Synergistic Effect of Eugenol and Probiotic Lactobacillus Plantarum Zs2058 against Salmonella Infection in C57bl/6 Mice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacteria and Cells
2.2. Assay for Antibacterial Activity In Vitro
2.3. Animal Experiments
2.4. 16S rDNA Sequencing and Bioinformatic Analysis
2.5. Adhesion and Invasion Assay
2.6. Determination of Virulence Genes
2.7. Cytokines Measurements
2.8. Statistics
3. Results
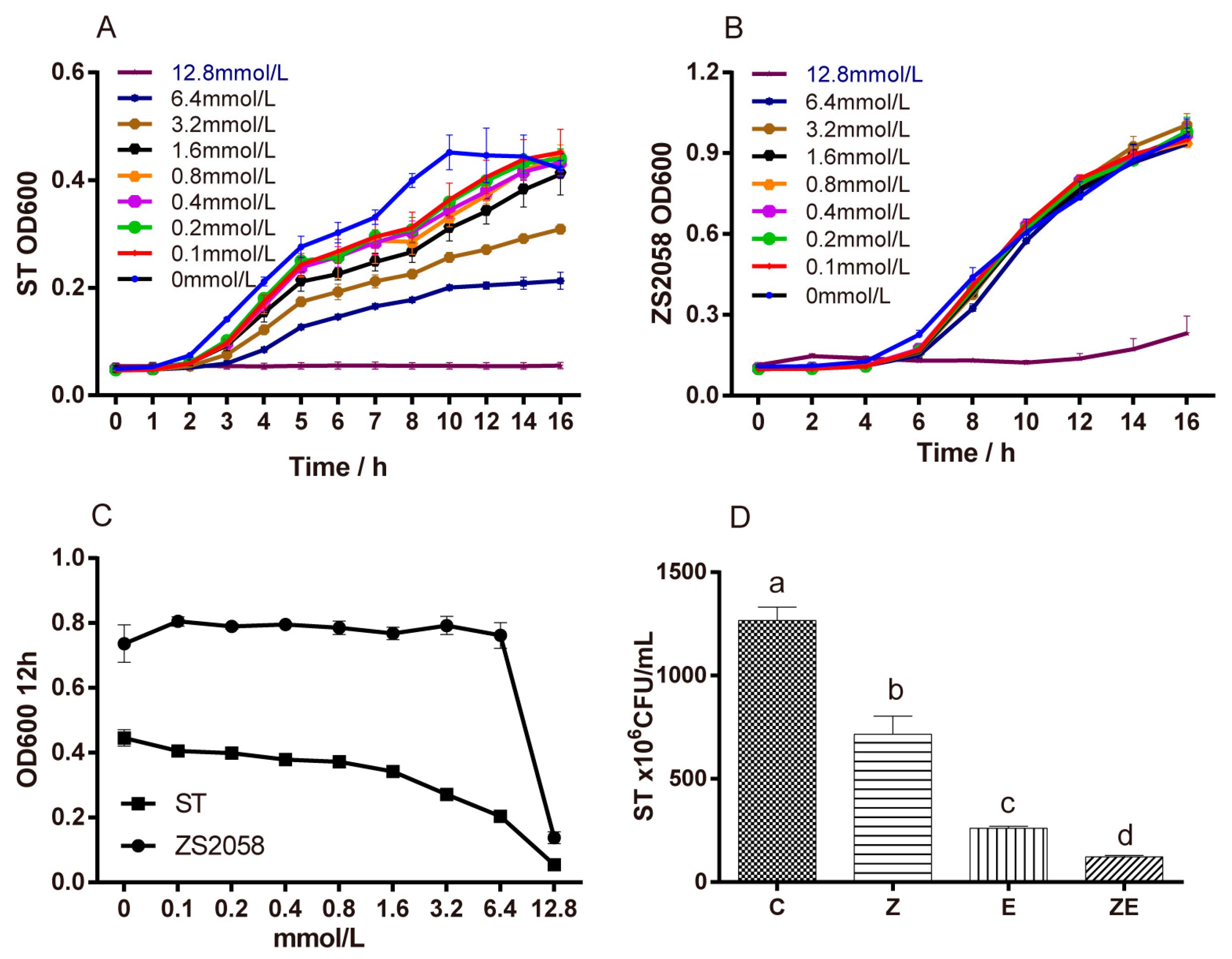
3.1. Combination of l. Plantarum ZS2058 and Eugenol (Clpze) Exerted Synergistic Effect on the Inhibition of ST Growth In Vitro
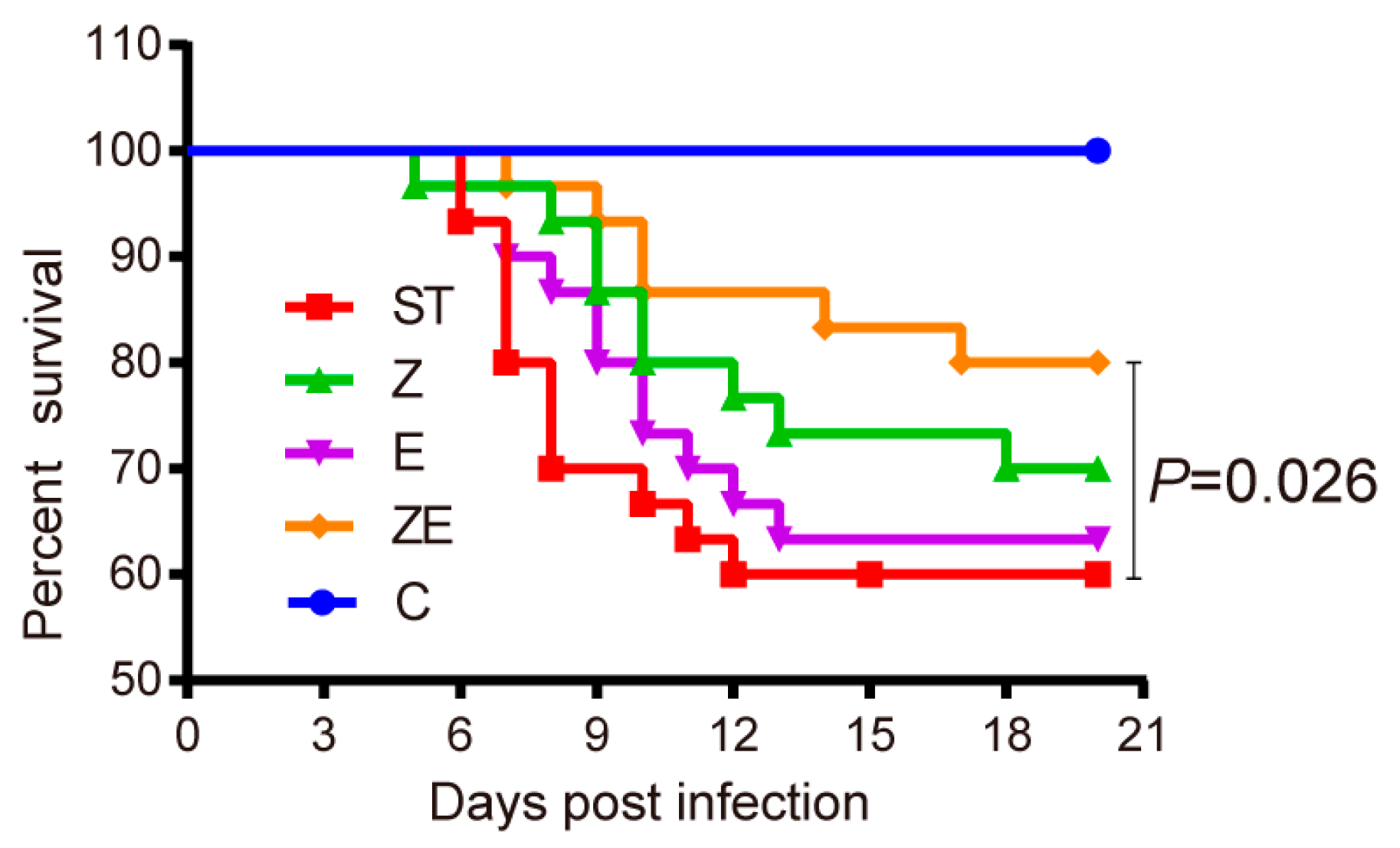
3.2. CLPZE Significantly Increased the Survival Rate of ST-Infected Mice
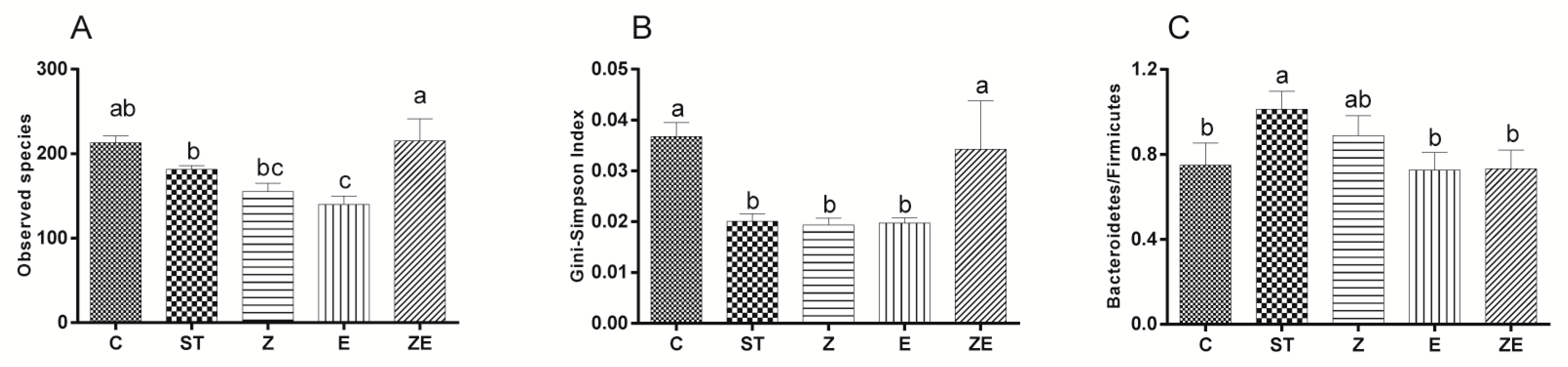
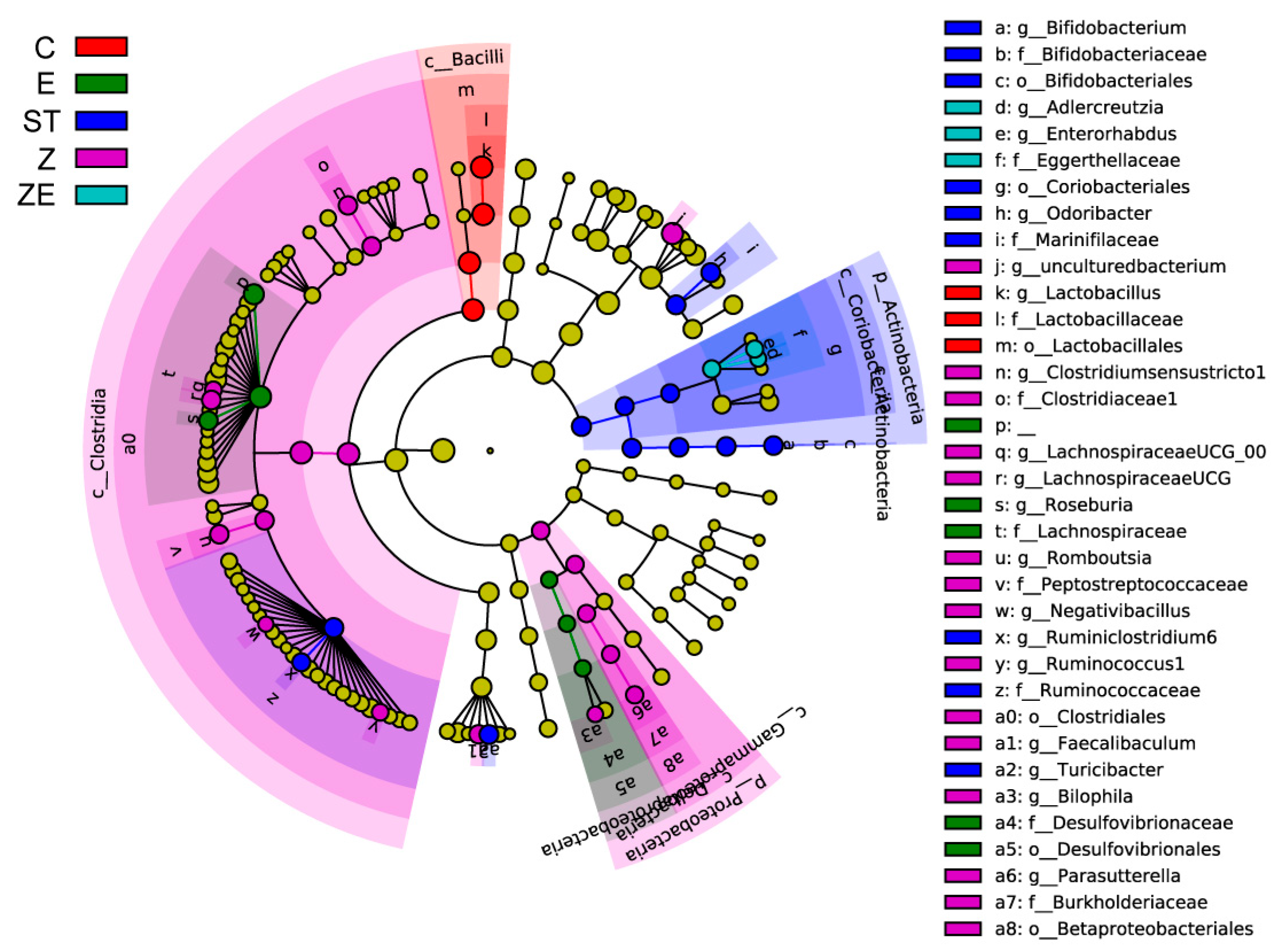
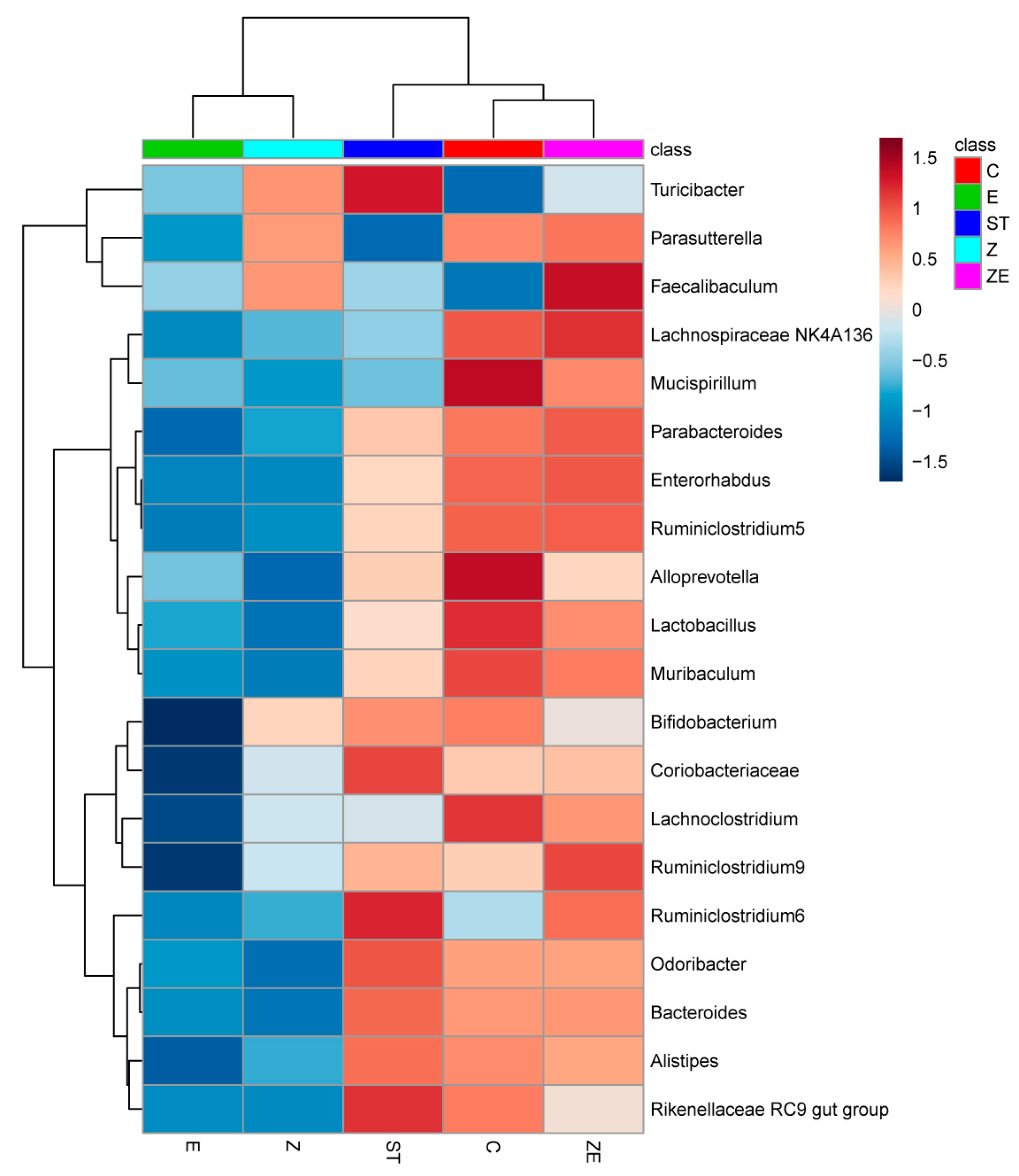
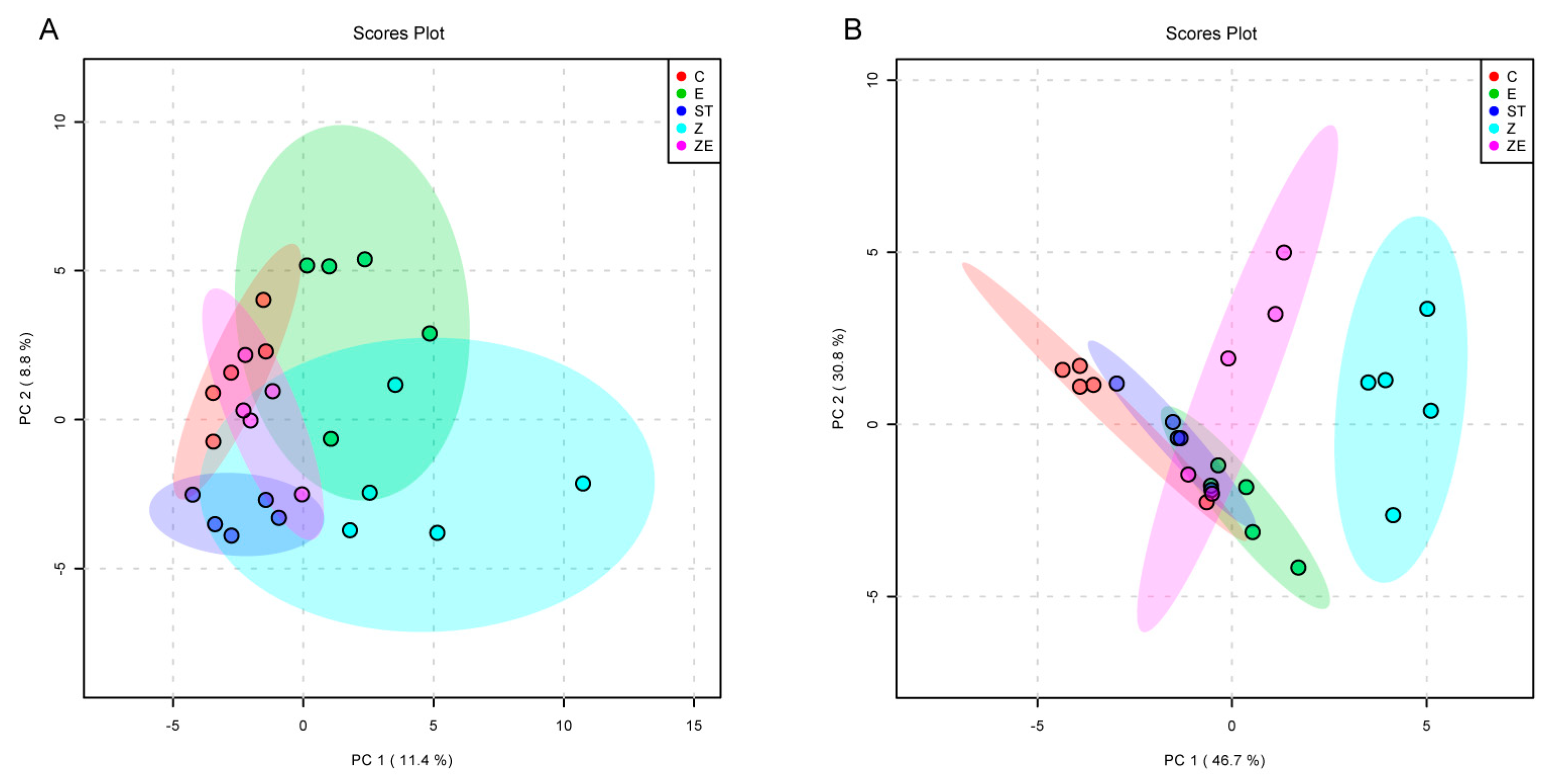
3.3. CLPZE Improves Intestinal Microbiota Structure of ST-Infected Mice
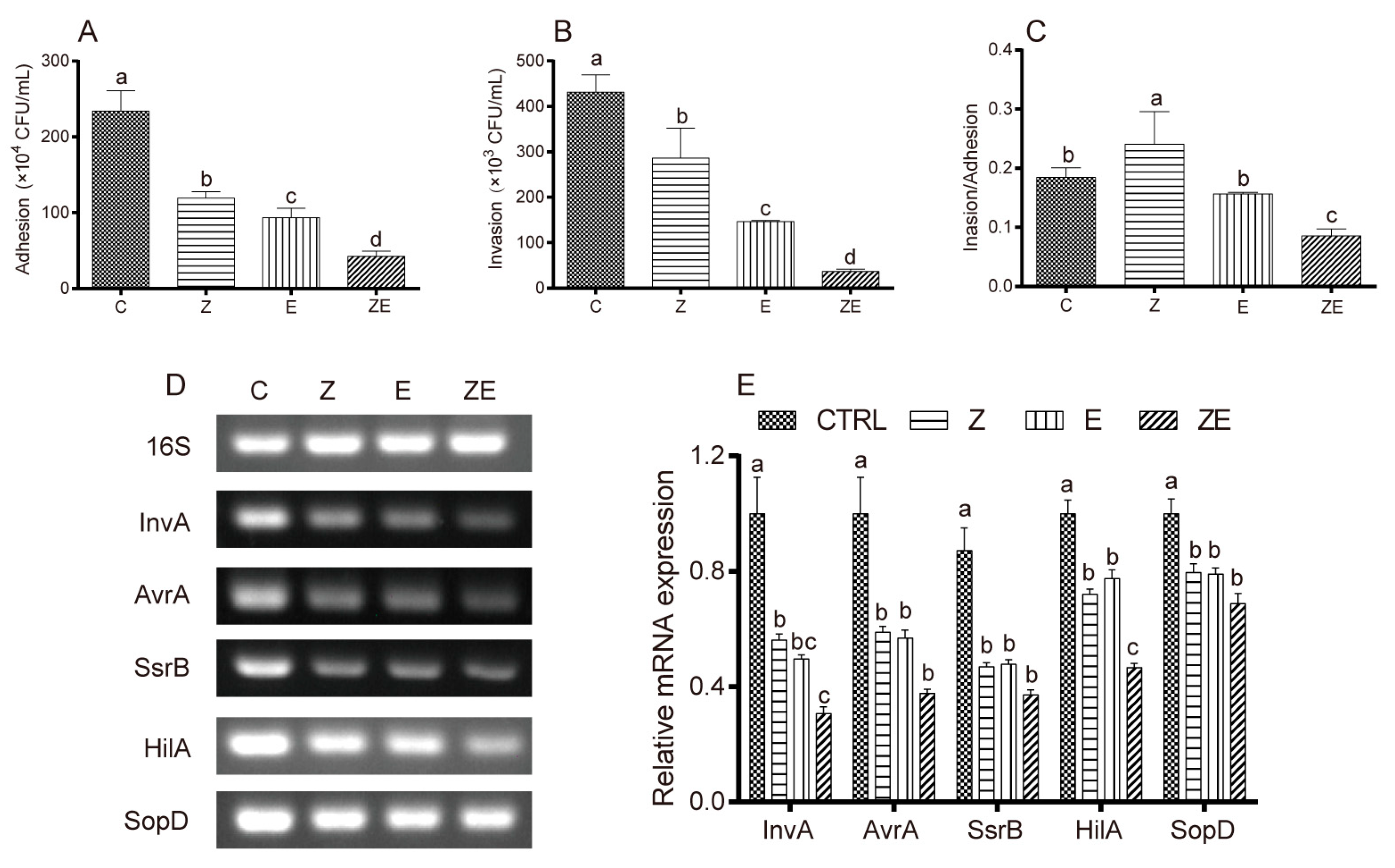
3.4. CLPZE Reduces Salmonella Invasiveness by Inhibiting its Virulence
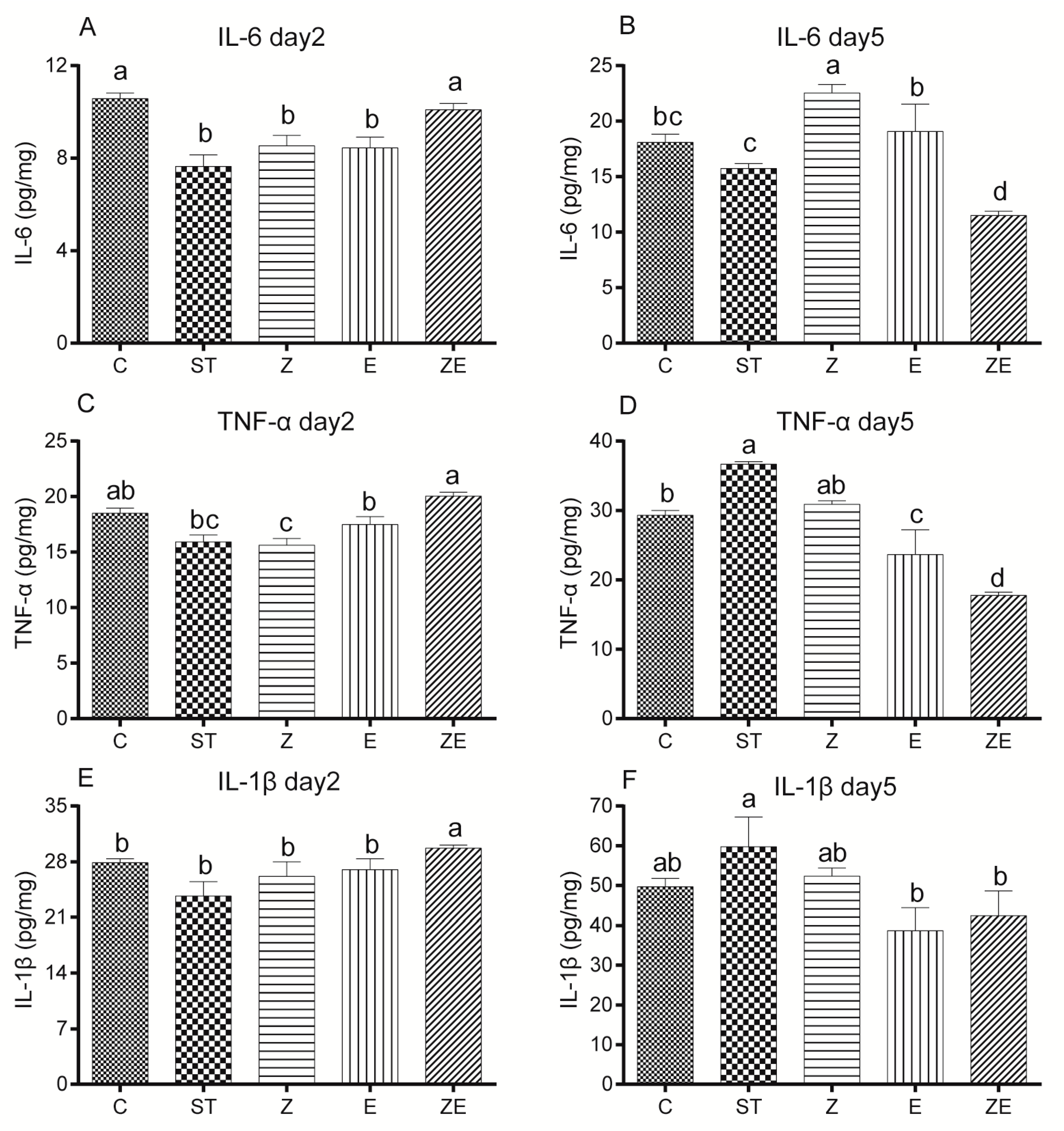
3.5. CLPZE Elicited Earlier Immune Responses to Prevent Systemic Infection by ST
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Antillon, M.; Warren, J.L.; Crawford, F.W.; Weinberger, D.M.; Kurum, E.; Pak, G.D.; Marks, F.; Pitzer, V.E. The burden of typhoid fever in low- and middle-income countries: A meta-regression approach. PLoS Negl. Trop. Dis. 2017, 11, e0005376. [Google Scholar] [CrossRef] [PubMed]
- Mogasale, V.; Maskery, B.; Ochiai, R.L.; Lee, J.S.; Mogasale, V.V.; Ramani, E.; Kim, Y.E.; Park, J.K.; Wierzba, T.F. Burden of typhoid fever in low-income and middle-income countries: A systematic, literature-based update with risk-factor adjustment. Lancet Glob. Health 2014, 2, e570–e580. [Google Scholar] [CrossRef] [Green Version]
- Parry, C.M. Typhoid fever. N. Engl. J. Med. 2002, 347, 1770–1782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunn, J.S.; Marshall, J.M.; Baker, S.; Dongol, S.; Charles, R.C.; Ryan, E.T. Salmonella chronic carriage: Epidemiology, diagnosis, and gallbladder persistence. Trends Microbiol. 2014, 22, 648–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitzer, V.E.; Bowles, C.C.; Baker, S.; Kang, G.; Balaji, V.; Farrar, J.J.; Grenfell, B.T. Predicting the impact of vaccination on the transmission dynamics of typhoid in South Asia: A mathematical modeling study. PLoS Negl. Trop. Dis. 2014, 8, e2642. [Google Scholar] [CrossRef]
- Saul, A.; Smith, T.; Maire, N. Stochastic simulation of endemic Salmonella enterica serovar Typhi: The importance of long lasting immunity and the carrier state. PLoS ONE 2013, 8, e74097. [Google Scholar] [CrossRef] [Green Version]
- Lam, L.H.; Monack, D.M. Intraspecies competition for niches in the distal gut dictate transmission during persistent Salmonella infection. PLoS Pathog. 2014, 10, e1004527. [Google Scholar] [CrossRef]
- Johnson, R.; Mylona, E.; Frankel, G. Typhoidal Salmonella: Distinctive virulence factors and pathogenesis. Cell Microbiol. 2018, 20, e12939. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Gong, L.; Wang, B.; Wu, Y.; Wang, Y.; Mei, X.; Xu, H.; Tang, L.; Liu, R.; Zeng, Z.; et al. Glycyrrhizin Attenuates Salmonella enterica Serovar Typhimurium Infection: New Insights Into Its Protective Mechanism. Front. Immunol. 2018, 9, 2321. [Google Scholar] [CrossRef] [Green Version]
- Daphne, A.C.; Stapels, P.W.S.H.; Westermann, A.J.; Fisher, R.A.; Thurston, T.L.; Saliba, A.; Blommestein, I.; Vogel, J.; Helaine, S. Salmonella persisters undermine host immune defenses during antibiotic treatment. Science 2018, 362, 1156–1160. [Google Scholar]
- Tanner, S.A.; Chassard, C.; Rigozzi, E.; Lacroix, C.; Stevens, M.J. Bifidobacterium thermophilum RBL67 impacts on growth and virulence gene expression of Salmonella enterica subsp. enterica serovar Typhimurium. BMC Microbiol. 2016, 16, 46. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Gu, Z.; Song, F.; Zhang, H.; Zhao, J.; Chen, W. Lactobacillus plantarum ZS2058 and Lactobacillus rhamnosus GG Use Different Mechanisms to Prevent Salmonella Infection in vivo. Front. Microbiol. 2019, 10, 299. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Hu, D.; Chen, Y.; Huang, H.; Zhang, H.; Zhao, J.; Gu, Z.; Chen, W. Strain-specific properties of Lactobacillus plantarum for prevention of Salmonella infection. Food Funct. 2018, 9, 3673–3682. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ding, Y.; Wang, S.; Chen, H.; Zhang, H.; Chen, W.; Gu, Z.; Chen, Y.Q. Extract of Syzygium aromaticum suppress eEF1A protein expression and fungal growth. J. Appl. Microbiol. 2017, 123, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Barboza, J.N.; da Silva Maia Bezerra Filho, C.; Silva, R.O.; Medeiros, J.V.R.; de Sousa, D.P. An Overview on the Anti-inflammatory Potential and Antioxidant Profile of Eugenol. Oxid. Med. Cell Longev. 2018, 2018, 3957262. [Google Scholar] [CrossRef] [PubMed]
- Api, A.M.; Belsito, D.; Bhatia, S.; Bruze, M.; Calow, P.; Dagli, M.L.; Dekant, W.; Fryer, A.D.; Kromidas, L.; La Cava, S.; et al. RIFM fragrance ingredient safety assessment, Eugenol, CAS Registry Number 97-53-0. Food Chem. Toxicol. 2016, 97S, S25–S37. [Google Scholar] [CrossRef]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods--a review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef]
- Vimal, A.; Jha, A.; Kumar, A. Eugenol derivatives prospectively inhibit l-asparaginase: A heady target protein of Salmonella typhimurium. Microb. Pathog. 2018, 114, 8–16. [Google Scholar] [CrossRef]
- Upadhyay, A.; Upadhyaya, I.; Mooyottu, S.; Venkitanarayanan, K. Eugenol in combination with lactic acid bacteria attenuates Listeria monocytogenes virulence in vitro and in invertebrate model Galleria mellonella. J. Med. Microbiol. 2016, 65, 443–455. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, S.; Moran, A.; Martinez-Blanco, H.; Ferrero, M.A.; Rodriguez-Aparicio, L.B. The Usefulness of Non-Toxic Plant Metabolites in the Control of Bacterial Proliferation. Probiotics Antimicrob. Proteins 2017, 9, 323–333. [Google Scholar] [CrossRef]
- Hawrelak, J.A.; Cattley, T.; Myers, S.P. Essential oils in the treatment of intestinal dysbiosis: A preliminary in vitro study. Altern. Med. Rev. A J. Clin. Ther. 2009, 14, 380–384. [Google Scholar]
- Deng, X.; Luo, Z.; Liu, Y.; Zhang, Y.; Feng, H.; Wang, J. Application of eugenol in the preparation of drugs to treat salmonella infection. Chinese patent, 2017; 201610935684.3. [Google Scholar]
- Barthel, M.; Hapfelmeier, S.; Quintanilla-Martinez, L.; Kremer, M.; Rohde, M.; Hogardt, M.; Pfeffer, K.; Russmann, H.; Hardt, W.D. Pretreatment of Mice with Streptomycin Provides a Salmonella enterica Serovar Typhimurium Colitis Model That Allows Analysis of Both Pathogen and Host. Infect. Immun. 2003, 71, 2839–2858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bassanetti, I.; Carcelli, M.; Buschini, A.; Montalbano, S.; Leonardi, G.; Pelagatti, P.; Tosi, G.; Massi, P.; Fiorentini, L.; Rogolino, D. Investigation of antibacterial activity of new classes of essential oils derivatives. Food Control 2017, 73, 606–612. [Google Scholar] [CrossRef]
- Mao, B.; Li, D.; Ai, C.; Zhao, J.; Zhang, H.; Chen, W. Lactulose Differently Modulates the Composition of Luminal and Mucosal Microbiota in C57BL/6J Mice. J. Agric. Food Chem. 2016, 64, 6240–6247. [Google Scholar] [CrossRef] [PubMed]
- Gang, W.; Yu, Z.; Tian, F.; Xing, J.; Chen, H.; Liu, X.; Zhang, Q.; Zhao, J.; Chen, Y.; Hao, Z. Screening of adhesive lactobacilli with antagonistic activity against Campylobacter jejuni. Food Control 2014, 44, 49–57. [Google Scholar]
- Jacobson, A.; Lam, L.; Rajendram, M.; Tamburini, F.; Honeycutt, J.; Pham, T.; Van Treuren, W.; Pruss, K.; Stabler, S.R.; Lugo, K.; et al. A Gut Commensal-Produced Metabolite Mediates Colonization Resistance to Salmonella Infection. Cell Host Microbe 2018, 24, 296–307 e297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, O.; van Berkel, L.A.; Chain, F.; Tanweer Khan, M.; Taverne, N.; Sokol, H.; Duncan, S.H.; Flint, H.J.; Harmsen, H.J.; Langella, P.; et al. Faecalibacterium prausnitzii A2-165 has a high capacity to induce IL-10 in human and murine dendritic cells and modulates T cell responses. Sci. Rep. 2016, 6, 18507. [Google Scholar] [CrossRef] [Green Version]
- Herp, S.; Brugiroux, S.; Garzetti, D.; Ring, D.; Jochum, L.M.; Beutler, M.; Eberl, C.; Hussain, S.; Walter, S.; Gerlach, R.G.; et al. Mucispirillum schaedleri Antagonizes Salmonella Virulence to Protect Mice against Colitis. Cell Host Microbe 2019, 25, 681–694.e688. [Google Scholar] [CrossRef]
- Wu, M.; Li, J.; An, Y.; Li, P.; Xiong, W.; Li, J.; Yan, D.; Wang, M.; Zhong, G. Chitooligosaccharides Prevents the Development of Colitis-Associated Colorectal Cancer by Modulating the Intestinal Microbiota and Mycobiota. Front Microbiol. 2019, 10, 2101. [Google Scholar] [CrossRef]
- Lievin-Le Moal, V.; Servin, A.L. Anti-infective activities of lactobacillus strains in the human intestinal microbiota: From probiotics to gastrointestinal anti-infectious biotherapeutic agents. Clin. Microbiol. Rev. 2014, 27, 167–199. [Google Scholar] [CrossRef] [Green Version]
- Andreevskaya, M.; Jaaskelainen, E.; Johansson, P.; Ylinen, A.; Paulin, L.; Bjorkroth, J.; Auvinen, P. Food Spoilage-Associated Leuconostoc, Lactococcus, and Lactobacillus Species Display Different Survival Strategies in Response to Competition. Appl. Environ. Microbiol. 2018, 84, e00554-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stecher, B.; Chaffron, S.; Kappeli, R.; Hapfelmeier, S.; Freedrich, S.; Weber, T.C.; Kirundi, J.; Suar, M.; McCoy, K.D.; von Mering, C.; et al. Like will to like: Abundances of closely related species can predict susceptibility to intestinal colonization by pathogenic and commensal bacteria. PLoS Pathog. 2010, 6, e1000711. [Google Scholar] [CrossRef] [PubMed]
- Foley, S.L.; Johnson, T.J.; Ricke, S.C.; Nayak, R.; Danzeisen, J. Salmonella pathogenicity and host adaptation in chicken-associated serovars. Microbiol. Mol. Biol. Rev. 2013, 77, 582–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LaRock, D.L.; Chaudhary, A.; Miller, S.I. Salmonellae interactions with host processes. Nat. Rev. Microbiol. 2015, 13, 191–205. [Google Scholar] [CrossRef]
- Laughlin, R.C.; Knodler, L.A.; Barhoumi, R.; Payne, H.R.; Wu, J.; Gomez, G.; Pugh, R.; Lawhon, S.D.; Baumler, A.J.; Steele-Mortimer, O.; et al. Spatial segregation of virulence gene expression during acute enteric infection with Salmonella enterica serovar Typhimurium. mBio 2014, 5, e00946-13. [Google Scholar] [CrossRef] [Green Version]
- Que, F.; Wu, S.; Huang, R. Salmonella pathogenicity island 1(SPI-1) at work. Curr. Microbiol. 2013, 66, 582–587. [Google Scholar] [CrossRef]
- Palmer, A.D.; Kim, K.; Slauch, J.M. PhoP-Mediated Repression of the SPI1 Type 3 Secretion System in Salmonella enterica Serovar Typhimurium. J. Bacteriol. 2019, 201, e00264-19. [Google Scholar] [CrossRef] [Green Version]
- Everest, P.; Ketley, J.; Hardy, S.; Douce, G.; Khan, S.; Shea, J.; Holden, D.; Maskell, D.; Dougan, G. Evaluation of Salmonella typhimurium Mutants in a Model of Experimental Gastroenteritis. Infect. Immun. 1999, 67, 2815–2821. [Google Scholar] [CrossRef] [Green Version]
- Keszei, A.F.; Tang, X.; McCormick, C.; Zeqiraj, E.; Rohde, J.R.; Tyers, M.; Sicheri, F. Structure of an SspH1-PKN1 complex reveals the basis for host substrate recognition and mechanism of activation for a bacterial E3 ubiquitin ligase. Mol. Cell. Biol. 2014, 34, 362–373. [Google Scholar] [CrossRef] [Green Version]
- Lu, R.; Wu, S.; Liu, X.; Xia, Y.; Zhang, Y.G.; Sun, J. Chronic effects of a Salmonella type III secretion effector protein AvrA in vivo. PLoS ONE 2010, 5, e10505. [Google Scholar] [CrossRef]
- Perez-Morales, D.; Banda, M.M.; Chau, N.Y.E.; Salgado, H.; Martinez-Flores, I.; Ibarra, J.A.; Ilyas, B.; Coombes, B.K.; Bustamante, V.H. The transcriptional regulator SsrB is involved in a molecular switch controlling virulence lifestyles of Salmonella. PLoS Pathog. 2017, 13, e1006497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steele-Mortimer, O. The Salmonella-containing vacuole: Moving with the times. Curr. Opin. Microbiol. 2008, 11, 38–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, M.W.; Jones, M.A.; Watson, P.R.; Siber, A.M.; McCormick, B.A.; Hedges, S.; Rosqvist, R.; Wallis, T.S.; Galyov, E.E. The secreted effector protein of Salmonella dublin, SopA, is translocated into eukaryotic cells and influences the induction of enteritis. Cell Microbiol. 2000, 2, 293–303. [Google Scholar] [CrossRef] [PubMed]
- Jennings, E.; Thurston, T.L.M.; Holden, D.W. Salmonella SPI-2 Type III Secretion System Effectors: Molecular Mechanisms And Physiological Consequences. Cell Host Microbe 2017, 22, 217–231. [Google Scholar] [CrossRef] [PubMed]
- Hitchcock, J.R.; Cook, C.N.; Bobat, S.; Ross, E.A.; Flores-Langarica, A.; Lowe, K.L.; Khan, M.; Dominguez-Medina, C.C.; Lax, S.; Carvalho-Gaspar, M.; et al. Inflammation drives thrombosis after Salmonella infection via CLEC-2 on platelets. J. Clin. Investig. 2015, 125, 4429–4446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islamuddin, M.; Chouhan, G.; Want, M.Y.; Ozbak, H.A.; Hemeg, H.A.; Afrin, F. Immunotherapeutic Potential of Eugenol Emulsion in Experimental Visceral Leishmaniasis. PLoS Negl. Trop. Dis. 2016, 10, e0005011. [Google Scholar] [CrossRef]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, F.; Liu, J.; Zhao, W.; Huang, H.; Hu, D.; Chen, H.; Zhang, H.; Chen, W.; Gu, Z. Synergistic Effect of Eugenol and Probiotic Lactobacillus Plantarum Zs2058 against Salmonella Infection in C57bl/6 Mice. Nutrients 2020, 12, 1611. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12061611
Song F, Liu J, Zhao W, Huang H, Hu D, Chen H, Zhang H, Chen W, Gu Z. Synergistic Effect of Eugenol and Probiotic Lactobacillus Plantarum Zs2058 against Salmonella Infection in C57bl/6 Mice. Nutrients. 2020; 12(6):1611. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12061611
Chicago/Turabian StyleSong, Fanfen, Junsheng Liu, Wenyu Zhao, Hongxuan Huang, Diangeng Hu, Haiqin Chen, Hao Zhang, Wei Chen, and Zhennan Gu. 2020. "Synergistic Effect of Eugenol and Probiotic Lactobacillus Plantarum Zs2058 against Salmonella Infection in C57bl/6 Mice" Nutrients 12, no. 6: 1611. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12061611