supplementation in formula modulates the neonatal gut microbiome and normalizes intestinal development. Sci. Rep. 2017, 7, 45274. [Google Scholar] [CrossRef]
- Huang, S.; Wu, Z.; Liu, C.; Han, D.; Feng, C.; Wang, S.; Wang, J. Milk fat globule membrane supplementation promotes neonatal growth and alleviates inflammation in low-birth-weight mice treated with lipopolysaccharide. Biomed. Res. Int. 2019, 2019, 4876078. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Wen, J.; Zhou, J.; Cai, W.; Qian, L. Milk fat globule membrane ameliorates necrotizing enterocolitis in neonatal rats and suppresses lipopolysaccharide-induced inflammatory response in IEC-6 enterocytes. JPEN J. Parenter. Enteral. Nutr. 2019, 43, 863–873. [Google Scholar] [CrossRef]
- Zavaleta, N.; Kvistgaard, A.S.; Graverholt, G.; Respicio, G.; Guija, H.; Valencia, N.; Lonnerdal, B. Efficacy of an MFGM-enriched complementary food in diarrhea, anemia, and micronutrient status in infants. J. Pediatr. Gastroenterol. Nutr. 2011, 53, 561–568. [Google Scholar] [CrossRef] [PubMed]
- Miyake, H.; Lee, C.; Chusilp, S.; Bhalla, M.; Li, B.; Pitino, M.; Seo, S.; O’Connor, D.L.; Pierro, A. Human breast milk exosomes attenuate intestinal damage. Pediatr. Surg. Int. 2020, 36, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Zhang, R.; Qian, T.; Peng, X.; He, W.; Zheng, S.; Cao, Y.; Pierro, A.; Shen, C. A comparison of exosomes derived from different periods breast milk on protecting against intestinal organoid injury. Pediatr. Surg. Int. 2019, 35, 1363–1368. [Google Scholar] [CrossRef] [PubMed]
- Matei, A.C.; Antounians, L.; Zani, A. Extracellular Vesicles as a potential therapy for neonatal conditions: State of the art and challenges in clinical translation. Pharmaceutics 2019, 11, 404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, C.; Patel, M.; Williams, S.; Arora, H.; Brawner, K.; Sims, B. Human breast milk-derived exosomes attenuate cell death in intestinal epithelial cells. Innate Immun. 2018, 24, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yan, X.; Zhang, L.; Cai, J.; Zhou, Y.; Liu, H.; Hu, Y.; Chen, W.; Xu, S.; Liu, P.; et al. Identification and peptidomic profiling of exosomes in preterm human milk: Insights into necrotizing enterocolitis prevention. Mol. Nutr. Food Res. 2019, 16, 1801247. [Google Scholar] [CrossRef]
- Norris, G.H.; Milard, M.; Michalski, M.C.; Blesso, C.N. Protective properties of milk sphingomyelin against dysfunctional lipid metabolism, gut dysbiosis, and inflammation. J. Nutr. Biochem. 2019, 73, 108224. [Google Scholar] [CrossRef]
- Nilsson, A. Role of sphingolipids in infant gut health and immunity. J. Pediatr. 2016, 173, S53–S59. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Cheng, Y.; Hansen, G.H.; Niels-Christiansen, L.L.; Koentgen, F.; Ohlsson, L.; Nilsson, A.; Duan, R.D. Crucial role of alkaline sphingomyelinase in sphingomyelin digestion: A study on enzyme knockout mice. J. Lipid Res. 2011, 52, 771–781. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, A. The presence of spingomyelin- and ceramide-cleaving enzymes in the small intestinal tract. Biochim. Biophys. Acta 1969, 176, 339–347. [Google Scholar] [CrossRef]
- Wu, J.; Nilsson, A.; Jonsson, B.A.; Stenstad, H.; Agace, W.; Cheng, Y.; Duan, R.D. Intestinal alkaline sphingomyelinase hydrolyses and inactivates platelet-activating factor by a phospholipase C activity. Biochem. J. 2006, 394, 299–308. [Google Scholar] [CrossRef] [Green Version]
- Andersson, D.; Kotarsky, K.; Wu, J.; Agace, W.; Duan, R.D. Expression of alkaline sphingomyelinase in yeast cells and anti-inflammatory effects of the expressed enzyme in a rat colitis model. Dig. Dis. Sci. 2009, 54, 1440–1448. [Google Scholar] [CrossRef]
- Duan, R.D.; Cheng, Y.; Jonsson, B.A.; Ohlsson, L.; Herbst, A.; Hellstrom-Westas, L.; Nilsson, A. Human meconium contains significant amounts of alkaline sphingomyelinase, neutral ceramidase, and sphingolipid metabolites. Pediatr. Res. 2007, 61, 61–66. [Google Scholar] [CrossRef] [Green Version]
- Norris, G.H.; Jiang, C.; Ryan, J.; Porter, C.M.; Blesso, C.N. Milk sphingomyelin improves lipid metabolism and alters gut microbiota in high fat diet-fed mice. J. Nutr. Biochem. 2016, 30, 93–101. [Google Scholar] [CrossRef]
- Miklavcic, J.J.; Schnabl, K.L.; Mazurak, V.C.; Thomson, A.B.; Clandinin, M.T. Dietary ganglioside reduces proinflammatory signaling in the intestine. J. Nutr. Metab. 2012, 2012, 280286. [Google Scholar] [CrossRef]
- Schnabl, K.L.; Larsen, B.; Van Aerde, J.E.; Lees, G.; Evans, M.; Belosevic, M.; Field, C.; Thomson, A.B.; Clandinin, M.T. Gangliosides protect bowel in an infant model of necrotizing enterocolitis by suppressing proinflammatory signals. J. Pediatr. Gastroenterol. Nutr. 2009, 49, 382–392. [Google Scholar] [CrossRef] [PubMed]
- Park, E.J.; Suh, M.; Thomson, B.; Thomson, A.B.; Ramanujam, K.S.; Clandinin, M.T. Dietary ganglioside decreases cholesterol content, caveolin expression and inflammatory mediators in rat intestinal microdomains. Glycobiology 2005, 15, 935–942. [Google Scholar] [CrossRef] [PubMed]
- Park, E.J.; Suh, M.; Thomson, B.; Ma, D.W.; Ramanujam, K.; Thomson, A.B.; Clandinin, M.T. Dietary ganglioside inhibits acute inflammatory signals in intestinal mucosa and blood induced by systemic inflammation of Escherichia coli lipopolysaccharide. Shock 2007, 28, 112–117. [Google Scholar] [CrossRef]
- Rueda, R.; Sabatel, J.L.; Maldonado, J.; Molina-Font, J.A.; Gil, A. Addition of gangliosides to an adapted milk formula modifies levels of fecal Escherichia coli in preterm newborn infants. J. Pediatr. 1998, 133, 90–94. [Google Scholar] [CrossRef]
- Calder, P.C. Functional roles of fatty acids and their effects on human health. JPEN J. Parenter. Enteral. Nutr. 2015, 39, 18S–32S. [Google Scholar] [CrossRef]
- Koletzko, B.; Bergmann, K.; Brenna, J.T.; Calder, P.C.; Campoy, C.; Clandinin, M.T.; Colombo, J.; Daly, M.; Decsi, T.; Demmelmair, H.; et al. Should formula for infants provide arachidonic acid along with DHA? A position paper of the European Academy of Paediatrics and the Child Health Foundation. Am. J Clin. Nutr. 2020, 111, 10–16. [Google Scholar] [CrossRef]


{kind=link}
| Term Infants | Preterm Infants | |||||
|---|---|---|---|---|---|---|
| Colostrum | Transitional | Mature | Colostrum | Transitional | Mature | |
| Saturated Fatty Acids | ||||||
| Caprylic acid (C8:0) | 0.07–0.19 | 0.2–0.31 | 0.2–0.3 | 0.03–0.03 | 0.09–0.11 | 0.16–0.16 |
| Capric acid (C10:0) | 0.5–1.04 | 1.2–1.6 | 1.5–1.8 | 0.09–0.09 | 1.0–1.7 | 1.2–2.1 |
| Lauric acid (C12:0) | 2.8–3.5 | 5.4–6.6 | 5.7–6.5 | 3.2–4.6 | 5.7–7.5 | 5.7–8.1 |
| Myristic acid (C14:0) | 5.4–6.0 | 6.6–7.5 | 6.5–7.1 | 5.8–7.2 | 8.0–9.2 | 7.4–9.0 |
| Palmitic acid (C16:0) | 24.3–25.5 | 21.9–23.3 | 21.7–22.7 | 22.5–24.1 | 21.5–23.5 | 20.9–22.3 |
| Stearic acid (C18:0) | 6.2–6.6 | 6.1–6.7 | 6.3–6.6 | 5.8–6.5 | 6.0–6.9 | 6.2–7.1 |
| Arachidic acid (C20:0) | 0.19–0.25 | 0.20–0.32 | 0.20–0.26 | 0.16–0.18 | 0.15–0.15 | 0.20–0.30 |
| Monounsaturated Fatty Acids | ||||||
| Myristoleic acid (C14:1n-5) | 0.13–0.23 | 0.19–0.25 | 0.18–0.22 | 0.11–0.13 | 0.22–0.22 | 0.21–0.21 |
| Palmitoleic acid (C16:1n-7) | 1.9–2.2 | 2.0–2.4 | 2.2–2.4 | 1.7–1.8 | 2.1–2.5 | 2.0–2.5 |
| Oleic acid (C18:1n-9) | 34.7–35.9 | 31.2–33.2 | 32.2–33.6 | 30.6–33.7 | 30.5–34.3 | 31.7–36.7 |
| Vaccenic acid (C18:1n-7) | 2.6–2.8 | 1.9–2.0 | 1.7–2.1 | 2.3–2.4 | 2.5–2.6 | 2.1–2.2 |
| Erucic acid (C22:1n-9) | 0.20–0.24 | 0.14–0.28 | 0.10–0.12 | 0.16–0.16 | 0.10–0.14 | 0.08–0.05 |
| n-3 Polyunsaturated fatty acids (n-3 LCPUFAs) | ||||||
| α-Linolenic acid (C18:3n-3) | 0.74–0.90 | 0.84–1.06 | 0.91–1.03 | 0.69–1.09 | 0.70–1.02 | 0.85–1.13 |
| Eicosapentaenoic acid (C20:5n-3) | 0.08–0.12 | 0.11–0.17 | 0.08–0.10 | 0.06–0.10 | 0.10–0.16 | 0.08–0.16 |
| Clupanodonic acid (C22:5n-3) | 0.27–0.33 | 0.19–0.25 | 0.14–0.16 | 0.30–0.34 | 0.24–0.36 | 0.16–0.24 |
| Docosahexaenoic acid (C22:6n-3) | 0.47–0.55 | 0.40–0.52 | 0.28–0.34 | 0.43–0.71 | 0.47–0.67 | 0.31–0.49 |
| n-6 Polyunsaturated fatty acids (n-6 LCPUFAs) | ||||||
| Linoleic acid (C18:2n-6) | 13.5–15.3 | 13.4–14.8 | 14.3–15.7 | 13.7–16.3 | 11.4–13.6 | 12.3–14.4 |
| γ-Linolenic acid (C18:3n-6) | 0.07–0.11 | 0.10–0.18 | 0.14–0.20 | 0.07–0.07 | 0.09–0.13 | 0.11–0.21 |
| Eicosadienoic acid (C20:2n-6) | 0.82–0.96 | 0.53–0.63 | 0.35–0.41 | 0.89–0.95 | 0.28–0.30 | 0.24–0.24 |
| Dihomo-γ-Linolenic acid (C20:3n-6) | 0.56–0.64 | 0.46–0.52 | 0.39–0.43 | 0.69–0.81 | 0.47–0.55 | 0.40–0.50 |
| Arachidonic acid (C20:4n-6) | 0.73–0.81 | 0.61–0.69 | 0.45–0.51 | 0.68–0.90 | 0.54–0.68 | 0.48–0.58 |
| Docosatetraenoic acid (C22:4n-6) | 0.29–0.39 | 0.19–0.25 | 0.09–0.11 | 0.44–0.49 | 0.22–0.22 | 0.13–0.17 |
| Adrenic acid (C22:5n-6) | 0.13–0.21 | 0.09–0.13 | 0.06–0.10 | 0.15–0.17 | 0.05–0.05 | 0.05–0.09 |
| Reference | Study Design | Population | n | Powerful and Prevalence of NEC | Principal Finding in NEC |
|---|---|---|---|---|---|
| Smithers et al. (2008) [86] | Systematic review | <37 GA | 1333 | RR = [0.62–2.04] | No benefit of n-3 LCPUFA supplemented formula |
| Zhang et al. (2014) [89] | Systemic review | <32 GA | 900 | RR = [0.23–1.10] | No benfit of n-3 LCPUFA supplementation |
| Double-blinded randomized clinical trials | |||||
| Carlson et al. (1998) [88] | Formula supplemented with 0.41% ARA + 0.13% DHA | <32 GA BW between 725–1375 g | 119 | Control = 17.6% Experimental = 2.9% | Significantly decreased |
| Fewtrell et al. (2002) [90] | Formula supplemented with 0.31% ARA + 0.17% DHA | <37 GA BW <1750 g | 197 | Control = 11% Experimental = 19% | No significant difference |
| Innis et al. (2002) [91] | BM supplemented with DHA BM supplemented with ARA + DHA | BW between 846–1560 g | 194 | Control = 1.6% Experimental = 1.5% | No significant difference |
| Fewtrell et al. (2004) [87] | Formula supplemented with 0.31% ARA + 0.17% DHA | <35 GA BW ≤2000 g | 238 | Control = 2% Experimental = 4% | No significant difference |
| Clandinin et al. (2005) [92] | Formula supplemented with DHA + ARA | <35 GA | 361 | Control = 3% Experimental = 5% | No significant difference |
| Henriksen et al. (2008) [93] | BM supplemented with 6.7% ARA + 6.9% DHA | BW <1500 g | 141 | Control = 3% Experimental = 1.5% | No significant difference |
| Makrides et al. (2009) [94] | High DHA (1%) Low DHA (0.3%) | <33 GA | 657 | Adj. OR = [0.87–5.22] | No significant difference |
| Collins et al. (2016) [95] | Formula supplemented with different doses of DHA | <30 GA | 53 | Control = 9% Experimental = 9% | No significant difference |
| Collins et al. (2017) [96] | BM supplemented with 60 mg/kg/day DHA | <29 WGA | 1273 | Adj. OR = [0.79–1.69] | No significant difference |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramiro-Cortijo, D.; Singh, P.; Liu, Y.; Medina-Morales, E.; Yakah, W.; Freedman, S.D.; Martin, C.R. Breast Milk Lipids and Fatty Acids in Regulating Neonatal Intestinal Development and Protecting against Intestinal Injury. Nutrients 2020, 12, 534. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12020534
Ramiro-Cortijo D, Singh P, Liu Y, Medina-Morales E, Yakah W, Freedman SD, Martin CR. Breast Milk Lipids and Fatty Acids in Regulating Neonatal Intestinal Development and Protecting against Intestinal Injury. Nutrients. 2020; 12(2):534. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12020534
Chicago/Turabian StyleRamiro-Cortijo, David, Pratibha Singh, Yan Liu, Esli Medina-Morales, William Yakah, Steven D. Freedman, and Camilia R. Martin. 2020. "Breast Milk Lipids and Fatty Acids in Regulating Neonatal Intestinal Development and Protecting against Intestinal Injury" Nutrients 12, no. 2: 534. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12020534