Low Levels of Serum Tryptophan Underlie Skeletal Muscle Atrophy

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Serum Tryptophan and Kynurenine Measurements
2.3. Muscle Mass Measurements
2.4. Materials and Cell Culture Conditions
2.5. Cell Proliferation Assay
2.6. Cell Differentiation and Myotube Formation Assay
2.7. Animal Experiments
2.8. Metabolome Analysis
2.9. Statistics
3. Results
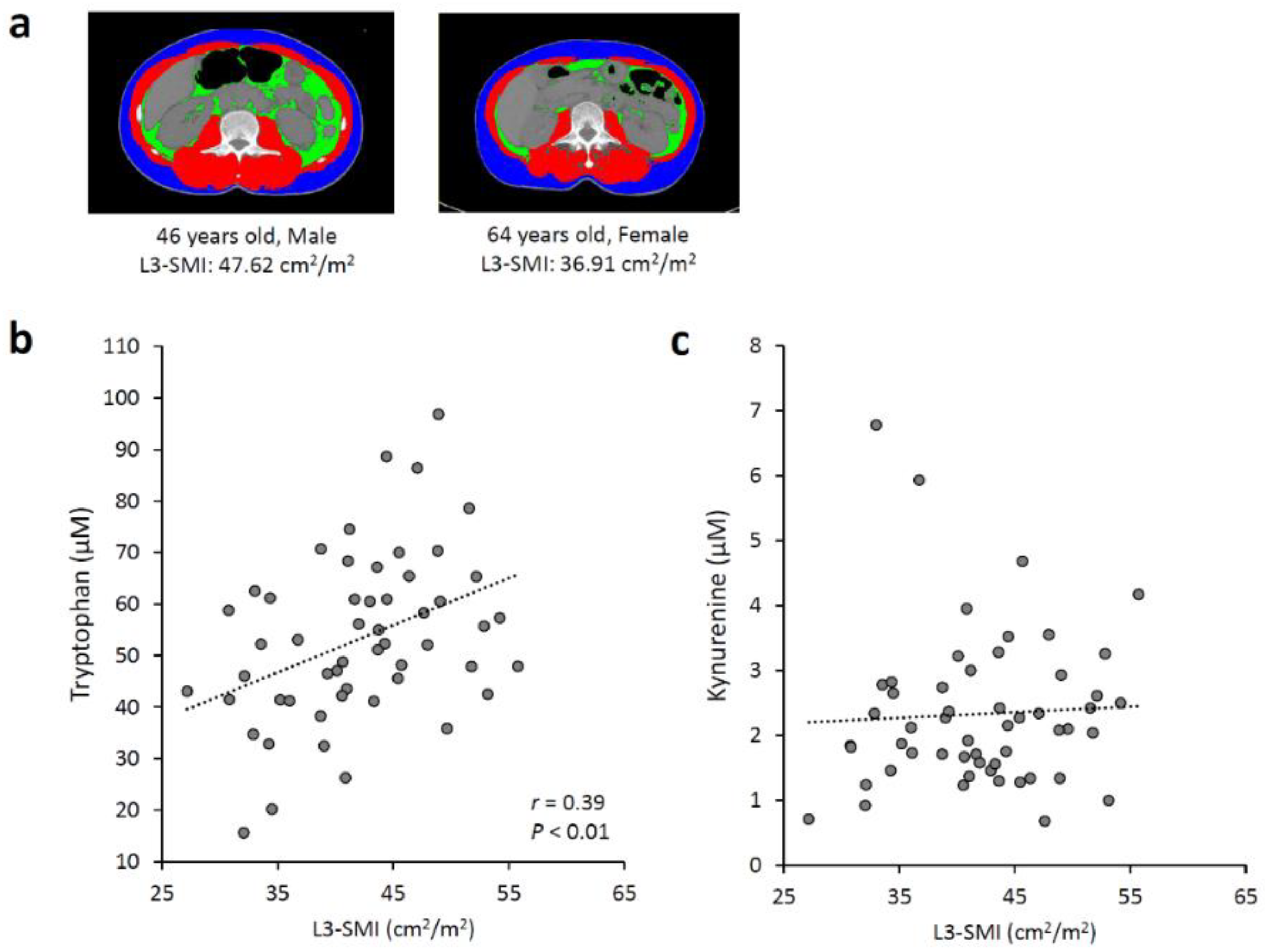
3.1. Associations between Serum Tryptophan and Kynurenine Levels and Skeletal Muscle Mass
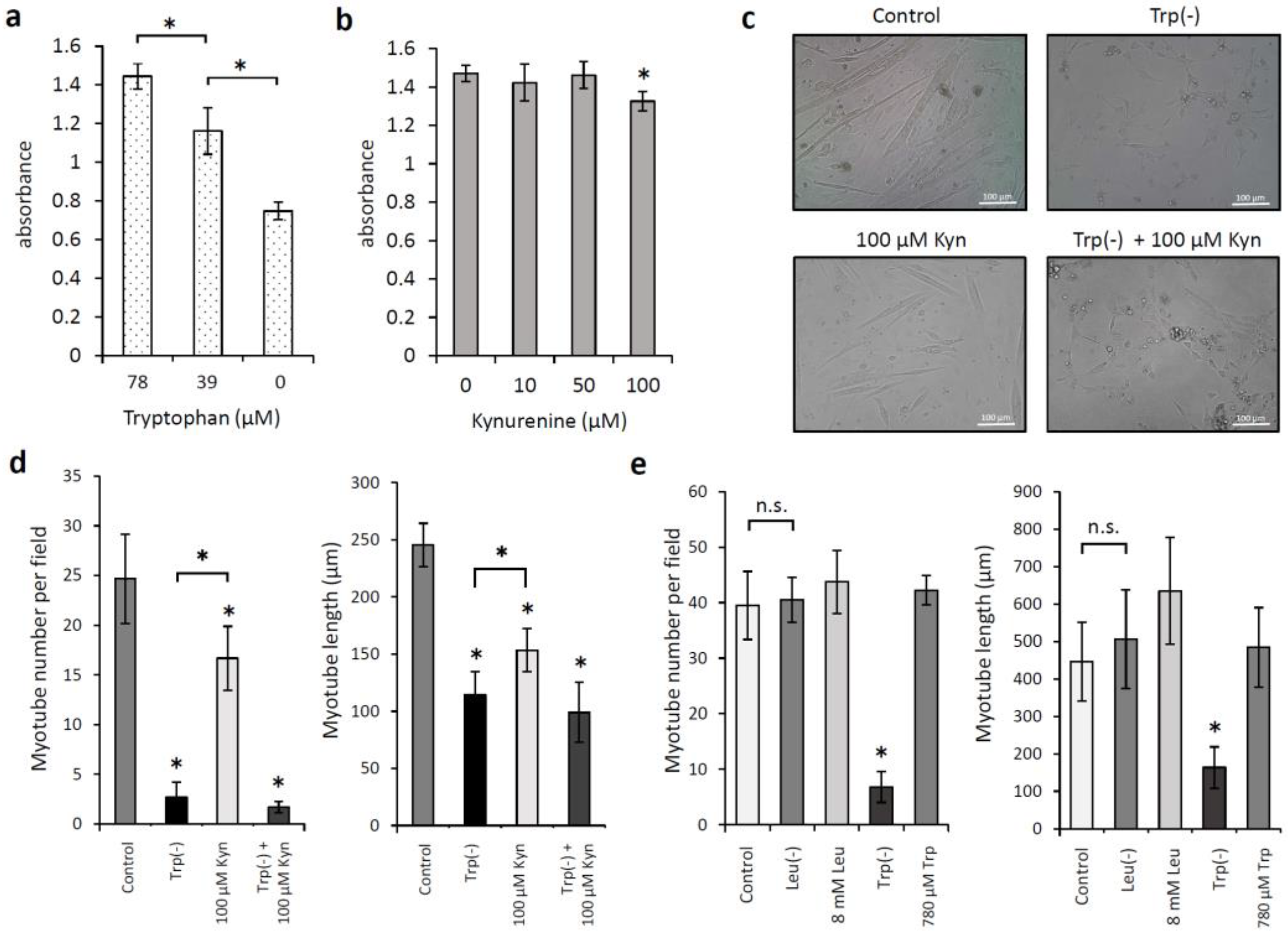
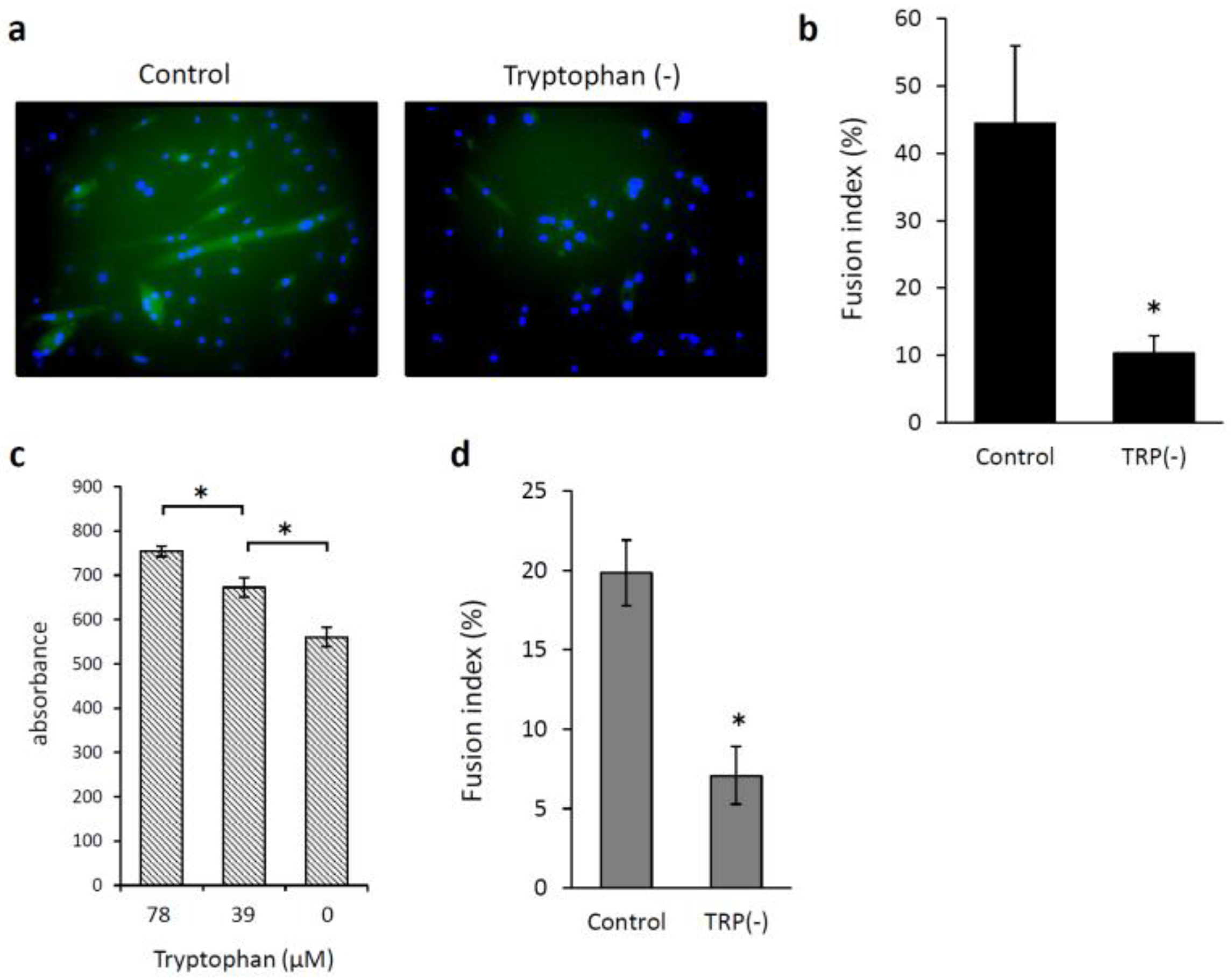
3.2. Effects of Low Levels of Tryptophan and Kynurenine on the Proliferation of and Myotube Formation by Skeletal Muscle Myoblast Cells
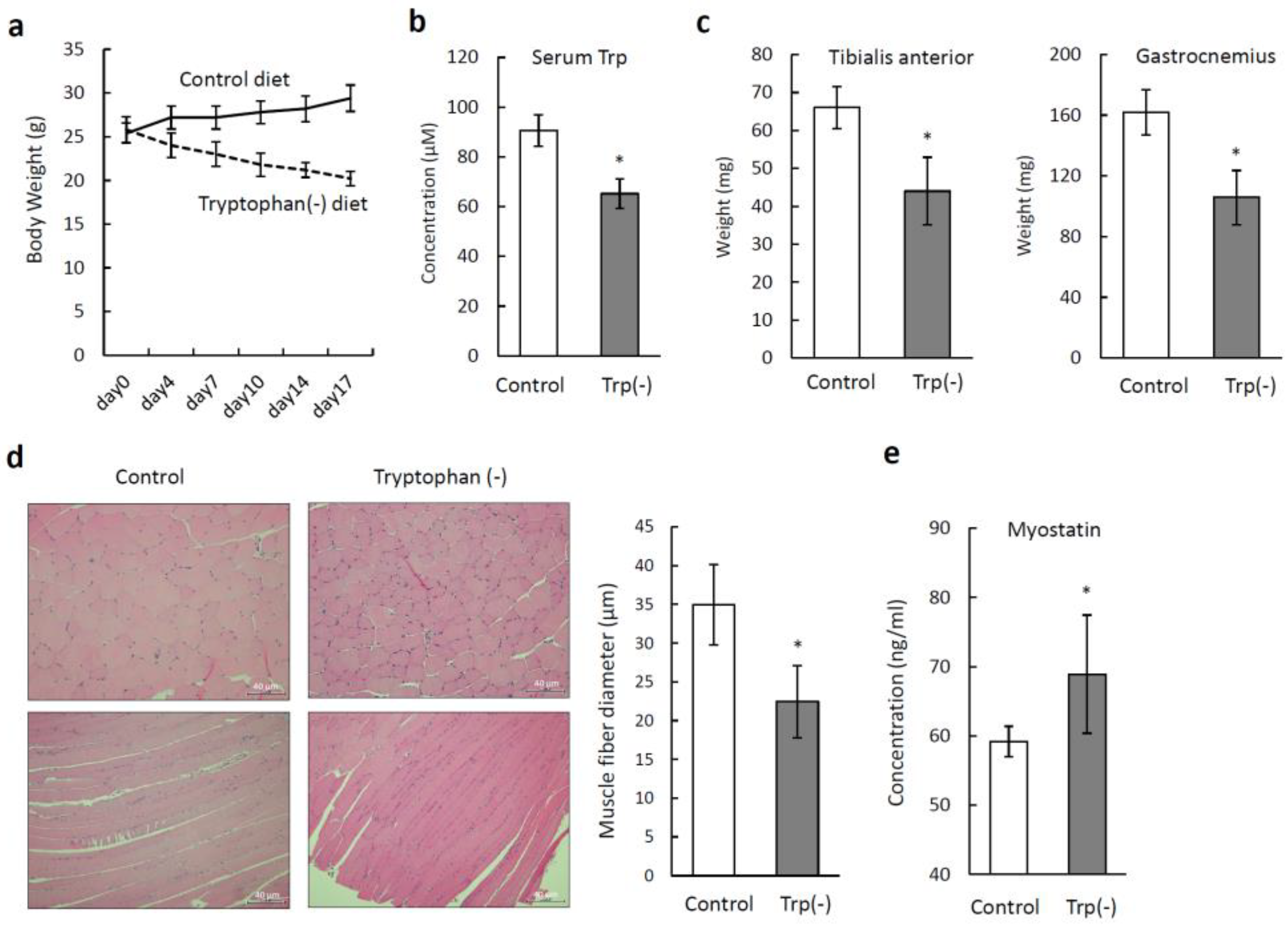
3.3. Tryptophan-Deficient Diet Induces Skeletal Muscle Atrophy in Mice
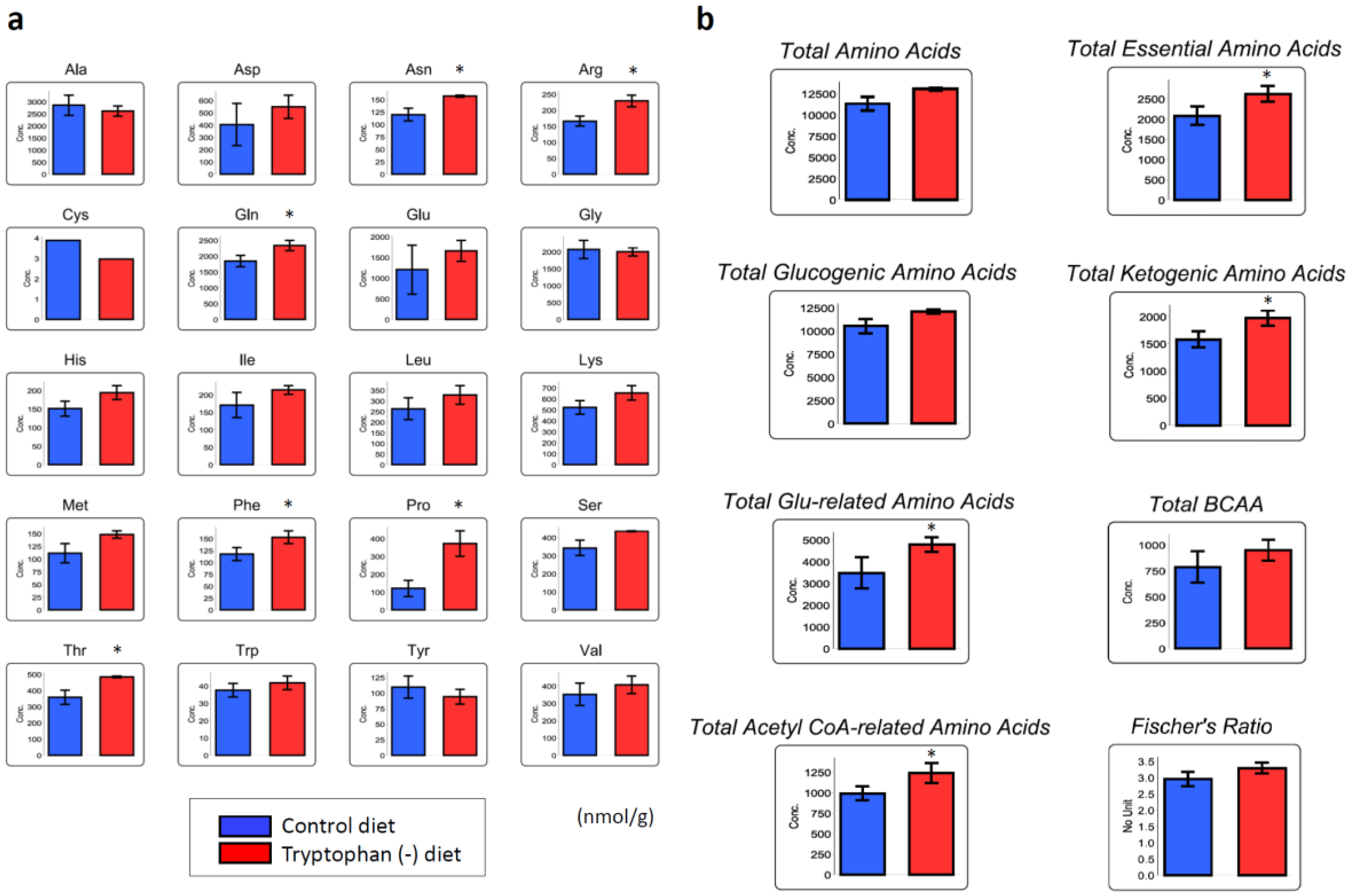
3.4. Tryptophan-Deficient Diet Changes the Amino Acid Profile in Skeletal Muscles
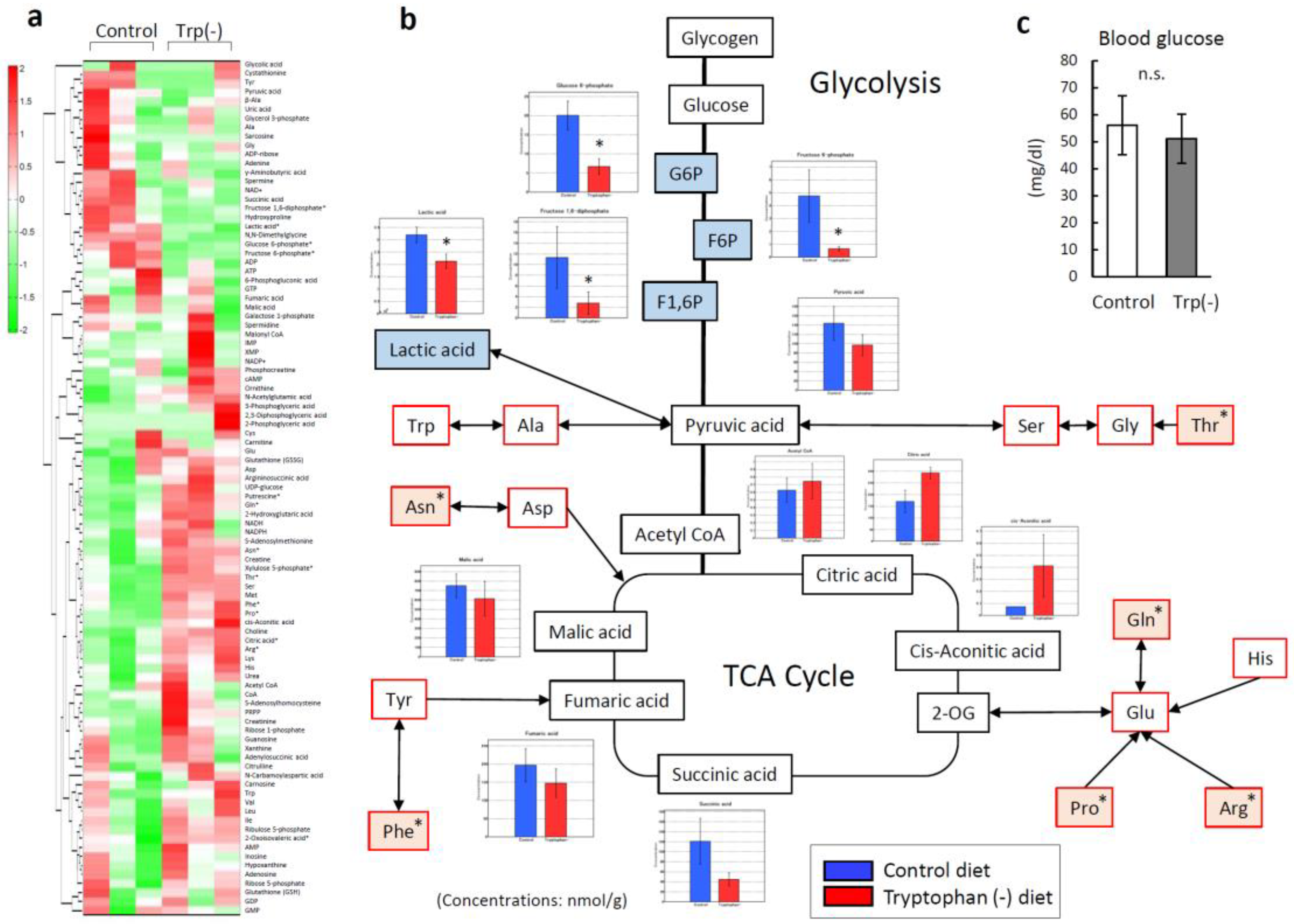
3.5. Serum Tryptophan Deficiency Induces Metabolic Alterations in Skeletal Muscles
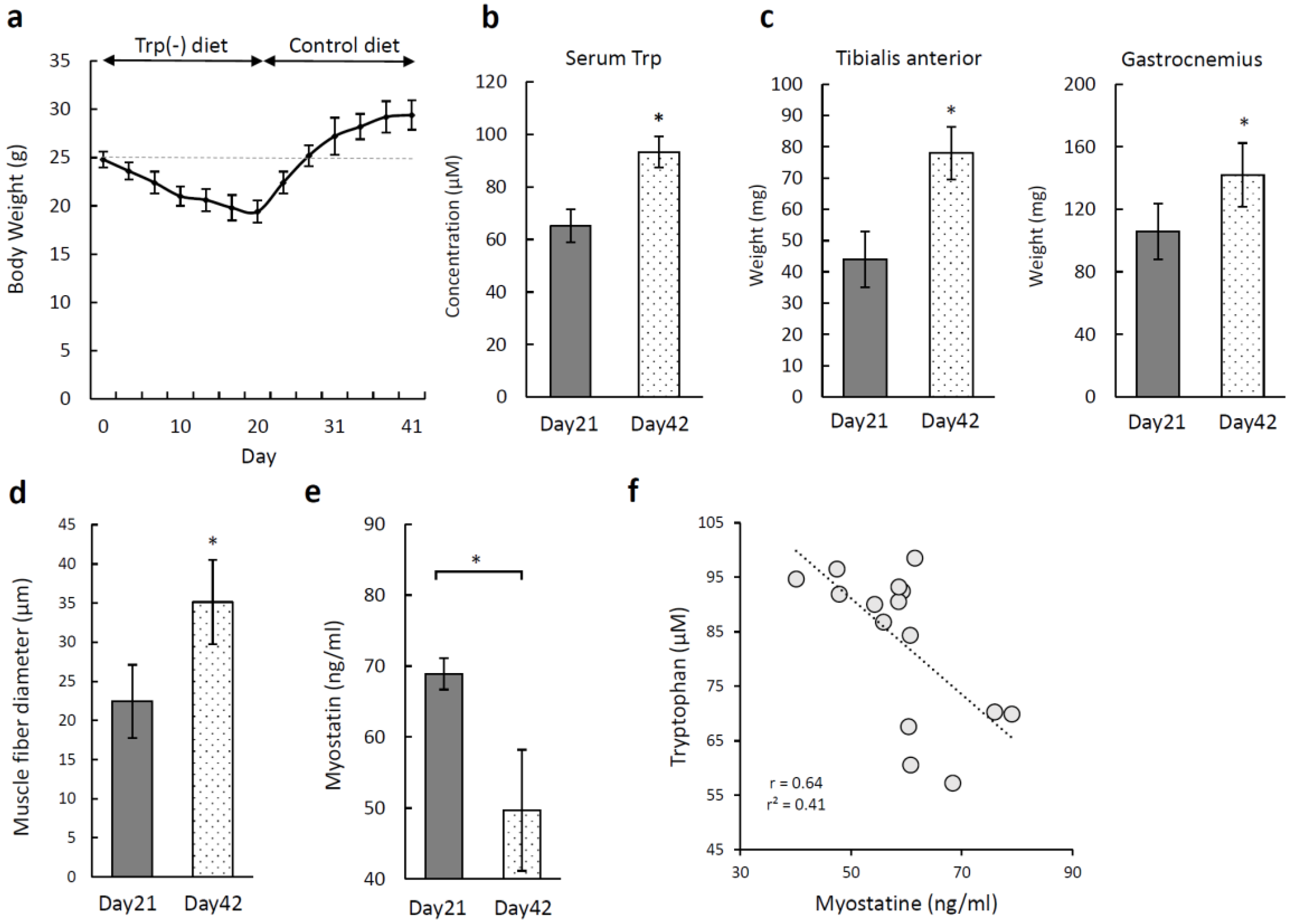
3.6. Recovery of Skeletal Muscle Atrophy Induced by a Tryptophan-Deficient Diet
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pamoukdjian, F.; Bouillet, T.; Lévy, V.; Soussan, M.; Zelek, L.; Paillaud, E. Prevalence and predictive value of pre-therapeutic sarcopenia in cancer patients: A systematic review. Clin. Nutr. 2017, 37, 1101–1113. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, N.; Hara, T.; Shibata, Y.; Matsumoto, T.; Nakamura, H.; Ninomiya, S.; Kito, Y.; Kitagawa, J.; Kanemura, N.; Goto, N.; et al. Sarcopenia is an independent prognostic factor in male patients with diffuse large B-cell lymphoma. Ann. Hematol. 2015, 94, 2043–2053. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, N.; Ninomiya, S.; Matsumoto, T.; Nakamura, H.; Kitagawa, J.; Shiraki, M.; Hara, T.; Shimizu, M.; Tsurumi, H. Prognostic impact of skeletal muscle assessed by computed tomography in patients with acute myeloid leukemia. Ann. Hematol. 2019, 98, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Takashina, C.; Tsujino, I.; Watanabe, T.; Sakaue, S.; Ikeda, D.; Yamada, A.; Sato, T.; Ohira, H.; Otsuka, Y.; Oyama-Manabe, N.; et al. Associations among the plasma amino acid profile, obesity, and glucose metabolism in Japanese adults with normal glucose tolerance. Nutr. Metab. 2016, 13, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Capri, M.; Martin, F.-P.J.; Montoliu, I.; DaSilva, L.; Salvioli, S.; Monti, D.; Kussmann, M.; Bucci, L.; Rezzi, S.; Biagi, E.; et al. Serum profiling of healthy aging identifies phospho- and sphingolipid species as markers of human longevity. Aging (Albany NY) 2016, 6, 9–25. [Google Scholar]
- Lai, H.S.; Lee, J.C.; Lee, P.H.; Wang, S.T.; Chen, W.J. Plasma free amino acid profile in cancer patients. Semin. Cancer Biol. 2005, 15, 267–276. [Google Scholar] [CrossRef]
- Toyoshima, K.; Nakamura, M.; Adachi, Y.; Imaizumi, A.; Hakamada, T.; Abe, Y.; Kaneko, E.; Takahashi, S.; Shimokado, K. Increased plasma proline concentrations are associated with sarcopenia in the elderly. PLoS ONE 2017, 12, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Miyagi, Y.; Higashiyama, M.; Gochi, A.; Akaike, M.; Ishikawa, T.; Miura, T.; Saruki, N.; Bando, E.; Kimura, H.; Imamura, F.; et al. Plasma free amino acid profiling of five types of cancer patients and its application for early detection. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [Green Version]
- Stone, T.W.; Stoy, N.; Darlington, L.G. An expanding range of targets for kynurenine metabolites of tryptophan. Trends Pharmacol. Sci. 2013, 34, 136–143. [Google Scholar] [CrossRef] [Green Version]
- Cervenka, I.; Agudelo, L.Z.; Ruas, J.L. Kynurenines: Tryptophan’s metabolites in exercise, inflammation, and mental health. Science 2017, 357. [Google Scholar] [CrossRef] [Green Version]
- Munn, D.H.; Mellor, A.L. IDO in the Tumor Microenvironment: Inflammation, Counter-Regulation, and Tolerance. Trends Immunol. 2016, 37, 193–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prendergast, G.C.; Mondal, A.; Dey, S.; Laury-Kleintop, L.D.; Muller, A.J. Inflammatory Reprogramming with IDO1 Inhibitors: Turning Immunologically Unresponsive “Cold” Tumors “Hot”. Trends Cancer 2018, 4, 38–58. [Google Scholar] [CrossRef] [PubMed]
- Ninomiya, S.; Hara, T.; Tsurumi, H.; Hoshi, M.; Kanemura, N.; Goto, N.; Kasahara, S.; Shimizu, M.; Ito, H.; Saito, K.; et al. Indoleamine 2,3-dioxygenase in tumor tissue indicates prognosis in patients with diffuse large B-cell lymphoma treated with R-CHOP. Ann. Hematol. 2011, 90, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Fukuno, K.; Hara, T.; Tsurumi, H.; Shibata, Y.; Mabuchi, R.; Nakamura, N.; Kitagawa, J.; Shimizu, M.; Ito, H.; Saito, K.; et al. Expression of indoleamine 2,3-dioxygenase in leukemic cells indicates an unfavorable prognosis in acute myeloid leukemia patients with intermediate-risk cytogenetics. Leuk. Lymphoma 2015, 56, 1398–1405. [Google Scholar] [CrossRef]
- Hoshi, M.; Ito, H.; Fujigaki, H.; Takemura, M.; Takahashi, T.; Tomita, E.; Ohyama, M.; Tanaka, R.; Saito, K.; Seishima, M. Indoleamine 2,3-dioxygenase is highly expressed in human adult T-cell leukemia/lymphoma and chemotherapy changes tryptophan catabolism in serum and reduced activity. Leuk. Res. 2009, 33, 39–45. [Google Scholar] [CrossRef]
- Lanic, H.; Kraut-Tauzia, J.; Modzelewski, R.; Clatot, F.; Mareschal, S.; Picquenot, J.M.; Stamatoullas, A.; Leprêtre, S.; Tilly, H.; Jardin, F. Sarcopenia is an independent prognostic factor in elderly patients with diffuse large B-cell lymphoma treated with immunochemotherapy. Leuk. Lymphoma 2014, 55, 817–823. [Google Scholar] [CrossRef]
- Uyttenhove, C.; Pilotte, L.; Théate, I.; Stroobant, V.; Colau, D.; Parmentier, N.; Boon, T.; Van den Eynde, B.J. Evidence for a tumoral immune resistance mechanism based on tryptophan degradation by indoleamine 2,3-dioxygenase. Nat. Med. 2003, 9, 1269–1274. [Google Scholar] [CrossRef]
- Komrokji, R.S.; Wei, S.; Mailloux, A.W.; Zhang, L.; Padron, E.; Sallman, D.; Lancet, J.E.; Tinsley, S.; Nardelli, L.A.; Pinilla-Ibarz, J.; et al. A phase II study to determine the safety and efficacy of the oral inhibitor of indoleamine 2,3-dioxygenase (IDO) enzyme INCB024360 in patients with myelodysplastic syndromes. Clin. Lymphoma Myeloma Leuk. 2019, 19, 157–161. [Google Scholar] [CrossRef]
- Löb, S.; Königsrainer, A.; Rammensee, H.G.; Opelz, G.; Terness, P. Inhibitors of indoleamine-2,3-dioxygenase for cancer therapy: Can we see the wood for the trees? Nat. Rev. Cancer 2009, 9, 445–452. [Google Scholar] [CrossRef]
- Brochez, L.; Chevolet, I.; Kruse, V. The rationale of indoleamine 2,3-dioxygenase inhibition for cancer therapy. Eur. J. Cancer 2017, 76, 167–182. [Google Scholar] [CrossRef]
- Moro, T.; Ebert, S.M.; Adams, C.M.; Rasmussen, B.B. Amino Acid Sensing in Skeletal Muscle. Trends Endocrinol. Metab. 2016, 27, 796–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, Y.; Sato, Y.; Obeng, K.A.; Yoshizawa, F. Acute oral administration of L-leucine upregulates slow-fiber– and mitochondria-related genes in skeletal muscle of rats. Nutr. Res. 2018, 57, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Atherton, P.J.; Smith, K.; Etheridge, T.; Rankin, D.; Rennie, M.J. Distinct anabolic signalling responses to amino acids in C2C12 skeletal muscle cells. Amino Acids 2010, 38, 1533–1539. [Google Scholar] [CrossRef]
- Dideriksen, K.; Reitelseder, S.; Holm, L. Influence of amino acids, dietary protein, and physical activity on muscle mass development in humans. Nutrients 2013, 5, 852–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imbesi, R.; Castrogiovanni, P. Embryonic and postnatal development in experimental tryptophan deprived rats. A preliminary study. J. Mol. Histol. 2008, 39, 487–498. [Google Scholar] [CrossRef]
- Lee, H.; Ohno, M.; Ohta, S.; Mikami, T. Regular Moderate or Intense Exercise Prevents Depression-Like Behavior without Change of Hippocampal Tryptophan Content in Chronically Tryptophan-Deficient and Stressed Mice. PLoS ONE 2013, 8, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.N.; Ha, S.H.; Kim, J.; Koh, A.; Lee, C.S.; Kim, J.H.; Jeon, H.; Kim, D.-H.; Suh, P.-G.; Ryu, S.H. Glycolytic flux signals to mTOR through glyceraldehyde-3-phosphate dehydrogenase-mediated regulation of Rheb. Mol. Cell. Biol. 2009, 29, 3991–4001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, J.; Vernus, B.; Chelh, I.; Cassar-Malek, I.; Gabillard, J.C.; Hadj Sassi, A.; Seiliez, I.; Picard, B.; Bonnieu, A. Myostatin and the skeletal muscle atrophy and hypertrophy signaling pathways. Cell. Mol. Life Sci. 2014, 71, 4361–4371. [Google Scholar] [CrossRef]
- Díaz, B.B.; González, D.A.; Gannar, F.; Pérez, M.C.R.; de León, A.C. Myokines, physical activity, insulin resistance and autoimmune diseases. Immunol. Lett. 2018, 203, 1–5. [Google Scholar] [CrossRef]
- Dukes, A.; Davis, C.; El Refaey, M.; Upadhyay, S.; Mork, S.; Arounleut, P.; Johnson, M.H.; Hill, W.D.; Isales, C.M.; Hamrick, M.W. The aromatic amino acid tryptophan stimulates skeletal muscle IGF1/p70s6k/mTor signaling in vivo and the expression of myogenic genes in vitro. Nutrition 2015, 31, 1018–1024. [Google Scholar] [CrossRef] [Green Version]
- Kanungo, S.; Wells, K.; Tribett, T.; El-Gharbawy, A. Glycogen metabolism and glycogen storage disorders. Ann. Transl. Med. 2018, 6, 474. [Google Scholar] [CrossRef] [PubMed]
- Eleftheriadis, T.; Pissas, G.; Liakopoulos, V.; Stefanidis, I. IDO decreases glycolysis and glutaminolysis by activating GCN2K, while it increases fatty acid oxidation by activating AhR, thus preserving CD4+ T-cell survival and proliferation. Int. J. Mol. Med. 2018, 42, 557–568. [Google Scholar] [CrossRef] [PubMed]
- Cosgrove, B.D.; Gilbert, P.M.; Porpiglia, E.; Mourkioti, F.; Lee, S.P.; Corbel, S.Y.; Llewellyn, M.E.; Delp, S.L.; Blau, H.M. Rejuvenation of the muscle stem cell population restores strength to injured aged muscles. Nat. Med. 2014, 20, 255–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murach, K.A.; Fry, C.S.; Kirby, T.J.; Jackson, J.R.; Lee, J.D.; White, S.H.; Dupont-Versteegden, E.E.; McCarthy, J.J.; Peterson, C.A. Starring or Supporting Role? Satellite Cells and Skeletal Muscle Fiber Size Regulation. Physiology 2017, 33, 26–38. [Google Scholar] [CrossRef]
- Platten, M.; Nollen, E.A.A.; Röhrig, U.F.; Fallarino, F.; Opitz, C.A. Tryptophan metabolism as a common therapeutic target in cancer, neurodegeneration and beyond. Nat. Rev. Drug Discov. 2019, 18, 379–401. [Google Scholar] [CrossRef]
- Musumeci, G.; Imbesi, R.; Trovato, F.M.; Szychlinska, M.A.; Aiello, F.C.; Buffa, P.; Castrogiovanni, P. Importance of serotonin (5-HT) and its precursor L-tryptophan for homeostasis and function of skeletal muscle in rats. A morphological and endocrinological study. Acta Histochem. 2015, 117, 267–274. [Google Scholar] [CrossRef]
- Shen, W. Total body skeletal muscle and adipose tissue volumes: Estimation from a single abdominal cross-sectional image. J. Appl. Physiol. 2004, 97, 2333–2338. [Google Scholar] [CrossRef] [Green Version]
- Pamoukdjian, F.; Paillaud, E.; Zelek, L.; Laurent, M.; Lévy, V.; Landre, T.; Sebbane, G. Measurement of gait speed in older adults to identify complications associated with frailty: A systematic review. J. Geriatr. Oncol. 2015, 6, 484–496. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | No. | Serum Tryptophan (µ M) | |||
|---|---|---|---|---|---|
| Median | Range | P-Value | |||
| All patients | 54 | 58.17 | 15.63–96.83 | ||
| Sex | male | 34 | 56.13 | 15.63–96.83 | <0.01 |
| female | 20 | 43.06 | 20.19–70.71 | ||
| Age (years) | <60 | 14 | 55.71 | 26.3–86.41 | 0.33 |
| ≥60 | 40 | 51.16 | 15.63–96.83 | ||
| PS | 0–1 | 42 | 52.09 | 15.63–86.41 | 0.28 |
| 2–4 | 12 | 52.25 | 26.3–96.83 | ||
| LDH | normal | 24 | 54.99 | 15.63–96.83 | 0.22 |
| increased | 30 | 48.8 | 26.3–86.41 | ||
| Extranodal sites | 0–1 | 40 | 51.62 | 15.63–86.41 | 0.30 |
| ≥2 | 14 | 53.62 | 26.3–96.83 | ||
| CS | I/II | 25 | 56.13 | 15.63–78.58 | 0.18 |
| III/IV | 29 | 47.93 | 26.3–96.83 | ||
| B symptoms | Absent | 36 | 56.13 | 20.19–96.83 | <0.05 |
| Present | 18 | 43.56 | 15.63–74.55 | ||
| L3-SMI (cm2/m2) | <41 | 27 | 42.64 | 15.63–70.71 | <0.01 |
| ≥41 | 27 | 59.42 | 35.85–96.83 | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ninomiya, S.; Nakamura, N.; Nakamura, H.; Mizutani, T.; Kaneda, Y.; Yamaguchi, K.; Matsumoto, T.; Kitagawa, J.; Kanemura, N.; Shiraki, M.; et al. Low Levels of Serum Tryptophan Underlie Skeletal Muscle Atrophy. Nutrients 2020, 12, 978. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12040978
Ninomiya S, Nakamura N, Nakamura H, Mizutani T, Kaneda Y, Yamaguchi K, Matsumoto T, Kitagawa J, Kanemura N, Shiraki M, et al. Low Levels of Serum Tryptophan Underlie Skeletal Muscle Atrophy. Nutrients. 2020; 12(4):978. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12040978
Chicago/Turabian StyleNinomiya, Soranobu, Nobuhiko Nakamura, Hiroshi Nakamura, Taku Mizutani, Yuto Kaneda, Kimihiro Yamaguchi, Takuro Matsumoto, Junichi Kitagawa, Nobuhiro Kanemura, Makoto Shiraki, and et al. 2020. "Low Levels of Serum Tryptophan Underlie Skeletal Muscle Atrophy" Nutrients 12, no. 4: 978. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12040978