1. Introduction
Secondary sclerosing cholangitis in critically ill patients (SC-CIP) is a rare, often rapidly advancing, cholestatic liver disease observed in patients without known liver disease after long-term treatment in an intensive care unit (ICU). Septic, cardiogenic or hypovolemic shock, mechanical ventilation, polytrauma, complicated surgeries, burns and severe (co-)morbidities such as obesity have been described as potential initiating factors [
1,
2,
3]. Usually, long-term ICU treatment with a mean duration of 30 to 40 days precedes the onset of this disease, but in an individual case, development of SC-CIP after only nine days of ICU stay has been reported [
1,
4].
The pathogenesis of SC-CIP remains widely enigmatic, and several potentially causative factors are of major interest: (i) ischemic injury of the biliary system, (ii) bile cast formation with protein-rich sludge and (iii) impaired innate immunity leading to recurrent biliary infections with bacteria and fungi [
5,
6]. The disease may cause destruction of the intra- and extra-hepatic biliary system with evolution of strictures leading to biliary-type liver fibrosis. Some cases may rapidly progress to cirrhosis within months, with the need for liver transplantation as the ultimate treatment option [
6]. Generally, prognosis is bad and therefore, transplant-free survival is only around 40 months [
7]. Antibiotic treatment of bacterial cholangitis and endoscopic removal of biliary casts can occasionally lead to temporary improvement [
6], but currently, liver transplantation is the only healing treatment, which, in selected patients, has excellent outcomes, similar to other indications [
8,
9].
In the last three years, data about the microbial changes of the gut and of other locations of the human body in critically ill patients has emerged [
10,
11,
12,
13]. Data about the long-term effects of an ICU stay on the gut microbiome are, however, scarce [
13]. Microbiological bile analysis from patients with SC-CIP and primary sclerosing cholangitis (PSC) showed an altered microbiological profile in these two groups, with predominance of drug-resistant organisms in the bile of SC-CIP [
14]. Data on the gut microbiome in SC-CIP is not available to date, but other chronic liver diseases have distinct changes in microbiome composition with potential impact on inflammation (non-alcoholic fatty liver disease (NAFLD), alcoholic liver disease, PSC and cirrhosis) [
15,
16,
17,
18]. Altered gut microbiome composition is thought to increase intestinal permeability [
19]. It is most commonly seen in cirrhosis but was also described in alcohol-induced injury, NAFLD and hepatits C virus (HCV)-mediated liver injury [
20,
21,
22,
23,
24]. When gut permeability is increased, bacteria from the intestinal lumen can translocate into extraintestinal locations of the body (lymph nodes, blood) and trigger inflammatory reactions, leading to disease progression [
16,
25].
Previous studies convincingly showed that bile acids have a pivotal impact on the composition of the microbiome and on gut permeability. In addition, the microbiome influences vice versa bile acid amount and composition [
26,
27]. Since the severity of liver disease was shown to correlate with impaired bile acid secretion, several factors may overlap which significantly hinder exact discrimination of cause and consequence in critically ill patients with liver disease [
27]. Gut permeability is regulated via farnesoid-X-receptor (FXR), being most commonly activated by primary bile acids [
28,
29]. Furthermore, in PSC patients with concomitant inflammatory bowel disease (IBD), the stool bile acid profile seems to allow the separation of such patients from those with solely IBD [
30]. Taken together, these findings and arguments led us to hypothesize that SC-CIP patients show alterations in the gut microbiome composition and the serum bile acid profile and that these are associated with elevated gut permeability and bacterial translocation as a basis of a pathogenic gut–liver axis.
Consequently, our current study aim was to compare the gut microbiome, gut permeability, inflammatory response and serum bile acid profile in detail in patients with SC-CIP to three different control groups. We therefore included patients suffering from SC-CIP, patients after an ICU stay without liver disease and two additional control groups consisting of patients suffering from alcohol-induced cirrhosis and of healthy probands at three different centers.
4. Discussion
Since SC-CIP is a relatively young disease entity and its occurrence is, despite growing intensive care treatment possibilities, still rare, sufficient evidence on the pathophysiology of this disease is lacking. Meanwhile, it is common knowledge that other liver diseases show alterations of the gut microbiome, gut permeability and bile acid composition [
38]. Still, the connection of these deviations with the underlying liver disease remains uncertain. Are these changes the drivers for the development of the disease or the consequence of the disease itself? To address these questions in SC-CIP, we conducted the present clinical study comparing SC-CIP patients with three different control groups. Applying current state-of-the-art methods, we could demonstrate for the first time that patients with SC-CIP have distinct changes in the stool microbiome composition, disturbed gut permeability, increased bacterial translocation and an altered serum bile acid profile compared to healthy controls. Patients treated in an ICU, who were at risk for SC-CIP, but who did not develop the disease (CIP controls), also show changes of the gut microbiome composition, increased gut permeability, as well as a distinct serum bile acid profile relative to healthy controls, which may be interpreted as long-term effects after ICU stay.
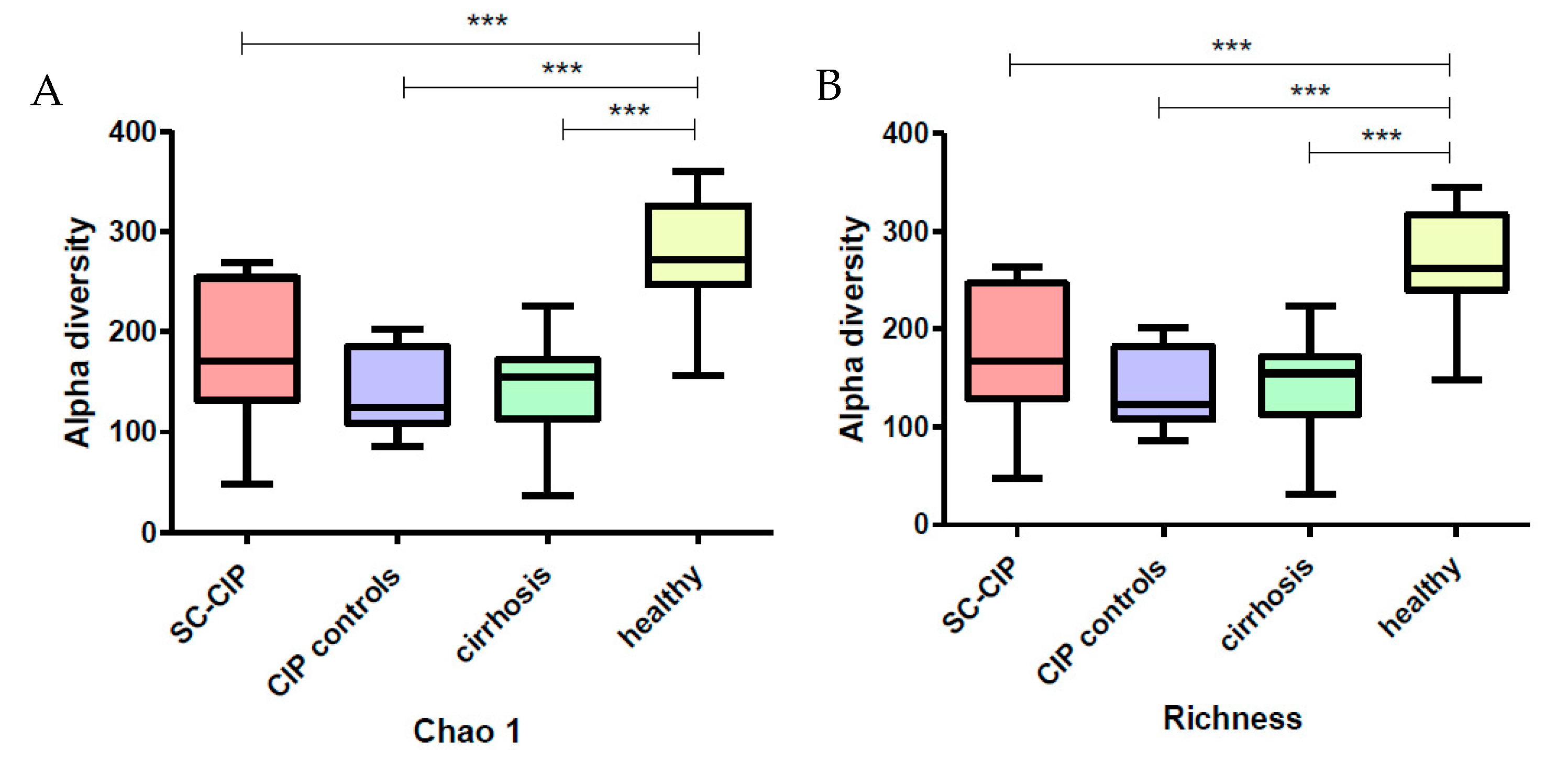
Alpha diversity was significantly reduced in SC-CIP and cirrhosis, a finding which was also observed in other liver diseases, including cirrhosis cohorts [
39,
40], as well as in obesity, inflammatory bowel disease, antibiotic treatment and ICU patients [
41,
42,
43,
44]. Interestingly, CIP controls also showed reduced alpha diversity, highlighting a potential lasting effect of the ICU stay itself on microbial diversity even more than one year after the intensive care treatment. In contrast, data from Aardema et al. suggest a faster recovery of the microbiome in their cohort of ICU patients after cardiac surgery, but the mean ICU stay of these patients was only one day [
13].
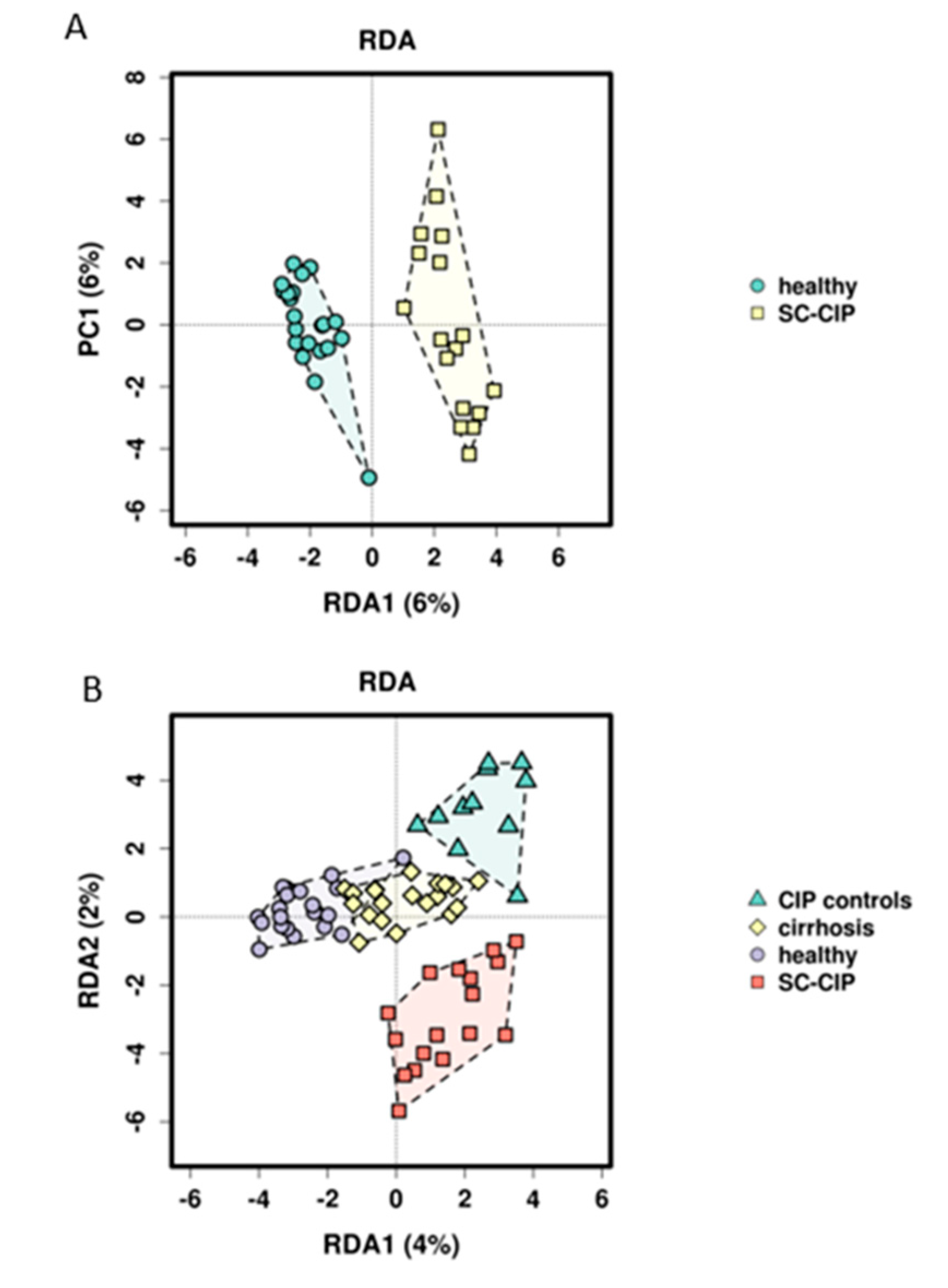
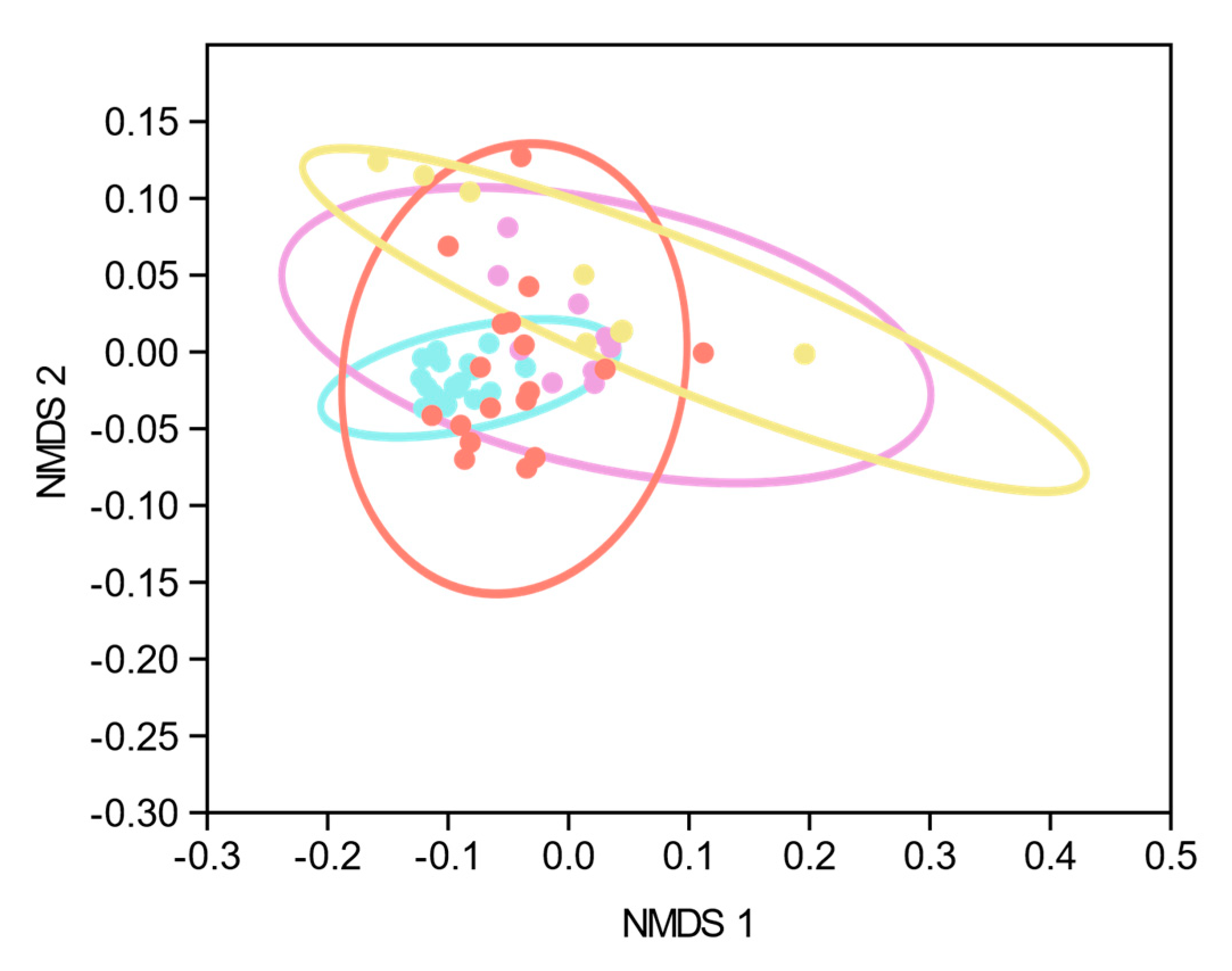
Using RDA analysis, differential clustering of SC-CIP and CIP controls compared to healthy controls could be observed. Comparable changes in beta diversity have been shown as early as 48 h after admission in ICU patients suffering from critical illnesses of different origins [
45] or in patients with early sepsis without organ failure [
46]. However, it is currently unclear how long such changes persist. Our data suggests that in SC-CIP, dysbiosis persists, since the mean time span between ICU admission and study inclusion was more than three years. The distinctly different microbiome composition of CIP control compared to healthy controls indicates that these changes persist at least partly from the critical illness and the ICU stay rather than being solely driven by the liver disease itself, and may also be influenced by co-factors like diet and drug intake.
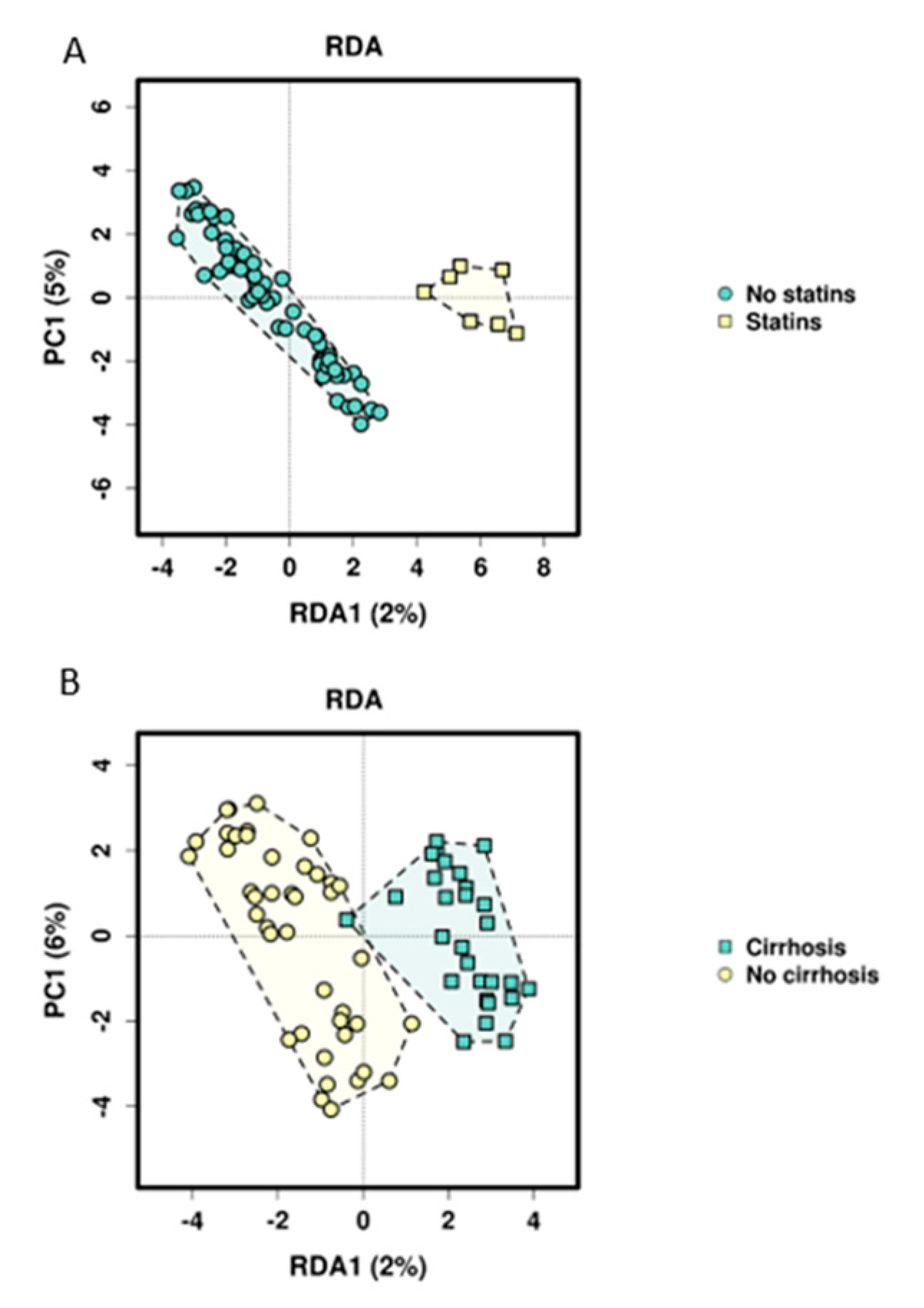
Searching for influencing factors for microbiome composition other than just group assignment, we found that statins significantly influence the microbiome in our study cohort in both applied multivariate models. This influence was also recently shown in a mouse model, indicating changes in the gut microbiome by statins and as a consequence, improving metabolic disorders such as hyperglycemia [
47]. Cirrhosis, platelet functions inhibitor intake, the BMI and calprotectin could be identified as other potential microbiome-modulating factors. The link of obesity, nutritional status, diet and microbiome composition has been described and discussed [
48,
49,
50], and the influence of cirrhosis on the microbiome as well as the role of microbiome alterations and signatures as biomarkers for diagnosis and disease progression of cirrhosis have been shown [
51]. More recent studies focused on the role of dysbiosis in the emergence of cirrhosis to hepatocellular carcinoma and its potential role as a biomarker for early carcinoma detection [
52,
53,
54,
55,
56]. The fact that NSAIDs could not be associated with microbiome composition in our study, contrary to previous literature [
57], might be caused by the small sample size and the low number of patients taking NSAIDs. Characteristics of the ICU stay could not explain differences in microbiome composition. This may also be due to the relatively small sample size and the long timespan between sample acquisition and ICU treatment.
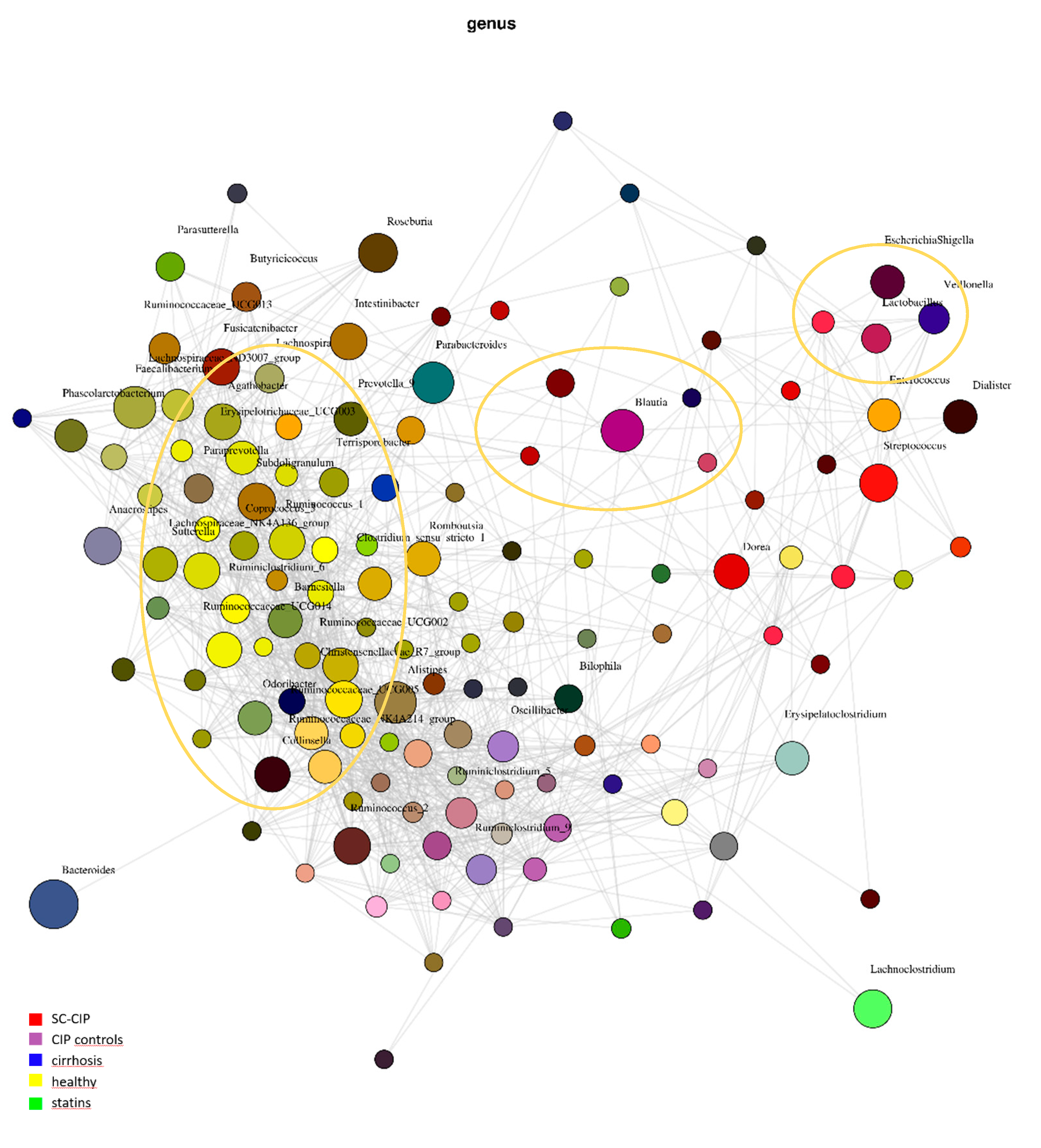
As expected from the literature [
40,
58], differences on all taxonomic levels could be observed in SC-CIP and cirrhosis compared to healthy controls, even though the patients in our cohort were stable outpatients with a rather mild course of disease. Only 7 patients were suffering from cirrhosis at the time of the study and none of them had decompensated cirrhosis. Furthermore, the magnitude of differentially abundant species in CIP controls compared to healthy controls was striking, because these patients were included into the study on average more than 1 year after intensive care treatment and did not suffer from liver disease. The dominant phylum observed in all four groups were the phyla Firmicutes and Bacteriodetes, followed by Proteobacteria and Actinobacteria. SC-CIP patients had an increased Firmicutes/Bacteriodetes ratio compared to all three other groups. This increased ratio has recently been associated with obesity, irritable bowel syndrome, leaky gut, NAFLD, cirrhosis and ICU mortality, although it was found not to be a good biomarker for critically ill patients because the Firmicutes/Bacteriodetes ratio may be normal when both phyla are equally affected [
38,
40,
59,
60].
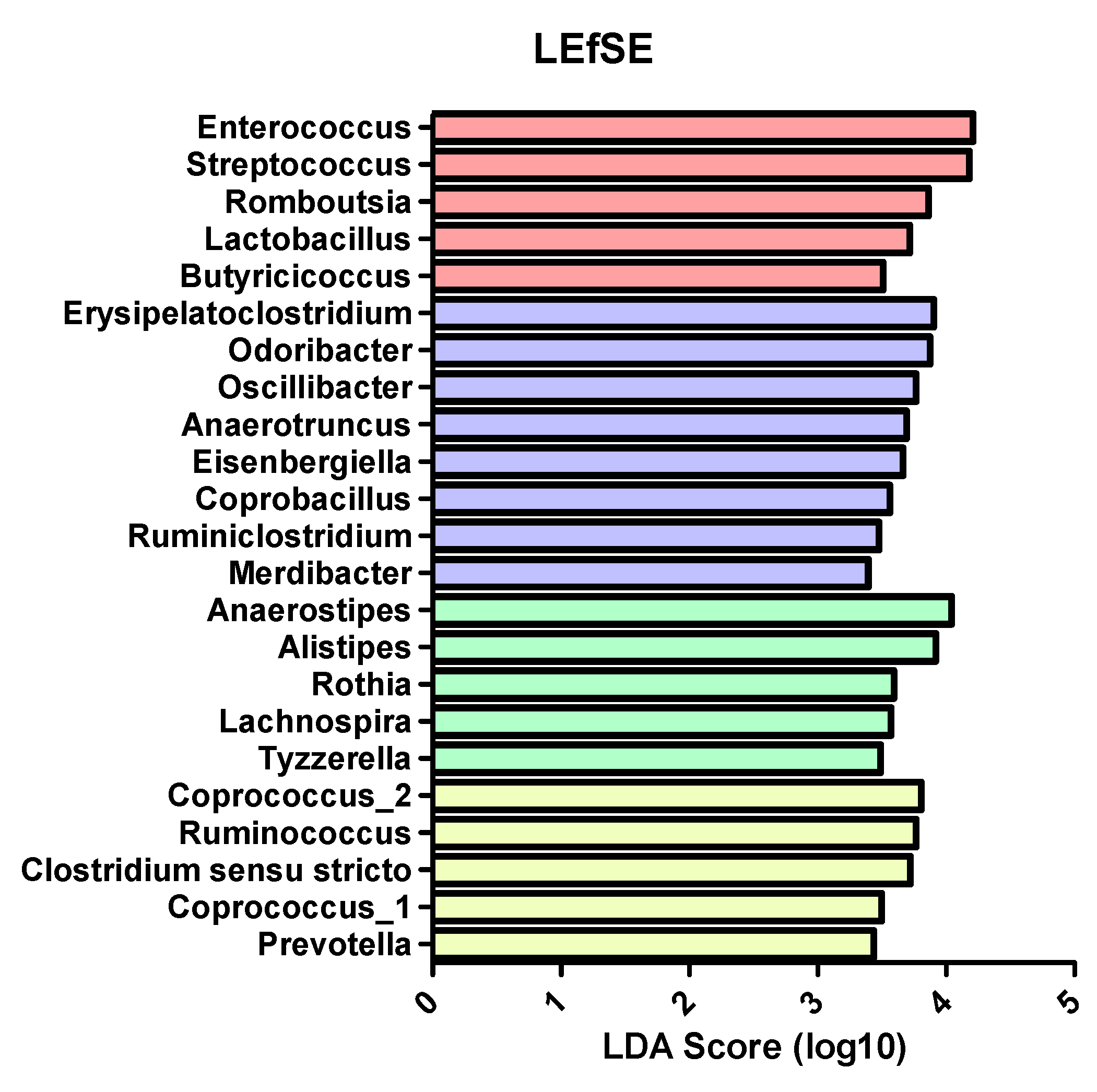
In more detail, there seems to be a shift towards pathogenic taxa in SC-CIP compared to healthy controls, highlighted by a high abundance of
Streptococcus and
Enterococcus in SC-CIP. High abundances of these genera have previously been observed in cirrhosis, acute on chronic liver failure and NAFLD, and is discussed to influence the clinical phenotype and prognosis [
18,
38]. Freedberg et al. [
61] showed, in their work on the influences of PPI on the microbiome, that these microbiome alterations may increase the risk for infections with
Clostridioides difficile. An oralization of the stool microbiome in SC-CIP with species that are normally abundant in the mouth and the upper gastrointestinal tract became evident.
Rothia dentocariosa, a bacterium normally found in the mouth of healthy humans with a potential pathogenic potential [
62], and
Streptococcus parasanguinis, a species associated with a healthy oral microflora [
63], were elevated in the stool of SC-CIP. The partially antibiotic-resistant pathogen
Enterococcus faecium [
64] was more abundant in SC-CIP, which was also previously described in long-stay ICU patients [
44]. In SC-CIP, the probiotic strain
Streptococcus thermophiles was elevated, and in healthy controls, the butyrate producing strain
Anaerostipes hadrus was elevated. Butyrate plays a crucial role in the gut as one of the most important metabolites, being an energy source of the intestinal epithelium and inhibiting inflammation in a healthy gut. In a mouse model,
Anaerostipes hadrus showed the ability to harm the inflamed intestinal epithelium, suggesting that bacterial strains can play different roles depending on the setting [
65].
A proportion of critically ill patients at an ICU need long-term treatment, sometimes over weeks, with invasive ventilation, vasoactive drugs and invasive procedures. Further on, they receive multiple medications including antibiotics, antiviral and antifungal therapies and enteric or parenteral nutrition. That all these interventions together with the underlying critical illness alter the microbiome on different body sites has been shown recently, as well as that these changes may trigger hospital-acquired infections, sepsis, multi-organ failure, energy homeostasis disturbance, muscle wasting and cachexia [
10,
12,
13,
44,
66,
67,
68]. One reason for this may be the slower bowel transit time in critically ill patients caused by electrolyte and glucose disturbances as well as by therapeutics like sedatives and opiates [
66]. Therefore, therapeutic efforts with approaches such as probiotics, symbiotics or fecal microbiota transplants have been investigated [
68,
69,
70,
71,
72,
73,
74]. Our cohort of critically ill patients who did not develop SC-CIP had all been discharged from ICU a year before study inclusion on average, but still, taxonomic differences remained on all levels compared to healthy controls. The reduction of
Faecalibacterium prausnitzii, a species thought to be beneficial for survival and function of enterocytes due to the production of short-chain fatty acids [
75] and known to show reduced abundance in many intestinal disorders [
76], as well as elevation of
Erysipelatoclostridium ramosum, a species involved in the development of metabolic syndrome [
77], were observed. With the knowledge that both SC-CIP and CIP controls independent of the presence of a liver disease show dysbiosis compared to healthy controls, the question remains if certain microbial differences could trigger the development of SC-CIP. Indeed, we could show that the microbiome of these two cohorts differs widely. A shift of highly abundant classes like
Bacilli and
Bacteroidia could be shown as well as changes going down to the species level, with reduction of an unnamed
Neisseria species and
Prevotella melaninogenica in SC-CIP. Dysbiosis of the intestinal microbiome is a known feature of alcohol-induced cirrhosis.
Bifidobacteria, Streptococci, Veillonella and
Enterobacteria were found to be elevated in cirrhosis, whereas
Faecalibacterium showed a reduction [
58,
76,
78]. In our cohort, we could observe reduced diversity, and on a genus level, also increased abundance of
Veillonella compared to healthy controls. This species could cause joint infections, endocarditis and was associated with hepatic encephalopathy [
66,
67,
68], but can also be an effect of PPI intake of some cirrhotic patients [
79].
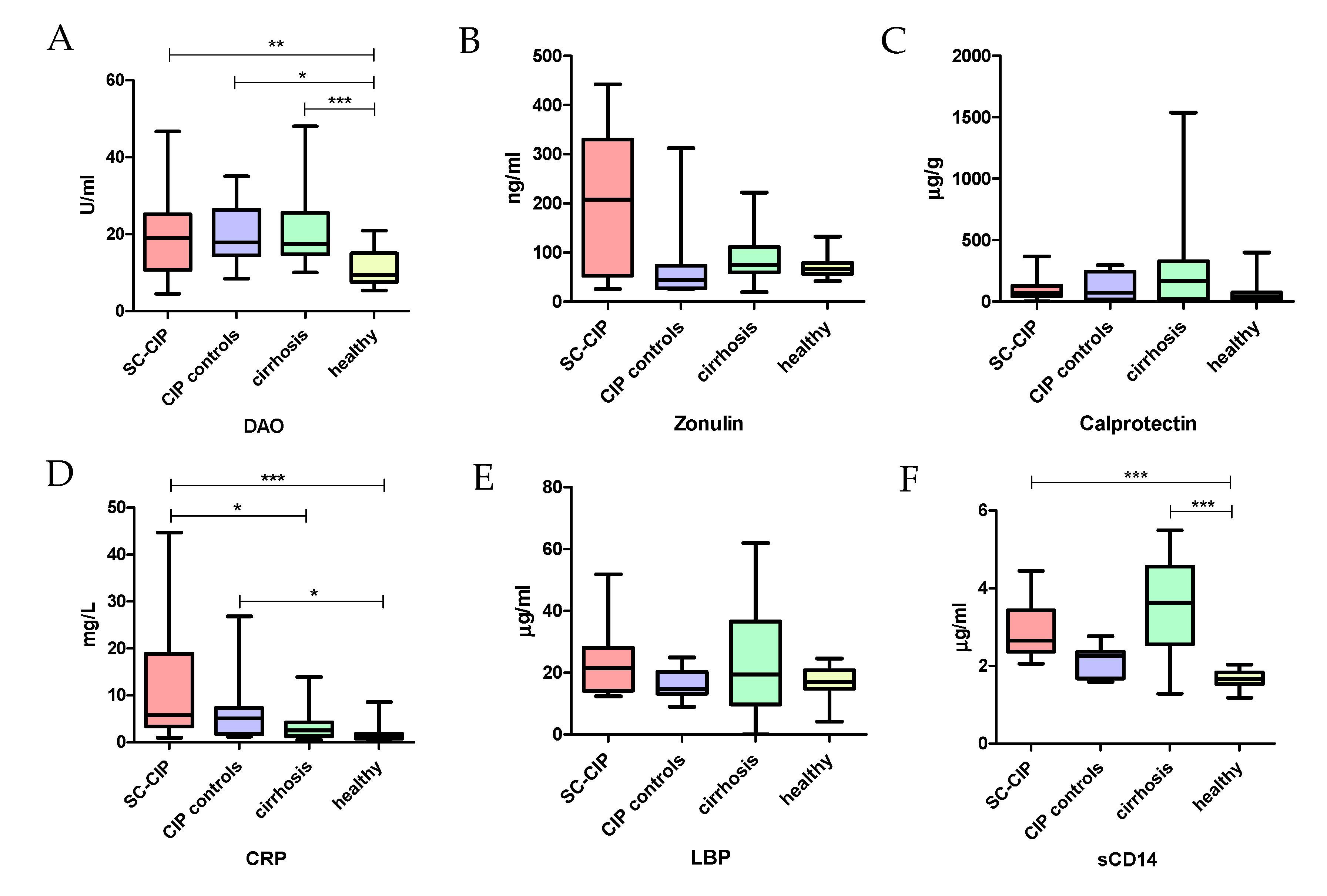
Alterations of the gut microbiome lead to increased inflammation, a leaky gut and endotoxemia, not only in gastrointestinal pathologies but also in liver disease [
80]. In our study, we could demonstrate this association and potential pathophysiological link for the first time in SC-CIP. Compared to healthy controls, SC-CIP patients had higher biomarkers of gut permeability (DAO), systemic inflammation (CRP) and bacterial translocation (sCD14), although none of the patients had clinically apparent signs of infection when the samples were taken. DAO can be found in several human tissues but is particularly present in the intestine and correlates with the integrity of the intestinal mucosa. Elevated DAO levels are a serum biomarker for increased intestinal permeability [
81]. sCD14 is an LPS binding glycoprotein which can be found on the surface of macrophages and monocytes and is produced during inflammation and sepsis. Elevated serum levels are considered to be a surrogate biomarker for endotoxemia [
82]. Our study suggests that this may be a consequence of the intensive care treatment rather than an effect of the liver disease alone, since increased gut permeability could also be shown in CIP controls. Enterocyte damage plays a role in the outcome of critically ill patients, as suggested by increased plasma intestinal fatty acid-binding protein and citrulline concentrations in plasma [
83]. The positive correlation of inflammation and gut permeability markers with important parameters of liver dysfunction, as observed in our current study, suggests a connection between gut inflammation and the severity of the liver disease. This was also shown previously by our study group [
84].
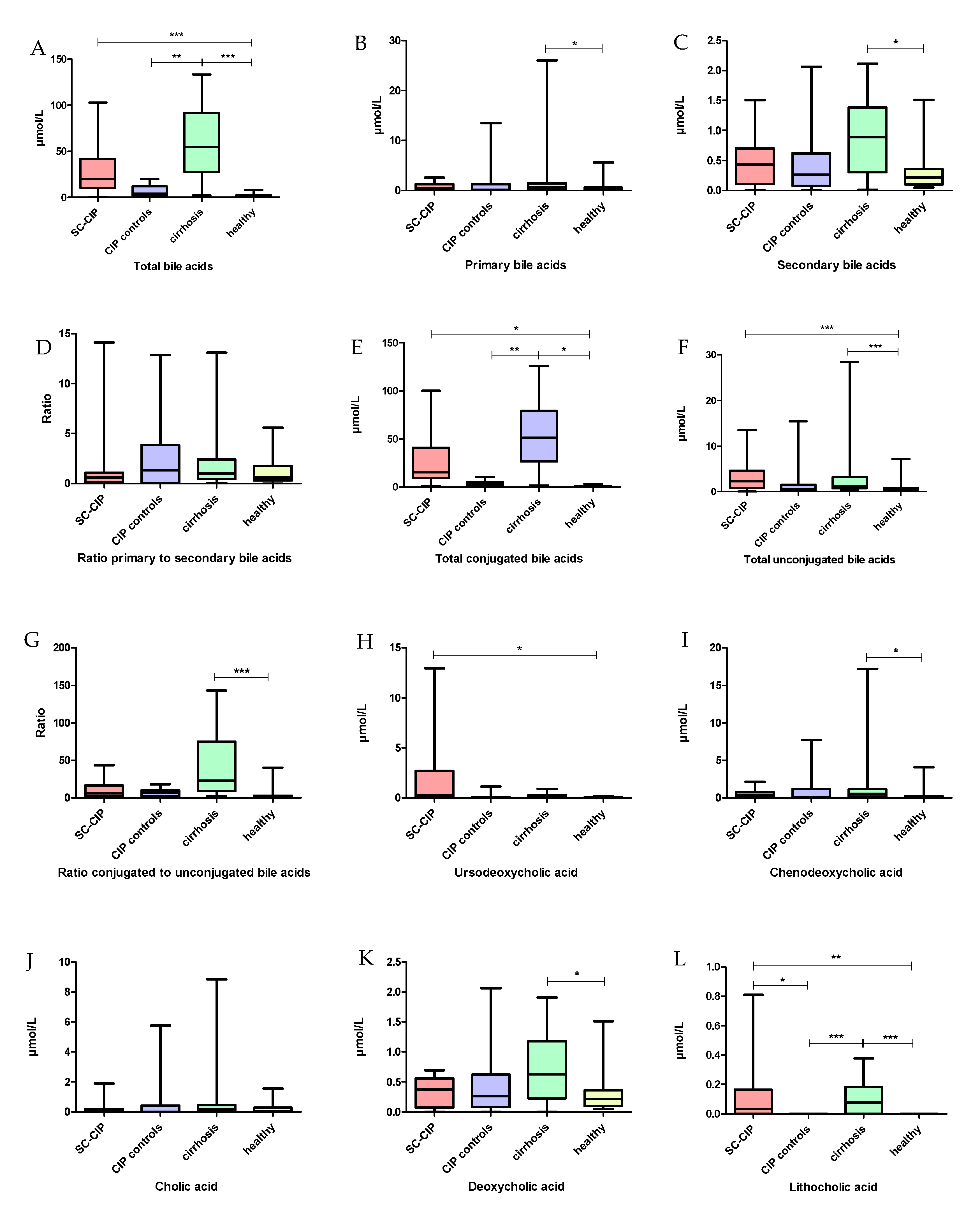
When searching for factors that may connect dysbiosis and intestinal wall integrity in a cholestatic disease, bile acids need to be considered. Bile acids consist of steroid molecules and can be divided into primary bile acids, which are produced in the liver, and into secondary bile acids, which are the result of primary bile acids being metabolized by the gut microbiome [
85]. This bile acid homeostasis is controlled by the intestinal microbiome [
86]. Thus, changes in the microbiome by inflammation, diet or antibiotic intake may also lead to changes in the bile acid profile [
87]. As expected, in a cholestatic liver disease, an increase in total serum bile acids was found in SC-CIP. This has previously also been published for other cholestatic liver diseases, such as primary biliary cholangitis, primary sclerosing cholangitis and NAFLD [
88,
89]. Furthermore, bile acid profiles were altered in SC-CIP and cirrhosis compared to healthy controls. Other than in primary and secondary, bile acids can also be divided into conjugated and unconjugated bile acids. This conjugation in adults aims to improve hydrophily and protects against toxic effects of the bile acids. In the intestine, primary conjugated bile acids are deconjugated by bacterial bile salt hydrolase [
90,
91]. As in primary biliary cholangitis and primary sclerosing cholangitis, conjugated bile acids were elevated in SC-CIP and cirrhosis serum [
89]. An increase in taurine-conjugated bile acids in gall bladder aspirates of mice was associated with the expansion of pathobionts and the development of colitis [
92], and may therefore be part of the cause and/or consequence of the increase of potential harmful bacterial species. Germ-free or antibiotic-treated rodents also showed an elevation of taurine-conjugated primary BA species in plasma and in the liver; therefore, the observed elevation of conjugated bile acids in the SC-CIP population may be linked to the loss in microbiome diversity and antibiotic use in critically ill patients [
93].
Our study has several important limitations. The cross-sectional study design does not allow to establish causal relationships. SC-CIP is a rare disease; therefore, the sample size is rather small, although three centers contributed to the study. Also, recruiting patients as CIP controls was challenging. Collection of stool samples at only one time point from each proband makes the analysis more vulnerable for daily fluctuations of the microbiome, and the cross-sectional design does not allow conclusions on longitudinal effects. Furthermore, 16S sequencing, although the current standard method for microbiome analysis, provides less microbial resolution compared to whole metagenome sequencing techniques, which can lead to considerable limitations in species level identification. Therefore, only those sequences which unambiguously and exactly matched a reference genome were analyzed on this level.

 ,
, 








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}