The Impact of FTO Genetic Variants on Obesity and Its Metabolic Consequences is Dependent on Daily Macronutrient Intake

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Anthropometric and Body Composition Measurements
2.3. Blood Collection, Biochemical Analysis, and Calculations
2.4. Daily Physical Activity and Dietary Intake Analyses
2.5. Genetic Analyses
2.6. Ethics Statement
2.7. Statistical Analysis
3. Results
3.1. Dietary Assessment
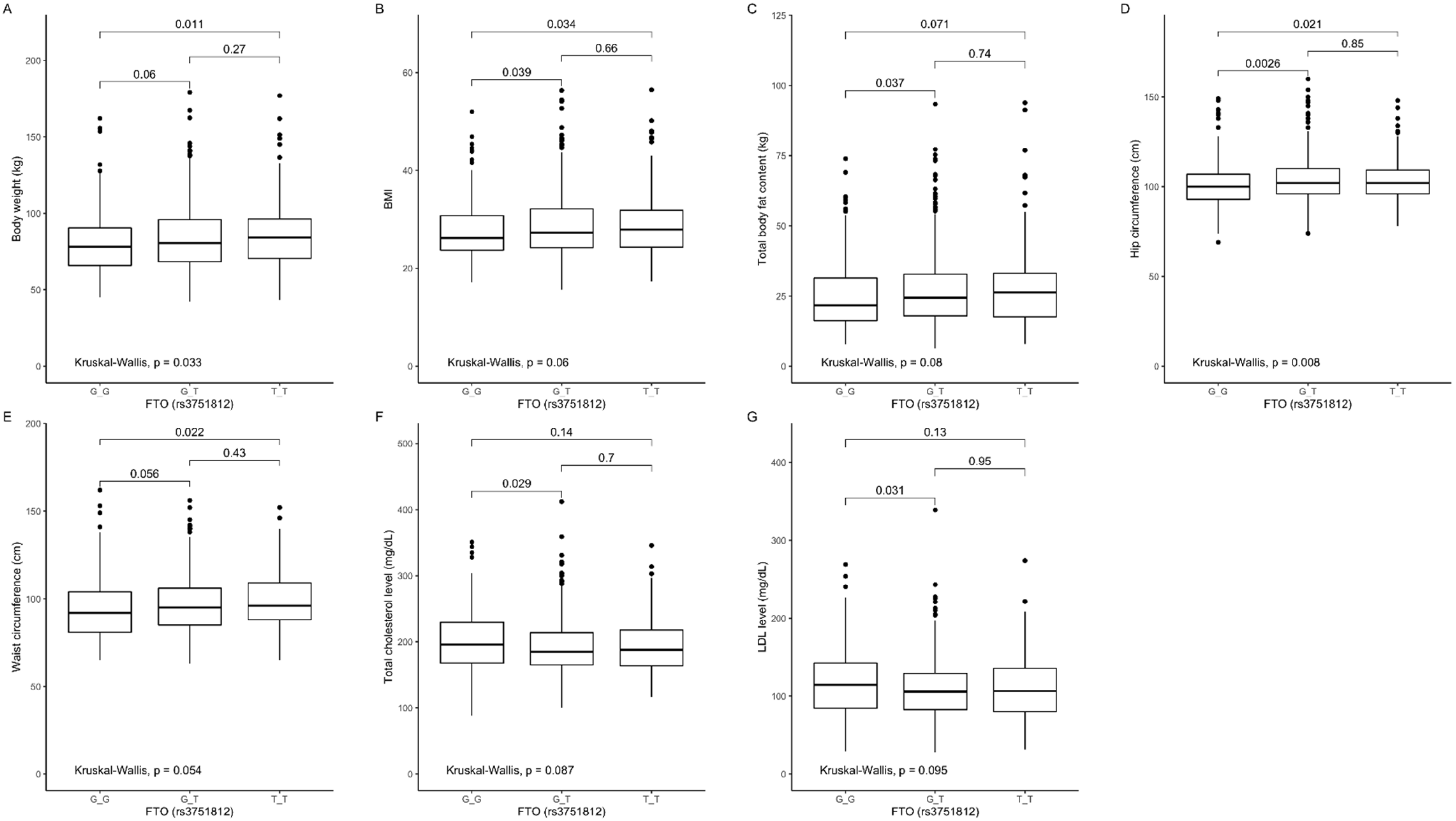
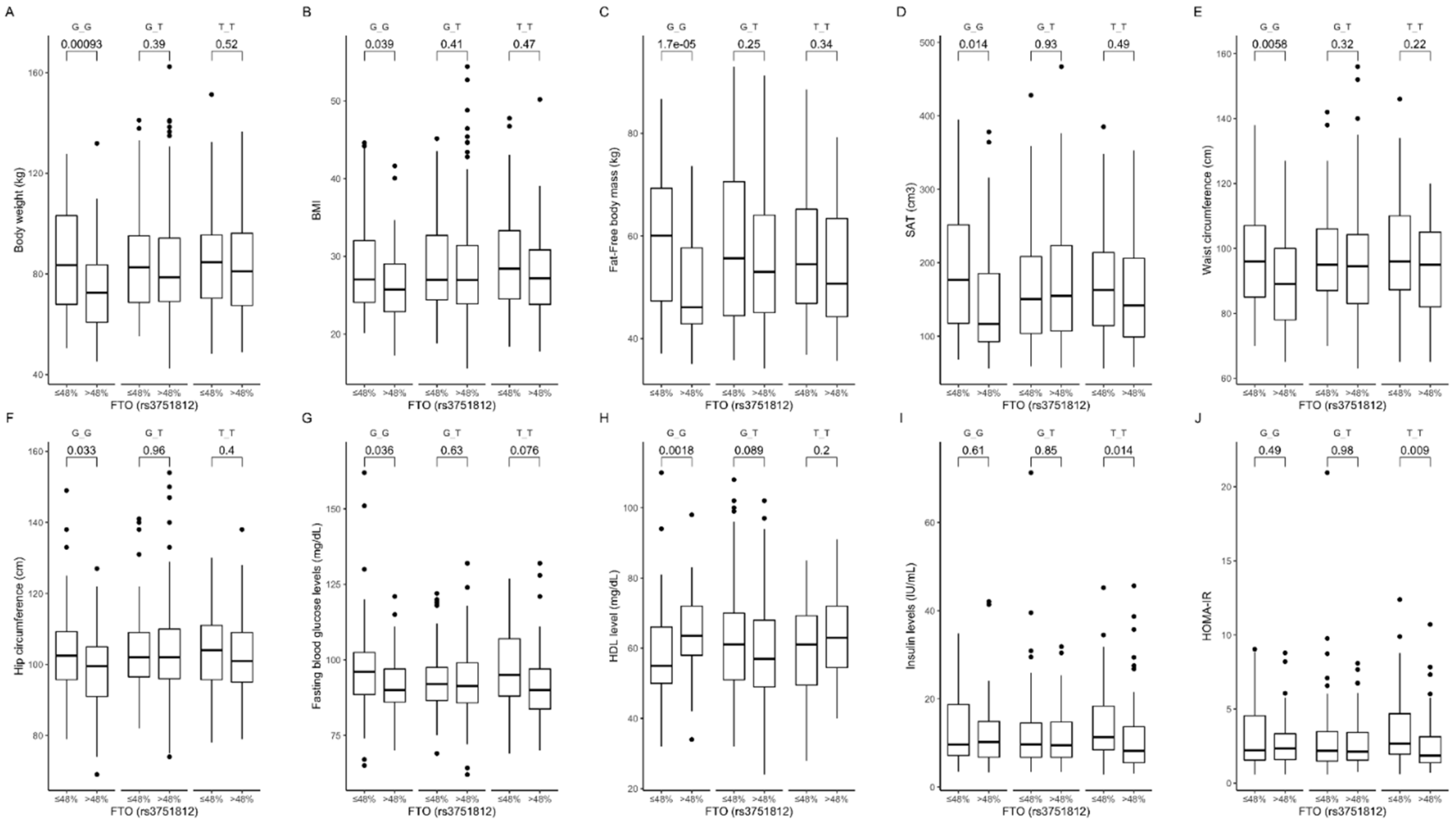
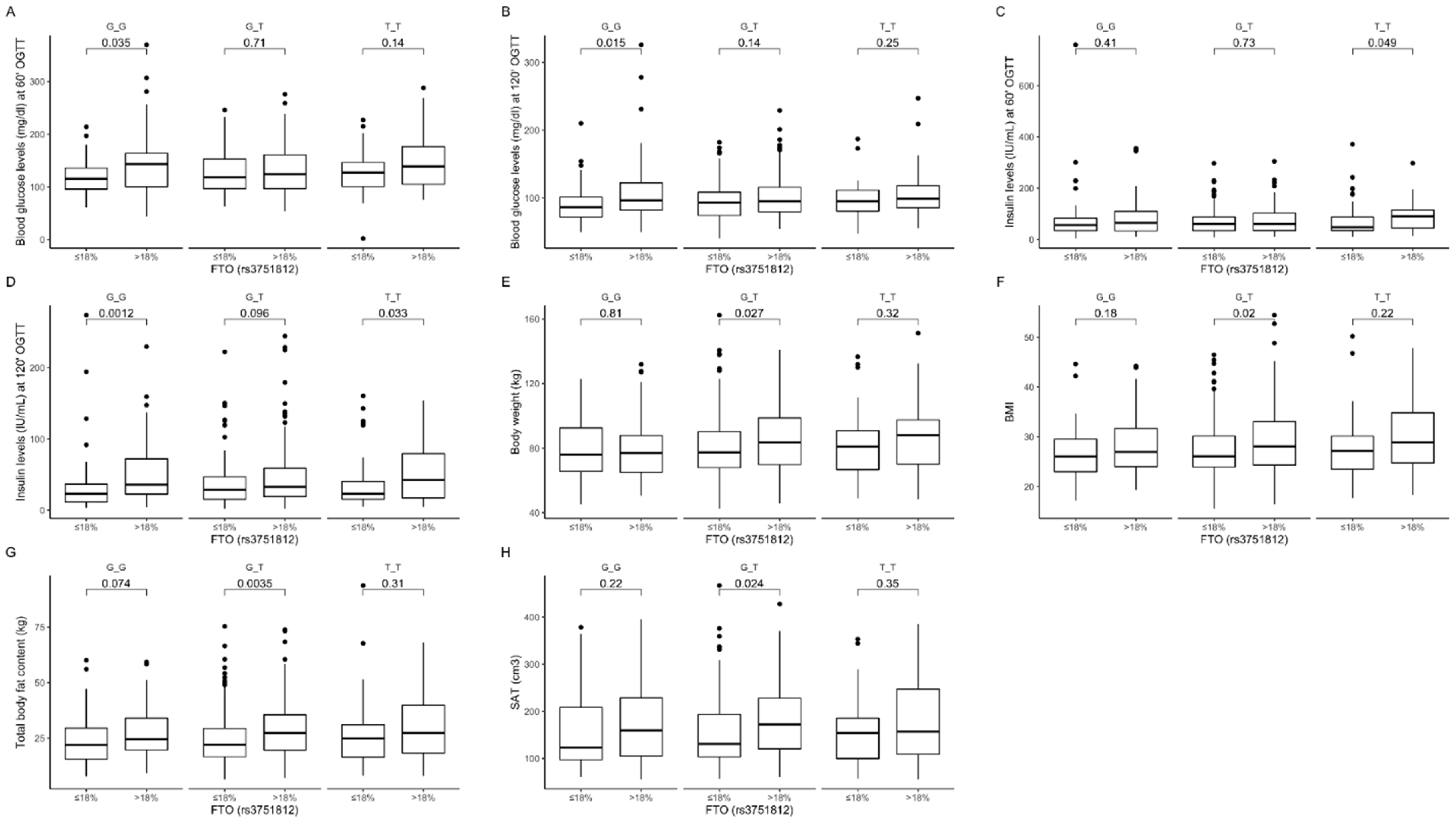
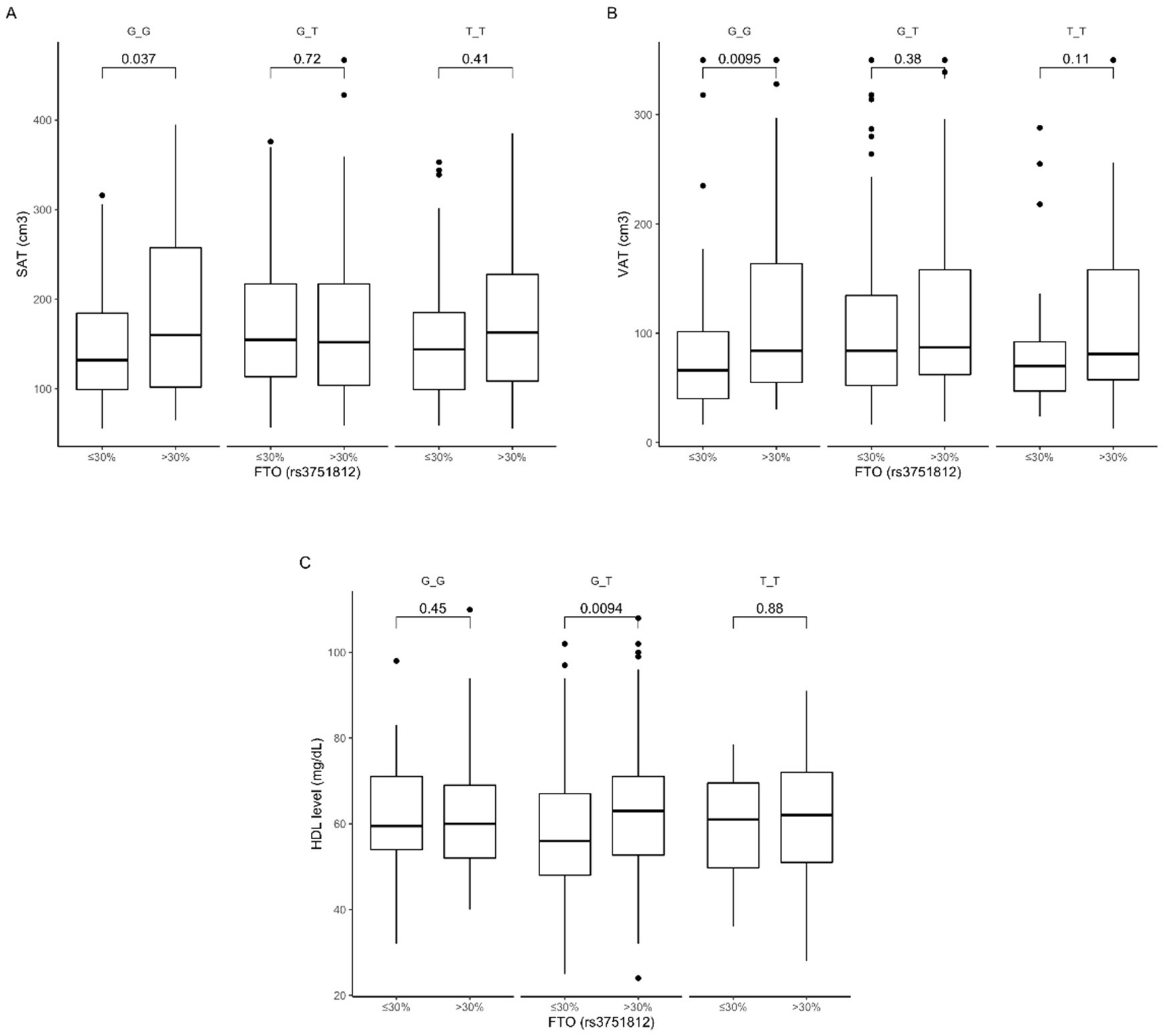
3.2. Association of rs3751812 Genetic Variants with Obesity, Anthropometric Measures, Lipid Profile, and Dietary Intake
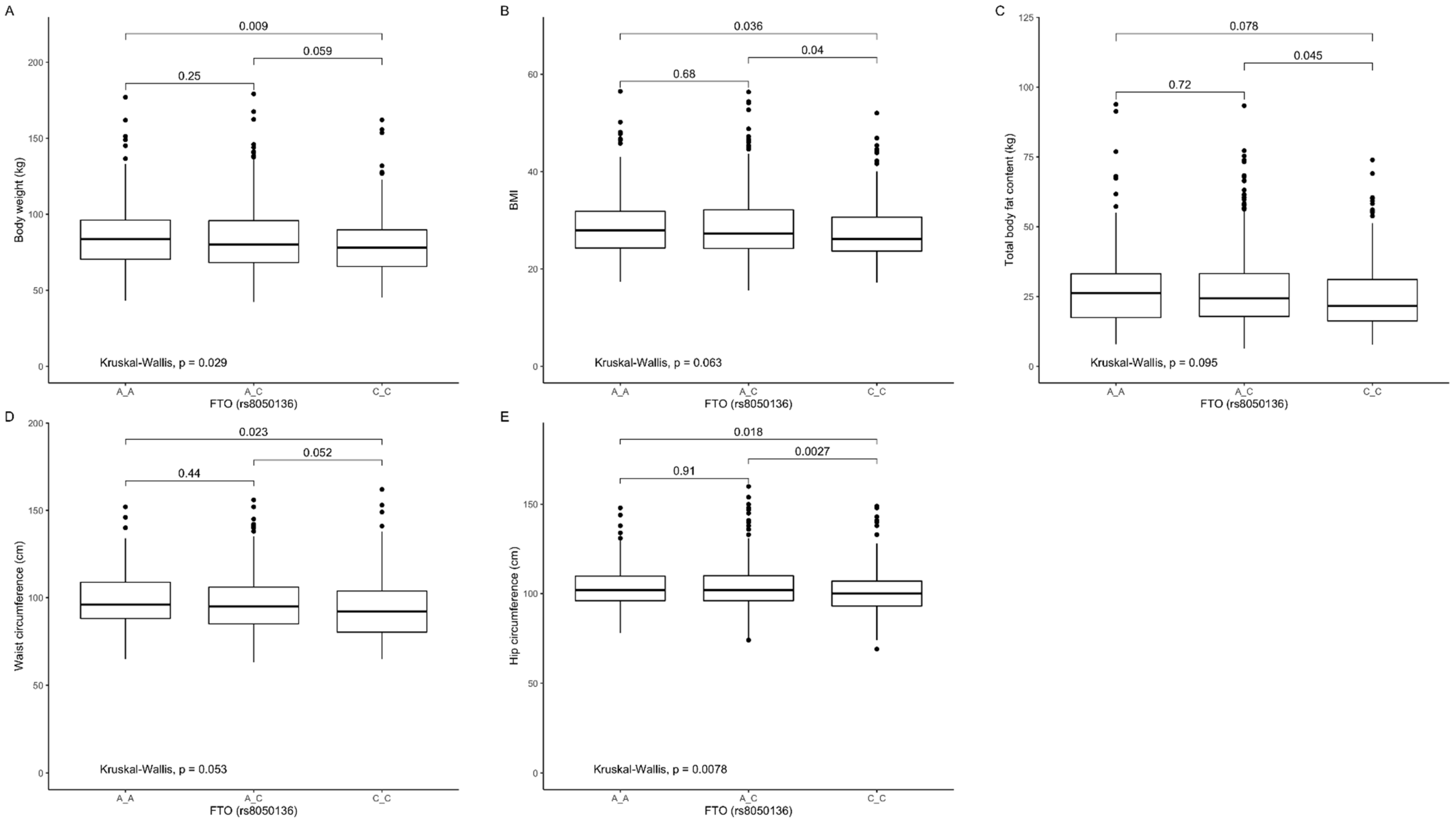
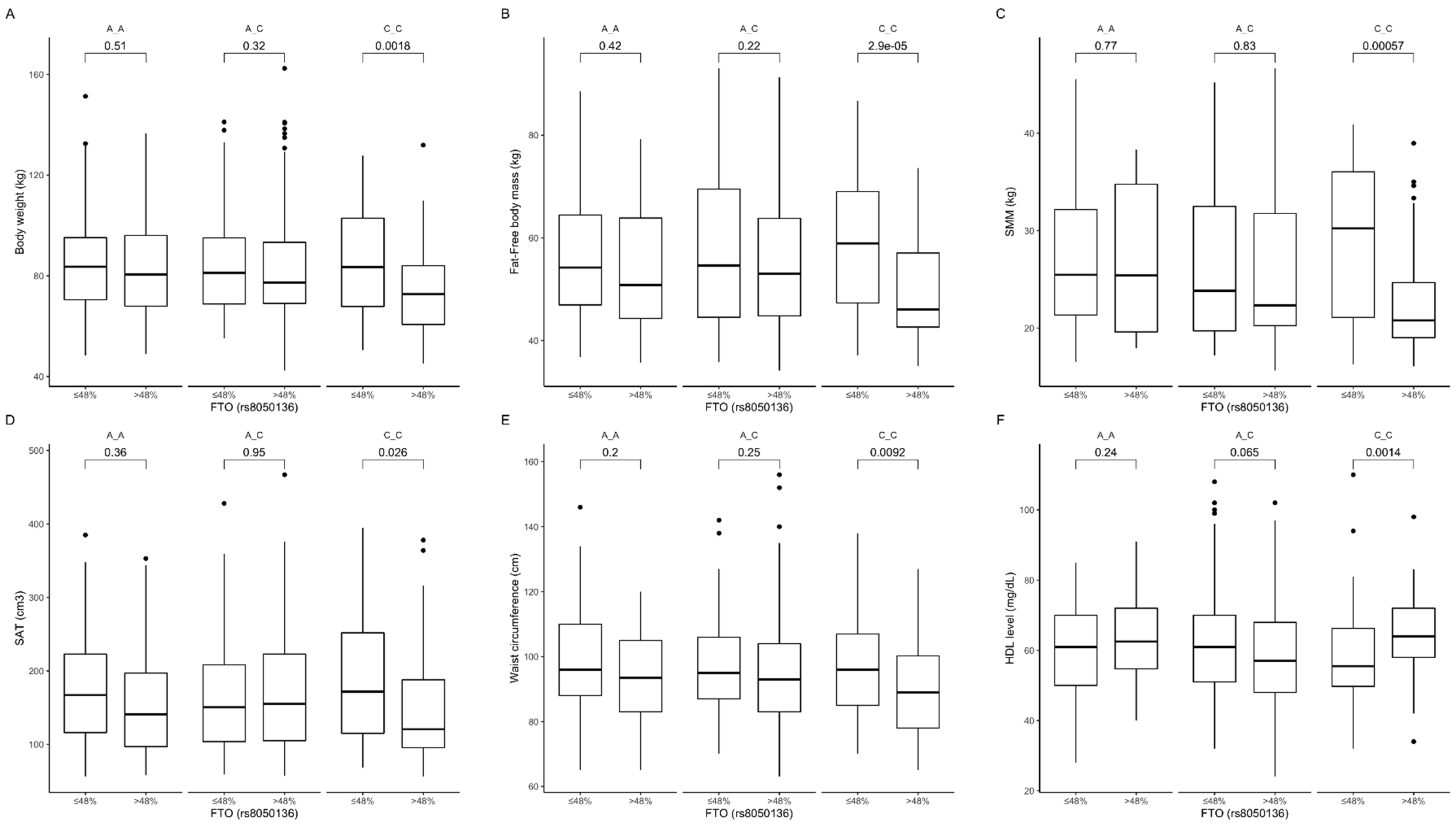
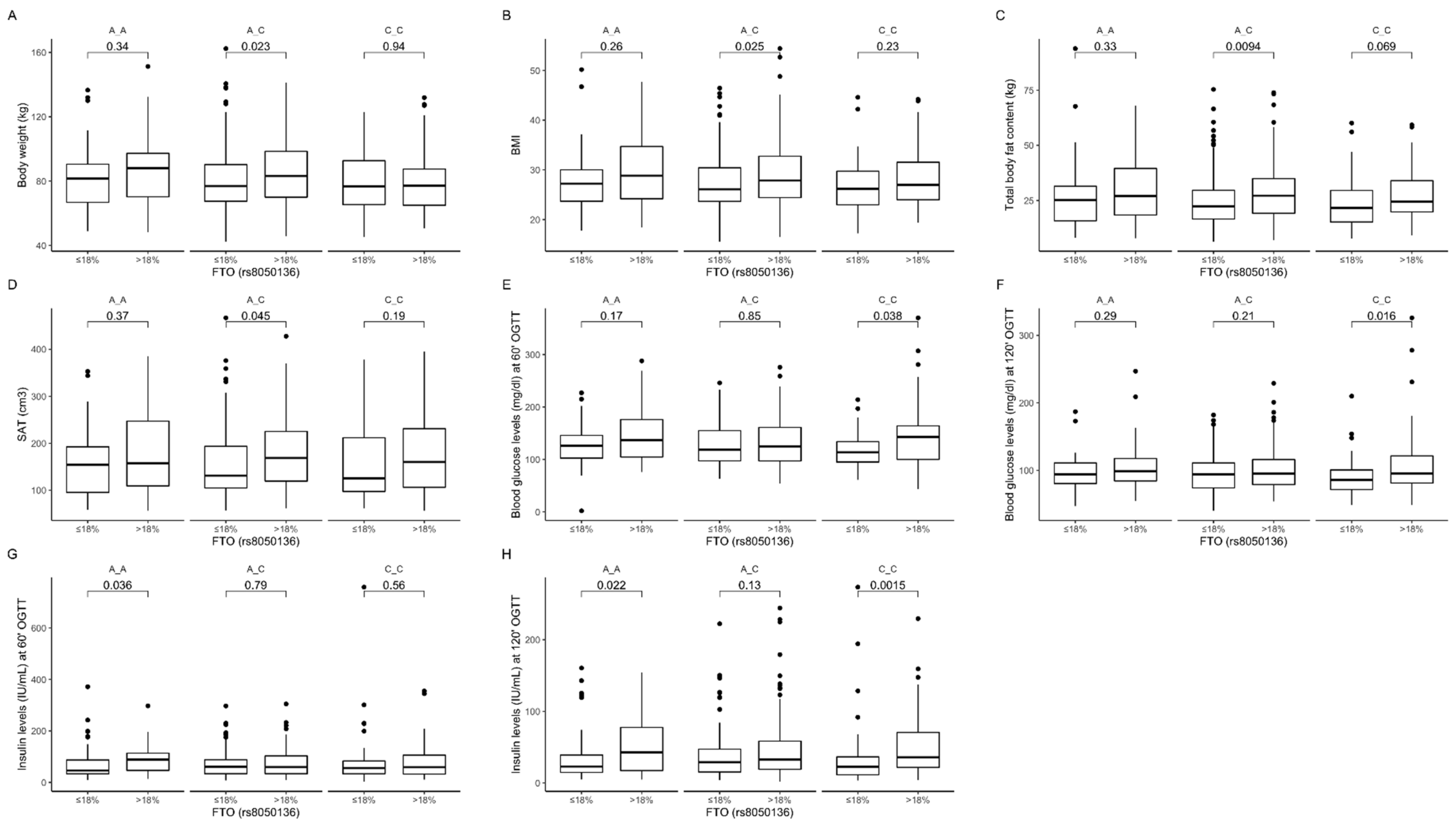
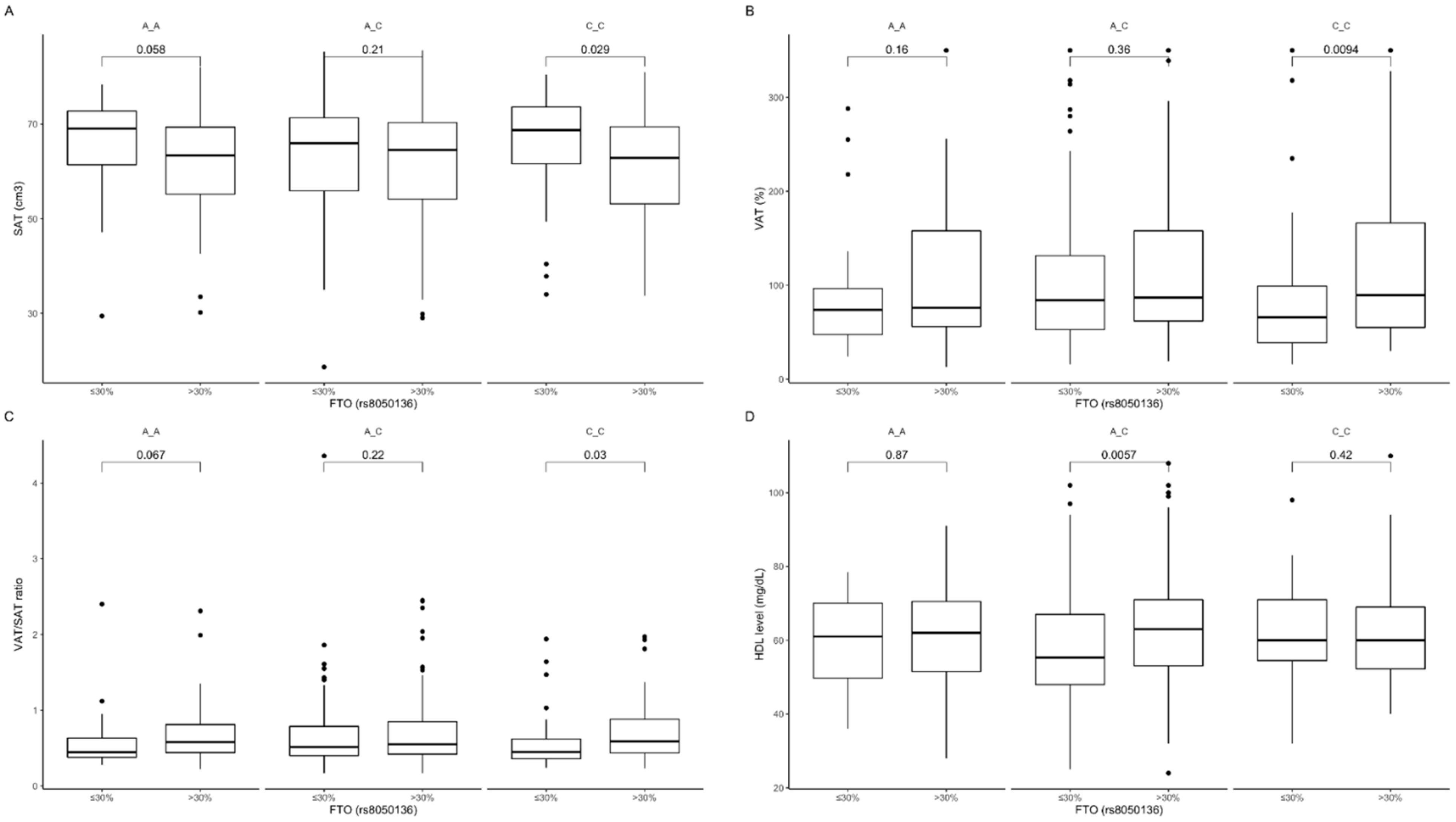
3.3. Association of rs8050136 Genetic Variants with Obesity, Anthropometric Measures, Lipid Profile, and Dietary Intake
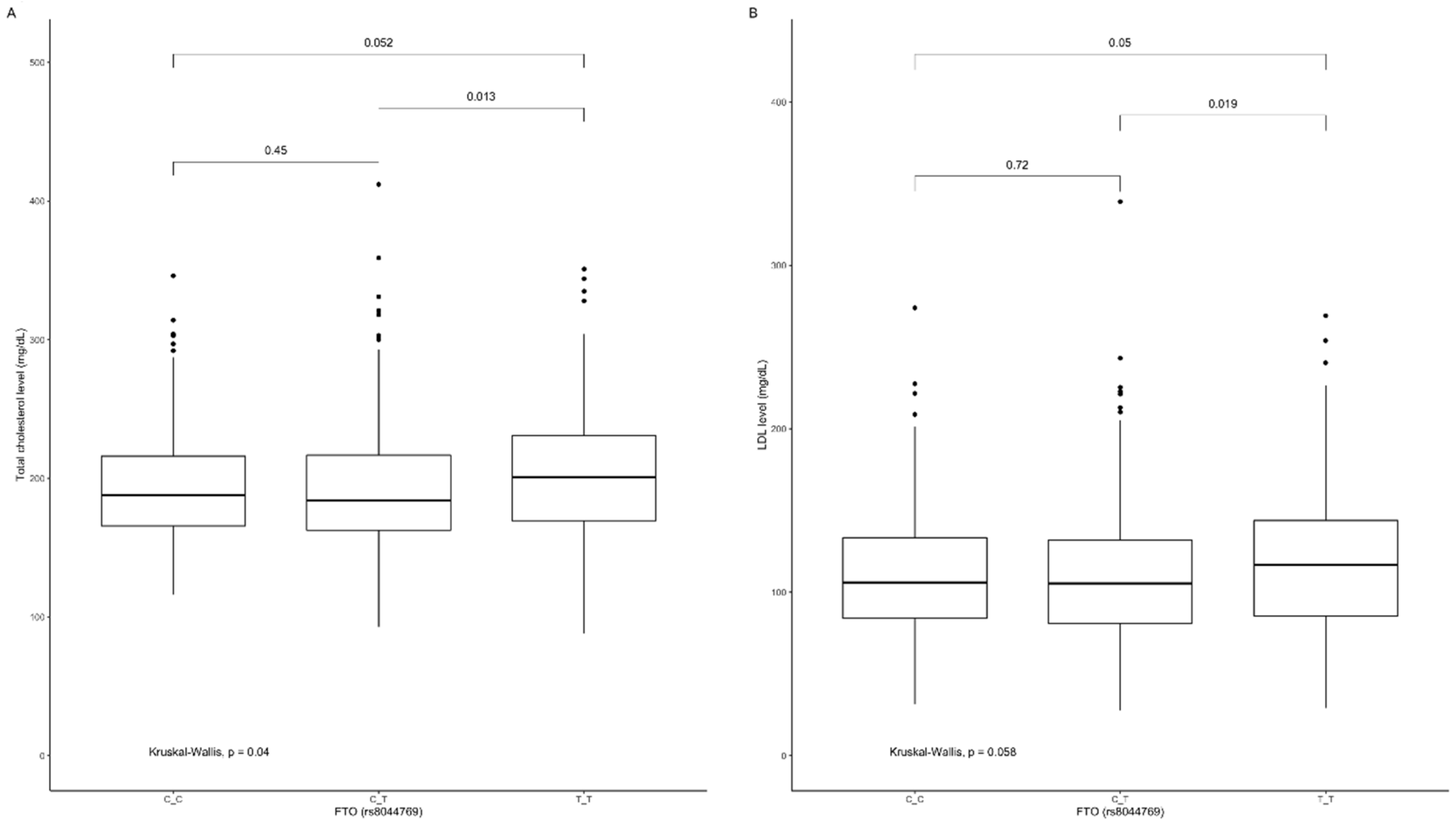
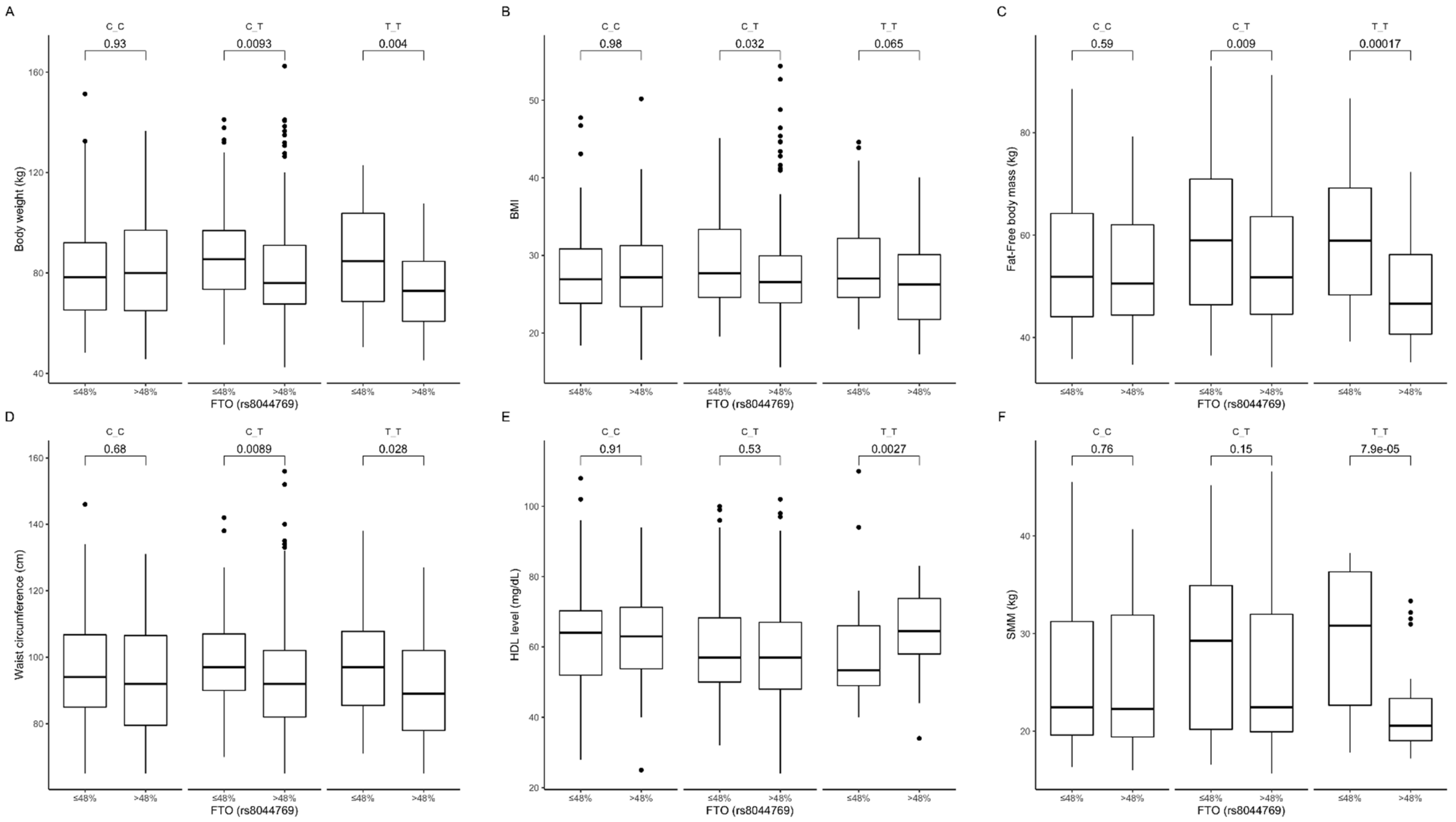
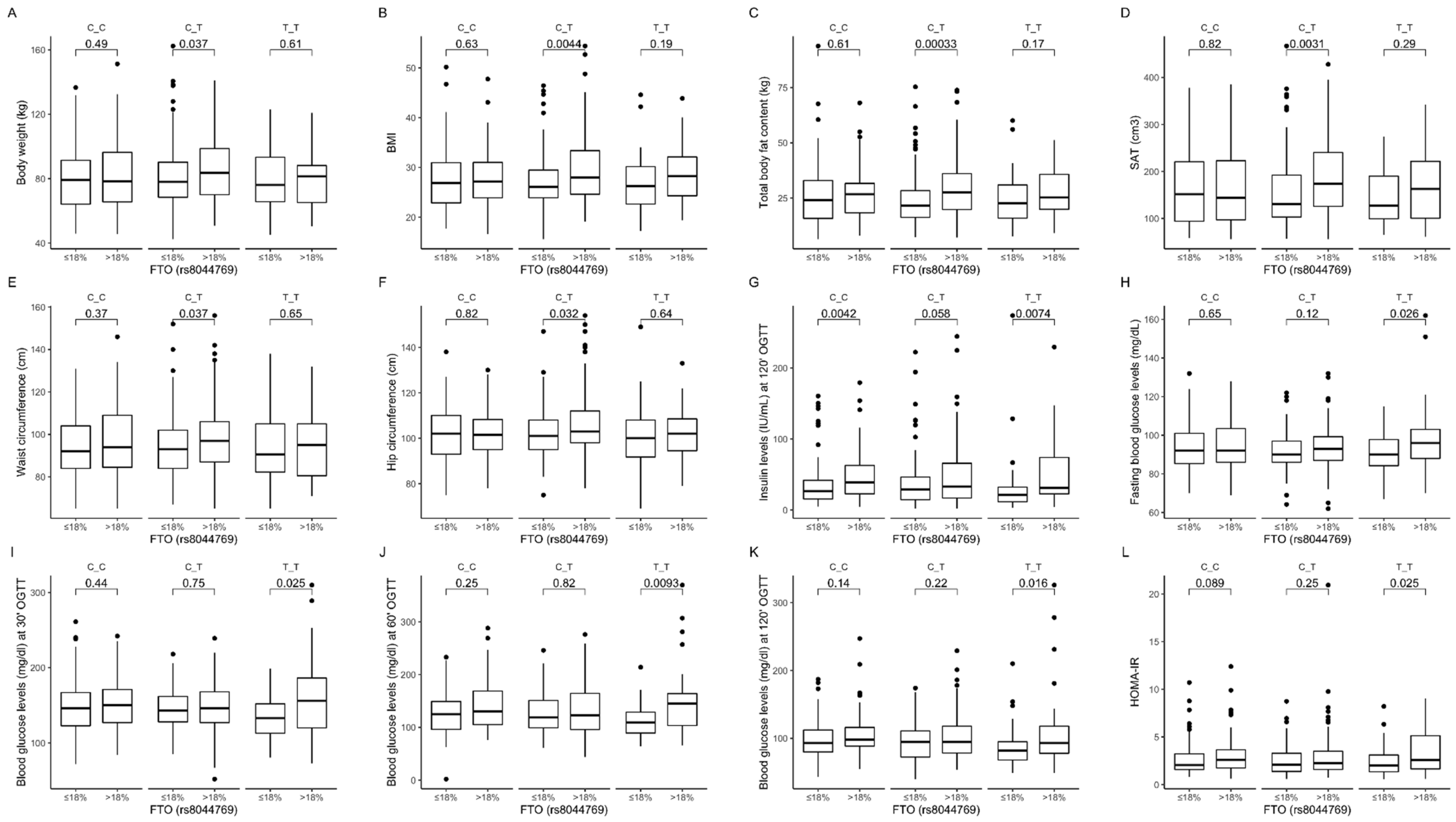
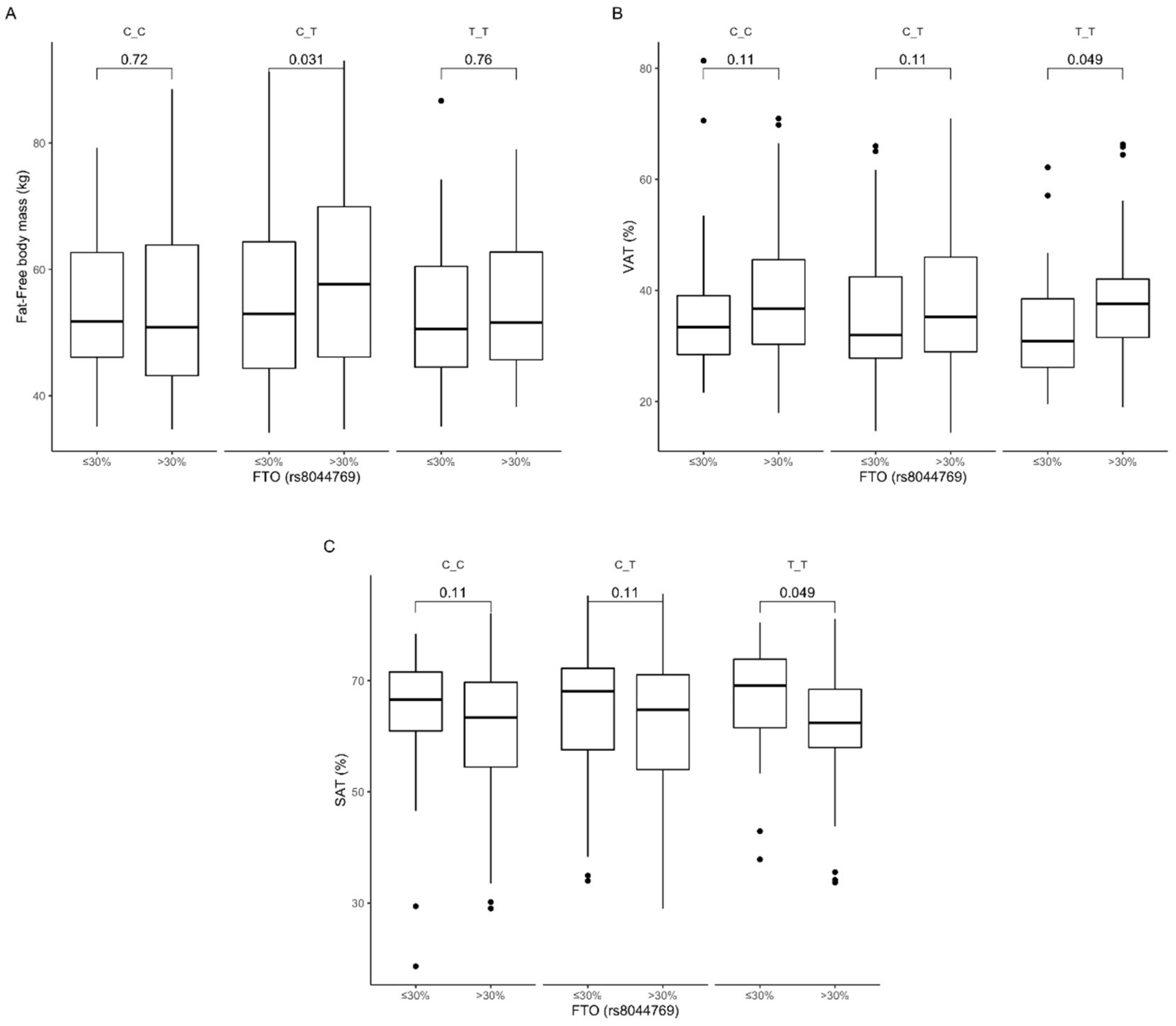
3.4. Association of rs8044769 Genetic Variants with Obesity, Anthropometric Measures, Lipid Profile, and Dietary Intake
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jaacks, L.M.; Vandevijvere, S.; Pan, A.; McGowan, C.J.; Wallace, C.; Imamura, F.; Mozaffarian, D.; Swinburn, B.; Ezzati, M. The obesity transition: Stages of the global epidemic. Lancet Diabetes Endocrinol. 2019, 7, 231–240. [Google Scholar] [CrossRef]
- Loh, M.; Zhou, L.; Ng, H.K.; Chambers, J.C. Epigenetic disturbances in obesity and diabetes: Epidemiological and functional insights. Mol. Metab. 2019, 27S, S33–S41. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.F. Big challenges: Obesity and type 2 diabetes in children and adolescents. World J. Pediatr. 2019, 15, 313–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Kelly, A.S. Review of Childhood Obesity: From Epidemiology, Etiology, and Comorbidities to Clinical Assessment and Treatment. Mayo Clin. Proc. 2017, 92, 251–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K.; et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas, 9(th) edition. Diabetes Res. Clin. Pract. 2019, 157, 107843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.R.; Zhang, M.Y.; Ni, J.W.; Cheng, R.Q.; Zheng, Z.Q.; Xi, L.; Luo, F. Clinical characteristics and beta-cell function of Chinese children and adolescents with type 2 diabetes from 2009 to 2018. World J. Pediatr. 2019, 15, 405–411. [Google Scholar] [CrossRef] [PubMed]
- Dal Canto, E.; Ceriello, A.; Rydén, L.; Ferrini, M.; Hansen, T.B.; Schnell, O.; Standl, E.; Beulens, J.W. Diabetes as a cardiovascular risk factor: An overview of global trends of macro and micro vascular complications. Eur. J. Prev. Cardiol. 2019, 26 (Suppl. 2), 25–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorleifsson, G.; Walters, G.B.; Gudbjartsson, D.F.; Steinthorsdottir, V.; Sulem, P.; Helgadottir, A.; Styrkarsdottir, U.; Gretarsdottir, S.; Thorlacius, S.; Jonsdottir, I.; et al. Genome-wide association yields new sequence variants at seven loci that associate with measures of obesity. Nat. Genet. 2009, 41, 18–24. [Google Scholar] [CrossRef]
- Loos, R.J. Genetic determinants of common obesity and their value in prediction. Best Pract. Res. Clin. Endocrinol. Metab. 2012, 26, 211–226. [Google Scholar] [CrossRef]
- Tan, L.J.; Zhu, H.; He, H.; Wu, K.H.; Li, J.; Chen, X.D.; Zhang, J.G.; Shen, H.; Tian, Q.; Krousel-Wood, M.; et al. Replication of 6 obesity genes in a meta-analysis of genome-wide association studies from diverse ancestries. PLoS ONE 2014, 9, e96149. [Google Scholar] [CrossRef] [Green Version]
- Sonestedt, E.; Roos, C.; Gullberg, B.; Ericson, U.; Wirfält, E.; Orho-Melander, M. Fat and carbohydrate intake modify the association between genetic variation in the FTO genotype and obesity. Am. J. Clin. Nutr. 2009, 90, 1418–1425. [Google Scholar] [CrossRef] [Green Version]
- Haupt, A.; Thamer, C.; Staiger, H.; Tschritter, O.; Kirchhoff, K.; Machicao, F.; Häring, H.-U.; Stefan, N.; Fritsche, A. Variation in the FTO gene influences food intake but not energy expenditure. Exp. Clin. Endocrinol. Diabetes 2009, 117, 194–197. [Google Scholar] [CrossRef]
- Wardle, J.; Carnell, S.; Haworth, C.M.; Farooqi, I.S.; O’Rahilly, S.; Plomin, R. Obesity associated genetic variation in FTO is associated with diminished satiety. J. Clin. Endocrinol. Metab. 2008, 93, 3640–3643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wardle, J.; Llewellyn, C.; Sanderson, S.; Plomin, R. The FTO gene and measured food intake in children. Int. J. Obes. 2009, 33, 42–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Speakman, J.R.; Rance, K.A.; Johnstone, A.M. Polymorphisms of the FTO gene are associated with variation in energy intake, but not energy expenditure. Obesity 2008, 16, 1961–1965. [Google Scholar] [CrossRef] [PubMed]
- Steemburgo, T.; Azevedo, M.J.; Gross, J.L.; Milagro, F.I.; Campion, J.; Martinez, J.A. The rs9939609 polymorphism in the FTO gene is associated with fat and fiber intakes in patients with type 2 diabetes. J. Nutr. Nutr. 2013, 6, 97–106. [Google Scholar]
- Mu, M.; Xu, L.F.; Hu, D.; Wu, J.; Bai, M.J. Dietary Patterns and Overweight/Obesity: A Review Article. Iran. J. Public Health 2017, 46, 869–876. [Google Scholar] [PubMed]
- Sales, N.M.; Pelegrini, P.B.; Goersch, M.C. Nutrigenomics: Definitions and advances of this new science. J. Nutr. Metab. 2014, 2014, 202759. [Google Scholar] [CrossRef] [Green Version]
- Vimaleswaran, K.S.; Bodhini, D.; Lakshmipriya, N.; Ramya, K.; Anjana, R.M.; Sudha, V.; Lovegrove, J.A.; Kinra, S.; Mohan, V.; Radha, V. Interaction between FTO gene variants and lifestyle factors on metabolic traits in an Asian Indian population. Nutr. Metab. 2016, 13, 39. [Google Scholar] [CrossRef] [Green Version]
- Kretowski, A.; Adamska, E.; Maliszewska, K.; Wawrusiewicz-Kurylonek, N.; Citko, A.; Goscik, J.; Bauer, W.; Wilk, J.; Golonko, A.; Waszczeniuk, M.; et al. The rs340874 PROX1 type 2 diabetes mellitus risk variant is associated with visceral fat accumulation and alterations in postprandial glucose and lipid metabolism. Genes Nutr. 2015, 10, 4. [Google Scholar] [CrossRef] [Green Version]
- Maliszewska, K.; Adamska-Patruno, E.; Goscik, J.; Lipinska, D.; Citko, A.; Krahel, A.; Miniewska, K.; Fiedorczuk, J.; Moroz, M.; Gorska, M.; et al. The Role of Muscle Decline in Type 2 Diabetes Development: A 5-Year Prospective Observational Cohort Study. Nutrients 2019, 11, 834. [Google Scholar] [CrossRef] [Green Version]
- Adamska-Patruno, E.; Goscik, J.; Czajkowski, P.; Maliszewska, K.; Golonko, A.; Wawrusiewicz-Kurylonek, N.; Citko, A.; Waszczeniuk, M.; Kretowski, A.; Gorska, M.; et al. The MC4R genetic variants are associated with lower visceral fat accumulation and higher postprandial relative increase in carbohydrate utilization in humans. Eur. J. Nutr. 2019, 58, 2929–2941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagstromer, M.; Oja, P.; Sjostrom, M. The International Physical Activity Questionnaire (IPAQ): A study of concurrent and construct validity. Public Health Nutr. 2006, 9, 755–762. [Google Scholar] [CrossRef]
- R Core Team (2020). R: A language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: https://www.R-project.org/ (accessed on 22 July 2020).
- Peng, S.; Zhu, Y.; Xu, F.; Ren, X.; Li, X.; Lai, M. FTO gene polymorphisms and obesity risk: A meta-analysis. BMC Med. 2011, 9, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frayling, T.M.; Timpson, N.J.; Weedon, M.N.; Zeggini, E.; Freathy, R.M.; Lindgren, C.M.; Perry, J.R.B.; Elliott, K.S.; Lango, H.; Rayner, N.W.; et al. A common variant in the FTO gene is associated with body mass index and predisposes to childhood and adult obesity. Science 2007, 316, 889–894. [Google Scholar] [CrossRef] [Green Version]
- Merritt, D.C.; Jamnik, J.; El-Sohemy, A. FTO genotype, dietary protein intake, and body weight in a multiethnic population of young adults: A cross-sectional study. Genes Nutr. 2018, 13, 4. [Google Scholar] [CrossRef]
- Wing, M.R.; Ziegler, J.; Langefeld, C.D.; Ng, M.C.Y.; Haffner, S.M.; Norris, J.M.; Goodarzi, M.O.; Bowden, D.W. Analysis of FTO gene variants with measures of obesity and glucose homeostasis in the IRAS Family Study. Hum. Genet. 2009, 125, 615–626. [Google Scholar] [CrossRef] [Green Version]
- Hosseini-Esfahani, F.; Koochakpoor, G.; Daneshpour, M.S.; Sedaghati-Khayat, B.; Mirmiran, P.; Azizi, F. Mediterranean Dietary Pattern Adherence Modify the Association between FTO Genetic Variations and Obesity Phenotypes. Nutrients 2017, 9, 1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chauhan, G.; Tabassum, R.; Mahajan, A.; Dwivedi, O.P.; Mahendran, Y.; Kaur, I.; Nigam, S.; Dubey, H.; Varma, B.; Madhu, S.V.; et al. Common variants of FTO and the risk of obesity and type 2 diabetes in Indians. J. Hum. Genet. 2011, 56, 720–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graff, M.; Gordon-Larsen, P.; Lim, U.; Fowke, J.H.; Love, S.A.; Fesinmeyer, M.; Wilkens, L.R.; Vertilus, S.; Ritchie, M.D.; Prentice, R.L.; et al. The influence of obesity-related single nucleotide polymorphisms on BMI across the life course: The PAGE study. Diabetes 2013, 62, 1763–1767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Xu, J.; Zhang, Z.; Ren, J.; Li, Y.; Wang, J.; Cao, Y.; Rong, F.; Zhao, R.; Huang, X.; et al. Association of FTO polymorphisms with obesity and metabolic parameters in Han Chinese adolescents. PLoS ONE 2014, 9, e98984. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.; Zeng, X.; Quan, L.; Zhu, J. Correlation between polymorphism of FTO gene and type 2 diabetes mellitus in Uygur people from northwest China. Int. J. Clin. Exp. Med. 2015, 8, 9744–9750. [Google Scholar]
- Bego, T.; Causevic, A.; Dujic, T.; Malenica, M.; Velija-Asimi, Z.; Prnjavorac, B.; Marc, J.; Nekvindová, J.; Palička, V.; Semiz, S. Association of FTO Gene Variant (rs8050136) with Type 2 Diabetes and Markers of Obesity, Glycaemic Control and Inflammation. J. Med. Biochem. 2019, 38, 153–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez, M.L.; West, K.L. Mechanisms by which dietary fatty acids modulate plasma lipids. J. Nutr. 2005, 135, 2075–2078. [Google Scholar] [CrossRef] [PubMed]
- Park, S.L.; Cheng, I.; Pendergrass, S.A.; Kucharska-Newton, A.M.; Lim, U.; Ambite, J.L.; Caberto, C.P.; Monroe, K.R.; Schumacher, F.; Hindorff, L.A.; et al. Association of the FTO obesity risk variant rs8050136 with percentage of energy intake from fat in multiple racial/ethnic populations: The PAGE study. Am. J. Epidemiol. 2013, 178, 780–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wing, M.R.; Ziegler, J.M.; Langefeld, C.D.; Roh, B.H.; Palmer, N.D.; Mayer-Davis, E.J.; Rewers, M.J.; Haffner, S.M.; E Wagenknecht, L.; Bowden, D.W. Analysis of FTO gene variants with obesity and glucose homeostasis measures in the multiethnic Insulin Resistance Atherosclerosis Study cohort. Int. J. Obes. 2011, 35, 1173–1182. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Bradfield, J.P.; Li, M.; Wang, K.; Zhang, H.; Kim, C.E.; Annaiah, K.; Glessner, J.T.; Thomas, K.; Garris, M.; et al. The role of obesity-associated loci identified in genome-wide association studies in the determination of pediatric BMI. Obesity 2009, 17, 2254–2257. [Google Scholar] [CrossRef]
- Dai, J.; Ying, P.; Shi, D.; Hou, H.; Sun, Y.; Xu, Z.; Chen, D.; Zhang, G.; Ni, M.; Teng, H.; et al. FTO variant is not associated with osteoarthritis in the Chinese Han population: Replication study for a genome-wide association study identified risk loci. J. Orthop. Surg. Res. 2018, 13, 65. [Google Scholar] [CrossRef]
- Cecil, J.E.; Tavendale, R.; Watt, P.; Hetherington, M.M.; Palmer, C.N. An obesity-associated FTO gene variant and increased energy intake in children. N. Engl. J. Med. 2008, 359, 2558–2566. [Google Scholar] [CrossRef]
- Lappalainen, T.; Lindstrom, J.; Paananen, J.; Eriksson, J.G.; Karhunen, L.; Tuomilehto, J.; Uusitupa, M.; Jääskeläinen, T. Association of the fat mass and obesity-associated (FTO) gene variant (rs9939609) with dietary intake in the Finnish Diabetes Prevention Study. Br. J. Nutr. 2012, 108, 1859–1865. [Google Scholar] [CrossRef] [Green Version]
- Ranzenhofer, L.; Mayer, L.E.; Davis, H.A.; Mielke-Maday, H.K.; McInerney, H.; Korn, R.; Gupta, N.; Brown, A.J.; Schebendach, J.; Tanofsky-Kraff, M.; et al. The FTO Gene and Measured Food Intake in 5- to 10-Year-Old Children Without Obesity. Obesity 2019, 27, 1023–1029. [Google Scholar] [CrossRef]
- Bauer, F.; Elbers, C.C.; Adan, R.A.; Loos, R.J.; Onland-Moret, N.C.; E Grobbee, D.; Van Vliet-Ostaptchouk, J.V.; Wijmenga, C.; Van Der Schouw, Y.T. Obesity genes identified in genome-wide association studies are associated with adiposity measures and potentially with nutrient-specific food preference. Am. J. Clin. Nutr. 2009, 90, 951–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCaffery, J.M.; Papandonatos, G.D.; Peter, I.; Huggins, G.S.; A Raynor, H.; Delahanty, L.M.; Cheskin, L.J.; Balasubramanyam, A.; E Wagenknecht, L.; Wing, R.R.; et al. Obesity susceptibility loci and dietary intake in the Look AHEAD Trial. Am. J. Clin. Nutr. 2012, 95, 1477–1486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heitmann, B.L.; Lissner, L. Dietary underreporting by obese individuals--is it specific or non-specific? BMJ 1995, 311, 986–989. [Google Scholar] [CrossRef] [PubMed] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Group Characteristics | |
|---|---|
| N | 819 |
| Age | 42.1 (14.5) |
| Female/male (%) | 52.5/47.5 |
| BMI (kg/m2) | 28.5 (6.6) |
| <25.0 | 273 (33.9%) |
| 25.0–29.9 | 278 (34.5%) |
| ≥30.0 | 255 (31.6%) |
| Total body fat content (kg) | 27.1 (13.8) |
| Total body fat content (%) | 31.4 (9.6) |
| Waist circumference (cm) | 96.2 (17.2) |
| Hip circumference (cm) | 103.3 (12.7) |
| WHR | 0.928 (0.088) |
| Visceral fat (cm3) | 108.4 (80.6) |
| Visceral fat (%) | 37.1 (12.1) |
| Subcutaneous fat (cm3) | 167.9 (81.7) |
| Subcutaneous fat (%) | 62.8 (12.3) |
| Visceral/subcutaneous fat ratio | 0.669 (0.443) |
| Total cholesterol | 195.4 (46.1) |
| HDL | 59.7 (14.9) |
| LDL | 112.0 (40.0) |
| TG | 118.8 (95.1) |
| Fasting blood glucose level (mg/dL) | 98.8 (23.9) |
| History of prediabetes or diabetes | |
| Yes | 411 (50.2%) |
| No | 408 (49.8%) |
| Dietary assessment (n) | 490 |
| Daily energy intake (kcal) | 1792.5 (697.4) |
| Daily energy from protein (%) | 18.9 (4.8) |
| Daily energy from fat (%) | 31.2 (7.5) |
| Daily energy from carbohydrates (%) | 47.6 (8.6) |
| Daily physical activity level | |
| Low | 60 (7.3%) |
| Moderate | 173 (21.1%) |
| High | 586 (71.6%) |
| rs3751812 | G/G | G/T | T/T | p-Value * |
|---|---|---|---|---|
| N | 211 | 420 | 181 | |
| Genotype frequency | 0.26 | 0.52 | 0.22 | >0.05 |
| Age | 40.5 (14.2) | 41.2 (14.7) | 39.5 (14.3) | 0.33 |
| Female (%) | 53.8% (0.49) | 53.8% (0.50) | 47.8% (0.50) | 0.36 |
| BMI (kg/m2) | 27.6 (6.0) | 28.7 (6.8) | 28.9 (6.8) | 0.060 |
| <25.0 | 81 (38.8%) | 136 (32.9%) | 53 (29.9%) | 0.410 |
| 25.0–29.9 | 69 (33.0%) | 141 (34.1%) | 66 (37.3%) | |
| ≥30.0 | 59 (28.2%) | 136 (32.9%) | 58 (32.8%) | |
| Total body fat content (kg) | 25.3 (12.4) | 27.6 (13.8) | 28.2 (15.2) | 0.080 |
| Total body fat content (%) | 30.6 (9.1) | 31.8 (9.6) | 31.6 (10.3) | 0.377 |
| Waist circumference (cm) | 94.3 (17.5) | 96.7 (17.2) | 97.5 (16.7) | 0.054 |
| Hip circumference (cm) | 101.3 (12.4) | 104.2 (13.0) | 103.8 (12.5) | 0.008 |
| WHR | 0.927 (0.091) | 0.925 (0.088) | 0.937 (0.085) | 0.327 |
| Visceral fat (cm3) | 103.0 (81.0) | 110.0 (79.9) | 112.3 (83.0) | 0.379 |
| Visceral fat (%) | 36.4 (11.8) | 37.5 (12.4) | 37.2 (11.7) | 0.587 |
| Subcutaneous fat (cm3) | 163.5 (83.1) | 167.2 (80.5) | 175.0 (82.7) | 0.401 |
| Subcutaneous fat (%) | 63.7 (11.7) | 62.3 (12.9) | 62.8 (11.7) | 0.557 |
| Visceral/subcutaneous fat ratio | 0.642 (0.406) | 0.687 (0.475) | 0.665 (0.413) | 0.554 |
| Total cholesterol | 202.7 (56.0) | 191.7 (41.3) | 194.0 (43.2) | 0.070 |
| HDL | 60.7 (14.1) | 59.8 (15.6) | 59.5 (14.5) | 0.662 |
| LDL | 117.3 (43.3) | 109.4 (37.8) | 111.2 (41.8) | 0.095 |
| TG | 123.8 (143.9) | 111.9 (69.7) | 116.3 (61.9) | 0.289 |
| Blood glucose level during OGTT (mg/dL) | ||||
| 0 min | 96.8 (24.1) | 95.6 (18.3) | 97.1 (20.8) | 0.914 |
| 30 min | 147.0 (44.3) | 145.0 (31.6) | 150.1 (35.6) | 0.312 |
| 60 min | 132.3 (56.0) | 129.5 (46.3) | 134.2 (46.3) | 0.380 |
| 120 min | 100.7 (46.1) | 99.1 (32.1) | 98.8 (31.0) | 0.621 |
| History of prediabetes or diabetes | ||||
| Yes | 103 (48.8%) | 209 (49.8%) | 95 (52.5%) | 0.751 |
| No | 108 (51.2%) | 211 (50.2%) | 86 (47.5%) | |
| Dietary assessment (n) | 126 | 259 | 101 | |
| Daily energy intake (kcal) | 1807.2 (732.3) | 1766.9 (676.0) | 1837.4 (713.4) | 0.849 |
| Daily energy from protein (%) | 18.7 (4.4) | 19.0 (4.9) | 19.1 (4.9) | 0.901 |
| Daily energy from fat (%) | 31.2 (7.2) | 30.9 (7.5) | 31.9 (7.8) | 0.568 |
| Daily energy from carbohydrates (%) | 47.6 (7.7) | 47.8 (9.1) | 46.8 (8.5) | 0.662 |
| Daily physical activity level | ||||
| Low | 16 (7.6%) | 25 (6.0%) | 18 (9.9%) | 0.302 |
| Moderate | 50 (23.7%) | 83 (19.8%) | 40 (22.1%) | |
| High | 145 (68.7%) | 312 (74.3%) | 123 (68.0%) | |
| rs8050136 | C/C | A/C | A/A | p-Value * |
|---|---|---|---|---|
| N | 209 | 424 | 182 | |
| Genotype frequency | 0.26 | 0.52 | 0.22 | >0.05 |
| Age | 40.2 (14.1) | 41.3 (14.8) | 39.4 (14.3) | 0.24 |
| Females (%) | 54.3% (0.49) | 53.4% (0.49) | 47.7% (0.50) | 0.36 |
| BMI (kg/m2) | 27.6 (6.1) | 28.7 (6.8) | 28.9 (6.8) | 0.063 |
| <25.0 | 80 (38.6%) | 138 (33.2%) | 54 (30.2%) | 0.422 |
| 25.0–29.9 | 69 (33.3%) | 140 (33.7%) | 67 (37.4%) | |
| ≥30.0 | 58 (28.0%) | 138 (33.2%) | 58 (32.4%) | |
| Total body fat content (kg) | 25.3 (12.4) | 27.5 (13.8) | 28.2 (15.2) | 0.095 |
| Total body fat content (%) | 30.6 (9.1) | 31.8 (9.6) | 31.6 (10.4) | 0.465 |
| Waist circumference (cm) | 94.2 (17.6) | 96.7 (17.2) | 97.4 (16.6) | 0.053 |
| Hip circumference (cm) | 101.2 (12.5) | 104.1 (13.0) | 103.8 (12.4) | 0.008 |
| WHR | 0.927 (0.092) | 0.925 (0.088) | 0.936 (0.085) | 0.382 |
| Visceral fat (cm3) | 103.3 (81.5) | 110.2 (80.0) | 111.8 (82.5) | 0.381 |
| Visceral fat (%) | 36.4 (11.7) | 37.6 (12.5) | 37.1 (11.6) | 0.570 |
| Subcutaneous fat (cm3) | 163.7 (83.5) | 166.9 (80.8) | 175.3 (83.1) | 0.405 |
| Subcutaneous fat (%) | 63.7 (11.6) | 62.2 (13.0) | 62.9 (11.6) | 0.540 |
| Visceral/subcutaneous fat ratio | 0.641 (0.404) | 0.690 (0.477) | 0.662 (0.410) | 0.536 |
| Total cholesterol | 201.9 (56.1) | 192.1 (41.4) | 193.7 (43.1) | 0.153 |
| HDL | 60.8 (14.0) | 59.6 (15.7) | 59.7 (14.4) | 0.422 |
| LDL | 116.3 (43.3) | 109.9 (37.9) | 111.1 (41.8) | 0.189 |
| TG | 124.1 (144.3) | 113.2 (71.1) | 115.1 (61.3) | 0.491 |
| Blood glucose level during OGTT (mg/dL) | ||||
| 0 min | 96.8 (24.2) | 95.8 (18.5) | 96.8 (20.4) | 0.922 |
| 30 min | 146.6 (44.5) | 145.2 (31.7) | 150.1 (35.4) | 0.263 |
| 60 min | 131.7 (56.4) | 129.9 (46.1) | 133.9 (46.1) | 0.413 |
| 120 min | 100.2 (46.3) | 99.3 (32.2) | 98.6 (30.9) | 0.493 |
| History of prediabetes or diabetes | ||||
| Yes | 100 (47.8%) | 213 (50.2%) | 96 (52.7%) | 0.628 |
| No | 109 (52.2%) | 211 (49.8%) | 86 (47.3%) | |
| Dietary assessment (n) | 103 | 264 | 123 | |
| Daily energy intake (kcal) | 1820.5 (734.9) | 1759.6 (673.3) | 1853.9 (716.3) | 0.645 |
| Daily energy from protein (%) | 18.7 (4.5) | 19.0 (4.9) | 19.0 (4.9) | 0.855 |
| Daily energy from fat (%) | 31.2 (7.2) | 30.9 (7.5) | 31.9 (7.8) | 0.506 |
| Daily energy from carbohydrates (%) | 47.6 (7.7) | 47.9 (9.1) | 46.8 (8.4) | 0.572 |
| Daily physical activity level | ||||
| Low | 16 (7.7%) | 25 (5.9%) | 19 (10.4%) | 0.179 |
| Moderate | 50 (23.9%) | 83 (19.6%) | 40 (22.0%) | |
| High | 143 (68.4%) | 316 (74.5%) | 123 (67.6%) | |
| rs8044769 | C/C | C/T | T/T | p-Value * |
|---|---|---|---|---|
| N | 270 | 406 | 138 | |
| Genotype frequency | 0.33 | 0.50 | 0.17 | >0.05 |
| Age | 40.4 (14.8) | 41.2 (14.6) | 39.6 (13.7) | 0.54 |
| Females (%) | 51.8% (0.50) | 51.9% (0.50) | 56.2% (0.49) | 0.65 |
| BMI (kg/m2) | 28.5 (6.8) | 28.7 (6.7) | 27.8 (6.1) | 0.534 |
| <25.0 | 88 (33.2%) | 131 (32.8%) | 51 (37.5%) | 0.862 |
| 25.0–29.9 (kg/m2) | 92 (34.7%) | 143 (35.8%) | 43 (31.6%) | |
| ≥30.0 (kg/m2) | 85 (32.1%) | 126 (31.5%) | 42 (30.9%) | |
| Total body fat content (kg) | 27.7 (15.0) | 27.2 (13.6) | 25.8 (11.9) | 0.676 |
| Total body fat content (%) | 31.7 (10.1) | 31.4 (9.6) | 31.1 (9.0) | 0.893 |
| Waist circumference (cm) | 96.2 (17.2) | 96.8 (17.3) | 94.6 (16.8) | 0.429 |
| Hip circumference (cm) | 103.3 (12.6) | 104.0 (13.0) | 101.5 (12.2) | 0.189 |
| WHR | 0.928 (0.089) | 0.927 (0.088) | 0.929 (0.089) | 0.980 |
| Visceral fat (cm3) | 109.2 (78.3) | 110.3 (83.7) | 99.8 (72.5) | 0.648 |
| Visceral fat (%) | 37.7 (11.8) | 36.9 (12.3) | 36.4 (12.0) | 0.617 |
| Subcutaneous fat (cm3) | 168.6 (83.3) | 169.7 (82.4) | 160.9 (74.2) | 0.773 |
| Subcutaneous fat (%) | 62.3 (11.8) | 63.0 (12.8) | 63.7 (11.8) | 0.598 |
| Visceral/subcutaneous fat ratio | 0.689 (0.492) | 0.662 (0.421) | 0.641 (0.408) | 0.590 |
| Total cholesterol | 193.8 (40.1) | 191.7 (42.7) | 206.7 (62.7) | 0.029 |
| LDL | 111.0 (38.6) | 109.6 (38.7) | 119.8 (46.5) | 0.058 |
| HDL | 60.7 (14.7) | 59.2 (15.5) | 60.4 (13.8) | 0.176 |
| TG | 110.5 (58.7) | 114.6 (73.4) | 132.6 (170.6) | 0.689 |
| Blood glucose level during OGTT (mg/dL) | ||||
| 0 min | 96.6 (20.0) | 96.1 (21.0) | 96.0 (20.1) | 0.910 |
| 30 min | 149.6 (35.1) | 143.7 (32.0) | 148.8 (47.8) | 0.229 |
| 60 min | 133.3 (45.4) | 128.7 (47.3) | 133.5 (59.3) | 0.303 |
| 120 min | 98.6 (31.8) | 99.3 (30.7) | 100.6 (53.0) | 0.147 |
| History of prediabetes or diabetes | ||||
| Yes | 143 (53.0%) | 201 (49.5%) | 64 (46.4%) | 0.420 |
| No | 127 (47.0%) | 205 (50.5%) | 74 (53.6%) | |
| Dietary assessment (n) | 157 | 248 | 83 | |
| Daily energy intake (kcal) | 1775.7 (646.2) | 1792.7 (735.8) | 1816.0 (686.3) | 0.791 |
| % of daily energy from protein | 18.9 (4.8) | 19.2 (5.0) | 18.4 (4.1) | 0.608 |
| % of daily energy from fat | 32.5 (7.3) | 30.1 (7.5) | 31.9 (7.4) | 0.005 |
| % of daily energy from carbohydrates | 46.5 (8.5) | 48.3 (8.9) | 47.1 (8.1) | 0.164 |
| Daily physical activity level | ||||
| Low | 23 (8.5%) | 26 (6.4%) | 10 (7.2%) | 0.060 |
| Moderate | 55 (20.4%) | 77 (19.0%) | 41 (29.7%) | |
| High | 192 (71.1%) | 303 (74.6%) | 87 (63.0%) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Czajkowski, P.; Adamska-Patruno, E.; Bauer, W.; Fiedorczuk, J.; Krasowska, U.; Moroz, M.; Gorska, M.; Kretowski, A. The Impact of FTO Genetic Variants on Obesity and Its Metabolic Consequences is Dependent on Daily Macronutrient Intake. Nutrients 2020, 12, 3255. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12113255
Czajkowski P, Adamska-Patruno E, Bauer W, Fiedorczuk J, Krasowska U, Moroz M, Gorska M, Kretowski A. The Impact of FTO Genetic Variants on Obesity and Its Metabolic Consequences is Dependent on Daily Macronutrient Intake. Nutrients. 2020; 12(11):3255. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12113255
Chicago/Turabian StyleCzajkowski, Przemyslaw, Edyta Adamska-Patruno, Witold Bauer, Joanna Fiedorczuk, Urszula Krasowska, Monika Moroz, Maria Gorska, and Adam Kretowski. 2020. "The Impact of FTO Genetic Variants on Obesity and Its Metabolic Consequences is Dependent on Daily Macronutrient Intake" Nutrients 12, no. 11: 3255. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12113255