Sodium Imbalance in Mice Results Primarily in Compensatory Gene Regulatory Responses in Kidney and Colon, but Not in Taste Tissue


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Experiments
2.2. Feeding Experiment and Amiloride Intervention
2.3. Short-term Preference Tests
2.4. Tissue Preparation
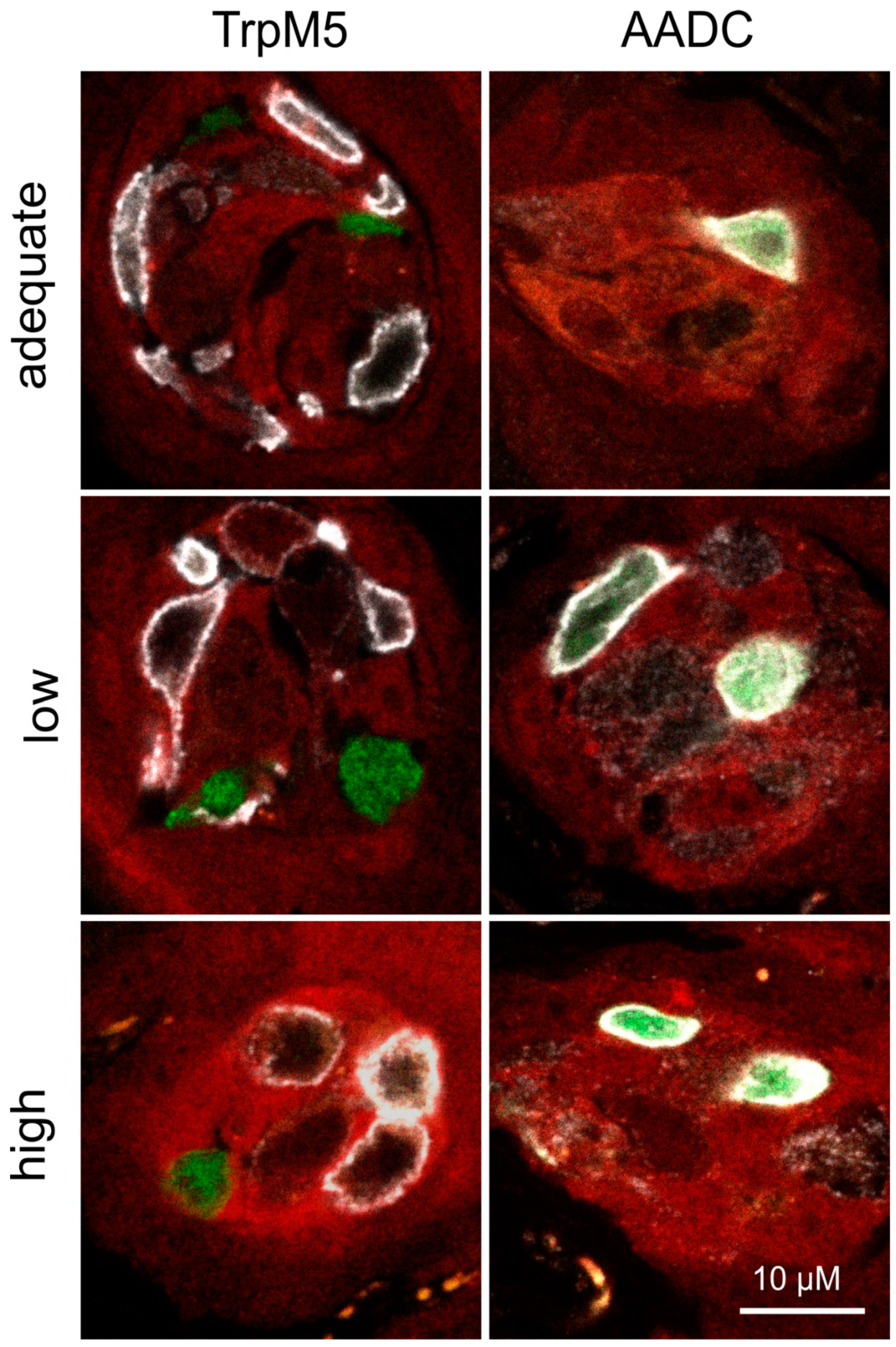
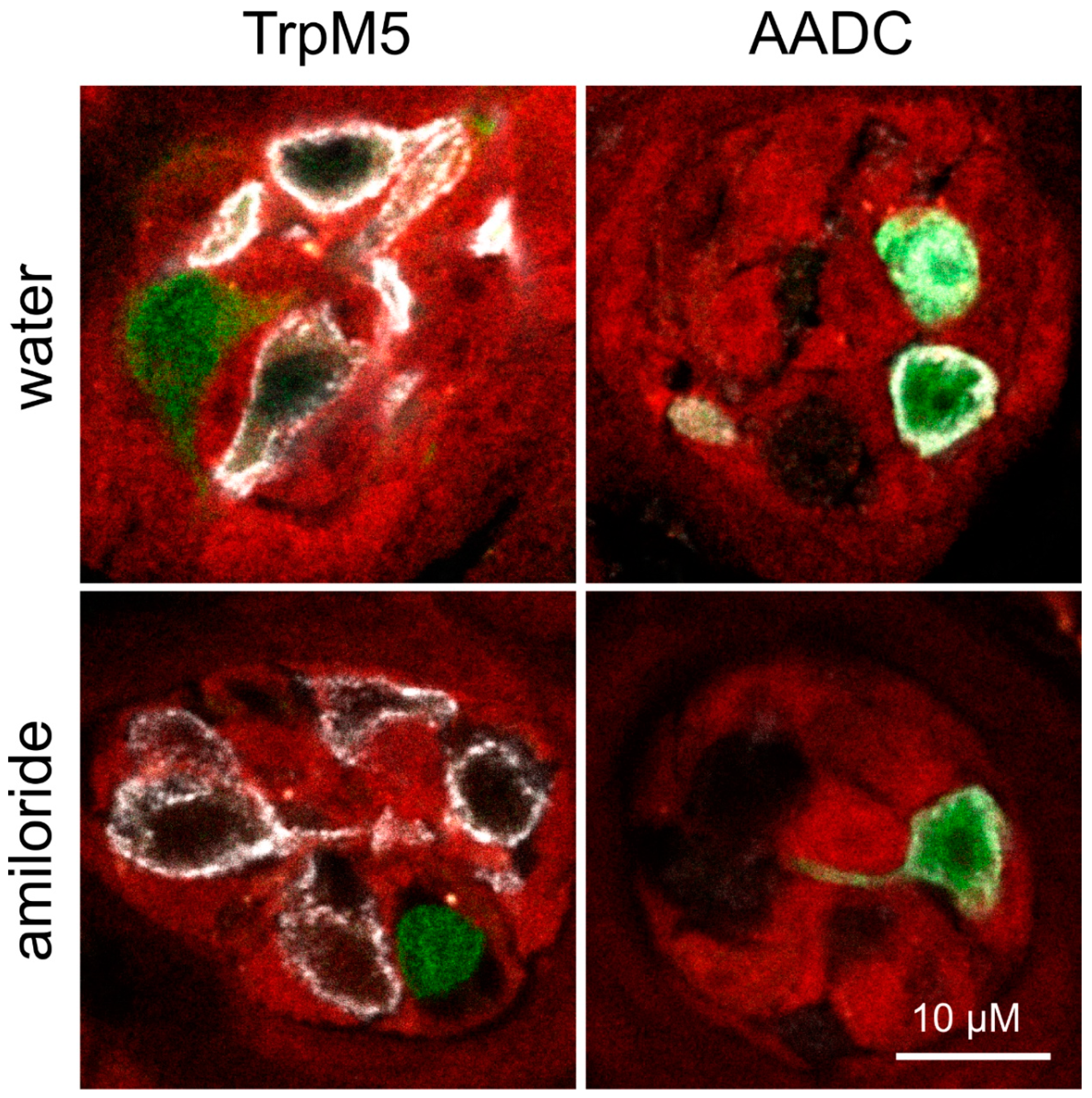
2.5. Immunohistochemistry
2.6. RNA Isolation and qRT-PCR
2.7. Functional Expression Analysis
2.8. Statistics
3. Results
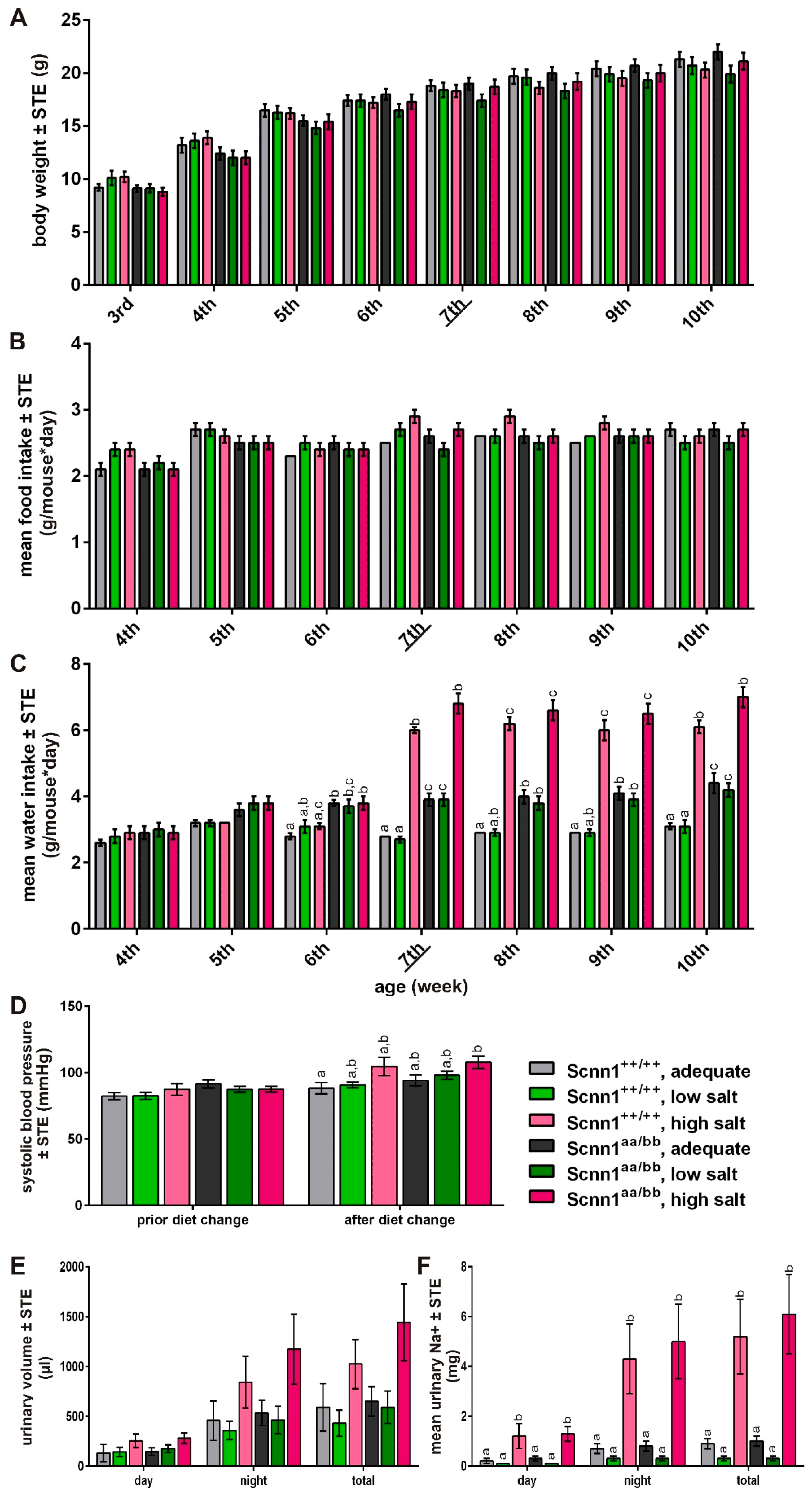
3.1. Feeding Experiment
3.2. Expression Analysis
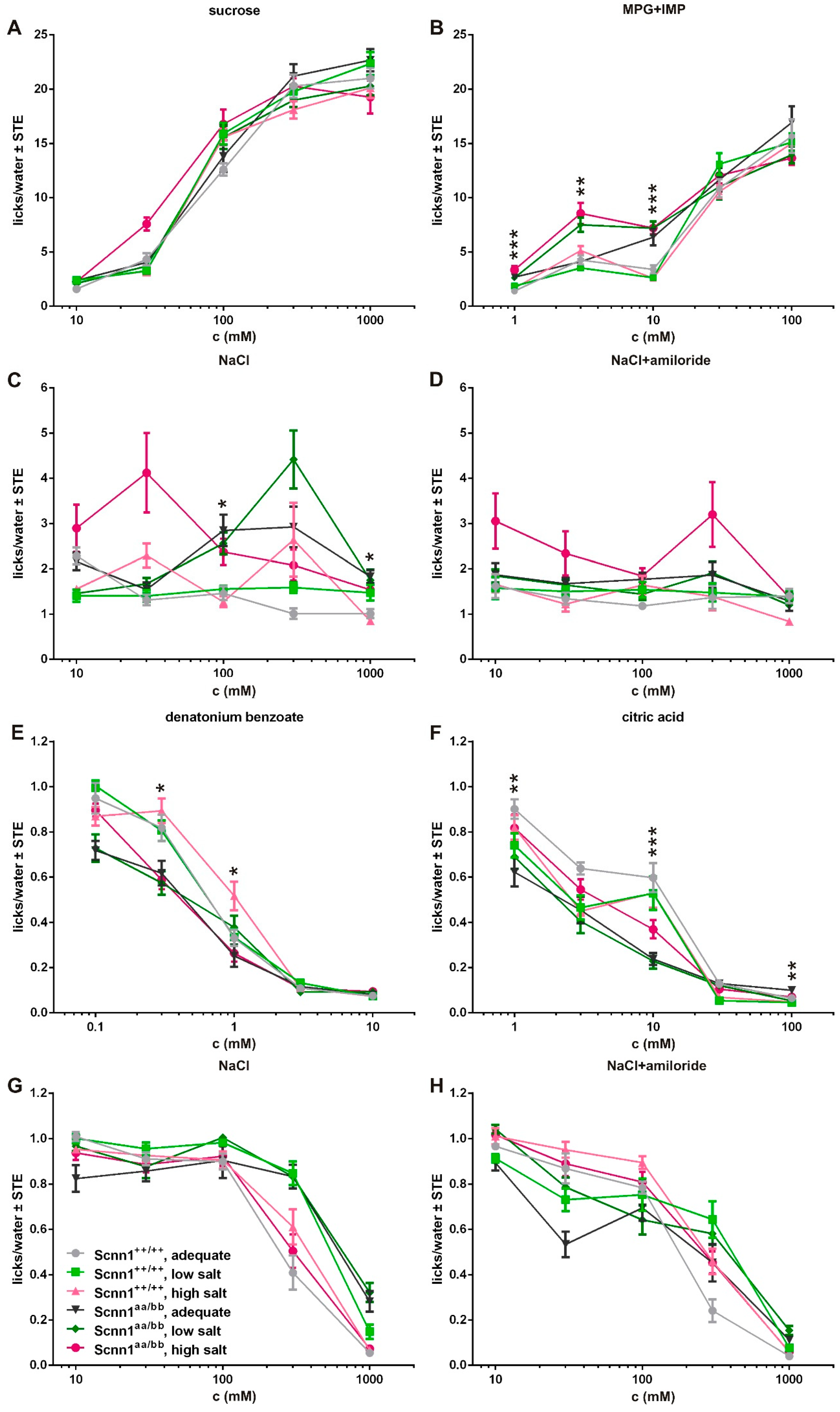
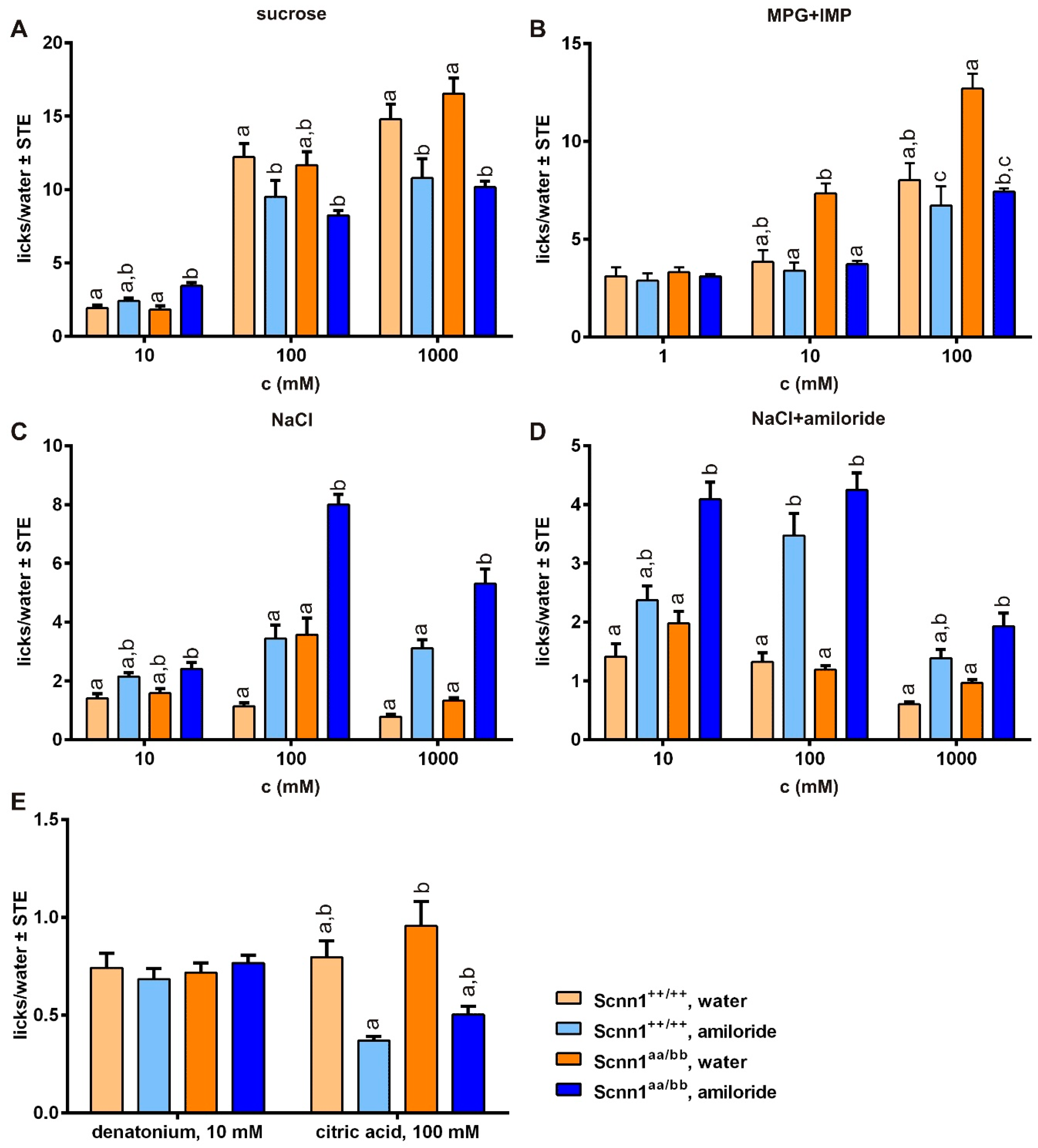
3.3. Short-term Preference Tests
3.4. Amiloride Intervention
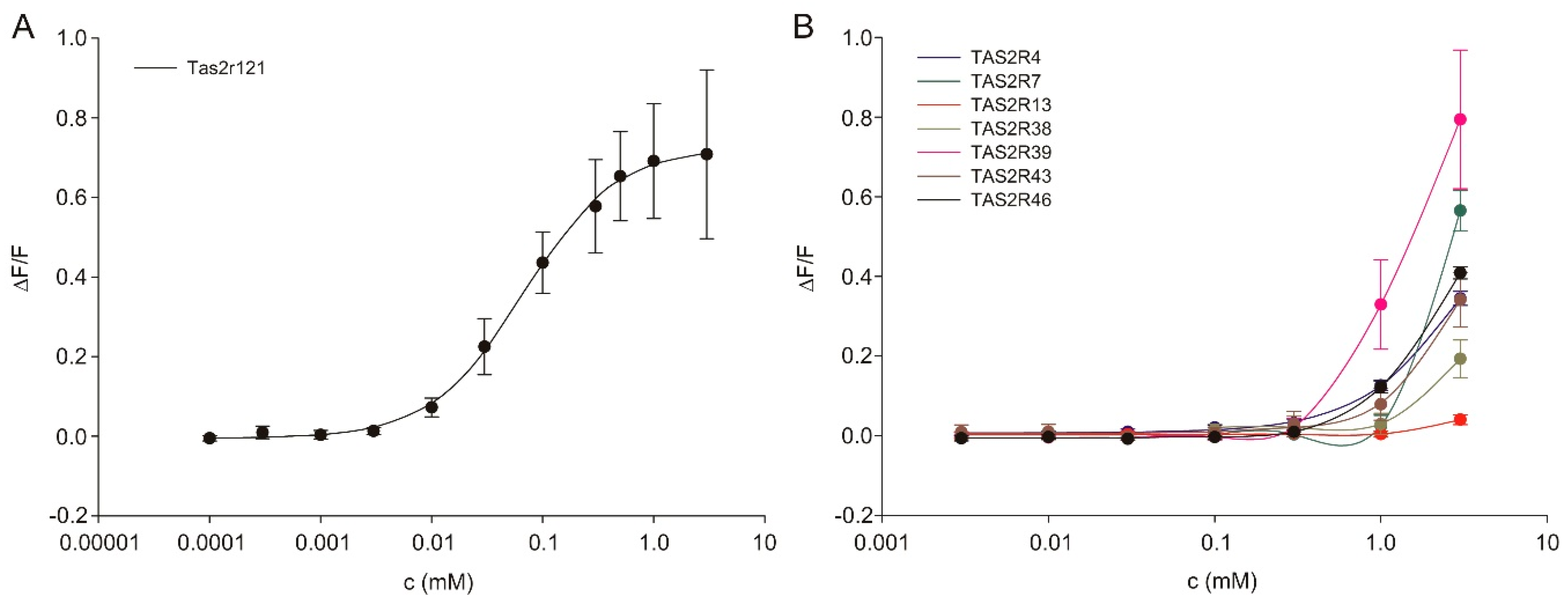
3.5. Amiloride Interaction with Bitter Taste Receptors
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hollenberg, N.K. Surfeit, deficit, and the set point for sodium homeostasis. Kidney Int. 1982, 21, 883–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damkjaer, M.; Jensen, P.H.; Schwammle, V.; Sprenger, R.R.; Jacobsen, I.A.; Jensen, O.N.; Bie, P. Selective renal vasoconstriction, exaggerated natriuresis and excretion rates of exosomic proteins in essential hypertension. Acta Physiol. (Oxf.) 2014, 212, 106–118. [Google Scholar] [CrossRef] [PubMed]
- Healy, V.; Thompson, C.; Johns, E.J. The adrenergic regulation of proximal tubular Na(+)/H(+) exchanger 3 in the rat. Acta Physiol. (Oxf.) 2014, 210, 678–689. [Google Scholar] [CrossRef] [PubMed]
- Fujita, T. Aldosterone in salt-sensitive hypertension and metabolic syndrome. J. Mol. Med. (Berl.) 2008, 86, 729–734. [Google Scholar] [CrossRef]
- Loffing, J.; Pietri, L.; Aregger, F.; Bloch-Faure, M.; Ziegler, U.; Meneton, P.; Rossier, B.C.; Kaissling, B. Differential subcellular localization of ENaC subunits in mouse kidney in response to high- and low-Na diets. Am. J. Physiol. Ren. Physiol. 2000, 279, F252–258. [Google Scholar] [CrossRef]
- Duc, C.; Farman, N.; Canessa, C.M.; Bonvalet, J.P.; Rossier, B.C. Cell-specific expression of epithelial sodium channel alpha, beta, and gamma subunits in aldosterone-responsive epithelia from the rat: Localization by in situ hybridization and immunocytochemistry. J. Cell Biol. 1994, 127, 1907–1921. [Google Scholar] [CrossRef]
- Kunzelmann, K.; Mall, M. Electrolyte transport in the mammalian colon: Mechanisms and implications for disease. Physiol. Rev. 2002, 82, 245–289. [Google Scholar] [CrossRef] [Green Version]
- Koyama, K.; Sasaki, I.; Naito, H.; Funayama, Y.; Fukushima, K.; Unno, M.; Matsuno, S.; Hayashi, H.; Suzuki, Y. Induction of epithelial Na+ channel in rat ileum after proctocolectomy. Am. J. Physiol. 1999, 276, G975–984. [Google Scholar] [CrossRef]
- Heck, G.L.; Mierson, S.; DeSimone, J.A. Salt taste transduction occurs through an amiloride-sensitive sodium transport pathway. Science 1984, 223, 403–405. [Google Scholar] [CrossRef]
- Brand, J.G.; Teeter, J.H.; Silver, W.L. Inhibition by amiloride of chorda tympani responses evoked by monovalent salts. Brain Res. 1985, 334, 207–214. [Google Scholar] [CrossRef]
- Garty, H.; Palmer, L.G. Epithelial sodium channels: Function, structure, and regulation. Physiol. Rev. 1997, 77, 359–396. [Google Scholar] [CrossRef] [PubMed]
- Kashlan, O.B.; Kleyman, T.R. ENaC structure and function in the wake of a resolved structure of a family member. Am. J. Physiol. Ren. Physiol. 2011, 301, F684–F696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cragoe, E.J., Jr.; Woltersdorf, O.W., Jr.; Bicking, J.B.; Kwong, S.F.; Jones, J.H. Pyrazine diuretics. II. N-amidino-3-amino-5-substituted 6-halopyrazinecarboxamides. J. Med. Chem. 1967, 10, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Cohen, A.B. Hyperkalemic effects of triamterene. Ann. Intern. Med. 1966, 65, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Gombos, E.A.; Freis, E.D.; Moghadam, A. Effects of MK-870 in normal subjects and hypertensive patients. N. Engl. J. Med. 1966, 275, 1215–1220. [Google Scholar] [CrossRef] [PubMed]
- Saha, C.; Eckert, G.J.; Ambrosius, W.T.; Chun, T.Y.; Wagner, M.A.; Zhao, Q.; Pratt, J.H. Improvement in blood pressure with inhibition of the epithelial sodium channel in blacks with hypertension. Hypertension 2005, 46, 481–487. [Google Scholar] [CrossRef] [Green Version]
- Denton, D.A. The Hunger for Salt; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 1882. [Google Scholar]
- St John, S.J. The perceptual characteristics of sodium chloride to sodium-depleted rats. Chem. Senses 2017, 42, 93–103. [Google Scholar] [CrossRef] [Green Version]
- Geerling, J.C.; Loewy, A.D. Central regulation of sodium appetite. Exp. Physiol. 2008, 93, 177–209. [Google Scholar] [CrossRef]
- Johnson, A.K.; Thunhorst, R.L. The neuroendocrinology of thirst and salt appetite: Visceral sensory signals and mechanisms of central integration. Front. Neuroendocrinol. 1997, 18, 292–353. [Google Scholar] [CrossRef]
- Fischer, N.M.; Schiefer, C.P.; St John, S.J. Gustatory contributions to sodium appetite: A microstructural analysis. Chem. Senses 2016, 41, E60. [Google Scholar]
- Sato, T.; Miyamoto, T.; Okada, Y. Comparison of gustatory transduction mechanisms in vertebrate taste cells. Zool. Sci. 1994, 11, 767–780. [Google Scholar]
- Lindemann, B. Taste reception. Physiol. Rev. 1996, 76, 718–766. [Google Scholar] [CrossRef] [PubMed]
- Stewart, R.E.; DeSimone, J.A.; Hill, D.L. New perspectives in a gustatory physiology: Transduction, development, and plasticity. Am. J. Physiol. 1997, 272, C1–26. [Google Scholar] [CrossRef] [PubMed]
- Benos, D.J. Amiloride: A molecular probe of sodium transport in tissues and cells. Am. J. Physiol. 1982, 242, C131–145. [Google Scholar] [CrossRef] [Green Version]
- Lindemann, B. Receptors and transduction in taste. Nature 2001, 413, 219–225. [Google Scholar] [CrossRef]
- Ninomiya, Y.; Funakoshi, M. Amiloride inhibition of responses of rat single chorda tympani fibers to chemical and electrical tongue stimulations. Brain Res. 1988, 451, 319–325. [Google Scholar] [CrossRef]
- Lin, W.; Finger, T.E.; Rossier, B.C.; Kinnamon, S.C. Epithelial Na+ channel subunits in rat taste cells: Localization and regulation by aldosterone. J. Comp. Neurol. 1999, 405, 406–420. [Google Scholar] [CrossRef]
- Gilbertson, T.A.; Kinnamon, S.C. Making sense of chemicals. Chem. Biol. 1996, 3, 233–237. [Google Scholar] [CrossRef] [Green Version]
- Boughter, J.D., Jr.; Gilbertson, T.A. From channels to behavior: An integrative model of NaCl taste. Neuron 1999, 22, 213–215. [Google Scholar] [CrossRef] [Green Version]
- Kretz, O.; Barbry, P.; Bock, R.; Lindemann, B. Differential expression of RNA and protein of the three pore-forming subunits of the amiloride-sensitive epithelial sodium channel in taste buds of the rat. J. Histochem. Cytochem. Off. J. Histochem. Soc. 1999, 47, 51–64. [Google Scholar] [CrossRef]
- Chandrashekar, J.; Kuhn, C.; Oka, Y.; Yarmolinsky, D.A.; Hummler, E.; Ryba, N.J.; Zuker, C.S. The cells and peripheral representation of sodium taste in mice. Nature 2010, 464, 297–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lossow, K.; Hermans-Borgmeyer, I.; Meyerhof, W.; Behrens, M. Segregated expression of ENaC subunits in taste cells. Chem. Senses 2020. [Google Scholar] [CrossRef] [PubMed]
- Jasti, J.; Furukawa, H.; Gonzales, E.B.; Gouaux, E. Structure of acid-sensing ion channel 1 at 1.9 A resolution and low pH. Nature 2007, 449, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Noreng, S.; Bharadwaj, A.; Posert, R.; Yoshioka, C.; Baconguis, I. Structure of the human epithelial sodium channel by cryo-electron microscopy. Elife 2018, 7. [Google Scholar] [CrossRef]
- Huang, T.; Stahler, F. Effects of dietary Na+ deprivation on epithelial Na+ channel (ENaC), BDNF, and TrkB mRNA expression in the rat tongue. Bmc Neurosci. 2009, 10, 19. [Google Scholar] [CrossRef] [Green Version]
- Hedrich, H. The Laboratory Mouse, 1st ed.; Elsevier, Academic Press: Cambridge, MA, USA, 2004. [Google Scholar]
- Raymond, M.A.; Chowdhury, T.; Mast, T.G.; Breza, J.M. A simple method for a rapid induction of salt appetite in mice. Annu. Meet. Assoc. Chemorecept. Sci. 2017. [Google Scholar] [CrossRef]
- Lossow, K.; Hubner, S.; Roudnitzky, N.; Slack, J.P.; Pollastro, F.; Behrens, M.; Meyerhof, W. Comprehensive analysis of mouse bitter taste receptors reveals different molecular receptive ranges for orthologous receptors in mice and humans. J. Biol. Chem. 2016, 291, 15358–15377. [Google Scholar] [CrossRef] [Green Version]
- Kusumakshi, S.; Voigt, A.; Hubner, S.; Hermans-Borgmeyer, I.; Ortalli, A.; Pyrski, M.; Dorr, J.; Zufall, F.; Flockerzi, V.; Meyerhof, W.; et al. A binary genetic approach to characterize trpm5 cells in mice. Chem. Senses 2015, 40, 413–425. [Google Scholar] [CrossRef] [Green Version]
- Hill, D.L.; Formaker, B.K.; White, K.S. Perceptual characteristics of the amiloride-suppressed sodium chloride taste response in the rat. Behav. Neurosci. 1990, 104, 734–741. [Google Scholar] [CrossRef]
- Markison, S.; Spector, A.C. Amiloride is an ineffective conditioned stimulus in taste aversion learning. Chem. Senses 1995, 20, 559–563. [Google Scholar] [CrossRef]
- Eylam, S.; Tracy, T.; Garcea, M.; Spector, A.C. Amiloride is an ineffective conditioned stimulus in taste aversion learning in C57BL/6J and DBA/2J mice. Chem. Senses 2003, 28, 681–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desor, J.A.; Finn, J. Effects of amiloride on salt taste in humans. Chem. Senses 1989, 14, 793–803. [Google Scholar] [CrossRef]
- McCutcheon, N.B. Human psychophysical studies of saltiness suppression by amiloride. Physiol. Behav. 1992, 51, 1069–1074. [Google Scholar] [CrossRef]
- Smith, D.V.; Ossebaard, C.A. Amiloride suppression of the taste intensity of sodium chloride: Evidence from direct magnitude scaling. Physiol. Behav. 1995, 57, 773–777. [Google Scholar] [CrossRef]
- Halpern, B.P. Amiloride and vertebrate gustatory responses to NaCl. Neurosci. Biobehav. Rev. 1998, 23, 5–47. [Google Scholar] [CrossRef]
- Brown, I.J.; Tzoulaki, I.; Candeias, V.; Elliott, P. Salt intakes around the world: Implications for public health. Int. J. Epidemiol. 2009, 38, 791–813. [Google Scholar] [CrossRef] [Green Version]
- Mozaffarian, D.; Fahimi, S.; Singh, G.M.; Micha, R.; Khatibzadeh, S.; Engell, R.E.; Lim, S.; Danaei, G.; Ezzati, M.; Powles, J.; et al. Global sodium consumption and death from cardiovascular causes. N. Engl. J. Med. 2014, 371, 624–634. [Google Scholar] [CrossRef] [Green Version]
- O′Donnell, M.; Mente, A.; Rangarajan, S.; McQueen, M.J.; Wang, X.; Liu, L.; Yan, H.; Lee, S.F.; Mony, P.; Devanath, A.; et al. Urinary sodium and potassium excretion, mortality, and cardiovascular events. N. Engl. J. Med. 2014, 371, 612–623. [Google Scholar] [CrossRef] [Green Version]
- WHO. Guideline: Sodium Intake for Adults and Children; World Health Organization (WHO): Geneva, Switzerland, 2012; Available online: www.who.int (accessed on 1 October 2018).
- Meneton, P.; Jeunemaitre, X.; de Wardener, H.E.; MacGregor, G.A. Links between dietary salt intake, renal salt handling, blood pressure, and cardiovascular diseases. Physiol. Rev. 2005, 85, 679–715. [Google Scholar] [CrossRef]
- He, F.J.; MacGregor, G.A. Reducing population salt intake worldwide: From evidence to implementation. Prog. Cardiovasc. Dis. 2010, 52, 363–382. [Google Scholar] [CrossRef]
- He, F.J.; MacGregor, G.A. A comprehensive review on salt and health and current experience of worldwide salt reduction programmes. J. Hum. Hypertens. 2009, 23, 363–384. [Google Scholar] [CrossRef] [PubMed]
- Bibbins-Domingo, K.; Chertow, G.M.; Coxson, P.G.; Moran, A.; Lightwood, J.M.; Pletcher, M.J.; Goldman, L. Projected effect of dietary salt reductions on future cardiovascular disease. N. Engl. J. Med. 2010, 362, 590–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sacks, F.M.; Campos, H. Dietary therapy in hypertension. N. Engl. J. Med. 2010, 362, 2102–2112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsugane, S.; Sasazuki, S.; Kobayashi, M.; Sasaki, S. Salt and salted food intake and subsequent risk of gastric cancer among middle-aged Japanese men and women. Br. J. Cancer 2004, 90, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Devine, A.; Criddle, R.A.; Dick, I.M.; Kerr, D.A.; Prince, R.L. A longitudinal study of the effect of sodium and calcium intakes on regional bone density in postmenopausal women. Am. J. Clin. Nutr. 1995, 62, 740–745. [Google Scholar] [CrossRef] [PubMed]
- Asaria, P.; Chisholm, D.; Mathers, C.; Ezzati, M.; Beaglehole, R. Chronic disease prevention: Health effects and financial costs of strategies to reduce salt intake and control tobacco use. Lancet 2007, 370, 2044–2053. [Google Scholar] [CrossRef]
- Strazzullo, P.; D′Elia, L.; Kandala, N.B.; Cappuccio, F.P. Salt intake, stroke, and cardiovascular disease: Meta-analysis of prospective studies. BMJ 2009, 339, b4567. [Google Scholar] [CrossRef] [Green Version]
- Bosak, N.P.; Inoue, M.; Nelson, T.M.; Hummler, E.; Ishiwatari, Y.; Bachmanov, A.A. Epithelial sodium channel (ENaC) is involved in reception of sodium taste: Evidence from mice with a tissue-specific conditional targeted mutation of the ENaC gene (abstract), thirty-second annual meeting of the association for chemoreception sciences. Chem. Senses 2010, 35, A2. [Google Scholar]
- Canessa, C.M.; Schild, L.; Buell, G.; Thorens, B.; Gautschi, I.; Horisberger, J.D.; Rossier, B.C. Amiloride-sensitive epithelial Na+ channel is made of three homologous subunits. Nature 1994, 367, 463–467. [Google Scholar] [CrossRef]
- McDonald, F.J.; Price, M.P.; Snyder, P.M.; Welsh, M.J. Cloning and expression of the beta- and gamma-subunits of the human epithelial sodium channel. Am. J. Physiol. 1995, 268, C1157–C1163. [Google Scholar] [CrossRef]
- Giraldez, T.; Afonso-Oramas, D.; Cruz-Muros, I.; Garcia-Marin, V.; Pagel, P.; Gonzalez-Hernandez, T.; Alvarez de la Rosa, D. Cloning and functional expression of a new epithelial sodium channel delta subunit isoform differentially expressed in neurons of the human and monkey telencephalon. J. Neurochem. 2007, 102, 1304–1315. [Google Scholar] [CrossRef] [PubMed]
- Edelheit, O.; Hanukoglu, I.; Dascal, N.; Hanukoglu, A. Identification of the roles of conserved charged residues in the extracellular domain of an epithelial sodium channel (ENaC) subunit by alanine mutagenesis. Am. J. Physiol. Ren. Physiol. 2011, 300, F887–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edelheit, O.; Ben-Shahar, R.; Dascal, N.; Hanukoglu, A.; Hanukoglu, I. Conserved charged residues at the surface and interface of epithelial sodium channel subunits--roles in cell surface expression and the sodium self-inhibition response. Febs J. 2014, 281, 2097–2111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stricker, E.M.; Hoffmann, M.L.; Riccardi, C.J.; Smith, J.C. Increased water intake by rats maintained on high NaCl diet: Analysis of ingestive behavior. Physiol. Behav. 2003, 79, 621–631. [Google Scholar] [CrossRef]
- Ramsey, D.J.; Thrasher, T.N. Thirst and water balance. Neurobiology of food and fluid intake. In Handbook of Behavioral Neurobiology; Stricker, E.M., Ed.; Plenum Press: New York, NY, USA, 1990; Volume 10, pp. 353–386. [Google Scholar]
- Contreras, R.J.; Hatton, G.I. Gustatory adaptation as an explanation for dietary-induced sodium appetite. Physiol. Behav. 1975, 15, 569–576. [Google Scholar] [CrossRef]
- Stricker, E.M.; Thiels, E.; Verbalis, J.G. Sodium appetite in rats after prolonged dietary sodium deprivation: A sexually dimorphic phenomenon. Am. J. Physiol. 1991, 260, R1082–R1088. [Google Scholar] [CrossRef]
- Berridge, K.C.; Flynn, F.W.; Schulkin, J.; Grill, H.J. Sodium depletion enhances salt palatability in rats. Behav. Neurosci. 1984, 98, 652–660. [Google Scholar] [CrossRef]
- Nachman, M.; Valentino, D.A. Roles of taste and postingestional factors in the satiation of sodium appetite in rats. J. Comp. Physiol. Psychol. 1966, 62, 280–283. [Google Scholar] [CrossRef]
- Wolf, G.; Schulkin, J.; Simson, P.E. Multiple factors in the satiation of salt appetite. Behav. Neurosci. 1984, 98, 661–673. [Google Scholar] [CrossRef]
- Handal, P.J. Immediate acceptance of sodium salts by sodium deficient rats. Psychon. Sci. 1965, 3, 315–316. [Google Scholar] [CrossRef] [Green Version]
- Fortin, S.M.; Roitman, M.F. Physiological state tunes mesolimbic signaling: Lessons from sodium appetite and inspiration from Randall R. Sakai. Physiol. Behav. 2017, 178, 21–27. [Google Scholar] [CrossRef] [PubMed]
- St John, S.J.; McBrayer, A.M.; Krauskopf, E.E. Sodium carbonate is saltier than sodium chloride to sodium-depleted rats. Chem. Senses 2017, 42, 647–653. [Google Scholar] [CrossRef] [PubMed]
- Beauchamp, G.K.; Bertino, M.; Burke, D.; Engelman, K. Experimental sodium depletion and salt taste in normal human volunteers. Am. J. Clin. Nutr. 1990, 51, 881–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernstein, I.L.; Hennessy, C.J. Amiloride-sensitive sodium channels and expression of sodium appetite in rats. Am. J. Physiol. 1987, 253, R371–374. [Google Scholar] [CrossRef]
- McCutcheon, N.B. Sodium deficient rats are unmotivated by sodium chloride solutions mixed with the sodium channel blocker amiloride. Behav. Neurosci. 1991, 105, 764–766. [Google Scholar] [CrossRef]
- Oka, Y.; Butnaru, M.; von Buchholtz, L.; Ryba, N.J.; Zuker, C.S. High salt recruits aversive taste pathways. Nature 2013, 494, 472–475. [Google Scholar] [CrossRef] [Green Version]
- Ren, Z.; Rhyu, M.R.; Phan, T.H.; Mummalaneni, S.; Murthy, K.S.; Grider, J.R.; DeSimone, J.A.; Lyall, V. TRPM5-dependent amiloride- and benzamil-insensitive NaCl chorda tympani taste nerve response. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 305, G106–117. [Google Scholar] [CrossRef] [Green Version]
- Lewandowski, B.C.; Sukumaran, S.K.; Margolskee, R.F.; Bachmanov, A.A. Amiloride-insensitive salt taste is mediated by two populations of type iii taste cells with distinct transduction mechanisms. J. Neurosci. Off. J. Soc. Neurosci. 2016, 36, 1942–1953. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Hoon, M.A.; Chandrashekar, J.; Mueller, K.L.; Cook, B.; Wu, D.; Zuker, C.S.; Ryba, N.J. Coding of sweet, bitter, and umami tastes: Different receptor cells sharing similar signaling pathways. Cell 2003, 112, 293–301. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.; Li, X.; DiCicco-Bloom, E.; Bello, N.T. Altered salt taste response and increased tongue epithelium Scnna1 expression in adult Engrailed-2 null mice. Physiol. Behav. 2018, 194, 410–419. [Google Scholar] [CrossRef]
- Ribeiro, S.C.; Monteiro, G.A.; Prazeres, D.M. The role of polyadenylation signal secondary structures on the resistance of plasmid vectors to nucleases. J. Gene Med. 2004, 6, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Azzoni, A.R.; Ribeiro, S.C.; Monteiro, G.A.; Prazeres, D.M. The impact of polyadenylation signals on plasmid nuclease-resistance and transgene expression. J. Gene Med. 2007, 9, 392–402. [Google Scholar] [CrossRef] [PubMed]
- Lindemann, B.; Barbry, P.; Kretz, O.; Bock, R. Occurrence of ENaC subunit mRNA and immunocytochemistry of the channel subunits in taste buds of the rat vallate papilla. Ann. N. Y. Acad. Sci. 1998, 855, 116–127. [Google Scholar] [CrossRef] [PubMed]
- Shigemura, N.; Islam, A.A.; Sadamitsu, C.; Yoshida, R.; Yasumatsu, K.; Ninomiya, Y. Expression of amiloride-sensitive epithelial sodium channels in mouse taste cells after chorda tympani nerve crush. Chem. Senses 2005, 30, 531–538. [Google Scholar] [CrossRef] [Green Version]
- Lingueglia, E.; Renard, S.; Waldmann, R.; Voilley, N.; Champigny, G.; Plass, H.; Lazdunski, M.; Barbry, P. Different homologous subunits of the amiloride-sensitive Na+ channel are differently regulated by aldosterone. J. Biol. Chem. 1994, 269, 13736–13739. [Google Scholar]
- Renard, S.; Voilley, N.; Bassilana, F.; Lazdunski, M.; Barbry, P. Localization and regulation by steroids of the alpha, beta and gamma subunits of the amiloride-sensitive Na+ channel in colon, lung and kidney. Pflug. Arch. Eur. J. Physiol. 1995, 430, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Asher, C.; Wald, H.; Rossier, B.C.; Garty, H. Aldosterone-induced increase in the abundance of Na+ channel subunits. Am. J. Physiol. 1996, 271, C605–C611. [Google Scholar] [CrossRef]
- Ono, S.; Kusano, E.; Muto, S.; Ando, Y.; Asano, Y. A low-Na+ diet enhances expression of mRNA for epithelial Na+ channel in rat renal inner medulla. Pflug. Arch. Eur. J. Physiol. 1997, 434, 756–763. [Google Scholar] [CrossRef]
- Stokes, J.B.; Sigmund, R.D. Regulation of rENaC mRNA by dietary NaCl and steroids: Organ, tissue, and steroid heterogeneity. Am. J. Physiol. 1998, 274, C1699–C1707. [Google Scholar] [CrossRef]
- Masilamani, S.; Kim, G.H.; Mitchell, C.; Wade, J.B.; Knepper, M.A. Aldosterone-mediated regulation of ENaC alpha, beta, and gamma subunit proteins in rat kidney. J. Clin. Investig. 1999, 104, R19–R23. [Google Scholar] [CrossRef] [Green Version]
- MacDonald, P.; MacKenzie, S.; Ramage, L.E.; Seckl, J.R.; Brown, R.W. Corticosteroid regulation of amiloride-sensitive sodium-channel subunit mRNA expression in mouse kidney. J. Endocrinol. 2000, 165, 25–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frindt, G.; Palmer, L.G. Regulation of epithelial Na+ channels by adrenal steroids: Mineralocorticoid and glucocorticoid effects. Am. J. Physiol. Ren. Physiol. 2012, 302, F20–F26. [Google Scholar] [CrossRef] [Green Version]
- Malsure, S.; Wang, Q.; Charles, R.P.; Sergi, C.; Perrier, R.; Christensen, B.M.; Maillard, M.; Rossier, B.C.; Hummler, E. Colon-specific deletion of epithelial sodium channel causes sodium loss and aldosterone resistance. J. Am. Soc. Nephrol. JASN 2014, 25, 1453–1464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marunaka, Y.; Marunaka, R.; Sun, H.; Yamamoto, T.; Kanamura, N.; Taruno, A. Na(+) homeostasis by epithelial Na(+) channel (ENaC) and Nax channel (Nax): Cooperation of ENaC and Nax. Ann. Transl. Med. 2016, 4, S11. [Google Scholar] [CrossRef] [Green Version]
- Schiffman, S.S.; Lockhead, E.; Maes, F.W. Amiloride reduces the taste intensity of Na+ and Li+ salts and sweeteners. Proc. Natl. Acad. Sci. USA 1983, 80, 6136–6140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imada, T.; Misaka, T.; Fujiwara, S.; Okada, S.; Fukuda, Y.; Abe, K. Amiloride reduces the sweet taste intensity by inhibiting the human sweet taste receptor. Biochem. Biophys. Res. Commun. 2010, 397, 220–225. [Google Scholar] [CrossRef]
- Yee, K.K.; Sukumaran, S.K.; Kotha, R.; Gilbertson, T.A.; Margolskee, R.F. Glucose transporters and ATP-gated K+ (KATP) metabolic sensors are present in type 1 taste receptor 3 (T1r3)-expressing taste cells. Proc. Natl. Acad. Sci. USA 2011, 108, 5431–5436. [Google Scholar] [CrossRef] [Green Version]
- Hellekant, G.; DuBois, G.E.; Roberts, T.W.; van der Wel, H. On the gustatory effect of amiloride in the monkey (Macaca mulatto) Chem. Senses 1988, 13, 89–93. [Google Scholar] [CrossRef]
- DeSimone, J.A.; Callaham, E.M.; Heck, G.L. Chorda tympani taste response of rat to hydrochloric acid subject to voltage-clamped lingual receptive field. Am. J. Physiol. 1995, 268, C1295–C1300. [Google Scholar] [CrossRef]
- Ossebaard, C.A.; Smith, D.V. Effect of amiloride on the taste of NaCl, Na-gluconate and KCl in humans: Implications for Na+ receptor mechanisms. Chem. Senses 1995, 20, 37–46. [Google Scholar] [CrossRef]
- Miyamoto, T.; Fujiyama, R.; Okada, Y.; Sato, T. Sour transduction involves activation of NPPB-sensitive conductance in mouse taste cells. J. Neurophysiol. 1998, 80, 1852–1859. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ENaC Subunit | Adequate (mean ± STABW) | Low Salt (mean ± STABW) | High Salt (mean ± STABW) | p-Value | |
|---|---|---|---|---|---|
| fungiform papillae | α | 0.0151 ± 0.0053 | 0.0174 ± 0.0098 | 0.0201 ± 0.0104 | 0.735 |
| β | 0.0062 ± 0.0030 | 0.0101 ± 0.0059 | 0.0087 ± 0.0069 | 0.464 | |
| γ | 0.0093 ± 0.0026 | 0.0118 ± 0.0066 | 0.0117 ± 0.0075 | 0.719 | |
| vallate and foliate papillae | α | 0.0606 ± 0.0111 | 0.0577 ± 0.0217 | 0.0538 ± 0.0152 | 0.780 |
| β | 0.0027 ± 0.0007 | 0.0024 ± 0.0010 | 0.0036 ± 0.0009 | 0.084 | |
| γ | 0.0020 ± 0.0005 | 0.0018 ± 0.0007 | 0.0021 ± 0.0004 | 0.669 | |
| non-gustatory epithelium | α | 0.0228 ± 0.0061 | 0.0283 ± 0.0107 | 0.0205 ± 0.0037 | 0.212 |
| β | 0.0011 ± 0.0004 | 0.0010 ± 0.0003 | 0.0010 ± 0.0002 | 0.942 | |
| γ | 0.0008 ± 0.0001 | 0.0007 ± 0.0003 | 0.0007 ± 0.0002 | 0.891 | |
| kidney | α | 0.0698 ± 0.0235 | 0.0600 ± 0.0154 | 0.0420 ± 0.0069 | 0.034 * |
| β | 0.0260 ± 0.0086 | 0.0196 ± 0.0043 | 0.0215 ± 0.0043 | 0.211 | |
| γ | 0.0425 ± 0.0112 | 0.0296 ± 0.0036 | 0.0360 ± 0.0084 | 0.054 | |
| distal colon | α | 0.0357 ± 0.0104 | 0.0504 ± 0.0102 | 0.0266 ± 0.0063 | 0.002#,$ |
| β | 0.0051 ± 0.0017 | 0.0258 ± 0.0184 | 0.0026 ± 0.0010 | 0.003#,$ | |
| γ | 0.0091 ± 0.0068 | 0.0306 ± 0.0302 | 0.0027 ± 0.0026 | 0.041$ |
| ENaC Subunit | Water (mean ± STABW) | Amiloride (mean ± STABW) | p-Value | |
|---|---|---|---|---|
| fungiform papillae | α | 0.0104 ± 0.0019 | 0.0136 ± 0.0019 | 0.051 |
| β | 0.0030 ± 0.0014 | 0.0060 ± 0.0012 | 0.017 | |
| γ | 0.0056 ± 0.0025 | 0.0063 ± 0.0024 | 0.700 | |
| vallate and foliate papillae | α | 0.0529 ± 0.0089 | 0.0700 ± 0.0083 | 0.031 |
| β | 0.0024 ± 0.0003 | 0.0028 ± 0.0006 | 0.311 | |
| γ | 0.0018 ± 0.0004 | 0.0017 ± 0.0003 | 0.853 | |
| non-gustatory epithelium | α | 0.0229 ± 0.0041 | 0.0280 ± 0.0019 | 0.066 |
| β | 0.0008 ± 0.0004 | 0.0013 ± 0.0004 | 0.195 | |
| γ | 0.0007 ± 0.0002 | 0.0008 ± 0.0003 | 0.682 | |
| kidney | α | 0.0583 ± 0.0217 | 0.0744 ± 0.0199 | 0.317 |
| β | 0.0223 ± 0.0061 | 0.0238 ± 0.0057 | 0.738 | |
| γ | 0.0399 ± 0.0172 | 0.0326 ± 0.0034 | 0.432 | |
| distal colon | α | 0.0256 ± 0.0097 | 0.0597 ± 0.0164 | 0.012 |
| β | 0.0051 ± 0.0018 | 0.0428 ± 0.0105 | <0.001 | |
| γ | 0.0067 ± 0.0038 | 0.0684 ± 0.0227 | 0.002 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lossow, K.; Meyerhof, W.; Behrens, M. Sodium Imbalance in Mice Results Primarily in Compensatory Gene Regulatory Responses in Kidney and Colon, but Not in Taste Tissue. Nutrients 2020, 12, 995. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12040995
Lossow K, Meyerhof W, Behrens M. Sodium Imbalance in Mice Results Primarily in Compensatory Gene Regulatory Responses in Kidney and Colon, but Not in Taste Tissue. Nutrients. 2020; 12(4):995. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12040995
Chicago/Turabian StyleLossow, Kristina, Wolfgang Meyerhof, and Maik Behrens. 2020. "Sodium Imbalance in Mice Results Primarily in Compensatory Gene Regulatory Responses in Kidney and Colon, but Not in Taste Tissue" Nutrients 12, no. 4: 995. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12040995