Maternal Diet Influences the Reinstatement of Cocaine-Seeking Behavior and the Expression of Melanocortin-4 Receptors in Female Offspring of Rats


Abstract
:1. Introduction
2. Materials and Methods
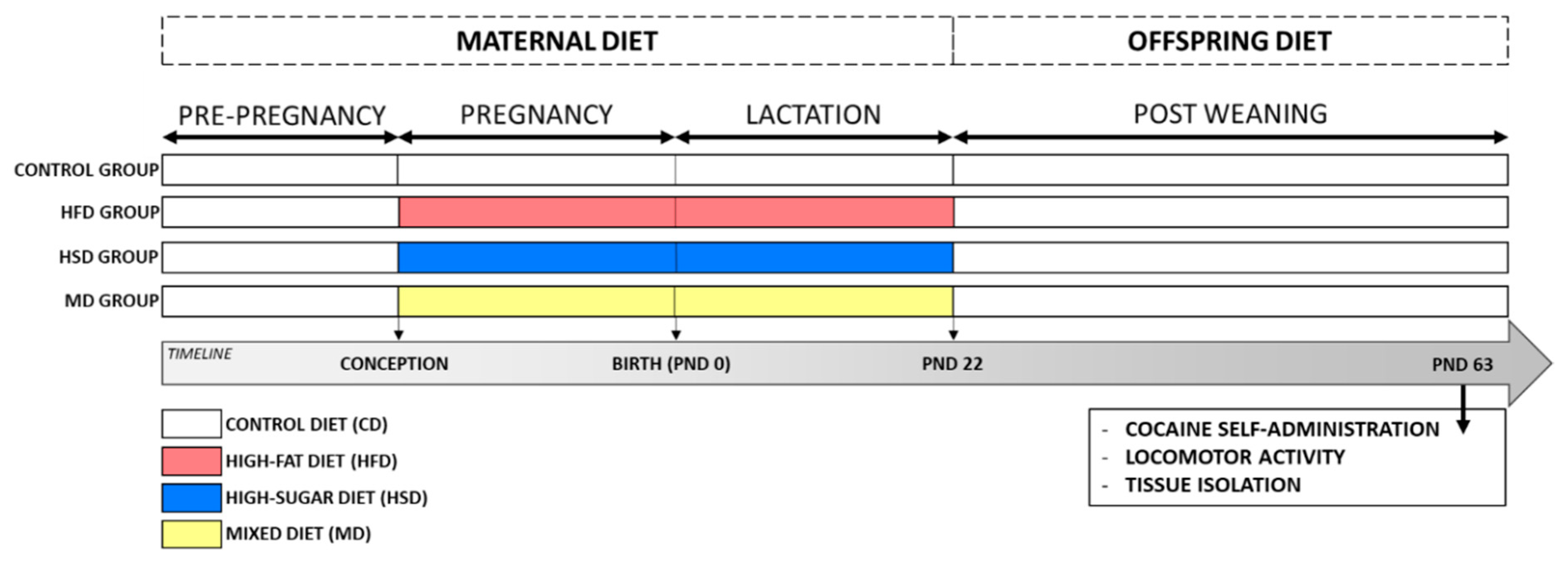
2.1. Animals and Diets
2.2. Behavioral Procedures
2.2.1. Drugs
2.2.2. Cocaine Self-Administration Procedure
Catheter Implantation and Initial Lever-Press Training
Cocaine Self-Administration
Progressive Ratio Test
Extinction
Reinstatement of Cocaine-Seeking
2.2.3. Locomotor Activity
2.3. Biochemical Analysis
2.3.1. Brain Tissue Collection
2.3.2. Melancortin-4 Receptor Expression
2.4. Statistical Analysis
3. Results
3.1. Maternal Body Weight, Caloric Intake and Litter Size
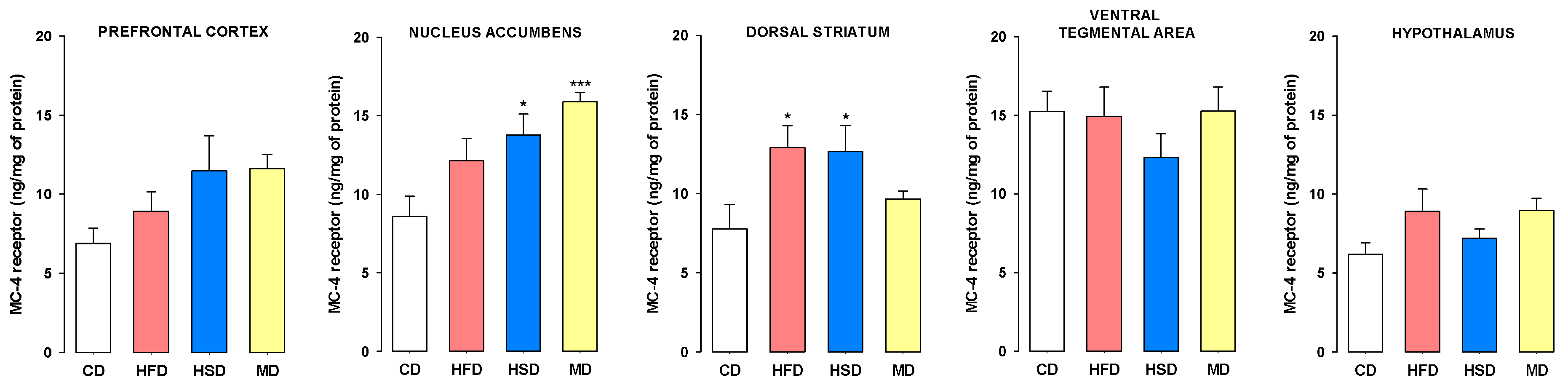
3.2. Expression of MC-4 Receptors
3.3. Locomotor Activity
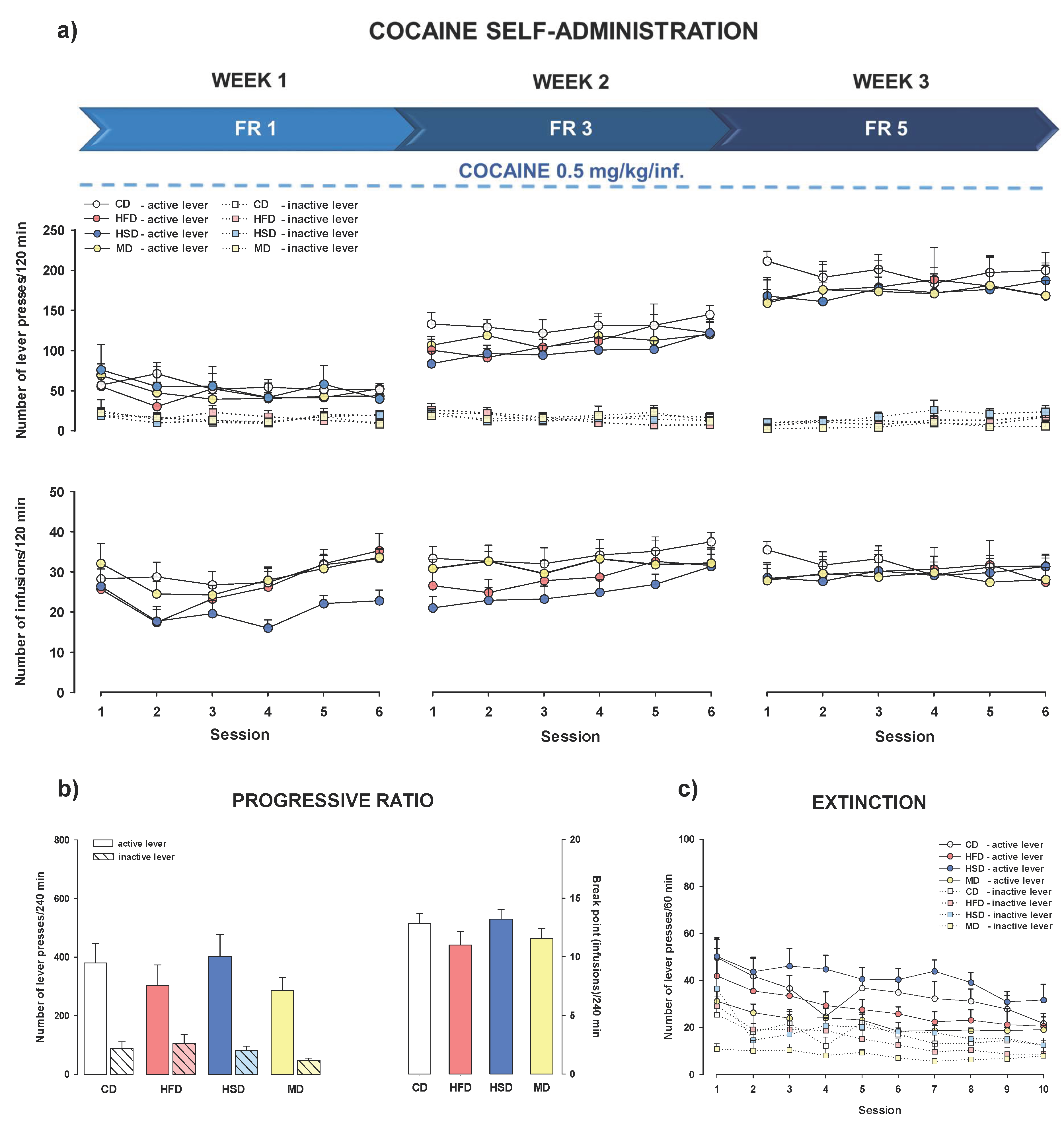
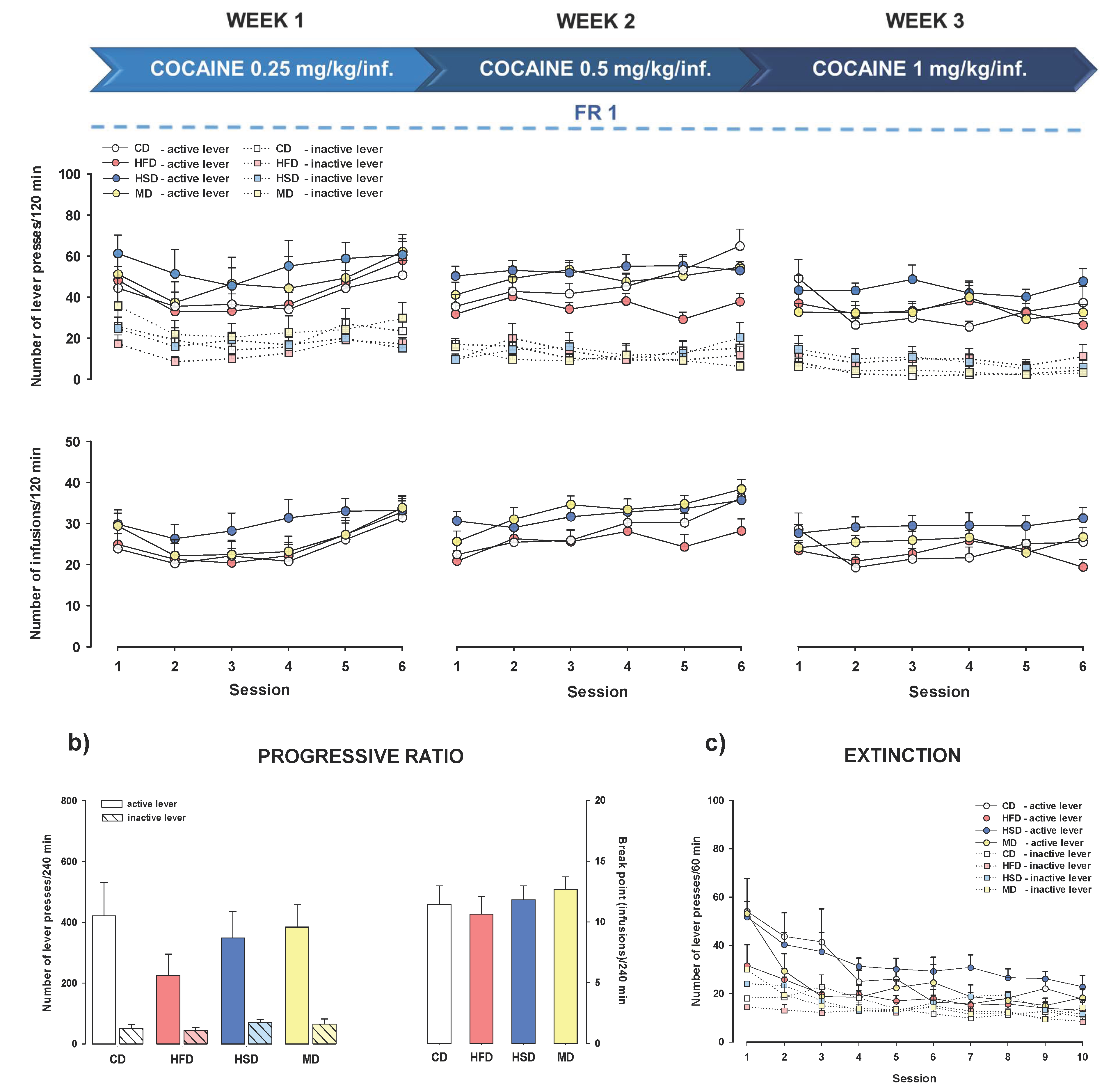
3.4. Cocaine Self-Administration
3.4.1. Stable Cocaine Dose and Increased Reinforcement Schedule
3.4.2. Increased Cocaine Dose and Stable Reinforcement Schedule
3.5. PR Schedule of Cocaine Reinforcement
3.6. Extinction Training
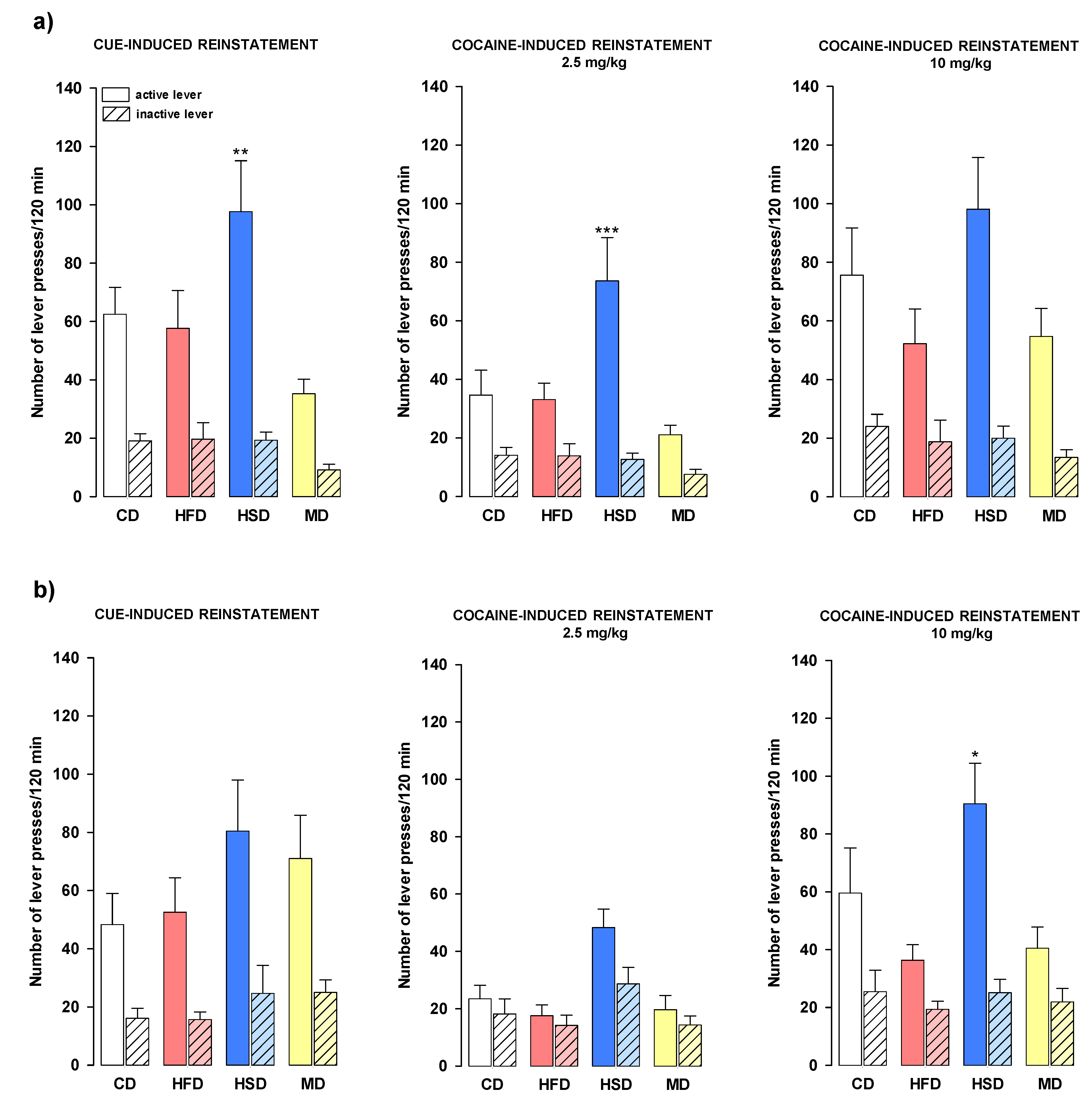
3.7. Reinstatement of Cocaine-Seeking Behavior
3.7.1. Relapse after Stable Cocaine Dose and Increased Reinforcement Schedule
3.7.2. Relapse after Increased Cocaine Dose and Stable Reinforcement Schedule
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Degenhardt, L.; Charlson, F.; Ferrari, A.; Santomauro, D.; Erskine, H.; Mantilla-Herrara, A.; Whiteford, H.; Leung, J.; Naghavi, M.; Griswold, M.; et al. The global burden of disease attributable to alcohol and drug use in 195 countries and territories, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Psychiatry 2018, 5, 987–1012. [Google Scholar] [CrossRef] [Green Version]
- O’Rourke, M.; Wrigley, C.; Hammond, S. Violence within mental health services: How to enhance risk management. Risk Manag. Healthc. Policy 2018, 11, 159–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uher, R.; Zwicker, A. Etiology in psychiatry: Embracing the reality of poly-gene-environmental causation of mental illness. World Psychiatry 2017, 16, 121–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukhara, D.; Banks, M.L.; Neigh, G.N. Stress as a risk factor for substance use disorders: A mini-review of molecular mediators. Front. Behav. Neurosci. 2018, 12, 309. [Google Scholar] [CrossRef] [Green Version]
- Edlow, A.G. Maternal obesity and neurodevelopmental and psychiatric disorders in offspring. Prenat. Diagn. 2017, 37, 95–110. [Google Scholar] [CrossRef] [Green Version]
- Gawlińska, K.; Gawliński, D.; Przegaliński, E.; Filip, M. Maternal high-fat diet during pregnancy and lactation provokes depressive-like behavior and influences the irisin/brain-derived neurotrophic factor axis and inflammatory factors in male and female offspring in rats. J. Physiol. Pharmacol. 2019, 70, 407–411. [Google Scholar] [CrossRef]
- Rivera, H.M.; Christiansen, K.J.; Sullivan, E.L. The role of maternal obesity in the risk of neuropsychiatric disorders. Front. Neurosci. 2015, 9, 194. [Google Scholar] [CrossRef]
- Sullivan, E.L.; Nousen, E.K.; Chamlou, K.A. Maternal high fat diet consumption during the perinatal period programs offspring behavior. Physiol. Behav. 2014, 123, 236–242. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, E.L.; Riper, K.M.; Lockard, R.; Valleau, J.C. Maternal high-fat diet programming of the neuroendocrine system and behavior. Horm. Behav. 2015, 76, 153–161. [Google Scholar] [CrossRef] [Green Version]
- Naef, L.; Moquin, L.; Dal Bo, G.; Giros, B.; Gratton, A.; Walker, C.D. Maternal high-fat intake alters presynaptic regulation of dopamine in the nucleus accumbens and increases motivation for fat rewards in the offspring. Neuroscience 2011, 176, 225–236. [Google Scholar] [CrossRef]
- Ong, Z.Y.; Muhlhausler, B.S. Maternal “junk-food” feeding of rat dams alters food choices and development of the mesolimbic reward pathway in the offspring. FASEB J. 2011, 25, 2167–2179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paradis, J.; Boureau, P.; Moyon, T.; Nicklaus, S.; Parnet, P.; Paillé, V. Perinatal western diet consumption leads to profound plasticity and GABAergic phenotype changes within hypothalamus and reward pathway from birth to sexual maturity in rat. Front. Endocrinol. 2017, 8, 216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karatayev, O.; Lukatskaya, O.; Moon, S.H.; Guo, W.R.; Chen, D.; Algava, D.; Abedi, S.; Leibowitz, S.F. Nicotine and ethanol co-use in Long-Evans rats: Stimulatory effects of perinatal exposure to a fat-rich diet. Alcohol 2015, 49, 479–489. [Google Scholar] [CrossRef] [Green Version]
- Morganstern, I.; Lukatskaya, O.; Moon, S.-H.; Guo, W.-R.; Shaji, J.; Karatayev, O.; Leibowitz, S.F. Stimulation of nicotine reward and central cholinergic activity in Sprague–Dawley rats exposed perinatally to a fat-rich diet. Psychopharmacology 2013, 230, 509–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naef, L.; Srivastava, L.; Gratton, A.; Hendrickson, H.; Owens, S.M.; Walker, C.D. Maternal high fat diet during the perinatal period alters mesocorticolimbic dopamine in the adult rat offspring: Reduction in the behavioral responses to repeated amphetamine administration. Psychopharmacology (Berl.) 2008, 197, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Bocarsly, M.E.; Barson, J.R.; Hauca, J.M.; Hoebel, B.G.; Leibowitz, S.F.; Avena, N.M. Effects of perinatal exposure to palatable diets on body weight and sensitivity to drugs of abuse in rats. Physiol. Behav. 2012, 107, 568–575. [Google Scholar] [CrossRef] [Green Version]
- Favrod-Coune, T.; Broers, B. The Health Effect of Psychostimulants: A Literature Review. Pharmaceuticals 2010, 3, 2333–2361. [Google Scholar] [CrossRef] [Green Version]
- Alvaro, J.D.; Tatro, J.B.; Quillan, J.M.; Fogliano, M.; Eisenhard, M.; Lerner, M.R.; Nestler, E.J.; Duman, R.S. Morphine down-regulates melanocortin-4 receptor expression in brain regions that mediate opiate addiction. Mol. Pharmacol. 1996, 50, 583–591. [Google Scholar]
- Lindblom, J.; Schiöth, H.B.; Larsson, A.; Wikberg, J.E.S.; Bergström, L. Autoradiographic discrimination of melanocortin receptors indicates that the MC3 subtype dominates in the medial rat brain. Brain Res. 1998, 810, 161–171. [Google Scholar] [CrossRef]
- Adan, R.A.H.; Tiesjema, B.; Hillebrand, J.J.G.; La Fleur, S.E.; Kas, M.J.H.; De Krom, M. The MC4 receptor and control of appetite. Br. J. Pharmacol. 2006, 149, 815–827. [Google Scholar] [CrossRef] [Green Version]
- Chaki, S.; Okuyama, S. Involvement of melanocortin-4 receptor in anxiety and depression. Peptides 2005, 26, 1952–1964. [Google Scholar] [CrossRef]
- Cone, R.D. Anatomy and regulation of the central melanocortin system. Nat. Neurosci. 2005, 8, 571–578. [Google Scholar] [CrossRef]
- Gawliński, D.; Gawlińska, K.; Frankowska, M.; Filip, M. Maternal high-sugar diet changes offspring vulnerability to reinstatement of cocaine-seeking behavior: Role of melanocortin-4 receptors. FASEB J. (in press). [CrossRef]
- Jastrzȩbska, J.; Frankowska, M.; Szumiec, Ł.; Sadakierska-Chudy, A.; Haduch, A.; Smaga, I.; Bystrowska, B.; Daniel, W.A.; Filip, M. Cocaine self-administration in Wistar-Kyoto rats: A behavioral and biochemical analysis. Behav. Brain Res. 2015, 293, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Frankowska, M.; Miszkiel, J.; Pomierny-Chamioło, L.; Pomierny, B.; Giannotti, G.; Suder, A.; Filip, M. Alternation in dopamine D2-like and metabotropic glutamate type 5 receptor density caused by differing housing conditions during abstinence from cocaine self-administration in rats. J. Psychopharmacol. 2019, 33, 372–382. [Google Scholar] [CrossRef] [PubMed]
- Richardson, N.R.; Roberts, D.C.S. Progressive ratio schedules in drug self-administration studies in rats: A method to evaluate reinforcing efficacy. J. Neurosci. Methods 1996, 66, 1–11. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 4th ed.; Academic Press: San Diego, CA, USA, 1998. [Google Scholar]
- Kamat, P.K.; Kalani, A.; Tyagi, N. Method and validation of synaptosomal preparation for isolation of synaptic membrane proteins from rat brain. MethodsX 2014, 1, 102–107. [Google Scholar] [CrossRef]
- Becker, J.B.; McClellan, M.L.; Reed, B.G. Sex differences, gender and addiction. J. Neurosci. Res. 2017, 95, 136–147. [Google Scholar] [CrossRef] [Green Version]
- Kesse, E.; Clavel-Chapelon, F.; Slimani, N.; van Liere, M.; E3N Group. Do eating habits differ according to alcohol consumption? Results of a study of the French cohort of the European Prospective Investigation into Cancer and Nutrition (E3N-EPIC). Am. J. Clin. Nutr. 2001, 74, 322–327. [Google Scholar] [CrossRef]
- Sansone, R.A.; Sansone, L.A. Obesity and substance misuse: Is there a relationship? Innov. Clin. Neurosci. 2013, 10, 30–35. [Google Scholar]
- Hussaini, A.E.; Nicholson, L.M.; Shera, D.; Stettler, N.; Kinsman, S. Adolescent Obesity as a Risk Factor for High-Level Nicotine Addiction in Young Women. J. Adolesc. Health 2011, 49, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Chang, G.-Q.; Gaysinskaya, V.; Karatayev, O.; Leibowitz, S.F. Maternal high-fat diet and fetal programming: Increased proliferation of hypothalamic peptide-producing neurons that increase risk for overeating and obesity. J. Neurosci. 2008, 28, 12107–12119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabanes, A.; de Assis, S.; Gustafsson, J.-A.; Hilakivi-Clarke, L. Maternal High n-6 Polyunsaturated Fatty Acid Intake during Pregnancy Increases Voluntary Alcohol Intake and Hypothalamic Estrogen Receptor Alpha and Beta Levels among Female Offspring. Dev. Neurosci. 2000, 22, 488–493. [Google Scholar] [CrossRef] [PubMed]
- Peleg-Raibstein, D.; Sarker, G.; Litwan, K.; Krämer, S.D.; Ametamey, S.M.; Schibli, R.; Wolfrum, C. Enhanced sensitivity to drugs of abuse and palatable foods following maternal overnutrition. Transl. Psychiatry 2016, 6, e911. [Google Scholar] [CrossRef]
- McLellan, A.T.; Lewis, D.C.; O’Brien, C.P.; Kleber, H.D. Drug dependence, a chronic medical illness implications for treatment, insurance, and outcomes evaluation. J. Am. Med. Assoc. 2000, 284, 1689–1695. [Google Scholar] [CrossRef]
- Hecht, G.S.; Spear, N.E.; Spear, L.P. Alterations in the reinforcing efficacy of cocaine in adult rats following prenatal exposure to cocaine. Behav. Neurosci. 1998, 112, 410–418. [Google Scholar] [CrossRef]
- Keller, R.W.; LeFevre, R.; Raucci, J.; Carlson, J.N.; Glick, S.D. Enhanced cocaine self-administration in adult rats prenatally exposed to cocaine. Neurosci. Lett. 1996, 205, 153–156. [Google Scholar] [CrossRef]
- Rocha, B.A.; Mead, A.N.; Kosofsky, B.E. Increased vulnerability to self-administer cocaine in mice prenatally exposed to cocaine. Psychopharmacology 2002, 163, 221–229. [Google Scholar] [CrossRef]
- Kelley, B.M.; Middaugh, L.D. Ethanol self-administration and motor deficits in adult C57BL/6J mice exposed prenatally to cocaine. Pharmacol. Biochem. Behav. 1996, 55, 575–584. [Google Scholar] [CrossRef]
- Delaney-Black, V.; Chiodo, L.M.; Hannigan, J.H.; Greenwald, M.K.; Janisse, J.; Patterson, G.; Huestis, M.A.; Partridge, R.T.; Ager, J.; Sokol, R.J. Prenatal and postnatal cocaine exposure predict teen cocaine use. Neurotoxicol. Teratol. 2011, 33, 110–119. [Google Scholar] [CrossRef] [Green Version]
- Frank, D.A.; Rose-Jacobs, R.; Crooks, D.; Cabral, H.J.; Gerteis, J.; Hacker, K.A.; Martin, B.; Weinstein, Z.B.; Heeren, T. Adolescent initiation of licit and illicit substance use: Impact of intrauterine exposures and post-natal exposure to violence. Neurotoxicol. Teratol. 2011, 33, 100–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minnes, S.; Singer, L.; Min, M.O.; Wu, M.; Lang, A.; Yoon, S. Effects of prenatal cocaine/polydrug exposure on substance use by age 15. Drug Alcohol. Depend. 2014, 134, 201–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richardson, G.A.; De Genna, N.M.; Goldschmidt, L.; Larkby, C.; Donovan, J.E. Prenatal cocaine exposure: Direct and indirect associations with 21-year-old offspring substance use and behavior problems. Drug Alcohol Depend. 2019, 195, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Bassareo, V.; Cucca, F.; Frau, R.; Di Chiara, G. Differential activation of accumbens shell and core dopamine by sucrose reinforcement with nose poking and with lever pressing. Behav. Brain Res. 2015, 294, 215–223. [Google Scholar] [CrossRef]
- Bassareo, V.; Cucca, F.; Frau, R.; Di Chiara, G. Monitoring dopamine transmission in the rat nucleus accumbens shell and core during acquisition of nose-poking for sucrose. Behav. Brain Res. 2015, 287, 200–206. [Google Scholar] [CrossRef]
- Hajnal, A.; Smith, G.P.; Norgren, R. Oral sucrose stimulation increases accumbens dopamine in the rat. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 286, 31–37. [Google Scholar] [CrossRef]
- Rada, P.; Avena, N.M.; Hoebel, B.G. Daily bingeing on sugar repeatedly releases dopamine in the accumbens shell. Neuroscience 2005, 134, 737–744. [Google Scholar] [CrossRef]
- Bello, N.T.; Lucas, L.R.; Hajnal, A. Repeated sucrose access influences dopamine D2 receptor density in the striatum. Neuroreport 2002, 13, 1575–1578. [Google Scholar] [CrossRef]
- Colantuoni, C.; Schwenker, J.; McCarthy, J.; Rada, P.; Ladenheim, B.; Cadet, J.-L.; Schwartz, G.J.; Moran, T.H.; Hoebel, B.G. Excessive sugar intake alters binding to dopamine and mu-opioid receptors in the brain. Neuroreport 2001, 12, 3549–3552. [Google Scholar] [CrossRef]
- Spangler, R.; Wittkowski, K.M.; Goddard, N.L.; Avena, N.M.; Hoebel, B.G.; Leibowitz, S.F. Opiate-like effects of sugar on gene expression in reward areas of the rat brain. Mol. Brain Res. 2004, 124, 134–142. [Google Scholar] [CrossRef]
- Ahmed, S.; Kashem, M.A.; Sarker, R.; Ahmed, E.U.; Hargreaves, G.A.; McGregor, I.S. Neuroadaptations in the striatal proteome of the rat following prolonged excessive sucrose intake. Neurochem. Res. 2014, 39, 815–824. [Google Scholar] [CrossRef] [PubMed]
- Avena, N.M.; Long, K.A.; Hoebel, B.G. Sugar-dependent rats show enhanced responding for sugar after abstinence: Evidence of a sugar deprivation effect. Physiol. Behav. 2005, 84, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Grimm, J.W.; Fyall, A.M.; Osincup, D.P. Incubation of sucrose craving: Effects of reduced training and sucrose pre-loading. Physiol. Behav. 2005, 84, 73–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayol, S.A.; Farrington, S.J.; Stickland, N.C. A maternal “junk food” diet in pregnancy and lactation promotes an exacerbated taste for “junk food” and a greater propensity for obesity in rat offspring. Br. J. Nutr. 2007, 98, 843–851. [Google Scholar] [CrossRef] [Green Version]
- Gosnell, B.A. Sucrose intake enhances behavioral sensitization produced by cocaine. Brain Res. 2005, 1031, 194–201. [Google Scholar] [CrossRef]
- Lenoir, M.; Serre, F.; Cantin, L.; Ahmed, S.H. Intense sweetness surpasses cocaine reward. PLoS ONE 2007, 2, e698. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.Q.; LE, Q.M.; Yu, X.C.; Ma, L.; Wang, F.F. Sucrose reward promotes rats’ motivation for cocaine. Sheng Li Xue Bao 2016, 68, 233–240. [Google Scholar] [CrossRef]
- Caprioli, D.; Zeric, T.; Thorndike, E.B.; Venniro, M. Persistent palatable food preference in rats with a history of limited and extended access to methamphetamine self-administration. Addict. Biol. 2015, 20, 913–926. [Google Scholar] [CrossRef] [Green Version]
- Culleré, M.E.; Spear, N.E.; Molina, J.C. Prenatal ethanol increases sucrose reinforcement, an effect strengthened by postnatal association of ethanol and sucrose. Alcohol 2014, 48, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Pacchioni, A.M.; Gabriele, A.; See, R.E. Dorsal striatum mediation of cocaine-seeking after withdrawal from short or long daily access cocaine self-administration in rats. Behav. Brain Res. 2011, 218, 296–300. [Google Scholar] [CrossRef] [Green Version]
- Scofield, M.D.; Heinsbroek, J.A.; Gipson, C.D.; Kupchik, Y.M.; Spencer, S.; Smith, A.C.W.; Roberts-Wolfe, D.; Kalivas, P.W. The Nucleus Accumbens: Mechanisms of Addiction across Drug Classes Reflect the Importance of Glutamate Homeostasis. Pharmacol. Rev. 2016, 68, 816–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, L.T.; Saad, S.; Tan, Y.; Pollock, C.; Chen, H. Maternal high-fat diet induces metabolic stress response disorders in offspring hypothalamus. J. Mol. Endocrinol. 2017, 59, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Everitt, B.J.; Wolf, M.E. Psychomotor stimulant addiction: A neural systems perspective. J. Neurosci. 2002, 22, 3312–3320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farrell, M.R.; Schoch, H.; Mahler, S.V. Modeling cocaine relapse in rodents: Behavioral considerations and circuit mechanisms. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2018, 87, 33–47. [Google Scholar] [CrossRef]
- Stewart, J. Psychological and neural mechanisms of relapse. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2008, 363, 3147–3158. [Google Scholar] [CrossRef] [Green Version]
- Alhadeff, A.L.; Goldstein, N.; Park, O.; Klima, M.L.; Vargas, A.; Betley, J.N. Natural and Drug Rewards Engage Distinct Pathways that Converge on Coordinated Hypothalamic and Reward Circuits. Neuron 2019, 103, 891–908. [Google Scholar] [CrossRef]
- Alvaro, J.D.; Taylor, J.R.; Duman, R.S.; Iadarola, M.; Nestler, E. Molecular and behavioral interactions between central melanocortins and cocaine. J. Pharmacol. Exp. Ther. 2003, 304, 391–399. [Google Scholar] [CrossRef] [Green Version]
- Hsu, R.; Taylor, J.R.; Newton, S.S.; Alvaro, J.D.; Haile, C.; Han, G.; Hruby, V.J.; Nestler, E.J.; Duman, R.S. Blockade of melanocortin transmission inhibits cocaine reward. Eur. J. Neurosci. 2005, 21, 2233–2242. [Google Scholar] [CrossRef] [Green Version]
- Alserda, E.; Adan, R.A.H.; Ramakers, G.M.J. Repeated agouti related peptide (83–132) injections inhibit cocaine-induced locomotor sensitisation, but not via the nucleus accumbens. Eur. J. Pharmacol. 2013, 719, 187–191. [Google Scholar] [CrossRef]
- Cui, H.; Lutter, M. The expression of MC4Rs in D1R neurons regulates food intake and locomotor sensitization to cocaine. Genes Brain Behav. 2013, 12, 658–665. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, M.O.; Bober, J.; Ferreira, J.G.; Tellez, L.A.; Mineur, Y.S.; Souza, D.O.; Gao, X.-B.; Picciotto, M.R.; Araújo, I.; Liu, Z.-W.; et al. AgRP neurons regulate development of dopamine neuronal plasticity and nonfood-associated behaviors. Nat. Neurosci. 2012, 15, 1108–1110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, B.K.; Huang, K.W.; Grueter, B.A.; Rothwell, P.E.; Malenka, R.C. Anhedonia requires MC4R-mediated synaptic adaptations in nucleus accumbens. Nature 2012, 487, 183–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindblom, J.; Opmane, B.; Mutulis, F.; Mutule, I.; Petrovska, R.; Klusa, V.; Bergström, L.; Wikberg, J.E. The MC4 receptor mediates alpha-MSH induced release of nucleus accumbens dopamine. Neuroreport 2001, 12, 2155–2158. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, M.S.; Barontini, M.; Armando, I.; Celis, M.E. Correlation of increased grooming behavior and motor activity with alterations in nigrostriatal and mesolimbic catecholamines after alpha-melanotropin and neuropeptide glutamine-isoleucine injection in the rat ventral tegmental area. Cell. Mol. Neurobiol. 2001, 21, 523–533. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control Diet (CD) | High-Fat Diet (HFD) | High-Sugar Diet (HSD) | Mixed Diet (MD) | |
|---|---|---|---|---|
| Carbohydrate Sucrose | 65% 4.6% | 25% | 70% 44% | 56% 18% |
| Fat | 13% | 60% | 12% | 28% |
| Protein | 22% | 15% | 18% | 16% |
| Total energy | 3.4 kcal/g | 5.3 kcal/g | 3.8 kcal/g | 3.9 kcal/g |
| CD | HFD | HSD | MD | |||
|---|---|---|---|---|---|---|
| Distance traveled (cm) | 5 min | 1064.93 ± 63.75 | 1031.16 ± 56.43 | 1112.13 ± 71.78 | 1018.53 ± 60.86 | F(3, 44) = 0.435, p = 0.729 |
| 30 min | 3130.32 ± 214.53 | 3332.69 ± 223.62 | 3210.91 ± 145.59 | 3070.44 ± 217.69 | F(3, 44) = 0.313, p = 0.816 | |
| 120 min | 5262.90 ± 380.66 | 4979.64 ± 284.89 | 5615.37 ± 603.51 | 5392.65 ± 562.38 | F(3, 44) = 0.311, p = 0.817 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gawliński, D.; Gawlińska, K.; Frankowska, M.; Filip, M. Maternal Diet Influences the Reinstatement of Cocaine-Seeking Behavior and the Expression of Melanocortin-4 Receptors in Female Offspring of Rats. Nutrients 2020, 12, 1462. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12051462
Gawliński D, Gawlińska K, Frankowska M, Filip M. Maternal Diet Influences the Reinstatement of Cocaine-Seeking Behavior and the Expression of Melanocortin-4 Receptors in Female Offspring of Rats. Nutrients. 2020; 12(5):1462. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12051462
Chicago/Turabian StyleGawliński, Dawid, Kinga Gawlińska, Małgorzata Frankowska, and Małgorzata Filip. 2020. "Maternal Diet Influences the Reinstatement of Cocaine-Seeking Behavior and the Expression of Melanocortin-4 Receptors in Female Offspring of Rats" Nutrients 12, no. 5: 1462. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12051462