Histidine: A Systematic Review on Metabolism and Physiological Effects in Human and Different Animal Species

 and
and 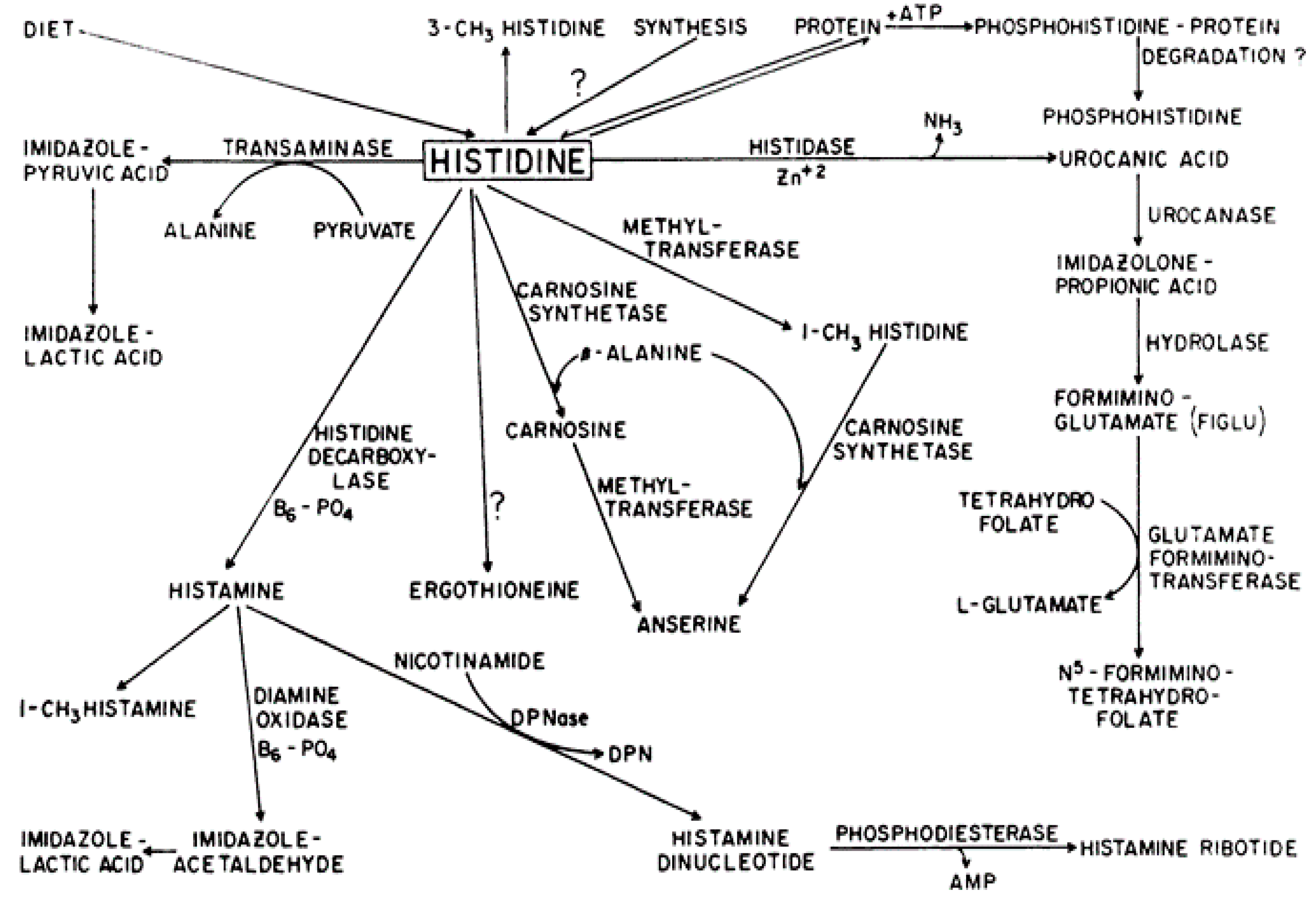{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Common Physiological Roles of Histidine in Different Species
2.1. Histidine Requirements and Metabolism
2.2. Physiological Role of Histidine is Common for All Species
2.3. Histidine Strongly Binds Metal (CO(II), Ni(II), Cd(II), Zn(II), Cu(II), and Fe(III) Ions)
3. Effect of Histidine Level and Supplementation in Humans
3.1. Histidine Intake and Consequences on Body Histidine Status
3.2. Histidine Supplementation and Eating Disorders
3.3. Histidine and Memory Disorders
3.4. Histidine and Metal Ion Status
3.5. Histidine and Skin Dysfunction
3.6. Histidine and Metabolic Syndrome
4. Effect of Histidine Level and Supplementation in Rodents (Rats and Mice)
4.1. Effect on Food Intake
4.2. Effect on Neuroprotection
4.3. Effect on Growth and Metabolic Dysfunctions
4.4. Effect of Histidine on Mineral Metabolism
4.5. Histidine and Cancer
4.6. Toxicity Dose
5. Effect of Histidine Level and Supplementation in Breeding Farmed, Monogastric Pigs and Chickens
5.1. Histidine in Pigs
5.2. Histidine in Chicken
6. Effect of Histidine Level and Supplementation in Fish
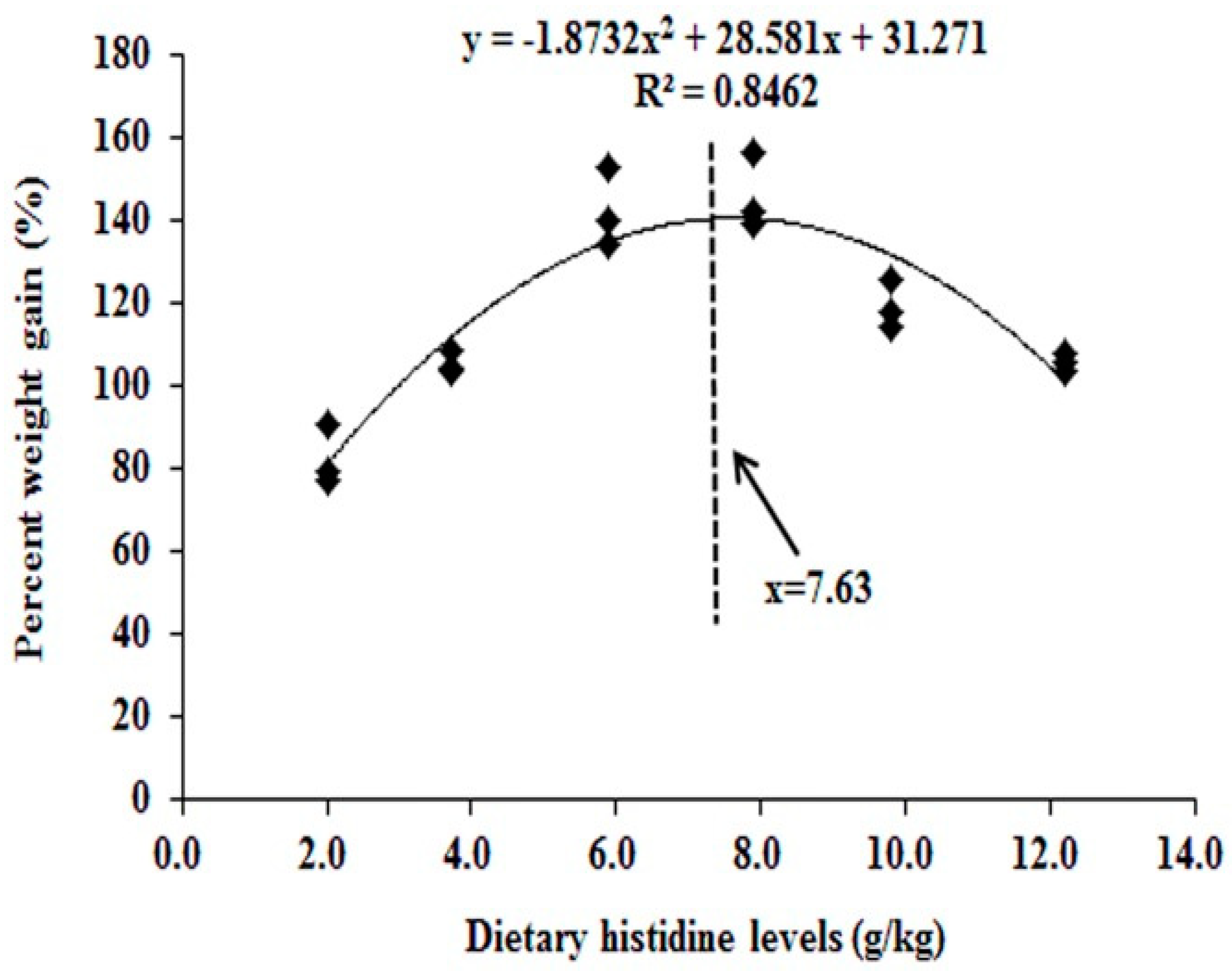
6.1. Histidine Effect on Fish Growth
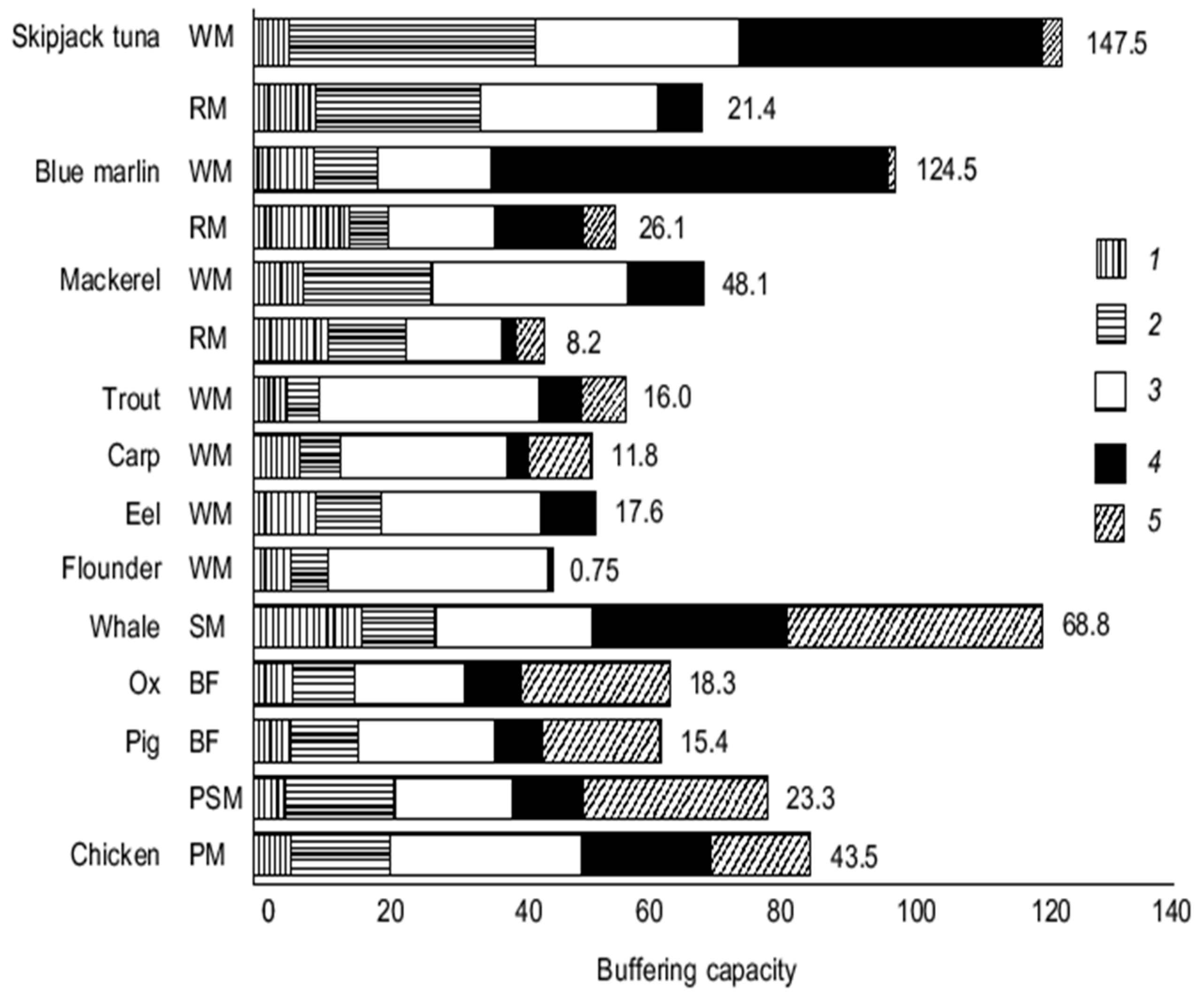
6.2. Buffering Role of Histidine in Fish
6.3. Role of Histidine in Preventing Cataracts in Fish
7. Effect of Histidine Level and Supplementation in Ruminants
7.1. Effect of Histidine on Growth
7.2. Effect of Histidine on Milk Protein Synthesis
8. Conclusions
- In human, the daily requirement for histidine is 8 to 12 mg/kg of body weight per day in adults, and the average intake in typical adult diets in Europe, USA and Japan was reported to be between 30 to 35 mg/kg of body weight per day (2.12 and 2.40 g per day). Daily histidine supplementation, under 2 g/daily, improves inflammation in overweight and obese people, decreases feelings of fatigue, and increases concentration and efficiency at work. A daily intake of 4 g decreases severity in atopic dermatitis disease. If the supplementation increases up to 8 g of histidine, a severe anorexia is induced. Finally, supplementation with a large dose, from 16 to 64 g/day, induces a decrease in taste and smell acuity, and the onset of headaches, weakness, drowsiness, nausea, and memory disorder.
- In rodents, histidine supplementation up to 25 g/kg of diet is beneficial. A large dose of 50 g/kg of diet can have different adverse effects, such as a large decrease in body weight gain, in food intake, and a growth retardation. Due to this negative effect, maximal tolerable dose of histidine was fixed to 25 g/kg of the diet.
- In pigs, histidine, when combined with other amino acids, can improve performance and amino acid balance in a context of low-protein diet.
- In chickens, the major benefit of histidine supplementation is the increase in carnosine and anserine in the muscle allowing an increase in anti-oxidant capacity.
- In salmonids, a beneficial effect of histidine supplementation is its capacity to decrease the incidence and severity of cataracts.
- In ruminants, histidine supplementation induces an increase in milk yield and an increase in milk protein synthesis.
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Rose, W.C. The amino acid requirements of adult man. Nutr. Abstr. Rev. 1957, 27, 631–647. [Google Scholar] [PubMed]
- Wu, G. Amino acids: Metabolism, functions, and nutrition. Amino Acids 2009, 37, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Kriengsinyos, W.; Rafii, M.; Wykes, L.J.; Ball, R.O.; Pencharz, P.B. Long-term effects of histidine depletion on whole-body protein metabolism in healthy adults. J. Nutr. 2002, 132, 3340–3348. [Google Scholar] [CrossRef] [Green Version]
- Onodera, R. Essentiality of histidine in ruminant and other animals including human Beings. Asian-Australas. J. Anim. Sci. 2003, 16, 445–454. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G. PRISMA group preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef] [Green Version]
- Taylor, R.G.; Levy, H.L.; McInnes, R.R. Histidase and histidinemia. Clinical and molecular considerations. Mol. Biol. Med. 1991, 8, 101–116. [Google Scholar] [PubMed]
- Mehler, A.H.; Tabor, H. Deamination of histidine to form urocanic acid in liver. J. Biol. Chem. 1953, 201, 775–784. [Google Scholar]
- Kinuta, M.; Ubuka, T.; Yao, W.B.; Zhao, Y.Q.; Shimizu, H. Isolation of S-[2-carboxy-1-(1H-imidazol-4-yl)ethyl]-3-thiolactic acid, a new metabolite of histidine, from normal human urine and its formation from S-[2-carboxy-1-(1H-imidazol-4-yl)ethyl]cysteine. Biochem. J. 1994, 297, 475–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hais, I.M.; Štrych, A. Increase in urocanic acid concentration in human epidermis following insolation. Collect. Czechoslov. Chem. Commun. 1969, 34, 649–655. [Google Scholar] [CrossRef]
- Zannoni, V.G.; La Du, B.N. Determination of histidine α-deaminase in human stratum corneum and its absence in histidinaemia. Biochem. J. 1963, 88, 160–162. [Google Scholar] [CrossRef] [Green Version]
- Stifel, F.B.; Herman, R.H. Histidine metabolism. Am. J. Clin. Nutr. 1971, 24, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Torres, N.; Beristain, L.; Bourges, H.; Tovar, A.R. Histidine-imbalanced diets stimulate hepatic histidase gene expression in rats. J. Nutr. 1999, 129, 1979–1983. [Google Scholar] [CrossRef] [Green Version]
- Tovar, A.R.; Santos, A.; Halhali, A.; Bourges, H.; Torres, N. Hepatic histidase gene expression responds to protein rehabilitation in undernourished growing rats. J. Nutr. 1998, 128, 1631–1635. [Google Scholar] [CrossRef] [PubMed]
- Kang-Lee, Y.A.; Harper, A.E. Effect of induction of histidase on histidine metabolism in vivo. J. Nutr. 1979, 109, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Feigelson, M. Estrogenic regulation of hepatic histidase during postnatal development and adulthood. J. Biol. Chem. 1968, 243, 5088–5093. [Google Scholar] [PubMed]
- Borsook, H.; Deasy, C.L.; Haagen-Smit, A.J.; Keighley, G.; Lowy, P.H. Metabolism of c14-labeled glycine, L-histidine, L-leucine, and L-lysine. J. Biol. Chem. 1950, 187, 839–848. [Google Scholar] [PubMed]
- Wolf, G. The metabolism of alpha-C14-histidine in the intact rat. I. Radioactivity in amino acids from protein. J. Biol. Chem. 1953, 200, 637–645. [Google Scholar]
- Harris, R.C.; Marlin, D.J.; Dunnett, M.; Snow, D.H.; Hultman, E. Muscle buffering capacity and dipeptide content in the thoroughbred horse, greyhound dog and man. Comp. Biochem. Physiol. A 1990, 97, 249–251. [Google Scholar] [CrossRef]
- Guiotto, A.; Calderan, A.; Borin, G.; Ruzza, P. Carnosine and Carnosine-Related Antioxidants: A Review. Available online: http://www.eurekaselect.com/60152/article (accessed on 28 February 2018).
- Asahi, R.; Tanaka, K.; Fujimi, T.J.; Kanzawa, N.; Nakajima, S. Proline decreases the suppressive effect of histidine on food intake and fat accumulation. J. Nutr. Sci. Vitaminol. (Tokyo) 2016, 62, 277–280. [Google Scholar] [CrossRef] [Green Version]
- Xie, H.; He, S.-H. Roles of histamine and its receptors in allergic and inflammatory bowel diseases. World J. Gastroenterol. WJG 2005, 11, 2851–2857. [Google Scholar] [CrossRef]
- Branco, A.C.C.C.; Yoshikawa, F.S.Y.; Pietrobon, A.J.; Sato, M.N. Role of histamine in modulating the immune response and inflammation. Mediat. Inflamm. 2018, 2018, 9524075. [Google Scholar] [CrossRef] [PubMed]
- Sheiner, J.B.; Morris, P.; Anderson, G.H. Food intake suppression by histidine. Pharmacol. Biochem. Behav. 1985, 23, 721–726. [Google Scholar] [CrossRef]
- Vaziri, P.; Dang, K.; Anderson, G.H. Evidence for histamine involvement in the effect of histidine loads on food and water intake in rats. J. Nutr. 1997, 127, 1519–1526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeitzer, J.M. Chapter Six—Control of Sleep and Wakefulness in Health and Disease. In Progress in Molecular Biology and Translational Science; Gillette, M.U., Ed.; Chronobiology: Biological Timing in Health and Disease; Academic Press: Cambridge, MA, USA, 2013; Volume 119, pp. 137–154. [Google Scholar]
- Lin, J.S. Brain structures and mechanisms involved in the control of cortical activation and wakefulness, with emphasis on the posterior hypothalamus and histaminergic neurons. Sleep Med. Rev. 2000, 4, 471–503. [Google Scholar] [CrossRef] [PubMed]
- Simons, F.E.R. Advances in H1-antihistamines. N. Engl. J. Med. 2004, 351, 2203–2217. [Google Scholar] [CrossRef]
- Scammell, T.E.; Jackson, A.C.; Franks, N.P.; Wisden, W.; Dauvilliers, Y. Histamine: Neural circuits and new medications. Sleep 2019, 42, zsy183. [Google Scholar] [CrossRef]
- Abe Details and Download Full Text PDF: Role of Histidine-Related Compounds as Intracellular Proton Buffering Constituents in Vertebrate Muscle. Available online: https://www.pubfacts.com/detail/10951092/Role-of-histidine-related-compounds-as-intracellular-proton-buffering-constituents-in-vertebrate-mus (accessed on 23 March 2018).
- Nair, N.G.; Perry, G.; Smith, M.A.; Reddy, V.P. NMR studies of zinc, copper, and iron binding to histidine, the principal metal ion complexing site of amyloid-β peptide. J. Alzheimers Dis. 2010, 20, 57–66. [Google Scholar] [CrossRef]
- Morris, P.J.; Bruce Martin, R. Stereoselective formation of cobalt(II), nickel(II) and zinc(II) chelates of histidine. J. Inorg. Nucl. Chem. 1970, 32, 2891–2897. [Google Scholar] [CrossRef]
- Schmidt, J.A.; Rinaldi, S.; Scalbert, A.; Ferrari, P.; Achaintre, D.; Gunter, M.J.; Appleby, P.N.; Key, T.J.; Travis, R.C. Plasma concentrations and intakes of amino acids in male meat-eaters, fish-eaters, vegetarians and vegans: A cross-sectional analysis in the EPIC-Oxford cohort. Eur. J. Clin. Nutr. 2016, 70, 306–312. [Google Scholar] [CrossRef] [Green Version]
- Iwasaki, M.; Ishihara, J.; Takachi, R.; Todoriki, H.; Yamamoto, H.; Miyano, H.; Yamaji, T.; Tsugane, S. Validity of a self-administered food-frequency questionnaire for assessing amino acid intake in japan: Comparison with intake from 4-day weighed dietary records and plasma levels. J. Epidemiol. 2016, 26, 36–44. [Google Scholar] [CrossRef] [Green Version]
- Steele, B.F.; Le Bovit, C.B. Leucine and histidine tolerance in the human. J. Nutr. 1951, 45, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Block, W.D.; Westhoff, M.H.; Steele, B.F. Histidine metabolism in the human adult: Histidine blood tolerance, and the effect of continued free L-histidine ingestion on the concentration of imidazole compounds in blood and urine. J. Nutr. 1967, 91, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Henkin, R.I. New aspects in the control of food intake and appetite. Ann. N. Y. Acad. Sci. 1977, 300, 321–334. [Google Scholar] [CrossRef] [PubMed]
- Okubo, H.; Sasaki, S. Histidine intake may negatively correlate with energy intake in human: A cross-sectional study in Japanese female students aged 18 years. J. Nutr. Sci. Vitaminol. (Tokyo) 2005, 51, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, S.; Hamada, M.; Tsuchiya, T.; Okuda, H. Suppression of food intake by histidine-rich protein under low energy intake. Nippon Eiyo Shokuryo Gakkaishi J. Jpn. Soc. Nutr. Food Sci. 2000, 53, 207–214. [Google Scholar] [CrossRef]
- Schechter, P.J.; Prakash, N.J. Failure of oral L-histidine to influence appetite or affect zinc metabolism in man: A double-blind study. Am. J. Clin. Nutr. 1979, 32, 1011–1014. [Google Scholar] [CrossRef] [Green Version]
- Geliebter, A.A.; Hashim, S.A.; Van Itallie, T.B. Oral L-histidine fails to reduce taste and smell acuity but induces anorexia and urinary zinc excretion. Am. J. Clin. Nutr. 1981, 34, 119–120. [Google Scholar] [CrossRef] [Green Version]
- Sasahara, I.; Fujimura, N.; Nozawa, Y.; Furuhata, Y.; Sato, H. The effect of histidine on mental fatigue and cognitive performance in subjects with high fatigue and sleep disruption scores. Physiol. Behav. 2015, 147, 238–244. [Google Scholar] [CrossRef] [Green Version]
- Layrisse, M.; Martínez-Torres, C.; Leets, I.; Taylor, P.; Ramírez, J. Effect of histidine, cysteine, glutathione or beef on iron absorption in humans. J. Nutr. 1984, 114, 217–223. [Google Scholar] [CrossRef]
- Schölmerich, J.; Freudemann, A.; Köttgen, E.; Wietholtz, H.; Steiert, B.; Löhle, E.; Häussinger, D.; Gerok, W. Bioavailability of zinc from zinc-histidine complexes. I. Comparison with zinc sulfate in healthy men. Am. J. Clin. Nutr. 1987, 45, 1480–1486. [Google Scholar] [CrossRef]
- Yoganathan, S.; Sudhakar, S.V.; Arunachal, G.; Thomas, M.; Subramanian, A.; George, R.; Danda, S. Menkes disease and response to copper histidine: An Indian case series. Ann. Indian Acad. Neurol. 2017, 20, 62. [Google Scholar] [CrossRef] [PubMed]
- Jain, P.; Kannan, L.; Chakrabarty, B.; Kumar, A.; Gupta, N.; Kabra, M.; Gulati, S. Menkes disease – An important cause of early onset refractory seizures. J. Pediatr. Neurosci. 2014, 9, 11–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christodoulou, J.; Danks, D.M.; Sarkar, B.; Baerlocher, K.E.; Casey, R.; Horn, N.; Tümer, Z.; Clarke, J.T. Early treatment of Menkes disease with parenteral copper-histidine: Long-term follow-up of four treated patients. Am. J. Med. Genet. 1998, 76, 154–164. [Google Scholar] [CrossRef]
- Yamane, T.; Inoue, Y.; Kobayashi-Hattori, K.; Oishi, Y. Effects of an Amino acid Deficiency on Hyaluronan Synthesis in Human Dermal Fibroblasts. Food Sci. Technol. Res. 2016, 22, 279–281. [Google Scholar] [CrossRef] [Green Version]
- Voorhees, J.J.; Chakrabarti, S.G.; Bernstein, I.A. The metabolism of “histidine-rich” protein in normal and psoriatic keratinization. J. Invest Dermatol 1968, 51, 344–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, S.P.; Brown, S.B.; Griffiths, C.E.; Weller, R.B.; Gibbs, N.K. Feeding filaggrin: Effects of l-histidine supplementation in atopic dermatitis. Clin. Cosmet. Investig. Dermatol. 2017, 10, 403–411. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.-C.; Li, C.-L.; Qi, J.-Y.; Huang, L.-N.; Shi, D.; Du, S.-S.; Liu, L.-Y.; Feng, R.-N.; Sun, C.-H. Relationships of dietary histidine and obesity in northern Chinese adults, an internet-based cross-sectional study. Nutrients 2016, 8, 420. [Google Scholar] [CrossRef] [Green Version]
- Feng, R.N.; Niu, Y.C.; Sun, X.W.; Li, Q.; Zhao, C.; Wang, C.; Guo, F.C.; Sun, C.H.; Li, Y. Histidine supplementation improves insulin resistance through suppressed inflammation in obese women with the metabolic syndrome: A randomised controlled trial. Diabetologia 2013, 56, 985–994. [Google Scholar] [CrossRef]
- Greenberg, A.S.; Obin, M.S. Obesity and the role of adipose tissue in inflammation and metabolism. Am. J. Clin. Nutr. 2006, 83, 461S–465S. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Hsu, C.; Lin, M.; Liu, K.; Yin, M. Histidine and carnosine delay diabetic deterioration in mice and protect human low density lipoprotein against oxidation and glycation. Eur. J. Pharmacol. 2005, 513, 145–150. [Google Scholar] [CrossRef]
- Ookuma, K.; Yoshimatsu, H.; Sakata, T.; Fujimoto, K.; Fukagawa, F. Hypothalamic sites of neuronal histamine action on food intake by rats. Brain Res. 1989, 490, 268–275. [Google Scholar] [CrossRef]
- Kasaoka, S.; Tsuboyama-Kasaoka, N.; Kawahara, Y.; Inoue, S.; Tsuji, M.; Ezaki, O.; Kato, H.; Tsuchiya, T.; Okuda, H.; Nakajima, S. Histidine supplementation suppresses food intake and fat accumulation in rats. Nutrition 2004, 20, 991–996. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, J.C.; Lampart, C.; Rose, C. Histamine formation in rat brain in vivo: Effects of histidine loads. J. Neurochem. 1972, 19, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Yoshimatsu, H.; Chiba, S.; Tajima, D.; Akehi, Y.; Sakata, T. Histidine suppresses food intake through its conversion into neuronal histamine. Exp. Biol. Med. 2002, 227, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Holeček, M.; Vodeničarovová, V. Effects of histidine supplementation on amino acid metabolism in rats. Physiol. Res. 2020, 69, 99–111. [Google Scholar] [CrossRef]
- Rossi, D.J.; Brady, J.D.; Mohr, C. Astrocyte metabolism and signaling during brain ischemia. Nat. Neurosci. 2007, 10, 1377–1386. [Google Scholar] [CrossRef]
- Yiu, G.; He, Z. Glial inhibition of CNS axon regeneration. Nat. Rev. Neurosci. 2006, 7, 617–627. [Google Scholar] [CrossRef] [Green Version]
- Liao, R.; Jiang, L.; Wang, R.; Zhao, H.; Chen, Y.; Li, Y.; Wang, L.; Jie, L.-Y.; Zhou, Y.; Zhang, X.; et al. Histidine provides long-term neuroprotection after cerebral ischemia through promoting astrocyte migration. Sci. Rep. 2015, 5, 15356. [Google Scholar] [CrossRef]
- Zhao, Y.-Y.; Yuan, Y.; Chen, Y.; Jiang, L.; Liao, R.-J.; Wang, L.; Zhang, X.-N.; Ohtsu, H.; Hu, W.-W.; Chen, Z. Histamine promotes locomotion recovery after spinal cord hemisection via inhibiting astrocytic scar formation. CNS Neurosci. Ther. 2015, 21, 454–462. [Google Scholar] [CrossRef]
- Chen, Z.; Li, W.-D.; Zhu, L.-J.; Shen, Y.-J.; Wei, E.-Q. Effects of histidine, a precursor of histamine, on pentylenetetrazole-induced seizures in rats. Acta Pharmacol. Sin. 2002, 23, 361–366. [Google Scholar]
- Wada, Y. Biphasic action of the histamine precursor l-histidine in the rat kindling model of epilepsy. Neurosci. Lett. 1996, 204, 205–208. [Google Scholar] [CrossRef]
- Kaneko, J.; Enya, A.; Enomoto, K.; Ding, Q.; Hisatsune, T. Anserine (beta-alanyl-3-methyl-L-histidine) improves neurovascular-unit dysfunction and spatial memory in aged AβPPswe/PSEN1dE9 Alzheimer’s-model mice. Sci. Rep. 2017, 7, 12571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solomon, J.K.; Geison, R.L. L-histidine-induced hypercholesterolemia: Characteristics of cholesterol biosynthesis in rat livers. Proc. Soc. Exp. Biol. Med. Soc. Exp. Biol. Med. 1978, 159, 44–47. [Google Scholar] [CrossRef] [PubMed]
- Ohmura, E.; Aoyama, Y.; Yoshida, A. Changes in lipids in liver and serum of rats fed a histidine-excess diet or cholesterol-supplemented diets. Lipids 1986, 21, 748–753. [Google Scholar] [CrossRef] [PubMed]
- Harvey, P.W.; Hunsaker, H.A.; Allen, K.G. Dietary L-histidine-induced hypercholesterolemia and hypocupremia in the rat. J. Nutr. 1981, 111, 639–647. [Google Scholar] [CrossRef] [PubMed]
- Muramatsu, K.; Odagiri, H.; Morishita, S.; Takeuchi, H. Effect of excess levels of individual amino acids on growth of rats fed casein diets. J. Nutr. 1971, 101, 1117–1125. [Google Scholar] [CrossRef] [Green Version]
- Benevenga, N.J.; Steele, R.D. Adverse effects of excessive consumption of amino acids. Annu. Rev. Nutr. 1984, 4, 157–181. [Google Scholar] [CrossRef]
- Schirmer, M.D.; Harper, A.E. Adaptive responses of mammalian histidine-degrading enzymes. J. Biol. Chem. 1970, 245, 1204–1211. [Google Scholar]
- Harper, A.E.; Becker, R.V.; Stucki, W.P. Some effects of excessive intakes of indispensable amino acids. Proc. Soc. Exp. Biol. Med. Soc. Exp. Biol. Med. 1966, 121, 695–699. [Google Scholar] [CrossRef]
- Ikezaki, S.; Nishikawa, A.; Furukawa, F.; Imazawa, T.; Enami, T.; Mitsui, M.; Takahashi, M. 13-week subchronic toxicity study of L-histidine monohydrochloride in F344 rats. Eisei Shikenjo Hokoku 1994, 57–63. [Google Scholar]
- Sun, X.; Feng, R.; Li, Y.; Lin, S.; Zhang, W.; Li, Y.; Sun, C.; Li, S. Histidine supplementation alleviates inflammation in the adipose tissue of high-fat diet-induced obese rats via the NF-κB- and PPARγ-involved pathways. Br. J. Nutr. 2014, 112, 477–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snedeker, S.M.; Greger, J.L. Metabolism of Zinc, Copper and Iron as Affected by Dietary Protein, Cysteine and Histidine. J. Nutr. 1983, 113, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Van Campen, D.; Gross, E. Effect of Histidine and Certain Other Amino Acids on the Absorption of Iron-59 by Rats. J. Nutr. 1969, 99, 68–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darwish, H.M.; Cheney, J.C.; Schmitt, R.C.; Ettinger, M.J. Mobilization of copper(II) from plasma components and mechanisms of hepatic copper transport. Am. J. Physiol. 1984, 246, G72–G79. [Google Scholar] [CrossRef]
- Hartter, D.E.; Barnea, A. Brain tissue accumulates 67copper by two ligand-dependent saturable processes. A high affinity, low capacity and a low affinity, high capacity process. J. Biol. Chem. 1988, 263, 799–805. [Google Scholar]
- Mas, A.; Sarkar, B. Uptake of 67Cu by isolated human trophoblast cells. Biochim. Biophys. Acta BBA-Mol. Cell Res. 1992, 1135, 123–128. [Google Scholar] [CrossRef]
- McArdle, H.J.; Guthrie, J.R.; Ackland, M.L.; Danks, D.M. Albumin has no role in the uptake of copper by human fibroblasts. J. Inorg. Biochem. 1987, 31, 123–131. [Google Scholar] [CrossRef]
- Freeman, R.M.; Taylor, P.R. Influence of histidine administration on zinc metabolism in the rat. Am. J. Clin. Nutr. 1977, 30, 523–527. [Google Scholar] [CrossRef] [Green Version]
- Buxani-Rice, S.; Ueda, F.; Bradbury, M.W.B. Transport of Zinc-65 at the Blood-Brain Barrier During Short Cerebrovascular Perfusion in the Rat: Its Enhancement by Histidine. J. Neurochem. 2002, 62, 665–672. [Google Scholar] [CrossRef]
- Wensink, J.; Van den Hamer, C.J. Effect of excess dietary histidine on rate of turnover of 65Zn in brain of rat. Biol. Trace Elem. Res. 1988, 16, 137–150. [Google Scholar] [CrossRef]
- Van Wouwe, J.P.; Hoogenkamp, S.; Van den Hamer, C.J. A histidine supplement and regulation of the zinc status in Swiss random mice. Biol. Trace Elem. Res. 1990, 24, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Frezza, C. Histidine metabolism boosts cancer therapy. Nature 2018, 559, 484–485. [Google Scholar] [CrossRef] [PubMed]
- Kanarek, N.; Keys, H.R.; Cantor, J.R.; Lewis, C.A.; Chan, S.H.; Kunchok, T.; Abu-Remaileh, M.; Freinkman, E.; Schweitzer, L.D.; Sabatini, D.M. Histidine catabolism is a major determinant of methotrexate sensitivity. Nature 2018, 559, 632–636. [Google Scholar] [CrossRef] [PubMed]
- Ikezaki, S.; Nishikawa, A.; Furukawa, F.; Enami, T.; Mitsui, M.; Tanakamaru, Z.; Kim, H.C.; Lee, I.S.; Imazawa, T.; Takahashi, M. Long-term toxicity/carcinogenicity study of L-histidine monohydrochloride in F344 rats. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 1996, 34, 687–691. [Google Scholar] [CrossRef]
- Gullino, P.; Winitz, M.; Birnbaum, S.M.; Cornfield, J.; Otey, M.C.; Greenstein, J.P. Studies on the metabolism of amino acids and related compounds in vivo. I. Toxicity of essential amino acids, individually and in mixtures, and the protective effect of l-arginine. Arch. Biochem. Biophys. 1956, 64, 319–332. [Google Scholar] [CrossRef]
- Figueroa, J.L.; Lewis, A.J.; Miller, P.S.; Fischer, R.L.; Diedrichsen, R.M. Growth, carcass traits, and plasma amino acid concentrations of gilts fed low-protein diets supplemented with amino acids including histidine, isoleucine, and valine. J. Anim. Sci. 2003, 81, 1529–1537. [Google Scholar] [CrossRef] [Green Version]
- Park, S.W.; Kim, C.H.; Kim, J.W.; Shin, H.S.; Paik, I.K.; Kil, D.Y. Effect of dietary supplementation of blood meal and additional magnesium on carnosine and anserine concentrations of pig muscles. Korean J. Food Sci. Anim. Resour. 2014, 34, 252–256. [Google Scholar] [CrossRef] [Green Version]
- Edmonds, M.S.; Baker, D.H. Comparative effects of individual amino acid excesses when added to a corn-soybean meal diet: Effects on growth and dietary choice in the chick. J. Anim. Sci. 1987, 65, 699–705. [Google Scholar] [CrossRef] [Green Version]
- Auh, J.; Namgung, N.; Shin, K.; Park, S.; Paik, I. Effects of supplementary blood meal on the content of carnosine and anserine in broiler meat. J. Poult. Sci. 2010, 47, 302–309. [Google Scholar] [CrossRef] [Green Version]
- Park, S.W.; Kim, C.H.; Namgung, N.; Jung, B.Y.; Paik, I.K.; Kil, D.Y. Effects of dietary supplementation of histidine, β-alanine, magnesium oxide, and blood meal on carnosine and anserine concentrations of broiler breast meat. J. Poult. Sci. 2013, 50, 251–256. [Google Scholar] [CrossRef] [Green Version]
- Kralik, G.; Sak-Bosnar, M.; Kralik, Z.; Galović, O.; Grčević, M.; Kralik, I. Effect of β-alanine and l-histidine on concentration of carnosine in muscle tissue and oxidative stability of chicken meat. Poljoprivreda 2015, 21, 190–194. [Google Scholar] [CrossRef]
- Haug, A.; R⊘dbotten, R.; Mydland, L.T.; Christophersen, O.A. Increased broiler muscle carnosine and anserine following histidine supplementation of commercial broiler feed concentrate. Acta Agric. Scand. Sect.-Anim. Sci. 2008, 58, 71–77. [Google Scholar] [CrossRef]
- Kai, S.; Watanabe, G.; Kubota, M.; Kadowaki, M.; Fujimura, S. Effect of dietary histidine on contents of carnosine and anserine in muscles of broilers. Anim. Sci. J. Nihon Chikusan Gakkaiho 2015, 86, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Kopeć, W.; Jamroz, D.; Wiliczkiewicz, A.; Biazik, E.; Pudlo, A.; Hikawczuk, T.; Skiba, T.; Korzeniowska, M. Influence of different histidine sources and zinc supplementation of broiler diets on dipeptide content and antioxidant status of blood and meat. Br. Poult. Sci. 2013, 54, 454–465. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.-D.; Qu, B.; Feng, L.; Jiang, J.; Kuang, S.-Y.; Wu, P.; Tang, L.; Tang, W.-N.; Zhang, Y.-A.; Zhou, X.-Q.; et al. Histidine prevents cu-induced oxidative stress and the associated decreases in mrna from encoding tight junction proteins in the intestine of grass carp (Ctenopharyngodon idella). PLoS ONE 2016, 11, e0157001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, R.P.; Poe, W.E.; Robinson, E.H. Leucine, isoleucine, valine and histidine requirements of fingerling channel catfish. J. Nutr. 1980, 110, 627–633. [Google Scholar] [CrossRef]
- Michelato, M.; Zaminhan, M.; Boscolo, W.R.; Nogaroto, V.; Vicari, M.; Artoni, R.F.; Furuya, V.R.B.; Furuya, W.M. Dietary histidine requirement of Nile tilapia juveniles based on growth performance, expression of muscle-growth-related genes and haematological responses. Aquaculture 2017, 467, 63–70. [Google Scholar] [CrossRef]
- Abe, H.; Dobson, G.P.; Hoeger, U.; Parkhouse, W.S. Role of histidine-related compounds to intracellular buffering in fish skeletal muscle. Am. J. Physiol. 1985, 249, R449–R454. [Google Scholar] [CrossRef]
- Breck, O.; BjerkÅs, E.; Campbell, P.; Arnesen, P.; Haldorsen, P.; Waagbø, R. Cataract preventative role of mammalian blood meal, histidine, iron and zinc in diets for Atlantic salmon (Salmo salar L.) of different strains. Aquac. Nutr. 2003, 9, 341–350. [Google Scholar] [CrossRef]
- Bjerkås, E.; Breck, O.; Waagbo, R. The role of nutrition in cataract formation in farmed fish. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2006, 1. [Google Scholar] [CrossRef]
- Remø, S.C.; Olsvik, P.A.; Torstensen, B.E.; Amlund, H.; Breck, O.; Waagbø, R. Susceptibility of Atlantic salmon lenses to hydrogen peroxide oxidation ex vivo after being fed diets with vegetable oil and methylmercury. Exp. Eye Res. 2011, 92, 414–424. [Google Scholar] [CrossRef] [PubMed]
- Remø, S.C.; Hevrøy, E.M.; Olsvik, P.A.; Fontanillas, R.; Breck, O.; Waagbø, R. Dietary histidine requirement to reduce the risk and severity of cataracts is higher than the requirement for growth in Atlantic salmon smolts, independently of the dietary lipid source. Br. J. Nutr. 2014, 111, 1759–1772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trösse, C.; Waagbø, R.; Breck, O.; Stavrum, A.-K.; Petersen, K.; Olsvik, P.A. Genome-wide transcription analysis of histidine-related cataract in Atlantic salmon (Salmo salar L). Mol. Vis. 2009, 15, 1332–1350. [Google Scholar]
- Waagbø, R.; Trösse, C.; Koppe, W.; Fontanillas, R.; Breck, O. Dietary histidine supplementation prevents cataract development in adult Atlantic salmon, Salmo salar L., in seawater. Br. J. Nutr. 2010, 104, 1460–1470. [Google Scholar] [CrossRef] [Green Version]
- Schoof, A.; Gabel, M.; Voigt, J.; Schönhusen, U.; Kluth, H. Investigations on the influence of duodenal histidine infusion on nitrogen and amino acid turnover of growing German Holstein bulls. Arch. Tierernahr. 2000, 53, 303–321. [Google Scholar] [CrossRef] [PubMed]
- Alderman, G.; Jarrige, R. Feed Evaluation and Protein Requirement Systems for Ruminants. Available online: https://publications.europa.eu/en/publication-detail/-/publication/72fd322e-2083-4a19-9105-758eda4faf9b/language-en (accessed on 2 March 2018).
- Gabel, M.; Poppe, S. Intestinal amino acid in supply in relation to the amino acid requirements in growing bulls. Arch. Tierernahr. 1986, 36, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Hu, H.; Zheng, N.; Wang, J. Leucine and histidine independently regulate milk protein synthesis in bovine mammary epithelial cells via mTOR signaling pathway. J. Zhejiang Univ. Sci. B 2015, 16, 560–572. [Google Scholar] [CrossRef] [PubMed]
- Prizant, R.L.; Barash, I. Negative effects of the amino acids Lys, His, and Thr on S6K1 phosphorylation in mammary epithelial cells. J. Cell. Biochem. 2008, 105, 1038–1047. [Google Scholar] [CrossRef]
- Toerien, C.A.; Trout, D.R.; Cant, J.P. Nutritional stimulation of milk protein yield of cows is associated with changes in phosphorylation of mammary eukaryotic initiation factor 2 and ribosomal s6 kinase 1. J. Nutr. 2010, 140, 285–292. [Google Scholar] [CrossRef] [Green Version]
- Doelman, J.; Purdie, N.G.; Osborne, V.R.; Cant, J.P. Short communication: The effects of histidine-supplemented drinking water on the performance of lactating dairy cows. J. Dairy Sci. 2008, 91, 3998–4001. [Google Scholar] [CrossRef] [Green Version]
- Hadrová, S.; Krizova, L.; Richter, M.; Třináctý, J.; Dračková, M. The effect of duodenal infusion of histidine on milk yield, milk composition, and plasma amino acids in dairy cows. J. Anim. Feed Sci. 2012, 21, 555–565. [Google Scholar] [CrossRef] [Green Version]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moro, J.; Tomé, D.; Schmidely, P.; Demersay, T.-C.; Azzout-Marniche, D. Histidine: A Systematic Review on Metabolism and Physiological Effects in Human and Different Animal Species. Nutrients 2020, 12, 1414. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12051414
Moro J, Tomé D, Schmidely P, Demersay T-C, Azzout-Marniche D. Histidine: A Systematic Review on Metabolism and Physiological Effects in Human and Different Animal Species. Nutrients. 2020; 12(5):1414. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12051414
Chicago/Turabian StyleMoro, Joanna, Daniel Tomé, Philippe Schmidely, Tristan-Chalvon Demersay, and Dalila Azzout-Marniche. 2020. "Histidine: A Systematic Review on Metabolism and Physiological Effects in Human and Different Animal Species" Nutrients 12, no. 5: 1414. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12051414