Metabolism of cis- and trans-Resveratrol and Dihydroresveratrol in an Intestinal Epithelial Model

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
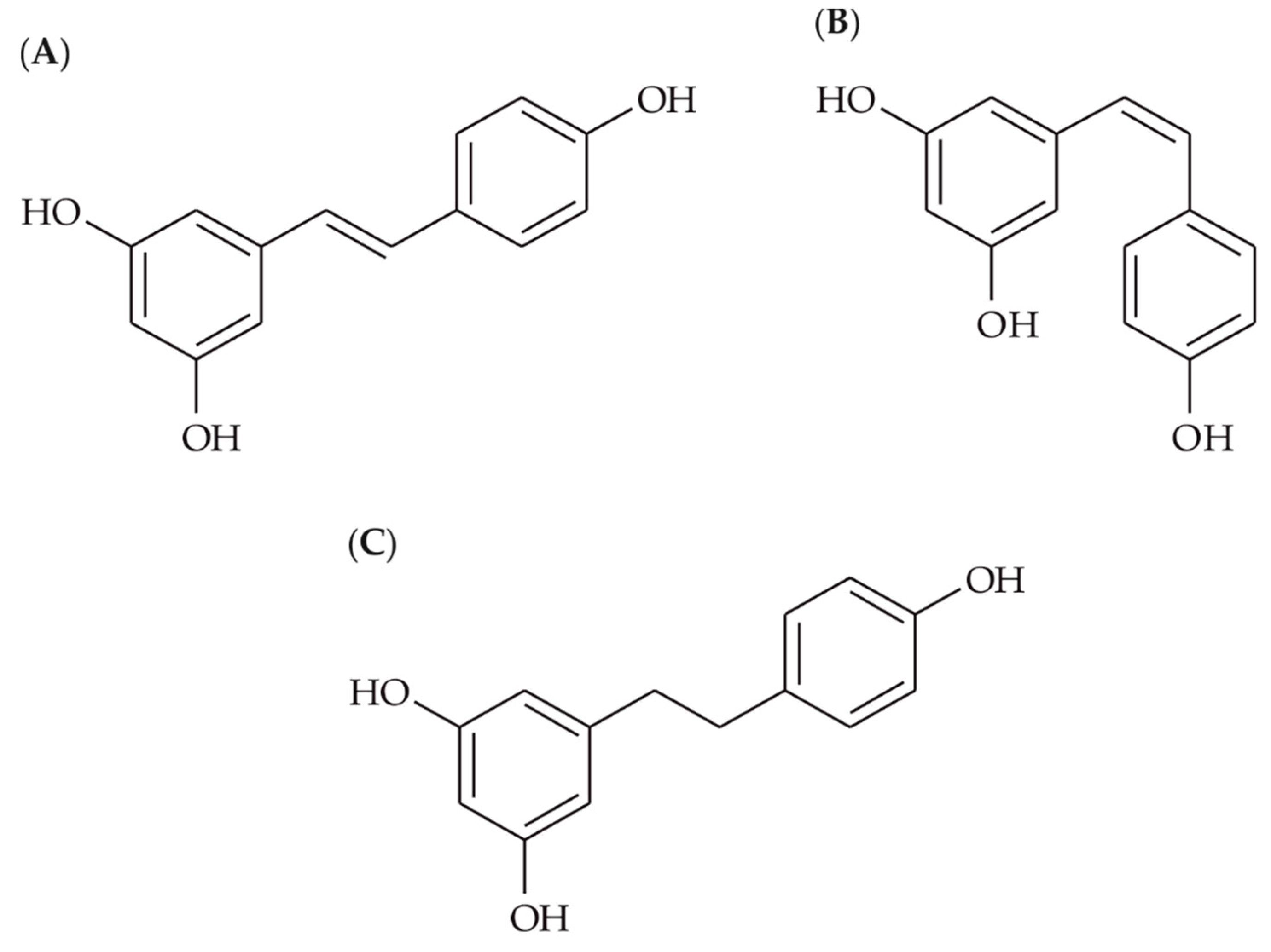
2.1. Preparation of Compounds
2.2. Cell Cultures
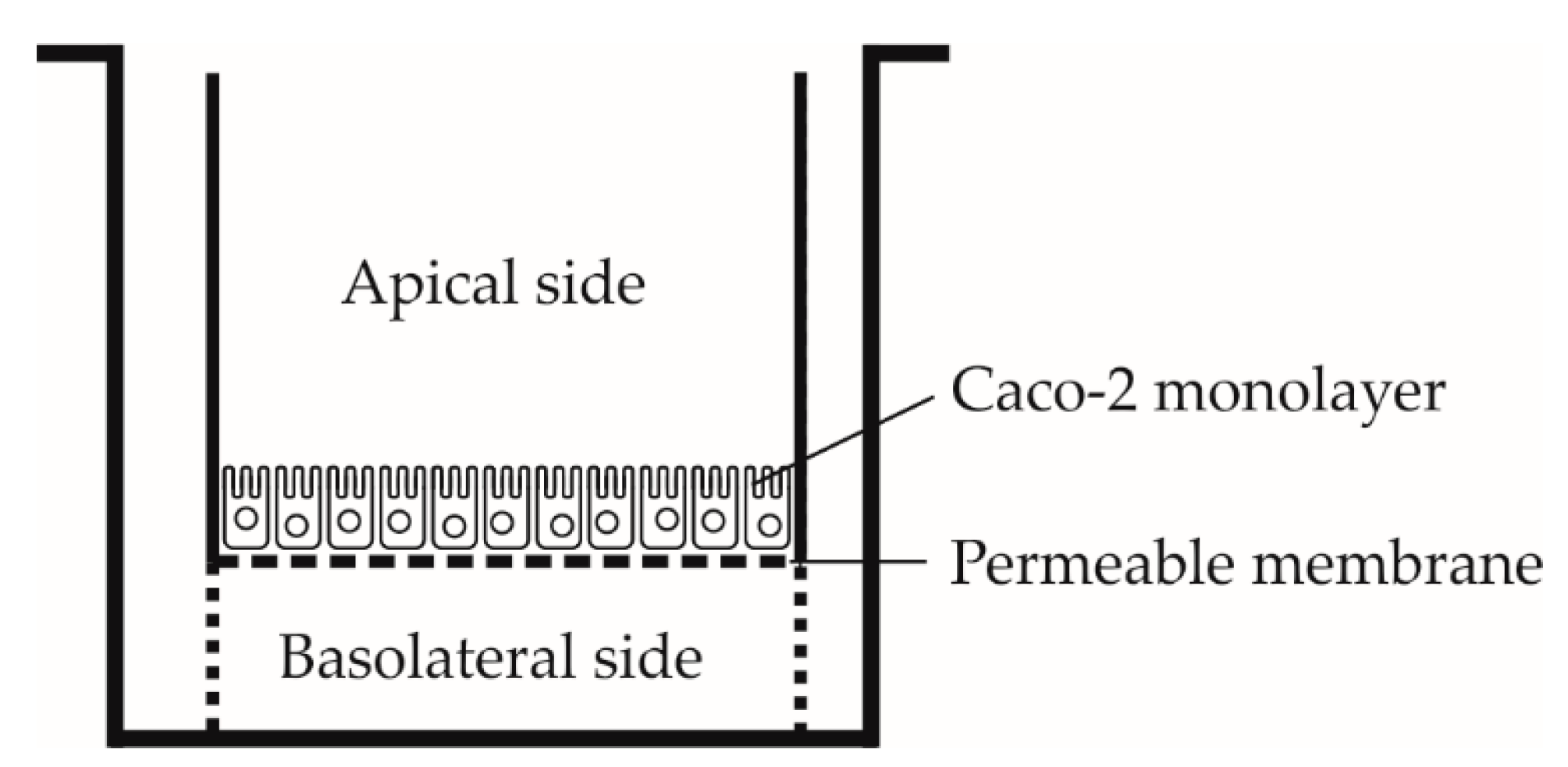
2.3. Permeability Assay
2.3.1. Preparation of Inserts with Caco-2 Cells
2.3.2. Measuring of the Monolayer Integrity
2.3.3. Metabolism and Absorption of Tested Compounds
2.4. LC/MS Analysis
2.4.1. Standards
2.4.2. Sample Purification
2.4.3. LC/MS Analysis of Metabolites
2.5. Statistical Analysis
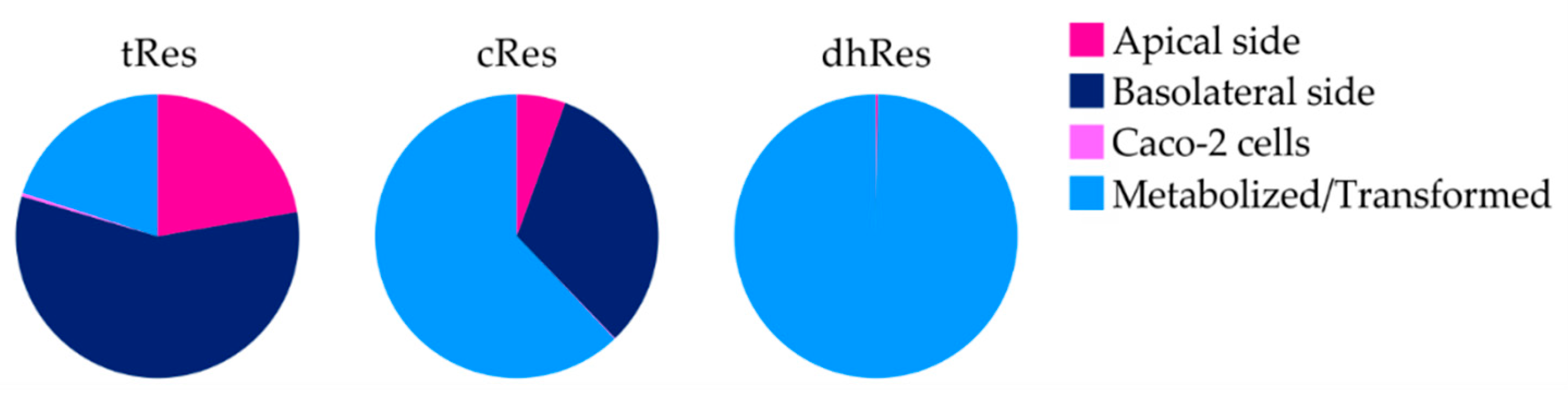
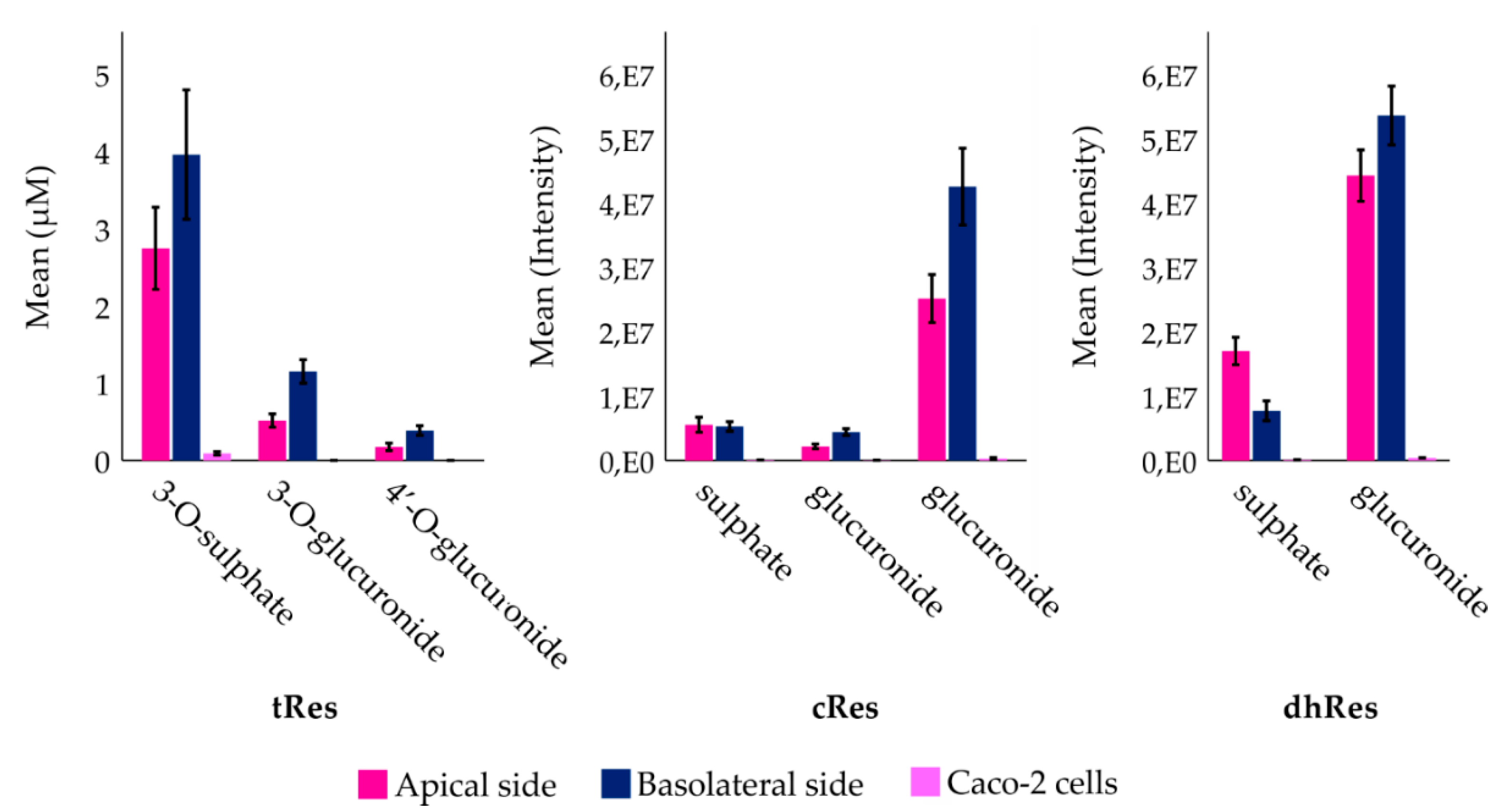
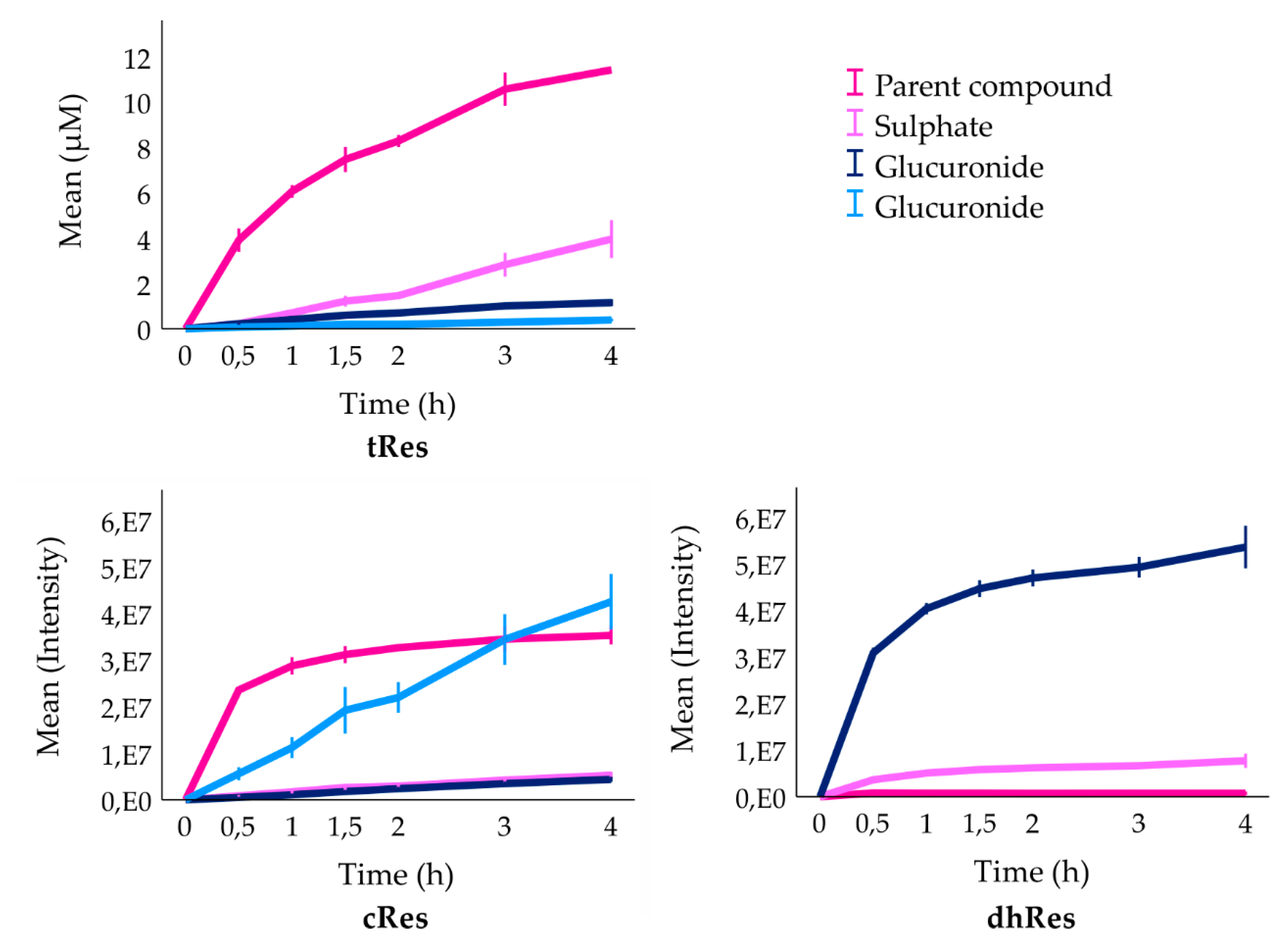
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Molecular Formula | Neutral Molecule Exact Mass: | Measured [M -H]- Exact Mass | Comparison with Standard | Detected |
|---|---|---|---|---|---|
| trans-resveratrol | C14H12O3 | 228.0786 | 227.0708 | YES | YES |
| cis-resveratrol | C14H12O3 | 228.0786 | 227.0708 | YES | YES |
| dihydroresvaratrol | C14H14O3 | 230.0943 | 229.0865 | YES | YES |
| trans-resveratrol-O-sulphate | C14H12O6S | 308.3064 | 307.0276 | YES | YES |
| cis-resveratrol-O-sulphate | C14H12O6S | 308.3064 | 307.0276 | NO | YES |
| dihydroresvaratrol-O-sulphate | C14H14O6S | 310.3223 | 309.0433 | NO | YES |
| trans-resveratrol-O-glucuronide | C20H20O9 | 404.3674 | 403.1029 | YES | YES |
| cis-resveratrol-O-glucuronide | C20H20O9 | 404.3674 | 403.1029 | NO | YES |
| dihydroresvaratrol-O-glucuronide | C20H22O9 | 406.3832 | 405.1186 | NO | YES |
| dihydroxymethylstilbene | C15H14O2 | 226.0994 | 225.0916 | NO | NO |
| methylresveratrol | C15H14O3 | 242.0943 | 241.0865 | NO | NO |
| dihydroxymethylstilbene | C15H14O2 | 212.0837 | 211.0759 | NO | NO |
| hydroxystilbene | C14H12O | 196.0888 | 195.0810 | NO | NO |
| stilbene | C14H12 | 180.0939 | 179.0861 | NO | NO |
| dihydroxymethyldihydrostilbene | C15H16O2 | 226.0994 | 225.0916 | NO | NO |
| methyl-dihydroresveratrol | C15H16O3 | 242.0943 | 241.0865 | NO | NO |
| dihydroxydihydrostilbene | C14H14O2 | 212.0837 | 211.0759 | NO | NO |
| hydroxydihydrostilbene | C14H14O | 196.0888 | 195.0810 | NO | NO |
| dihydrostilbene | C14H14 | 180.0939 | 179.0861 | NO | NO |
| phendiol | C6H6O2 | 110.0368 | 109.0290 | NO | NO |
| phenol | C6H6O | 94.0419 | 93.0340 | NO | NO |
| ethylenphenol | C8H10O | 122.0732 | 121.0653 | NO | NO |
| ethylphenol | C8H10O2 | 138.0681 | 137.0603 | NO | NO |
Appendix B
| Compound | Calibration Curve Equation | R^2 | Linear Range [ng/mL] | LOD [ng/mL] | LOQ [ng/mL] | RSD [%] of Injection Triplicate |
|---|---|---|---|---|---|---|
| tRes | Y = 1570.9X – 11845 | 0.9961 | 5-1000 | 8.2 | 9.8 | 1.68 |
| cRes | Y = 3587X – 24484 | 0.9990 | 5-500 | 7.1 | 7.8 | 0.12 |
| dhRes | Y = 612.13X – 5494.6 | 0.9906 | 5-800 | 10.4 | 13.6 | 3.15 |
| tRes-3-O-sulphate | Y = 543.14X – 8201.7 | 0.9902 | 100-600 | 17.4 | 22.6 | 2.41 |
| tRes-3-O-glucuronide | Y = 398.29X – 2733.6 | 0.9961 | 100-1000 | 9.4 | 15.4 | 2.98 |
References
- El Khawand, T.; Courtois, A.; Valls, J.; Richard, T.; Krisa, S. A review of dietary stilbenes: sources and bioavailability. Phytochem. Rev. 2018, 17, 1007–1029. [Google Scholar] [CrossRef]
- Berman, A.Y.; Motechin, R.A.; Wiesenfeld, M.Y.; Holz, M.K. The therapeutic potential of resveratrol: a review of clinical trials. NPJ Precis. Oncol. 2017, 35, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno, A.; Castro, M.; Falqué, E. Evolution of trans- and cis-resveratrol content in red grapes (Vitis vinifera L. cv Mencía, Albarello and Merenzao) during ripening. Eur. Food Res. Technol. 2008, 227, 667–674. [Google Scholar] [CrossRef]
- Renaud, S.; de Lorgeril, M. Wine, alcohol, platelets, and the French paradox for coronary heart disease. Lancet 1992, 339, 1523–1526. [Google Scholar] [CrossRef]
- Kopp, P. Resveratrol, a phytoestrogen found in red wine. A possible explanation for the conundrum of the’French paradox’? Eur. J. Endocrinol. 1998, 138, 619–620. [Google Scholar] [CrossRef] [Green Version]
- Walle, T.; Hsieh, F.; DeLegge, M.H.; Oatis, J.E.; Walle, U.K. High absorption but very low bioavailability of oral resveratrol in humans. Drug Metab. Dispos. 2004, 32, 1377–1382. [Google Scholar] [CrossRef] [Green Version]
- Wenzel, E.; Somoza, V. Metabolism and bioavailability of trans-resveratrol. Mol. Nutr. Food Res. 2005, 49, 472–481. [Google Scholar] [CrossRef]
- Jarosova, V.; Vesely, O.; Marsik, P.; Jaimes, J.D.; Smejkal, K.; Kloucek, P.; Havlik, J. Metabolism of stilbenoids by human faecal microbiota. Molecules 2019, 24, 1155. [Google Scholar] [CrossRef] [Green Version]
- Bode, L.M.; Bunzel, D.; Huch, M.; Cho, G.S.; Ruhland, D.; Bunzel, M.; Bub, A.; Franz, C.M.; Kulling, S.E. In vivo and in vitro metabolism of trans-resveratrol by human gut microbiota. Am. J. Clin. Nutr. 2013, 97, 295. [Google Scholar] [CrossRef]
- Juan, M.E.; González-Pons, E.; Planas, J.M. Multidrug Resistance Proteins Restrain the Intestinal Absorption of trans-Resveratrol in Rats. J. Nutr. 2010, 140, 489–495. [Google Scholar] [CrossRef]
- Maier-Salamon, A.; Hagenauer, B.; Wirth, M.; Gabor, F.; Szekeres, T.; Jäger, W. Increased transport of resveratrol across monolayers of the human intestinal Caco-2 cells is mediated by inhibition and saturation of metabolites. Pharm. Res. 2006, 23, 2107–2115. [Google Scholar] [CrossRef]
- Urpi-Sarda, M.; Zamora-Ros, R.; Lamuela-Raventos, R.; Cherubini, A.; Jauregui, O.; De La Torre, R.; Covas, M.I.; Estruch, R.; Jaeger, W.; Andres-Lacueva, C. HPLC-Tandem Mass Spectrometric Method to Characterize Resveratrol Metabolism in Humans. Clin. Chem. 2007, 53, 292–299. [Google Scholar] [CrossRef] [Green Version]
- Boocock, D.J.; Faust, G.E.S.; Patel, K.R.; Schinas, A.M.; Brown, V.A.; Ducharme, M.P.; Booth, T.D.; Crowell, J.A.; Perloff, M.; Gescher, A.J.; et al. Phase I Dose Escalation Pharmacokinetic Study in Healthy Volunteers of Resveratrol, a Potential Cancer Chemopreventive Agent. Cancer Epidemiol. Biomarkers Prev. 2007, 16, 1246–1252. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Yi, L.; Jin, X.; Xie, Q.; Zhang, T.; Zhou, X.; Chang, H.; Fu, Y.; Zhu, J.; Zhang, Q.; et al. Absorption of resveratrol by vascular endothelial cells through passive diffusion and an SGLT1-mediated pathway. J. Nutr. Biochem. 2013, 24, 1823–1829. [Google Scholar] [CrossRef]
- Maier-Salamon, A.; Böhmdorfer, M.; Riha, J.; Thalhammer, T.; Szekeres, T.; Jaeger, W. Interplay between metabolism and transport of resveratrol. Ann. N. Y. Acad. Sci. 2013, 1290, 98–106. [Google Scholar] [CrossRef]
- Lagouge, M.; Argmann, C.; Gerhart-Hines, Z.; Meziane, H.; Lerin, C.; Daussin, F.; Messadeq, N.; Milne, J.; Lambert, P.; Elliott, P.; et al. Resveratrol Improves Mitochondrial Function and Protects against Metabolic Disease by Activating SIRT1 and PGC-1α. Cell 2006, 127, 1109–1122. [Google Scholar] [CrossRef]
- Timmers, S.; Konings, E.; Bilet, L.; Houtkooper, R.H.; van de Weijer, T.; Goossens, G.H.; Hoeks, J.; van der Krieken, S.; Ryu, D.; Kersten, S.; et al. Calorie Restriction-like Effects of 30 Days of Resveratrol Supplementation on Energy Metabolism and Metabolic Profile in Obese Humans. Cell Metab. 2011, 14, 612–622. [Google Scholar] [CrossRef] [Green Version]
- Falomir, E.; Lucas, R.; Peñalver, P.; Martí-Centelles, R.; Dupont, A.; Zafra-Gómez, A.; Carda, M.; Morales, J.C. Cytotoxic, Antiangiogenic and Antitelomerase Activity of Glucosyl- and Acyl- Resveratrol Prodrugs and Resveratrol Sulfate Metabolites. ChemBioChem 2016, 17, 1343–1348. [Google Scholar] [CrossRef] [Green Version]
- Storniolo, C.E.; Quifer-Rada, P.; Lamuela-Raventos, R.M.; Moreno, J.J. Piceid presents antiproliferative effects in intestinal epithelial Caco-2 cells, effects unrelated to resveratrol release. Food Funct. 2014, 5, 2137–2144. [Google Scholar] [CrossRef]
- Su, D.; Cheng, Y.; Liu, M.; Liu, D.; Cui, H.; Zhang, B.; Zhou, S.; Yang, T.; Mei, Q. Comparision of Piceid and Resveratrol in Antioxidation and Antiproliferation Activities In Vitro. PLoS ONE 2013, 8, e54505. [Google Scholar] [CrossRef] [Green Version]
- Jarosova, V.; Doskocil, I.; Volstatova, T.; Havlik, J. Adhesive Property of Different Strains of Lactobacilli in the Presence of Resveratrol. Sci. Agric. Bohem. 2018, 49, 291–296. [Google Scholar] [CrossRef] [Green Version]
- Hubatsch, I.; Ragnarsson, E.G.E.; Artursson, P. Determination of drug permeability and prediction of drug absorption in Caco-2 monolayers. Nat. Protoc. 2007, 2, 2111–2119. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Shin, Y.; Yu, C.; Kosmeder, J.; Hirschelman, W.; Pezzuto, J.; van Breemen, R. Increasing the Throughput and Productivity of Caco-2 Cell Permeability Assays Using Liquid Chromatography-Mass Spectrometry: Application to Resveratrol Absorption and Metabolism. Comb. Chem. High Throughput Screen. 2012, 6, 757–767. [Google Scholar] [CrossRef] [PubMed]
- Henry, C.; Vitrac, X.; Decendit, A.; Ennamany, R.; Krisa, S.; Mérillon, J.M. Cellular uptake and efflux of trans-piceid and its aglycone trans-resveratrol on the apical membrane of human intestinal Caco-2 cells. J. Agric. Food Chem. 2005, 53, 798–803. [Google Scholar] [CrossRef] [PubMed]
- Kaldas, M.I.; Walle, U.K.; Walle, T. Resveratrol transport and metabolism by human intestinal Caco-2 cells. J. Pharm. Pharmacol. 2003, 55, 307–312. [Google Scholar] [CrossRef]
- Kuhnle, G.; Spencer, J.P.E.; Chowrimootoo, G.; Schroeter, H.; Debnam, E.S.; Srai, S.K.S.; Rice-Evans, C.; Hahn, U. Resveratrol is absorbed in the small intestine as resveratrol glucuronide. Biochem. Biophys. Res. Commun. 2000, 272, 212–217. [Google Scholar] [CrossRef]
- Azorín-Ortuño, M.; Yáñez-Gascón, M.J.; Vallejo, F.; Pallarés, F.J.; Larrosa, M.; Lucas, R.; Morales, J.C.; Tomás-Barberán, F.A.; García-Conesa, M.T.; Espín, J.C. Metabolites and tissue distribution of resveratrol in the pig. Mol. Nutr. Food Res. 2011, 55, 1154–1168. [Google Scholar] [CrossRef] [Green Version]
- Sabolovic, N.; Humbert, A.C.; Radominska-Pandya, A.; Magdalou, J. Resveratrol is efficiently glucuronidated by UDP-glucuronosyltransferases in the human gastrointestinal tract and in Caco-2 cells. Biopharm. Drug Dispos. 2006, 27, 181–189. [Google Scholar] [CrossRef]
- Juan, M.E.; Alfaras, I.; Planas, J.M. Determination of dihydroresveratrol in rat plasma by HPLC. J. Agric. Food Chem. 2010, 58, 7472–7475. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jarosova, V.; Vesely, O.; Doskocil, I.; Tomisova, K.; Marsik, P.; Jaimes, J.D.; Smejkal, K.; Kloucek, P.; Havlik, J. Metabolism of cis- and trans-Resveratrol and Dihydroresveratrol in an Intestinal Epithelial Model. Nutrients 2020, 12, 595. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12030595
Jarosova V, Vesely O, Doskocil I, Tomisova K, Marsik P, Jaimes JD, Smejkal K, Kloucek P, Havlik J. Metabolism of cis- and trans-Resveratrol and Dihydroresveratrol in an Intestinal Epithelial Model. Nutrients. 2020; 12(3):595. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12030595
Chicago/Turabian StyleJarosova, Veronika, Ondrej Vesely, Ivo Doskocil, Katerina Tomisova, Petr Marsik, Jose D. Jaimes, Karel Smejkal, Pavel Kloucek, and Jaroslav Havlik. 2020. "Metabolism of cis- and trans-Resveratrol and Dihydroresveratrol in an Intestinal Epithelial Model" Nutrients 12, no. 3: 595. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12030595