Phycobiliproteins from Arthrospira Platensis (Spirulina): A New Source of Peptides with Dipeptidyl Peptidase-IV Inhibitory Activity




{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Microalgae Biomass
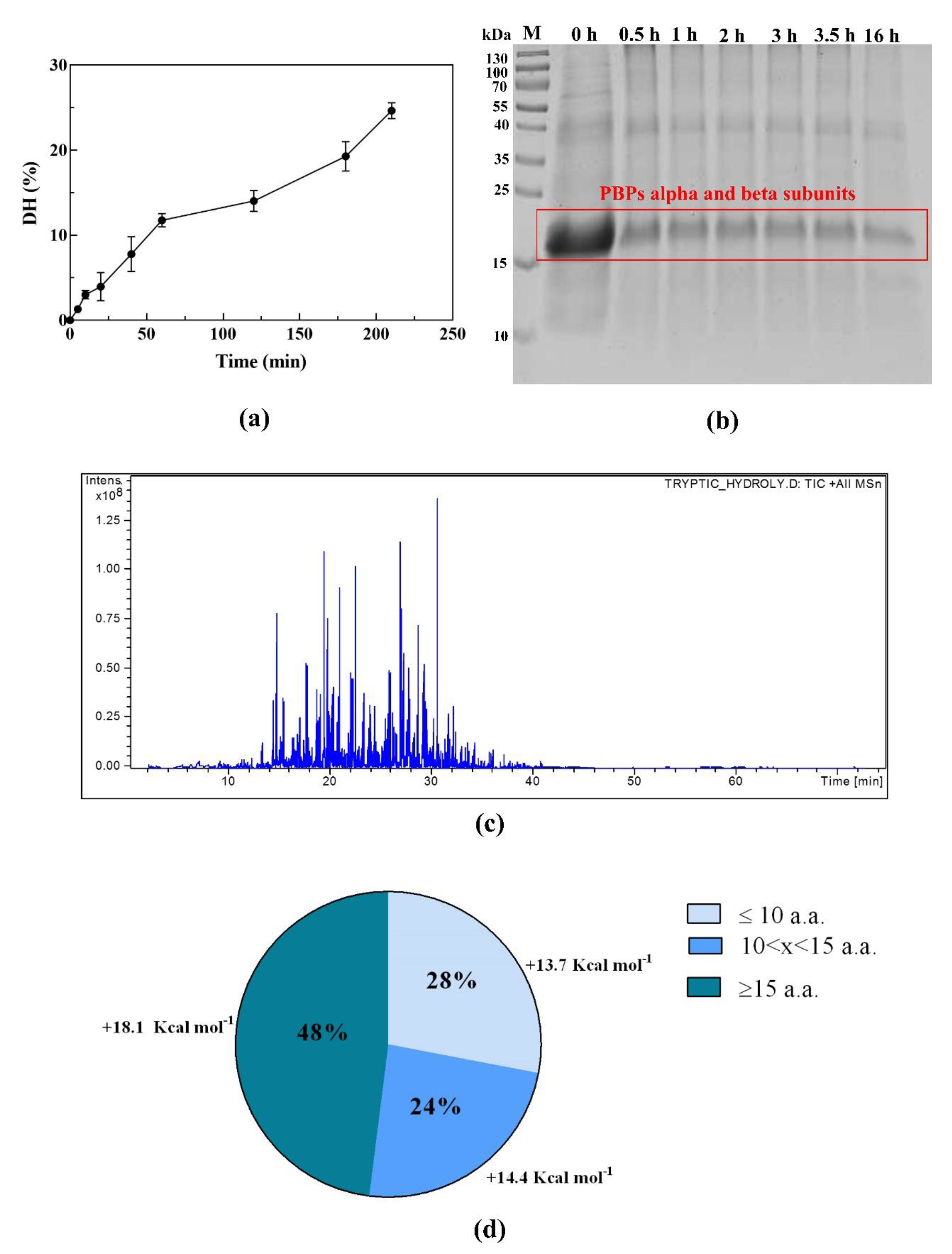
2.3. Enzymatic Hydrolysis of PBP
2.4. Analysis of the Hydrolysate by LC-ESI-MS/MS
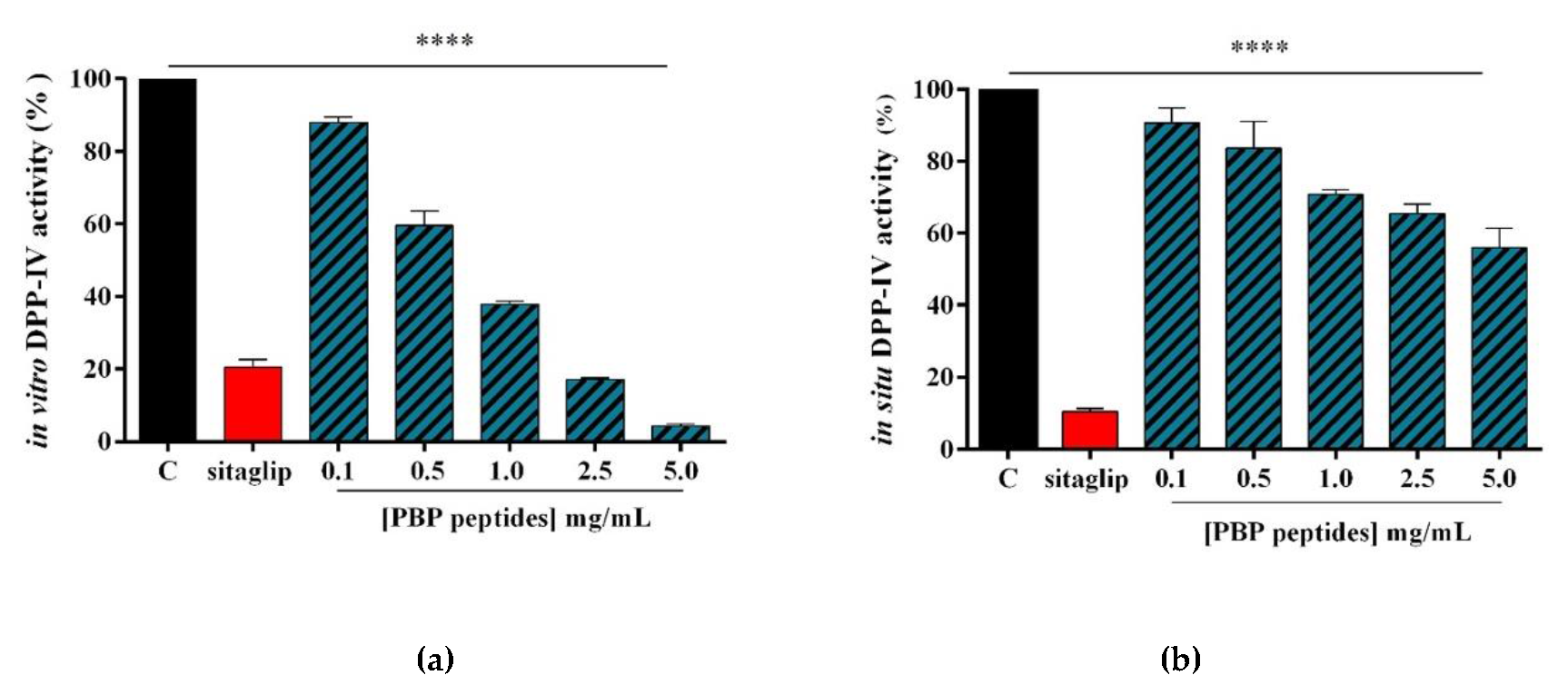
2.5. In Vitro DPP-IV Activity of Tryptic PBP Peptides
2.6. Cell Culture
2.7. MTT Assay
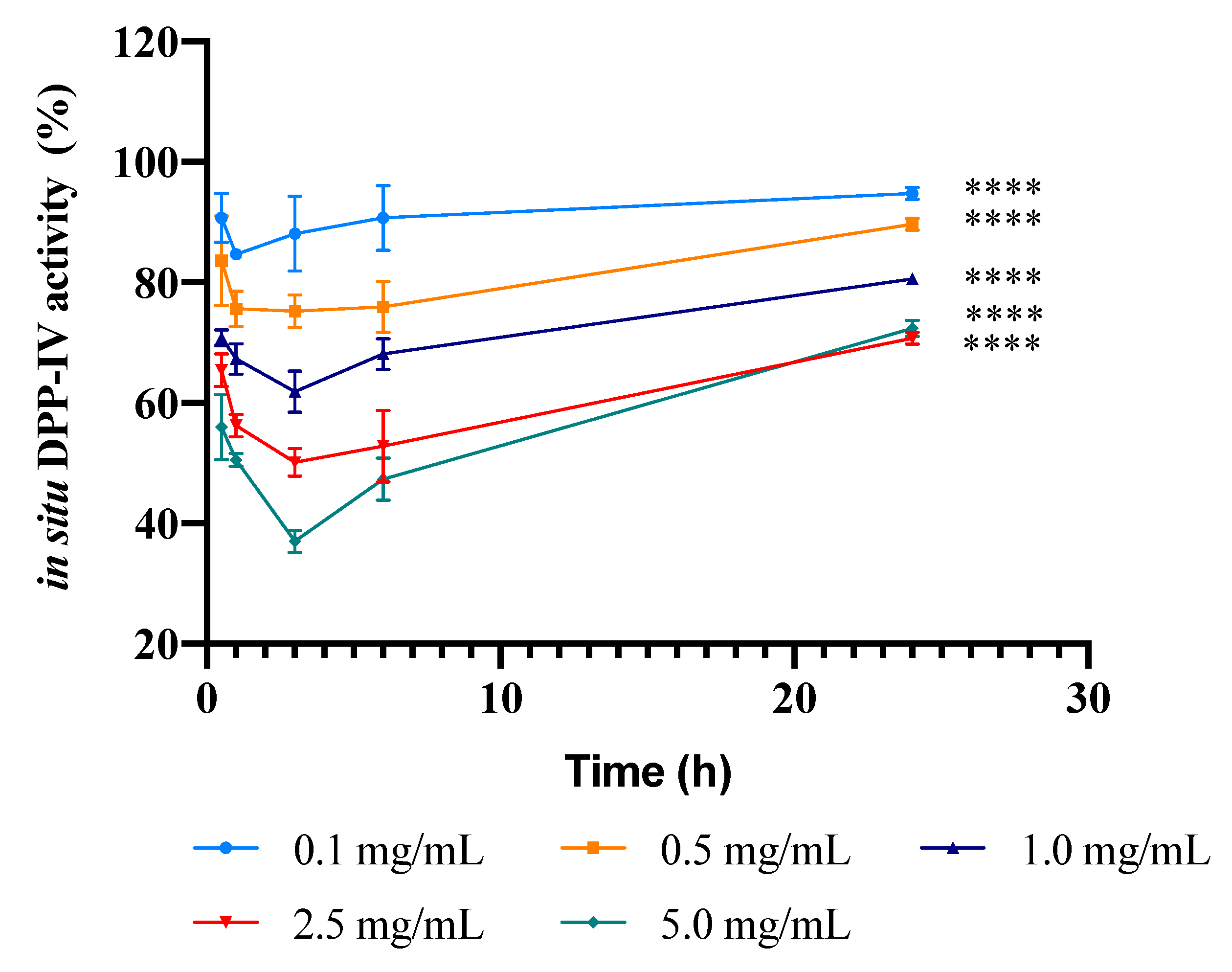
2.8. Evaluation of the Inhibitory Effect of PBP Peptides on the In Situ DPP-IV Activity Expressed by Caco-2 Cells
2.9. Statistical Analysis
3. Results
3.1. PBP In Vitro Digestion and Peptide Identification by Nano ESI-MS/MS
3.2. PBP Peptides Inhibit DPP-IV Activity In Vitro and at the Cellular Level
3.3. Kinetics of the Inhibition of DPP-IV Activity Expressed by Caco-2 Cells Induced by the PBP Hydrolysate
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ACE | angiotensin converting enzyme |
| ACN | acetonitrile |
| AMC | 7-Amido-4-methylcoumarin |
| APC | allophycocyanin |
| BSA | bovine serum albumin |
| C-PC | C-phycocyanin |
| DH | degree of hydrolysis |
| DMEM | Dulbecco Minimum Essential Medium |
| DMSO | dimethyl sulfoxide |
| DPP-IV | dipeptidyl peptidase IV |
| FBS | fetal bovine serum |
| GIP | insulinotropic peptide |
| GLP-1 | glucagon-like peptide |
| HMGCoAR | 3-hydroxy-3-methyl-glutaryl-coenzyme A reductase |
| HPLC-ESI-MS/MS | High-performance liquid chromatography electrospray ionization tandem mass spectrometry |
| INS-1 β-cells | rat insulinoma beta cells |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| MW | molecular weight |
| OPA | o-phthaldialdehyde |
| PBP | phycobiliproteins |
| PBS | phosphate buffered saline |
| SDS-PAGE | sodium dodecyl sulphate-polyacrylamide gel electrophoresis |
| T2D | type 2 diabetes |
| TIC | total ion chromatogram |
| UF | ultrafiltration |
References
- Patil, G.; Chethana, S.; Madhusudhan, M.; Raghavarao, K. Fractionation and purification of the phycobiliproteins from Spirulina platensis. Bioresour. Technol. 2008, 99, 7393–7396. [Google Scholar] [CrossRef] [PubMed]
- Glazer, A.N. Phycobiliproteins—A family of valuable, widely used fluorophores. Environ. Boil. Fishes 1994, 6, 105–112. [Google Scholar] [CrossRef]
- Yoshida, A.; Takagaki, Y.; Nishimune, T. Enzyme Immunoassay for Phycocyanin as the Main Component of Spirulina Color in Foods. Biosci. Biotechnol. Biochem. 1996, 60, 57–60. [Google Scholar] [CrossRef] [PubMed]
- Bhat, V.B.; Madyastha, K.M. Scavenging of peroxynitrite by phycocyanin and phycocyanobilin from Spirulina platensis: Protection against oxidative damage to DNA. Biochem. Biophys. Res. Commun. 2001, 285, 262–266. [Google Scholar] [CrossRef]
- Furuta, T.; Miyabe, Y.; Yasui, H.; Kinoshita, Y.; Kishimura, H. Angiotensin I Converting Enzyme Inhibitory Peptides Derived from Phycobiliproteins of Dulse Palmaria palmata. Mar. Drugs 2016, 14, 32. [Google Scholar] [CrossRef] [Green Version]
- Cermeno, M.; Stack, J.; Tobin, P.R.; O’Keeffe, M.; Harnedy, P.A.; Stengel, D.; Fitzgerald, R.J. Peptide identification from a Porphyra dioica protein hydrolysate with antioxidant, angiotensin converting enzyme and dipeptidyl peptidase IV inhibitory activities. Food Funct. 2019, 10, 3421–3429. [Google Scholar] [CrossRef]
- Cherng, S.-C.; Cheng, S.-N.; Tarn, A.; Chou, T.-C. Anti-inflammatory activity of c-phycocyanin in lipopolysaccharide-stimulated RAW 264.7 macrophages. Life Sci. 2007, 81, 1431–1435. [Google Scholar] [CrossRef]
- Rimbau, V.; Camins, A.; Pubill, D.; Sureda, F.X.; Romay, C.; González, R.; Jiménez, A.; Escubedo, E.; Camarasa, J.; Pallàs, M. C-phycocyanin protects cerebellar granule cells from low potassium/serum deprivation-induced apoptosis. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2001, 364, 96–104. [Google Scholar] [CrossRef]
- Estrada, J.P. Antioxidant activity of different fractions of Spirulina platensis protean extract. Il Farm. 2001, 56, 497–500. [Google Scholar] [CrossRef]
- Nagaoka, S.; Shimizu, K.; Kaneko, H.; Shibayama, F.; Morikawa, K.; Kanamaru, Y.; Otsuka, A.; Hirahashi, T.; Kato, T. A Novel Protein C-Phycocyanin Plays a Crucial Role in the Hypocholesterolemic Action of Spirulina platensis Concentrate in Rats. J. Nutr. 2005, 135, 2425–2430. [Google Scholar] [CrossRef] [Green Version]
- Ou, Y.; Lin, L.; Pan, Q.; Yang, X.; Cheng, X. Preventive effect of phycocyanin from Spirulina platensis on alloxan-injured mice. Environ. Toxicol. Pharmacol. 2012, 34, 721–726. [Google Scholar] [CrossRef]
- Ou, Y.; Lin, L.; Yang, X.; Pan, Q.; Cheng, X. Antidiabetic potential of phycocyanin: Effects on KKAy mice. Pharm. Boil. 2013, 51, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Ou, Y.; Ren, Z.; Wang, J.; Yang, X. Phycocyanin ameliorates alloxan-induced diabetes mellitus in mice: Involved in insulin signaling pathway and GK expression. Chem. Interact. 2016, 247, 49–54. [Google Scholar] [CrossRef]
- Gao, Y.N.; Liao, G.Y.; Xiang, C.X.; Yang, X.G.; Cheng, X.D.; Ou, Y. Effects of phycocyanin on INS-1 pancreatic beta-cell mediated by PI3K/Akt/FoxO1 signaling pathway. Int. J. Biol. Macromol. 2016, 83, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Nauck, M.A.; Baller, B.; Meier, J.J. Gastric inhibitory polypeptide and glucagon-like peptide-1 in the pathogenesis of type 2 diabetes. Diabetes 2004, 53, S190–S196. [Google Scholar] [CrossRef] [Green Version]
- Lacroix, I.M.E.; Li-Chan, E.C. Food-derived dipeptidyl-peptidase IV inhibitors as a potential approach for glycemic regulation—Current knowledge and future research considerations. Trends Food Sci. Technol. 2016, 54, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Nongonierma, A.B.; Fitzgerald, R.J. Structure activity relationship modelling of milk protein-derived peptides with dipeptidyl peptidase IV (DPP-IV) inhibitory activity. Peptides 2016, 79, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aiello, G.; Li, Y.; Arnoldi, A.; Bollati, C.; Arnoldi, A.; Lammi, C. Chemical and biological characterization of spirulina protein hydrolysates: Focus on ACE and DPP-IV activities modulation. J. Funct. Foods 2019, 63, 103592. [Google Scholar] [CrossRef]
- Howell, S.; Brewis, I.A.; Hooper, N.M.; Kenny, A.; Turner, A.J. Mosaic expression of membrane peptidases by confluent cultures of Caco-2 cells. FEBS Lett. 1993, 317, 109–112. [Google Scholar] [CrossRef] [Green Version]
- Aiello, G.; Lammi, C.; Boschin, G.; Zanoni, C.; Arnoldi, A. Exploration of Potentially Bioactive Peptides Generated from the Enzymatic Hydrolysis of Hempseed Proteins. J. Agric. Food Chem. 2017, 65, 10174–10184. [Google Scholar] [CrossRef]
- Natoli, M.; Leoni, B.D.; D’Agnano, I.; D’Onofrio, M.; Brandi, R.; Arisi, I.; Zucco, F.; Felsani, A. Cell growing density affects the structural and functional properties of Caco-2 differentiated monolayer. J. Cell. Physiol. 2011, 226, 1531–1543. [Google Scholar] [CrossRef] [PubMed]
- Lammi, C.; Zanoni, C.; Scigliuolo, G.M.; D’Amato, A.; Arnoldi, A. Lupin Peptides Lower Low-Density Lipoprotein (LDL) Cholesterol through an Up-regulation of the LDL Receptor/Sterol Regulatory Element Binding Protein 2 (SREBP2) Pathway at HepG2 Cell Line. J. Agric. Food Chem. 2014, 62, 7151–7159. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Acquah, C.; Aluko, R.E.; Udenigwe, C.C. Considering food matrix and gastrointestinal effects in enhancing bioactive peptide absorption and bioavailability. J. Funct. Foods 2020, 64, 103680. [Google Scholar] [CrossRef]
- Lammi, C.; Arnoldi, A.; Aiello, G. Soybean Peptides Exert Multifunctional Bioactivity Modulating 3-Hydroxy-3-Methylglutaryl-CoA Reductase and Dipeptidyl Peptidase-IV Targets in Vitro. J. Agric. Food Chem. 2019, 67, 4824–4830. [Google Scholar] [CrossRef]
- Lammi, C.; Bollati, C.; Gelain, F.; Arnoldi, A.; Pugliese, R. Enhancement of the Stability and Anti-DPPIV Activity of Hempseed Hydrolysates through Self-Assembling Peptide-Based Hydrogels. Front. Chem. 2019, 6. [Google Scholar] [CrossRef] [Green Version]
- Power, O.; Nongonierma, A.B.; Jakeman, P.M.; Fitzgerald, R.J. Food protein hydrolysates as a source of dipeptidyl peptidase IV inhibitory peptides for the management of type 2 diabetes. Proc. Nutr. Soc. 2013, 73, 34–46. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Chen, R.; Zuo, F.; Ma, H.; Zhang, Y.; Chen, S. Comparison of dipeptidyl peptidase IV-inhibitory activity of peptides from bovine and caprine milk casein by in silico and in vitro analyses. Int. Dairy J. 2016, 53, 37–44. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; Mazzocchi, C.; Paolella, S.; Fitzgerald, R.J. Release of dipeptidyl peptidase IV (DPP-IV) inhibitory peptides from milk protein isolate (MPI) during enzymatic hydrolysis. Food Res. Int. 2017, 94, 79–89. [Google Scholar] [CrossRef] [Green Version]
- Nongonierma, A.B.; Paolella, S.; Mudgil, P.; Maqsood, S.; Fitzgerald, R.J. Dipeptidyl peptidase IV (DPP-IV) inhibitory properties of camel milk protein hydrolysates generated with trypsin. J. Funct. Foods 2017, 34, 49–58. [Google Scholar] [CrossRef] [Green Version]
- Lammi, C.; Bollati, C.; Ferruzza, S.; Ranaldi, G.; Sambuy, Y.; Arnoldi, A. Soybean- and Lupin-Derived Peptides Inhibit DPP-IV Activity on In Situ Human Intestinal Caco-2 Cells and Ex Vivo Human Serum. Nutrients 2018, 10, 1082. [Google Scholar] [CrossRef] [Green Version]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Aiello, G.; Bollati, C.; Bartolomei, M.; Arnoldi, A.; Lammi, C. Phycobiliproteins from Arthrospira Platensis (Spirulina): A New Source of Peptides with Dipeptidyl Peptidase-IV Inhibitory Activity. Nutrients 2020, 12, 794. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12030794
Li Y, Aiello G, Bollati C, Bartolomei M, Arnoldi A, Lammi C. Phycobiliproteins from Arthrospira Platensis (Spirulina): A New Source of Peptides with Dipeptidyl Peptidase-IV Inhibitory Activity. Nutrients. 2020; 12(3):794. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12030794
Chicago/Turabian StyleLi, Yuchen, Gilda Aiello, Carlotta Bollati, Martina Bartolomei, Anna Arnoldi, and Carmen Lammi. 2020. "Phycobiliproteins from Arthrospira Platensis (Spirulina): A New Source of Peptides with Dipeptidyl Peptidase-IV Inhibitory Activity" Nutrients 12, no. 3: 794. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12030794