Effects of Pre-Sleep Whey vs. Plant-Based Protein Consumption on Muscle Recovery Following Damaging Morning Exercise

Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
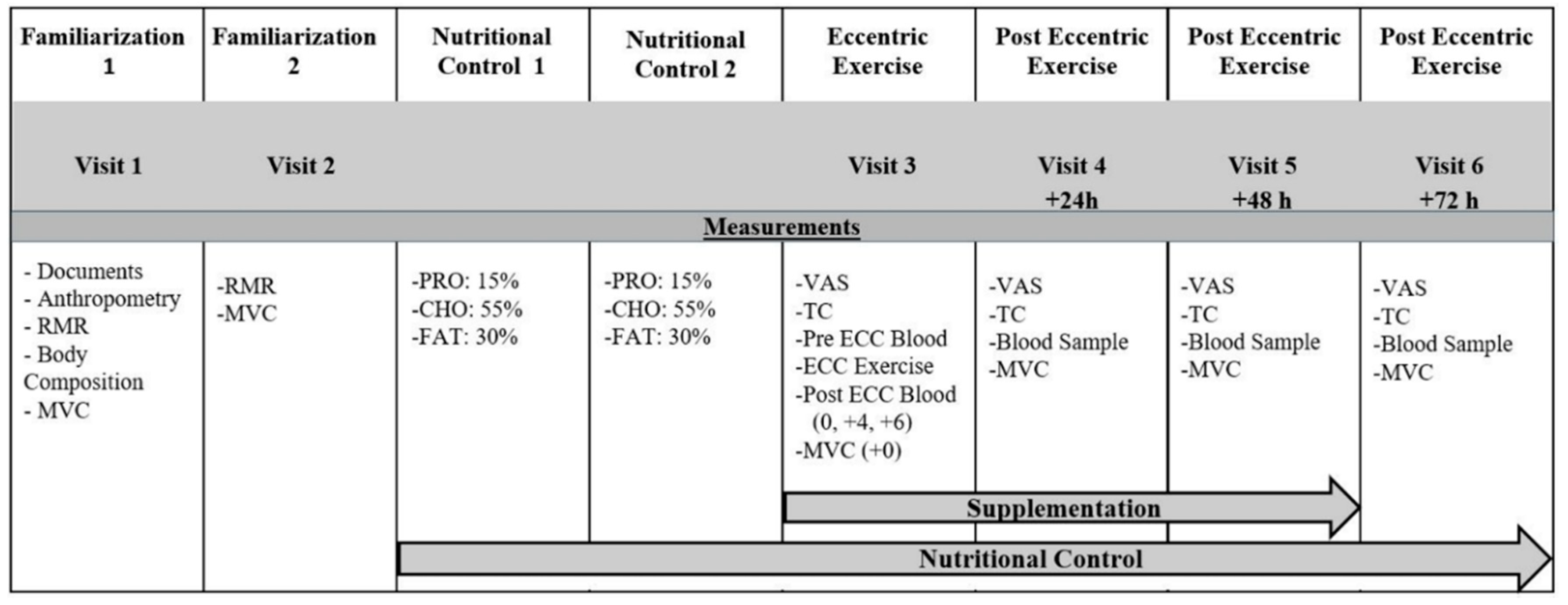
2.2. Experimental Protocol
2.3. Resting Metabolic Rate
2.4. Maximal Voluntary Contraction
2.5. Muscle Soreness
2.6. Thigh Circumference
2.7. Blood Sampling
2.8. Muscle Damage Protocol
2.9. Dietary and Exercise Control
2.10. Statistics
3. Results
3.1. Participant Characteristics
3.2. Nutrient Intake
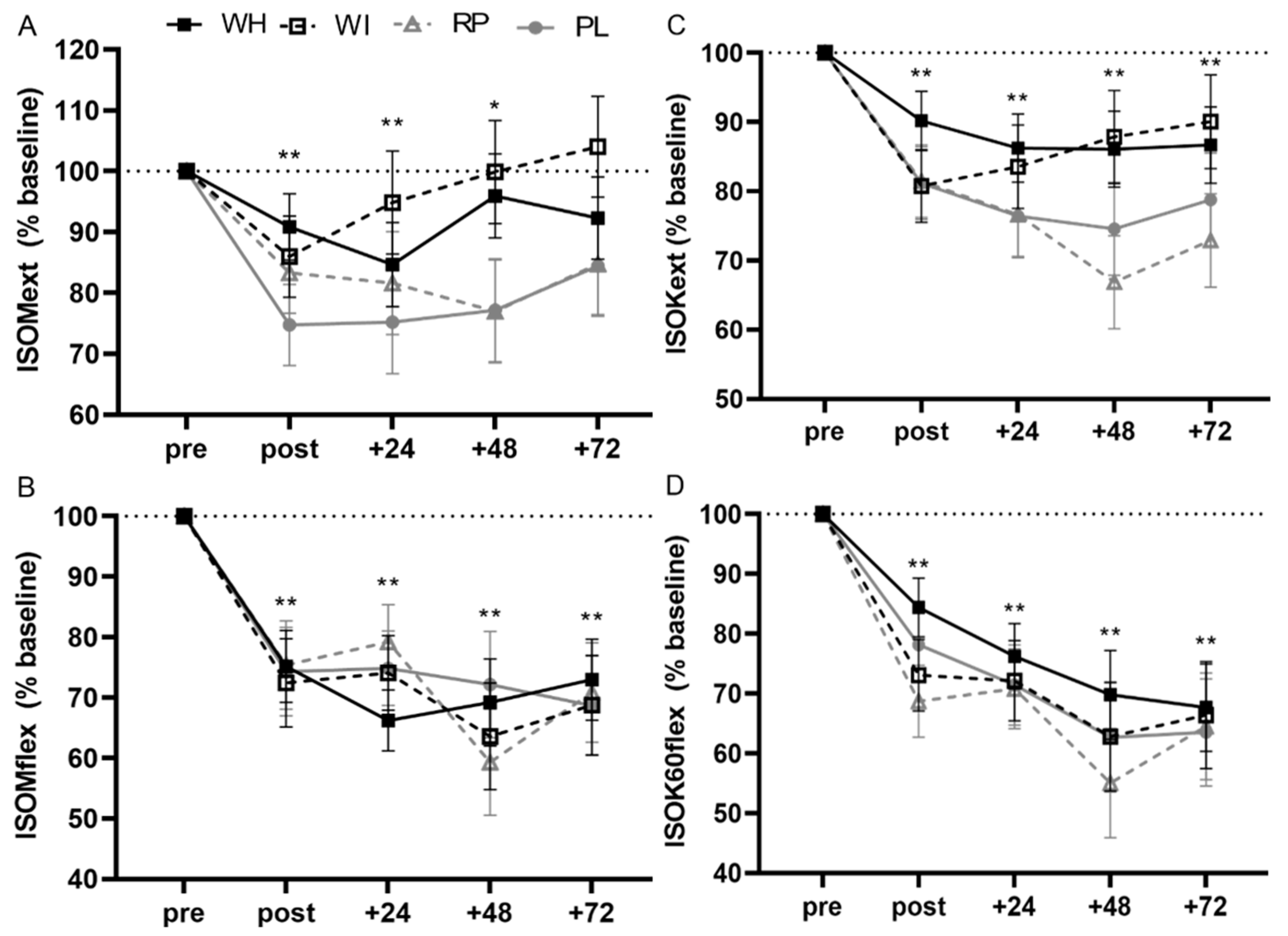
3.3. Muscle Function
3.3.1. ISOMext
3.3.2. ISOMflex
3.3.3. ISOK60ext
3.3.4. ISOK60flex
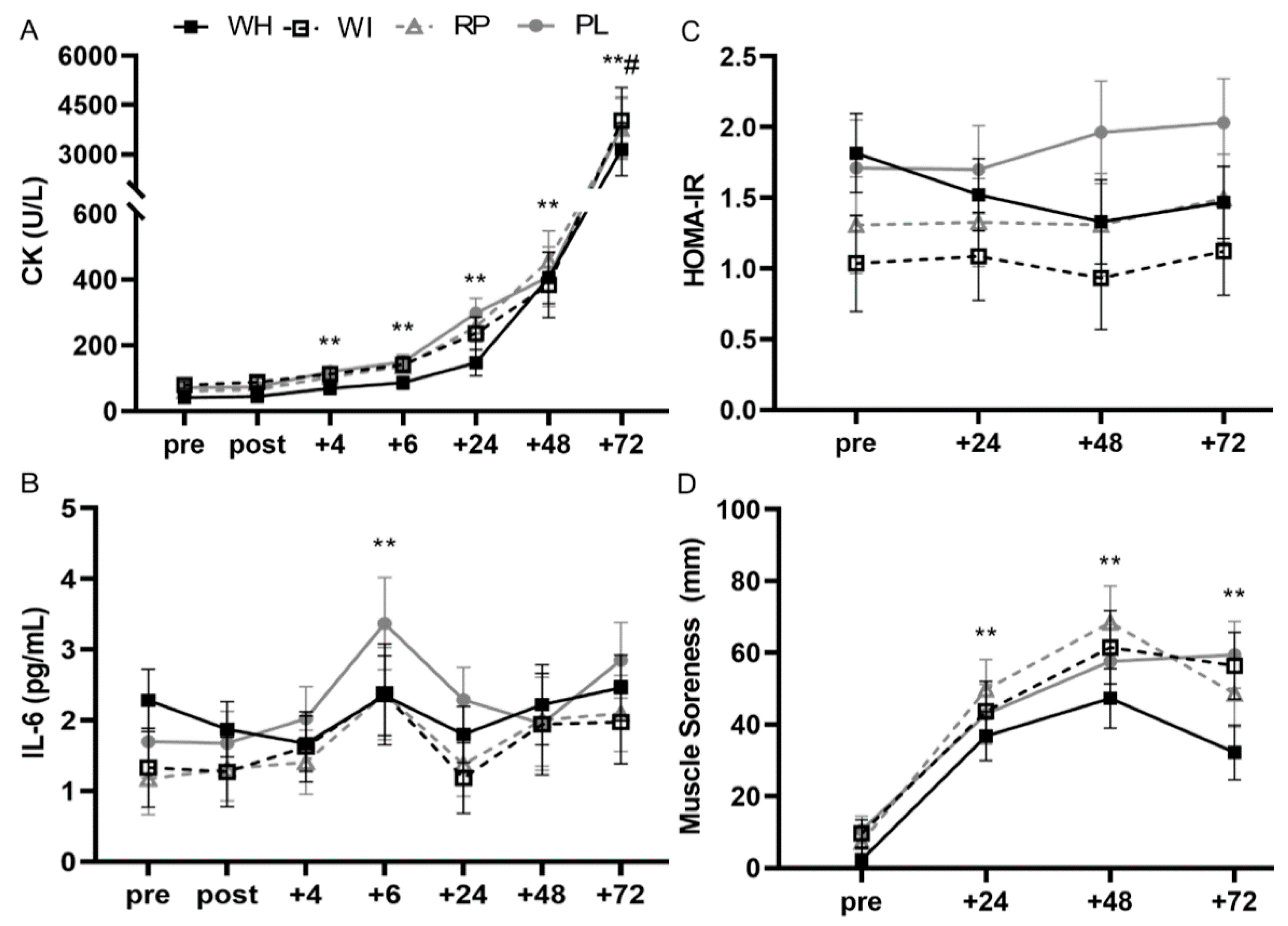
3.4. Blood Markers
3.4.1. Creatine Kinase (CK)
3.4.2. IL-6
3.4.3. IL-10
3.4.4. HOMA-IR
3.5. Muscle Soreness
3.6. Thigh Circumference
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Foster, C.; Wright, G.; Battista, R.A.; Porcari, J.P. Training in the aging athlete. Curr. Sports Med. Rep. 2007, 6, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, J.F.T.; Lamb, K.L.; Twist, C. Exercise-induced muscle damage and recovery in young and middle-aged males with different resistance training experience. Sports 2019, 7, 132. [Google Scholar] [CrossRef] [Green Version]
- Tee, J.C.; Bosch, A.N.; Lambert, M.I. Metabolic consequences of exercise-induced muscle damage. Sport. Med. 2007, 37, 827–836. [Google Scholar] [CrossRef]
- MacIntyre, D.L.; Reid, W.D.; McKenzie, D.C. Delayed muscle soreness. The inflammatory response to muscle injury and its clinical implications. Sport. Med. 1995, 20, 24–40. [Google Scholar] [CrossRef] [PubMed]
- Manfredi, T.G.; Fielding, R.A.; O’Reilly, K.P.; Meredith, C.N.; Lee, H.Y.; Evans, W.J. Plasma creatine kinase activity and exercise-induced muscle damage in older men. Med. Sci. Sport. Exerc. 1991, 23, 1028–1034. [Google Scholar] [CrossRef]
- Roth, S.M.; Martel, G.F.; Ivey, F.M.; Lemmer, J.T.; Metter, E.J.; Hurley, B.F.; Rogers, M.A. High-volume, heavy-resistance strength training and muscle damage in young and older women. J. Appl. Physiol. 2000, 88, 1112–1118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breen, L.; Phillips, S.M. Interactions between exercise and nutrition to prevent muscle waste during ageing. Br. J. Clin. Pharmacol. 2012, 75, 708–715. [Google Scholar] [CrossRef] [Green Version]
- Davies, R.W.; Carson, B.P.; Jakeman, P.M. The effect of whey protein supplementation on the temporal recovery of muscle function following resistance training: A systematic review and meta-analysis. Nutrients 2018, 10, 221. [Google Scholar] [CrossRef] [Green Version]
- Groen, B.B.L.; Res, P.T.; Pennings, B.; Hertle, E.; Senden, J.M.G.; Saris, W.H.M.; Van Loon, L.J.C. Intragastric protein administration stimulates overnight muscle protein synthesis in elderly men. AJP Endocrinol. Metab. 2012, 302, E52–E60. [Google Scholar] [CrossRef]
- Res, P.T.; Groen, B.; Pennings, B.; Beelen, M.; Wallis, G.A.; Gijsen, A.P.; Senden, J.M.G.; Van Loon, L.J.C. Protein ingestion before sleep improves postexercise overnight recovery. Med. Sci. Sports Exerc. 2012, 44, 1560–1569. [Google Scholar] [CrossRef] [Green Version]
- Snijders, T.; Res, P.T.; Smeets, J.S.J.; Vliet, S.V.; Kranenburg, J.V.; Maase, K.; Kies, A.K.; Verdijk, L.B.; Van Loon, L.J.C. Protein ingestion before sleep increases muscle mass and strength gains during prolonged resistance-type exercise training in healthy young men. J. Nutr. 2015, 145, 1178–1184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West, D.W.D.; Sawan, S.A.; Mazzulla, M.; Williamson, E.; Moore, D.R. Whey protein supplementation enhances whole body protein metabolism and performance recovery after resistance exercise: A double-blind crossover study. Nutrients 2017, 9, 735. [Google Scholar] [CrossRef] [PubMed]
- Abbott, W.; Brett, A.; Cockburn, E.; Clifford, T. Presleep casein protein ingestion acceleration of functional recovery in professional soccer players. Int. J. Sports Physiol. Perform. 2019, 14, 385–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trommelen, J.; Van Loon, L. Pre-Sleep protein ingestion to improve the skeletal muscle adaptive response to exercise training. Nutrients 2016, 8, 763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trommelen, J.; Holwerda, A.M.; Kouw, I.W.K.; Langer, H.; Halson, S.L.; Rollo, I.; Verdijk, L.B.; Van Loon, L.J.C. Resistance exercise augments postprandial overnight muscle protein synthesis rates. Med. Sci. Sports Exerc. 2016, 48, 2517–2525. [Google Scholar] [CrossRef] [Green Version]
- Kinsey, A.W.; Cappadona, S.R.; Panton, L.B.; Allman, B.R.; Contreras, R.J.; Hickner, R.C.; Ormsbee, M.J. The effect of casein protein prior to sleep on fat metabolism in obese men. Nutrients 2016, 8, 452. [Google Scholar] [CrossRef]
- Allman, B.R.; Morrissey, M.C.; Kim, J.S.; Panton, L.B.; Contreras, R.J.; Hickner, R.C.; Ormsbee, M.J. Lipolysis and fat oxidation are not altered with presleep compared with daytime caseinprotein intake in resistance-trained women. J. Nutr. 2019, 47–54. [Google Scholar] [CrossRef]
- Holwerda, A.M.; Kouw, I.W.; Trommelen, J.; Halson, S.L.; Wodzig, W.K.; Verdijk, L.B.; Van Loon, L.J. Physical activity performed in the evening increases the overnight muscle protein synthetic response to presleep protein ingestion in older men. J. Nutr. 2016, 146, 1307–1314. [Google Scholar] [CrossRef] [Green Version]
- Trommelen, J.; Kouw, I.W.K.; Holwerda, A.M.; Snijders, T.; Halson, S.L.; Rollo, I.; Verdijk, L.B.; Van Loon, L.J.C. Pre-sleep dietary protein-derived amino acids are incorporated in myofibrillar protein during post-exercise overnight recovery. Am. J. Physiol. Endocrinol. Metab. 2018. [Google Scholar] [CrossRef]
- Karagounis, L.G.; Beaumont, M.; Donato-Capel, L.; Godin, J.P.; Kapp, A.F.; Draganidis, D.; Pinaud, S.; Vuichoud, J.; Shevlyakova, M.; Rade-Kukic, K.; et al. Ingestion of a pre-bedtime protein containing beverage prevents overnight induced negative whole body protein balance in healthy middle-aged men: A randomized trial. Front. Nutr. 2019, 6. [Google Scholar] [CrossRef]
- Burd, N.A.; Yang, Y.; Moore, D.R.; Tang, J.E.; Tarnopolsky, M.A.; Phillips, S.M. Greater stimulation of myofibrillar protein synthesis with ingestion of whey protein isolate v. micellar casein at rest and after resistance exercise in elderly men. Br. J. Nutr. 2012, 108, 958–962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, S.B.; Tarnopolsky, M.A.; MacDonald, M.J.; MacDonald, J.R.; Armstrong, D.; Phillips, S.M. Consumption of fluid skim milk promotes greater muscle protein accretion after resistance exercise than does consumption of an isonitrogenous and isoenergetic soy-protein beverage. Am. J. Clin. Nutr. 2007, 85, 1031–1040. [Google Scholar] [CrossRef] [PubMed]
- Hartman, J.W.; Tang, J.E.; Wilkinson, S.B.; Tarnopolsky, M.A.; Lawrence, R.L.; Fullerton, A.V.; Phillips, S.M. Consumption of fat-free fluid milk after resistance exercise promotes greater lean mass accretion than does consumption of soy or carbohydrate in young, novice, male weightlifters. Am. J. Clin. Nutr. 2007, 86, 373–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, S.M.; Tang, J.E.; Moore, D.R. The role of milk- and soy-based protein in support of muscle protein synthesis and muscle protein accretion in young and elderly persons. J. Am. Coll. Nutr. 2009, 28, 343–354. [Google Scholar] [CrossRef] [PubMed]
- Van Vliet, S.; Burd, N.A.; Van Loon, L.J. The skeletal muscle anabolic response to plant- versus animal-based protein consumption. J. Nutr. 2015, 145, 1981–1991. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.M.; Hartman, J.W.; Wilkinson, S.B. Dietary protein to support anabolism with resistance exercise in young men. J. Am. Coll. Nutr. 2005, 24, 134–139. [Google Scholar] [CrossRef]
- Gorissen, S.H.M.; Witard, O.C. Characterising the muscle anabolic potential of dairy, meat and plant-based protein sources in older adults. Proc. Nutr. Soc. 2018, 77, 20–31. [Google Scholar] [CrossRef]
- Gorissen, S.H.M.; Crombag, J.J.R.; Senden, J.M.G.; Waterval, W.A.H.; Bierau, J.; Verdijk, L.B.; Van Loon, L.J.C. Protein content and amino acid composition of commercially available plant - based protein isolates. Amino Acids 2018, 50, 1685–1695. [Google Scholar] [CrossRef] [Green Version]
- Snijders, T.; Trommelen, J.; Kouw, I.W.K.; Holwerda, A.M.; Verdijk, L.B.; Van Loon, L.J.C. The impact of pre-sleep protein ingestion on the skeletal muscle adaptive response to exercise in humans: An update. Front. Nutr. 2019, 6, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Cleak, M.J.; Eston, R.G. Muscle soreness, swelling, stiffness and strength loss after intense eccentric exercise. Br. J. Sports Med. 1992, 26, 267–272. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.J.; Child, R.B.; Day, S.H.; Donnelly, A.E. Exercise-induced skeletal muscle damage and adaptation following repeated bouts of eccentric muscle contractions. J. Sports Sci. 1997, 15, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Stupka, N.; Tarnopolsky, M.A.; Yardley, N.J.; Phillips, S.M. Cellular adaptation to repeated eccentric exercise-induced muscle damage. J. Appl. Physiol. 2001, 91, 1669–1678. [Google Scholar] [CrossRef] [PubMed]
- Semmler, J.G.; Tucker, K.J.; Allen, T.J.; Proske, U. Eccentric exercise increases EMG amplitude and force fluctuations during submaximal contractions of elbow flexor muscles. J. Appl. Physiol. 2007, 103, 979–989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apweiler, E.; Wallace, D.; Stansfield, S.; Allerton, D.; Brown, M.; Stevenson, E.; Clifford, T. Pre-bed casein protein supplementation does not enhance acute functional recovery in physically active males and females when exercise is performed in the morning. Sports 2019, 7, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koopman, R.; Van Loon, L.J.C. Aging, exercise, and muscle protein metabolism. J. Appl. Physiol. 2009, 106, 2040–2048. [Google Scholar] [CrossRef]
- Phillips, S.M.; Chevalier, S.; Leidy, H.J. Protein “requirements” beyond the RDA: Implications for optimizing health. Appl. Physiol. Nutr. Metab. 2016, 41, 565–572. [Google Scholar] [CrossRef] [Green Version]
- Holwerda, A.M.; Overkamp, M.; Paulussen, K.J.M.; Smeets, J.S.J.; Van Kranenburg, J.; Backx, E.M.P.; Gijsen, A.P.; Goessens, J.P.B.; Verdijk, L.B.; Van Loon, L.J.C. Protein supplementation after exercise and before sleep does not further augment muscle mass and strength gains during resistance exercise training in active older men. J. Nutr. 2018, 148, 1723–1732. [Google Scholar] [CrossRef]
- Kouw, I.W.; Holwerda, A.M.; Trommelen, J.; Kramer, I.F.; Bastiaanse, J.; Halson, S.L.; Wodzig, W.K.; Verdijk, L.B.; Van Loon, L.J. Protein ingestion before sleep increases overnight muscle protein synthesis rates in healthy older men: A randomized controlled trial. J. Nutr. 2017, 147, 2252–2261. [Google Scholar] [CrossRef] [Green Version]
- Joy, J.M.; Vogel, R.M.; Shane Broughton, K.; Kudla, U.; Kerr, N.Y.; Davison, J.M.; Wildman, R.E.C.; DiMarco, N.M. Daytime and nighttime casein supplements similarly increase muscle size and strength in response to resistance training earlier in the day: A preliminary investigation. J. Int. Soc. Sports Nutr. 2018, 15, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Schoenfeld, B.J.; Aragon, A.A.; Krieger, J.W. The effect of protein timing on muscle strength and hypertrophy: A meta-analysis. J. Int. Soc. Sports Nutr. 2013, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Berryman, C.E.; Lieberman, H.R.; Fulgoni, V.L.; Pasiakos, S.M. Protein intake trends and conformity with the Dietary Reference Intakes in the United States: Analysis of the National Health and Nutrition Examination Survey, 2001–2014. Am. J. Clin. Nutr. 2018, 108, 405–413. [Google Scholar] [CrossRef]
- Campbell, B.; Kreider, R.; Ziegenfuss, T.; La Bounty, P.; Roberts, M.; Burke, D.; Landis, J.; Lopez, H.; Antonio, J. International society of sports nutrition position stand: Protein and exercise. J. Int. Soc. Sports Nutr. 2007, 4, 8. [Google Scholar] [CrossRef] [Green Version]
- Fulgoni, V.L. Current protein intake in America: Analysis of the National Health and Nutrition Examination Survey, 2003–2004. Am. J. Clin. Nutr. 2008, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsen, M.S.; Clausen, D.; Jørgensen, A.A.; Mikkelsen, U.R.; Hansen, M. Presleep protein supplementation does not improve recovery during consecutive days of intense endurance training: A randomized controlled trial. Int. J. Sport Nutr. Exerc. Metab. 2019, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ormsbee, M.J.; Kinsey, A.W.; Eddy, W.R.; Madzima, T.A.; Arciero, P.J.; Figueroa, A.; Panton, L.B. The influence of nighttime feeding of carbohydrate or protein combined with exercise training on appetite and cardiometabolic risk in young obese women. Appl. Physiol. Nutr. Metab. 2015, 9, 1–9. [Google Scholar] [CrossRef]
- Kinsey, A.W.; Eddy, W.R.; Madzima, T.A.; Panton, L.B.; Arciero, P.J.; Kim, J.S.; Ormsbee, M.J. Influence of night-time protein and carbohydrate intake on appetite and cardiometabolic risk in sedentary overweight and obese women. Br. J. Nutr. 2014, 112, 320–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theodorou, A.; Nikolaidis, M.; Paschalis, V.; Panayiotou, G.; Fatouris, I.; Koutedakis, Y.; Jamurtas, A. No effect of antioxidant supplementation on muscle performance and blood redox status adaptations to eccentric training. Am. J. Clin. Nutr. 2011, 93, 1373–1383. [Google Scholar] [CrossRef] [Green Version]
- Vanholder, R.; Sukru Sever, M.; Erek, E.; Lameire, N. Rhabdomyolysis. J. Am. Soc. Nephrol. 2000, 11, 1553–1561. [Google Scholar]
- Huerta-Alardín, A.L.; Varon, J.; Marik, P.E. Bench-to-bedside review: Rhabdomyolysis—An overview for clinicians. Crit. Care 2005, 9, 158–169. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Whey Hydrolysate | Whey Isolate | Rice/Pea | Placebo | Total | p-Value | |
|---|---|---|---|---|---|---|
| N | 9 | 6 | 6 | 6 | 27 | |
| Age, yr | 57 ± 2 | 53 ± 3 | 56 ± 2 | 52 ± 4 | 55 ± 1 | 0.655 |
| Height, cm | 179 ± 3 | 180 ± 3 | 179 ± 2 | 180 ± 3 | 179 ± 1 | 0.965 |
| Weight, kg | 86.1 ± 4.3 | 83.9 ± 4.4 | 79.1 ± 4.9 | 78.7 ± 5.5 | 82.4 ± 2.3 | 0.613 |
| Lean Mass, kg | 61.7 ± 1.9 | 60.7 ± 2.5 | 57.2 ± 3.2 | 55.9 ± 3.2 | 59.2 ± 1.3 | 0.346 |
| Body Fat, % | 24 ± 2 | 23 ± 2 | 23 ± 1 | 24 ± 2 | 23 ± 1 | 0.967 |
| Whey Hydrolysate | Whey Isolate | Rice/Pea | Placebo | Total | |
|---|---|---|---|---|---|
| Calories, kcal | 2405 ± 113 | 2439 ± 136 | 2293 ± 102 | 2240 ± 135 | 2351 ± 60 |
| CHO, g | 331 ± 16 | 335 ± 19 | 315 ± 14 | 308 ± 19 | 323 ± 8 |
| CHO, g/kg | 3.86 ± 0.14 | 4.02 ± 0.21 | 4.03 ± 0.22 | 3.96 ± 0.21 | 3.96 ± 0.09 |
| Fat, g | 80 ± 4 | 81 ± 5 | 76 ± 3 | 75 ± 5 | 78 ± 2 |
| Fat, g/kg | 0.94 ± 0.03 | 0.97 ± 0.05 | 0.98 ± 0.05 | 0.96 ± 0.05 | 0.96 ± 0.02 |
| PRO, g | 90 ± 4 | 91 ± 5 | 86 ± 4 | 84 ± 5 | 88 ± 2 |
| PRO, g/kg | 1.05 ± 0.04 | 1.10 ± 0.06 | 1.10 ± 0.06 | 1.08 ± 0.06 | 1.08 ± 0.02 |
| Plant PRO, % | 26.1 ± 2.0 | 28.1 ± 4.9 | 65.0 ± 2.6 ** | 31.6 ± 2.0 | 36.7 ± 3.4 |
| Animal, PRO, % | 74.0 ± 2.0 | 71.9 ± 4.9 | 35.1 ± 2.6 ** | 68.4 ± 2.0 | 63.3 ± 3.4 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saracino, P.G.; Saylor, H.E.; Hanna, B.R.; Hickner, R.C.; Kim, J.-S.; Ormsbee, M.J. Effects of Pre-Sleep Whey vs. Plant-Based Protein Consumption on Muscle Recovery Following Damaging Morning Exercise. Nutrients 2020, 12, 2049. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12072049
Saracino PG, Saylor HE, Hanna BR, Hickner RC, Kim J-S, Ormsbee MJ. Effects of Pre-Sleep Whey vs. Plant-Based Protein Consumption on Muscle Recovery Following Damaging Morning Exercise. Nutrients. 2020; 12(7):2049. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12072049
Chicago/Turabian StyleSaracino, Patrick G., Hannah E. Saylor, Brett R. Hanna, Robert C. Hickner, Jeong-Su Kim, and Michael J. Ormsbee. 2020. "Effects of Pre-Sleep Whey vs. Plant-Based Protein Consumption on Muscle Recovery Following Damaging Morning Exercise" Nutrients 12, no. 7: 2049. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12072049