
Is a Four-Week Ketogenic Diet an Effective Nutritional Strategy in CrossFit-Trained Female and Male Athletes?

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
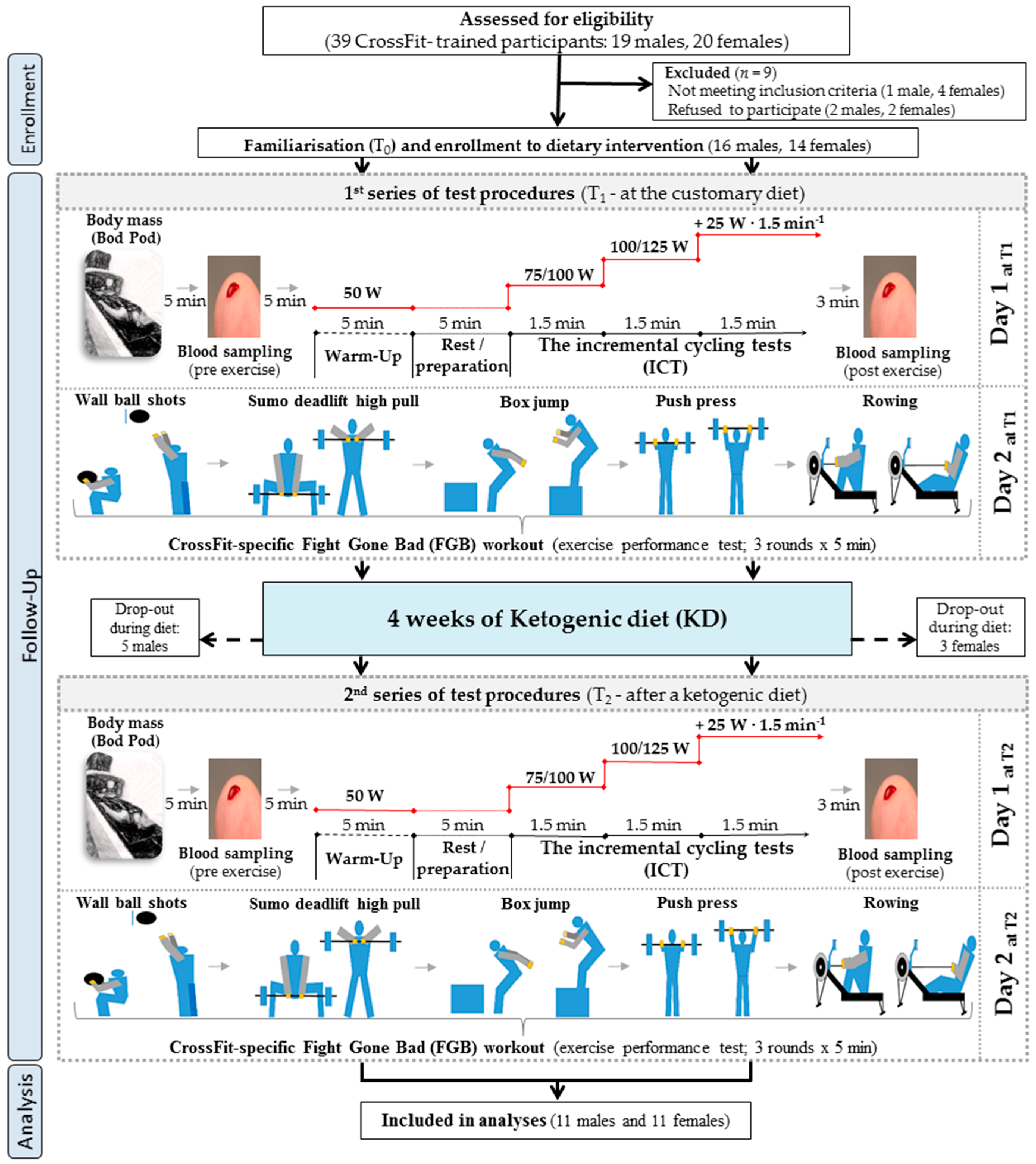
2. Materials and Methods
2.1. Participants
2.2. Study Design, Protocol, and Visits
2.3. Exercise Tests
2.3.1. ICT
2.3.2. CrossFit-Specific Physical Performance Test
2.4. Blood Collection and Sample Analysis
2.4.1. Blood Sampling
2.4.2. Blood Sample Analysis
2.5. Statistical Analysis
3. Results
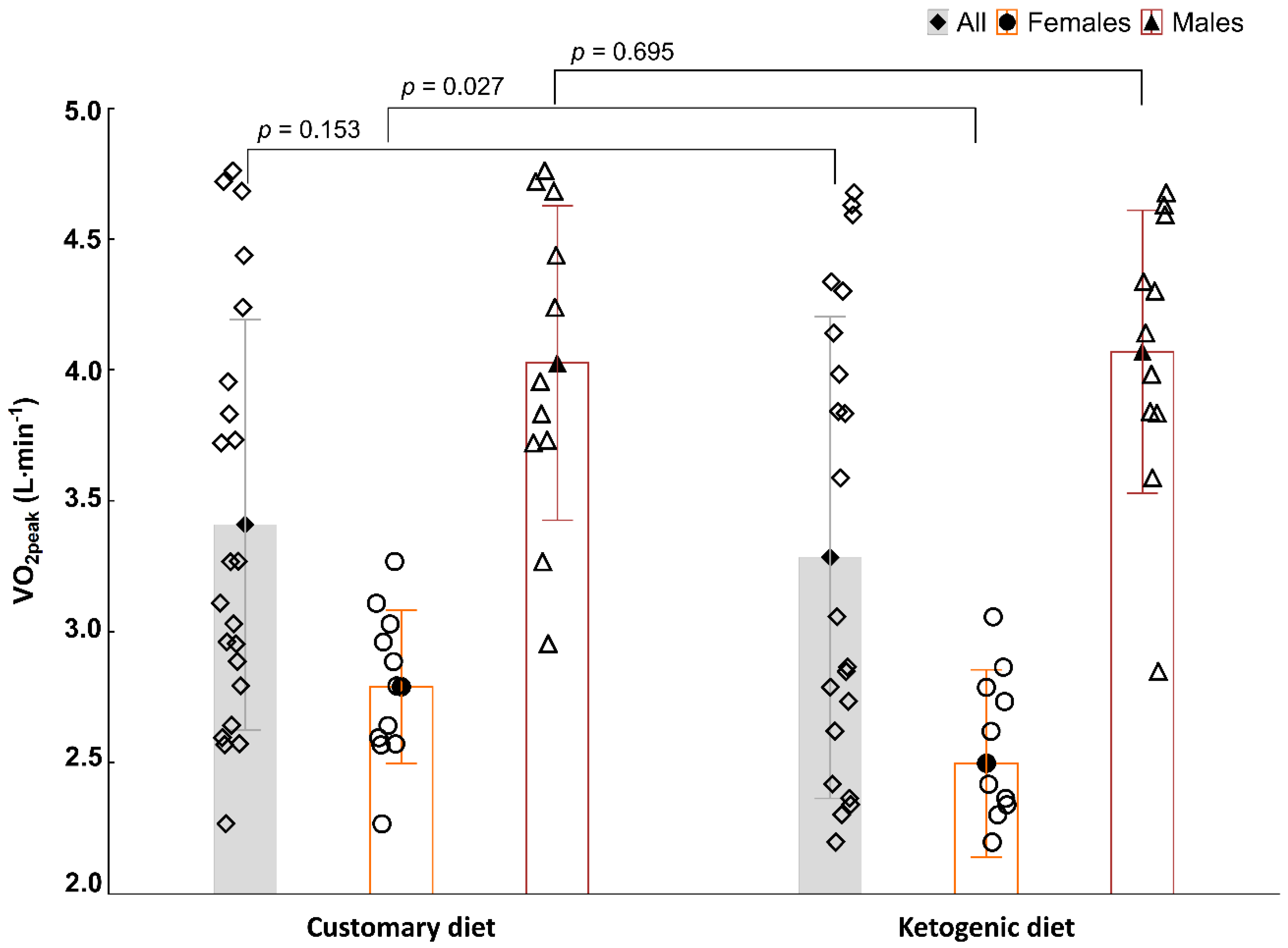
3.1. Aerobic Capacity in the ICT
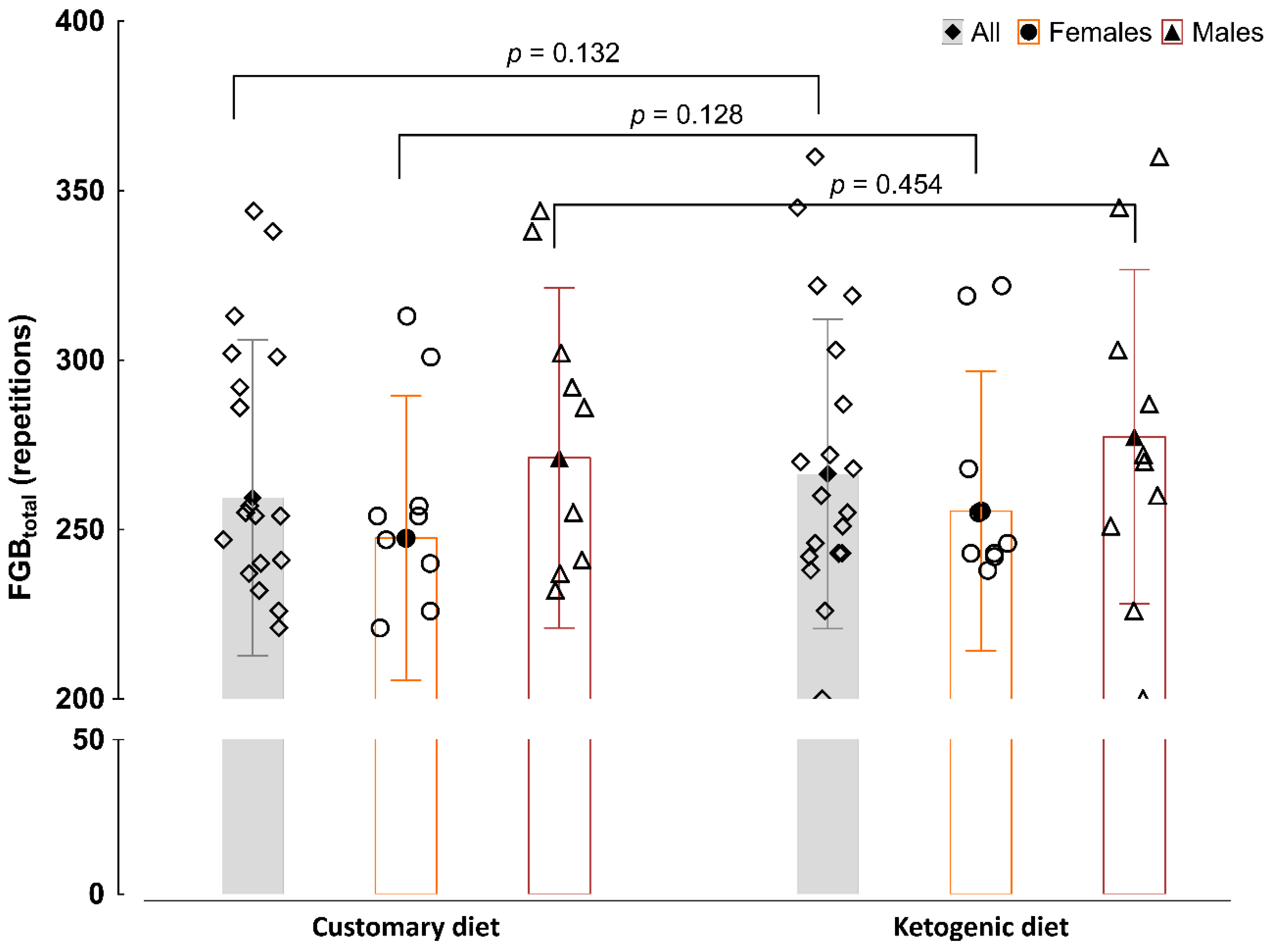
3.2. Discipline-Specific CrossFit Performance in the FGB Test
3.3. Blood Marker Analysis
3.3.1. Blood Acid–Base Balance
3.3.2. Blood Biochemical Parameters
3.3.3. Blood Haematological Markers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burke, L.M.; Kiens, B.; Ivy, J.L. Carbohydrates and fat for training and recovery. J. Sports Sci. 2004, 22, 15–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhillon, K.K.; Gupta, S. Biochemistry, Ketogenesis. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Burke, L.M.; Hawley, J.A.; Wong, S.H.S.; Jeukendrup, A.E. Carbohydrates for training and competition. J. Sports Sci. 2011, 29, S17–S27. [Google Scholar] [CrossRef] [PubMed]
- Kerksick, C.M.; Wilborn, C.D.; Roberts, M.D.; Smith-Ryan, A.; Kleiner, S.M.; Jäger, R.; Collins, R.; Cooke, M.; Davis, J.N.; Galvan, E.; et al. ISSN Exercise & sports nutrition review update: Research & recommendations. J. Int. Soc. Sports Nutr. 2018, 15, 38. [Google Scholar]
- Thomas, D.T.; Erdman, K.A.; Burke, L.M. American College of Sports Medicine joint position statement. Nutrition and Athletic Performance. Med. Sci. Sports Exerc. 2016, 48, 543–568. [Google Scholar]
- Spriet, L.L. New insights into the interaction of carbohydrate and fat metabolism during exercise. Sports Med. 2014, 44, 87–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moscatelli, F.; Valenzano, A.; Polito, R.; Francesco, S.; Montana, A.; Salerno, M.; Messina, A.; Monda, M.; Cibelli, G.; Monda, V.; et al. Ketogenic diet and sport performance. Sport Mont 2020, 18, 91–94. [Google Scholar]
- Durkalec-Michalski, K.; Nowaczyk, P.M.; Siedzik, K. Effect of a four-week ketogenic diet on exercise metabolism in crossfit-trained athletes. J. Int. Soc. Sports Nutr. 2019, 16, 16. [Google Scholar] [CrossRef] [Green Version]
- Harvey, K.L.; Holcomb, L.E.; Kolwicz, S.C. Ketogenic diets and exercise performance. Nutrients 2019, 11, 2296. [Google Scholar] [CrossRef] [Green Version]
- Gosmanov, A.R.; Gosmanova, E.O.; Dillard-Cannon, E. Management of adult diabetic ketoacidosis. Diabetes Metab. Syndr. Obes. 2014, 7, 255–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paoli, A. Ketogenic diet for obesity: Friend or foe? Int. J. Environ. Res. Public Health 2014, 11, 2092–2107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foster, G.D.; Wyatt, H.R.; Hill, J.O.; Makris, A.P.; Rosenbaum, D.L.; Brill, C.; Stein, R.I.; Mohammed, B.S.; Miller, B.; Rader, D.J.; et al. Weight and metabolic outcomes after 2 years on a low-carbohydrate versus low-fat diet. Ann. Intern. Med. 2010, 153, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Bueno, N.B.; de Melo, I.S.V.; de Oliveira, S.L.; da Rocha Ataide, T. Very-low-carbohydrate ketogenic diet v. low-fat diet for long-term weight loss: A meta-analysis of randomised controlled trials. Br. J. Nutr. 2013, 110, 1178–1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brouns, F. Overweight and diabetes prevention: Is a low-carbohydrate–high-fat diet recommendable? Eur. J. Nutr. 2018, 57, 1301–1312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, V.W.; Leung, K.; Hsu, A.; Luk, B.; Lai, J.; Shen, S.Y.; Minchinton, A.I.; Waterhouse, D.; Bally, M.B.; Lin, W.; et al. A low carbohydrate, high protein diet slows tumor growth and prevents cancer initiation. Cancer Res. 2011, 71, 4484–4493. [Google Scholar] [CrossRef] [Green Version]
- Otto, C.; Kaemmerer, U.; Illert, B.; Muehling, B.; Pfetzer, N.; Wittig, R.; Voelker, H.U.; Thiede, A.; Coy, J.F. Growth of human gastric cancer cells in nude mice is delayed by a ketogenic diet supplemented with omega-3 fatty acids and medium-chain triglycerides. BMC Cancer 2008, 8, 122. [Google Scholar] [CrossRef] [Green Version]
- Shaw, D.M.; Merien, F.; Braakhuis, A.; Maunder, E.D.; Dulson, D.K. Effect of a ketogenic diet on submaximal exercise capacity and efficiency in runners. Med. Sci. Sports Exerc. 2019, 51, 2135–2146. [Google Scholar] [CrossRef] [PubMed]
- Volek, J.S.; Noakes, T.; Phinney, S.D. Rethinking fat as a fuel for endurance exercise. Eur. J. Sport Sci. 2015, 15, 13–20. [Google Scholar] [CrossRef]
- Evans, M.; Cogan, K.E.; Egan, B. Metabolism of ketone bodies during exercise and training: Physiological basis for exogenous supplementation. J. Physiol. 2017, 595, 2857–2871. [Google Scholar] [CrossRef] [Green Version]
- Pinckaers, P.J.M.; Churchward-Venne, T.A.; Bailey, D.; Van Loon, L.J.C. Ketone Bodies and exercise performance: The next magic bullet or merely hype? Sports Med. 2017, 47, 383–391. [Google Scholar] [CrossRef] [Green Version]
- Hasan-Olive, M.M.; Lauritzen, K.H.; Ali, M.; Rasmussen, L.J.; Storm-Mathisen, J.; Bergersen, L.H. A ketogenic diet improves mitochondrial biogenesis and bioenergetics via the PGC1α-SIRT3-UCP2 axis. Neurochem. Res. 2019, 44, 22–37. [Google Scholar] [CrossRef] [PubMed]
- Hearris, M.A.; Hammond, K.M.; Fell, J.M.; Morton, J.P. Regulation of muscle glycogen metabolism during exercise: Implications for endurance performance and training adaptations. Nutrients 2018, 10, 298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egan, B.; Zierath, J.R. Exercise metabolism and the molecular regulation of skeletal muscle adaptation. Cell Metab. 2013, 17, 162–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perry, C.G.R.; Lally, J.; Holloway, G.P.; Heigenhauser, G.J.F.; Bonen, A.; Spriet, L.L. Repeated transient mRNA bursts precede increases in transcriptional and mitochondrial proteins during training in human skeletal muscle. J. Physiol. 2010, 588, 4795–4810. [Google Scholar] [CrossRef] [PubMed]
- Pilegaard, H.; Keller, C.; Steensberg, A.; Helge, J.W.; Pedersen, B.K.; Saltin, B.; Neufer, P.D. Influence of pre-exercise muscle glycogen content on exercise-induced transcriptional regulation of metabolic genes. J. Physiol. 2002, 541, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Durkalec-Michalski, K.; Zawieja, E.E.; Zawieja, B.E.; Podgórski, T.; Jurkowska, D.; Jeszka, J. Influence of low versus moderate glycemic index of diet on substrate oxidation and energy expenditure during incremental exercise in endurance athletes: A randomized counterbalanced cross-over trial. Int. J. Food Sci. Nutr. 2018, 69, 741–752. [Google Scholar] [CrossRef]
- Nakao, R.; Abe, T.; Yamamoto, S.; Oishi, K. Ketogenic diet induces skeletal muscle atrophy via reducing muscle protein synthesis and possibly activating proteolysis in mice. Sci. Rep. 2019, 9, 19652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burke, L.M.; Sharma, A.P.; Heikura, I.A.; Forbes, S.F.; Holloway, M.; McKay, A.K.A.; Bone, J.L.; Leckey, J.J.; Welvaert, M.; Ross, M.L. Crisis of confidence averted: Impairment of exercise economy and performance in elite race walkers by ketogenic low carbohydrate, high fat (LCHF) diet is reproducible. PLoS ONE 2020, 15, e0234027. [Google Scholar]
- Burke, L.M. Ketogenic Low-CHO, High-fat diet: The future of elite endurance sport? J. Physiol. 2020, 599, 819–843. [Google Scholar] [CrossRef] [PubMed]
- Paoli, A.; Bianco, A.; Grimaldi, K.A. The ketogenic diet and sport: A possible marriage? Exerc. Sport Sci. Rev. 2015, 43, 153–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burke, L.M. Re-examining high-fat diets for sports performance: Did we call the ‘nail in the coffin’ too soon? Sports Med. 2015, 45, 33–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burke, L.M.; Ross, M.L.; Garvican-Lewis, L.A.; Welvaert, M.; Heikura, I.A.; Forbes, S.G.; Mirtschin, J.G.; Cato, L.E.; Strobel, N.; Sharma, A.P.; et al. Low carbohydrate, high fat diet impairs exercise economy and negates the performance benefit from intensified training in elite race walkers. J. Physiol. 2017, 595, 2785–2807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McSwiney, F.T.; Wardrop, B.; Hyde, P.N.; Lafountain, R.A.; Volek, J.S.; Doyle, L. Keto-adaptation enhances exercise performance and body composition responses to training in endurance athletes. Metabolism 2018, 81, 25–34. [Google Scholar] [CrossRef]
- Wilson, J.M.; Lowery, R.P.; Roberts, M.D.; Sharp, M.H.; Joy, J.M.; Shields, K.A.; Partl, J.; Volek, J.S.; D’Agostino, D. The effects of ketogenic dieting on body composition, strength, power, and hormonal profiles in resistance training males. J. Strength Cond. Res. 2020, 34, 3463–3474. [Google Scholar] [CrossRef]
- Paoli, A.; Grimaldi, K.; D’Agostino, D.; Cenci, L.; Moro, T.; Bianco, A.; Palma, A. Ketogenic diet does not affect strength performance in elite artistic gymnasts. J. Int. Soc. Sports Nutr. 2012, 9, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaspar, M.B.; Austin, K.; Huecker, M.; Sarav, M. Ketogenic diet: From the historical records to use in elite athletes. Curr. Nutr. Rep. 2019, 8, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Durkalec-Michalski, K.; Zawieja, E.E.; Zawieja, B.E.; Jurkowska, D.; Buchowski, M.S.; Jeszka, J. Effects of low versus moderate glycemic index diets on aerobic capacity in endurance runners: Three-week randomized controlled crossover trial. Nutrients 2018, 10, 370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durkalec-Michalski, K.; Woźniewicz, M.; Bajerska, J.; Jeszka, J. Comparison of accuracy of various non-calorimetric methods measuring energy expenditure at different intensities. Hum. Mov. 2013, 14, 161–167. [Google Scholar] [CrossRef]
- Beaver, W.L.; Wasserman, K.; Whipp, B.J. A new method for detecting anaerobic threshold by gas exchange. J. Appl. Physiol. 1986, 60, 2020–2027. [Google Scholar] [CrossRef]
- Durkalec-Michalski, K.; Podgórski, T.; Sokołowski, M.; Jeszka, J. Relationship between body composition indicators and physical capacity of the combat sports athletes. Arch. Budo 2016, 12, 247–256. [Google Scholar]
- Durkalec-Michalski, K.; Zawieja, E.E.; Podgórski, T.; Łoniewski, I.; Zawieja, B.E.; Warzybok, M.; Jeszka, J. The effect of chronic progressive-dose sodium bicarbonate ingestion on crossfit-like performance: A double-blind, randomized cross-over trial. PLoS ONE 2018, 13, e0197480. [Google Scholar] [CrossRef]
- Podgorski, T.; Bartkowiak, U.; Pawlak, M. Comparison of hematological parameters of venous and capillary blood in athletes. Trends Sport Sci. 2014, 21, 39–45. [Google Scholar]
- Zajac, A.; Poprzecki, S.; Maszczyk, A.; Czuba, M.; Michalczyk, M.; Zydek, G. The effects of a ketogenic diet on exercise metabolism and physical performance in off-road cyclists. Nutrients 2014, 6, 2493–2508. [Google Scholar] [CrossRef] [PubMed]
- Zinn, C.; Wood, M.; Williden, M.; Chatterton, S.; Maunder, E. Ketogenic diet benefits body composition and well-being but not performance in a pilot case study of new zealand endurance athletes. J. Int. Soc. Sports Nutr. 2017, 14, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klement, R.J.; Frobel, T.; Albers, T.; Fikenzer, S.; Prinzhausen, J.; Kämmerer, U. A pilot case study on the impact of a self-prescribed ketogenic diet on biochemical parameters and running performance in healthy and physically active individuals. Nutr. Med. 2013, 1, 10. [Google Scholar]
- Kephart, W.C.; Pledge, C.D.; Roberson, P.A.; Mumford, P.W.; Romero, M.A.; Mobley, C.B.; Martin, J.S.; Young, K.C.; Lowery, R.P.; Wilson, J.M.; et al. The three-month effects of a ketogenic diet on body composition, blood parameters, and performance metrics in crossfit trainees: A pilot study. Sports 2018, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Phinney, S.D.; Bistrian, B.R.; Wolfe, R.R.; Blackburn, G.L. The human metabolic response to chronic ketosis without caloric restriction: Physical and biochemical adaptation. Metabolism 1983, 32, 757–768. [Google Scholar] [CrossRef]
- Gregory, R.M. A low-carbohydrate ketogenic diet combined with 6-weeks of crossfit training improves body composition and performance. Int. J. Sports Exerc. Med. 2017, 3, 054. [Google Scholar] [CrossRef]
- Carr, A.J.; Sharma, A.P.; Ross, M.L.; Welvaert, M.; Slater, G.J.; Burke, L.M. Chronic ketogenic low carbohydrate high fat diet has minimal effects on acid-base status in elite athletes. Nutrients 2018, 10, 236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langfort, J.; Pilis, W.; Zarzeczny, R.; Nazar, K.; Kaciuba-Uściłko, H. Effect of low-carbohydrate-ketogenic diet on metabolic and hormonal responses to graded exercise in men. J. Physiol. Pharm. 1996, 47, 361–371. [Google Scholar]
- Greenhaff, P.L.; Gleeson, M.; Maughan, R.J. The effects of diet on muscle ph and metabolism during high intensity exercise. Eur. J. Appl. Physiol. Occup. Physiol. 1988, 57, 531–539. [Google Scholar] [CrossRef]
- Helge, J.W.; Richter, E.A.; Kiens, B. Interaction of training and diet on metabolism and endurance during exercise in man. J. Physiol. 1996, 492, 293–306. [Google Scholar] [CrossRef] [PubMed]
- Volek, J.S.; Freidenreich, D.J.; Saenz, C.; Kunces, L.J.; Creighton, B.C.; Bartley, J.M.; Davitt, P.M.; Munoz, C.X.; Anderson, J.M.; Maresh, C.M.; et al. Metabolic characteristics of keto-adapted ultra-endurance runners. Metabolism 2016, 65, 100–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phinney, S.D.; Bistrian, B.R.; Evans, W.J.; Gervino, E.; Blackburn, G.L. The human metabolic response to chronic ketosis without caloric restriction: Preservation of submaximal exercise capability with reduced carbohydrate oxidation. Metabolism 1983, 32, 769–776. [Google Scholar] [CrossRef]
- Webster, C.C.; Noakes, T.D.; Chacko, S.K.; Swart, J.; Kohn, T.A.; Smith, J.A.H. Gluconeogenesis during endurance exercise in cyclists habituated to a long-term low carbohydrate high-fat diet. J. Physiol. 2016, 594, 4389–4405. [Google Scholar] [CrossRef] [PubMed]
- Helge, J.W. Long-term fat diet adaptation effects on performance, training capacity, and fat utilization. Med. Sci. Sports Exerc. 2002, 34, 1499–1504. [Google Scholar] [CrossRef]
- Urbain, P.; Strom, L.; Morawski, L.; Wehrle, A.; Deibert, P.; Bertz, H. Impact of a 6-week non-energy-restricted ketogenic diet on physical fitness, body composition and biochemical parameters in healthy adults. Nutr. Metab. 2017, 14, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greene, D.A.; Varley, B.J.; Hartwig, T.B.; Chapman, P.; Rigney, M. A low-carbohydrate ketogenic diet reduces body mass without compromising performance in powerlifting and olympic weightlifting athletes. J. Strength Cond. Res. 2018, 32, 3373–3382. [Google Scholar] [CrossRef] [PubMed]
- Rhyu, H.; Cho, S.-Y.; Roh, H.-T. The effects of ketogenic diet on oxidative stress and antioxidative capacity markers of taekwondo athletes. J. Exerc. Rehabil. 2014, 10, 362–366. [Google Scholar] [CrossRef]
- Banfi, G.; Colombini, A.; Lombardi, G.; Lubkowska, A. Metabolic markers in sports medicine. Adv. Clin. Chem. 2012, 56, 1–54. [Google Scholar]
- Hamoud, A.; Weaver, L.; Stec, D.E.; Hinds, T.D. Bilirubin in the liver to gut signaling axis. Trends Endocrinol. Metab. 2018, 29, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Kwak, M.-S.; Kim, D.; Chung, G.E.; Kang, S.J.; Park, M.J.; Kim, Y.J.; Yoon, J.-H.; Lee, H.-S. Serum bilirubin levels are inversely associated with nonalcoholic fatty liver disease. Clin. Mol. Hepatol. 2012, 18, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Wooten, J.S.; Biggerstaff, K.D.; Ben-Ezra, V. Responses of LDL and HDL Particle size and distribution to omega-3 fatty acid supplementation and aerobic exercise. J. Appl. Physiol. 2009, 107, 794–800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarosz, M. Normy Żywienia dla Populacji Polski; Instytut Żywności i Żywienia: Warszawa, Poland, 2017. [Google Scholar]
- Pagana, K.; Pagana, T.; Pagana, T. Mosby’s Diagnostic and Laboratory Test Reference, 14th ed.; Elsevier: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Kim, I.; Yetley, E.A.; Calvo, M.S. Variations in iron-status measures during the menstrual cycle. Am. J. Clin. Nutr. 1993, 58, 705–709. [Google Scholar] [CrossRef] [PubMed]
- McNulty, K.L.; Elliott-Sale, K.J.; Dolan, E.; Swinton, P.A.; Ansdell, P.; Goodall, S.; Thomas, K.; Hicks, K.M. The effects of menstrual cycle phase on exercise performance in eumenorrheic women: A systematic review and meta-analysis. Sports Med. 2020, 50, 1813–1827. [Google Scholar] [CrossRef] [PubMed]
- Hagstrom, A.D.; Yuwono, N.; Warton, K.; Ford, C.E. Sex bias in cohorts included in sports medicine research. Sports Med. 2021, in press. [Google Scholar] [CrossRef] [PubMed]
- Durkalec–Michalski, K.; Zawieja, E.E.; Zawieja, B.E.; Michałowska, P.; Podgórski, T. The gender dependent influence of sodium bicarbonate supplementation on anaerobic power and specific performance in female and male wrestlers. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Helge, J.W. A high carbohydrate diet remains the evidence based choice for elite athletes to optimise performance. J. Physiol. 2017, 595, 2775. [Google Scholar] [CrossRef] [Green Version]
- Erlenbusch, M.; Haub, M.; Munoz, K.; MacConnie, S.; Stillwell, B. Effect of high-fat or high-carbohydrate diets on endurance exercise: A meta-analysis. Int. J. Sport Nutr. Exerc. Metab. 2005, 15, 1–14. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Indicator | Diet Type | All | Females | Males |
|---|---|---|---|---|
| n | - | 22 | 11 | 11 |
| Age (years) | - | 29.5 ± 4.4 | 30.5 ± 3.3 | 28.5 ± 5.3 |
| Body height (cm) | - | 172 ± 8 | 166 ± 7 | 177 ± 5 |
| Body mass (kg) | Customary diet Ketogenic diet | 71.1 ± 12.8 70.1 ± 12.6 | 60.8 ± 4.9 60.0 ± 4.4 | 81.3 ± 9.5 80.1 ± 9.7 |
| Indicator | Diet Type | All | Females | Males |
|---|---|---|---|---|
| Texh (s) | Customary diet Ketogenic diet p | 636 ± 178 634 ± 182 0.903 * | 510 ± 104 496 ± 102 0.485 * | 761 ± 146 771 ± 134 0.706 * |
| Wmax (W) | Customary diet Ketogenic diet p | 252 ± 61 248 ± 60 0.388 ** | 205 ± 29 198 ± 26 0.345 ** | 300 ± 43 298 ± 38 0.753 ** |
| HRmax (bpm) | Customary diet Ketogenic diet p | 181 ± 9 184 ± 8 0.042 * | 176 ± 6 180 ± 6 0.014 * | 185 ± 10 187 ± 8 0.447 * |
| TGET (s) | Customary diet Ketogenic diet p | 486 ± 122 479 ± 126 0.414 * | 401 ± 79 386 ± 77 0.082 * | 571 ± 95 572 ± 92 0.936 * |
| VO2GET (L·min−1) | Customary diet Ketogenic diet p | 2.68 ± 0.63 2.64 ± 0.72 0.524 * | 2.18 ± 0.34 2.03 ± 0.26 0.105 * | 3.19 ± 0.41 3.26 ± 0.43 0.388 * |
| %VO2maxGET | Customary diet Ketogenic diet p | 82.7 ± 6.4 83.8 ± 8.5 0.592 ** | 83.9 ± 5.7 83.2 ± 9.7 0.929 ** | 81.4 ± 7.1 84.5 ± 7.6 0.385 * |
| WGET (W) | Customary diet Ketogenic diet p | 210 ± 44 211 ± 49 0.747 * | 175 ± 19 170 ± 27 0.463 ** | 245 ± 31 252 ± 26 0.082 * |
| HRGET (bpm) | Customary diet Ketogenic diet p | 171 ± 7 173 ± 7 0.044 * | 170 ± 7 171 ± 4 0.615 * | 171 ± 8 175 ± 8 0.029 * |
| Indicator | Diet Type | Pre-ICT Exercise | Post-ICT Exercise | ||||
|---|---|---|---|---|---|---|---|
| All | Females | Males | All | Females | Males | ||
| pH | Customary diet Ketogenic diet p | 7.405 ± 0.027 7.404 ± 0.023 0.895 * | 7.407 ± 0.021 7.405 ± 0.017 0.732 * | 7.402 ± 0.038 7.404 ± 0.033 0.753 ** | 7.239 ± 0.056 7.257 ± 0.046 0.178 * | 7.253 ± 0.059 7.259 ± 0.038 0.676 * | 7.213 ± 0.044 7.253 ± 0.062 0.028 ** |
| H+ (mmol·L−1) | Customary diet Ketogenic diet p | 39.41 ± 2.49 39.45 ± 2.02 0.944 * | 39.21 ± 1.95 39.42 ± 1.51 0.764 * | 39.77 ± 3.46 39.52 ± 2.92 0.753 ** | 58.16 ± 7.55 55.61 ± 6.00 0.155 * | 56.30 ± 7.82 55.24 ± 4.96 0.569 * | 61.57 ± 6.22 56.28 ± 8.07 0.028 ** |
| BE (mmol·L−1) | Customary diet Ketogenic diet p | −0.3 ± 1.8 −0.7 ± 2.0 0.502 * | −0.3 ± 1.9 −1.0 ± 2.1 0.411 * | −0.2 ± 1.8 −0.1 ± 1.6 0.753 ** | −13.8 ± 2.7 −13.1 ± 2.4 0.351 * | −13.5 ± 2.7 −13.1 ± 2.2 0.569 * | −14.4 ± 3.0 −13.1 ± 2.9 0.028 ** |
| HCO3− (mmol·L−1) | Customary diet Ketogenic diet p | 24.2 ± 1.4 23.9 ± 1.4 0.524 * | 24.2 ± 1.4 23.7 ± 1.5 0.393 * | 24.2 ± 1.4 24.4 ± 1.2 0.753 ** | 15.0 ± 1.7 15.4 ± 1.5 0.384 * | 15.2 ± 1.7 15.4 ± 1.4 0.620 * | 14.8 ± 1.7 15.5 ± 1.8 0.028 ** |
| Anion gap (mmol·L−1) | Customary diet Ketogenic diet p | 10.6 ± 1.6 10.8 ± 1.8 0.501 ** | 10.9 ± 1.8 10.8 ± 2.0 0.878 ** | 10.1 ± 1.0 10.9 ± 1.4 0.208 ** | 20.7 ± 5.9 22.0 ± 2.2 0.737 ** | 19.8 ± 7.2 21.8 ± 2.2 0.610 ** | 22.5 ± 1.7 22.4 ± 2.4 0.028 ** |
| Lactate (mmol·L−1) | Customary diet Ketogenic diet p | 1.8 ± 0.5 1.8 ± 0.3 0.568 * | 1.7 ± 0.3 1.8 ± 0.2 0.758 * | 2.1 ± 0.7 1.8 ± 0.3 0.345 ** | 12.4 ± 2.6 11.1 ± 2.1 0.042 * | 11.6 ± 2.7 10.8 ± 1.9 0.215 * | 13.9 ± 1.6 11.7 ± 2.4 0.028 ** |
| Osmolarity (mmol·kg−1) | Customary diet Ketogenic diet p | 290.7 ± 2.8 289.7 ± 3.9 0.362 * | 290.9 ± 3.1 289.9 ± 3.7 0.299 * | 290.2 ± 2.4 289.4 ± 4.5 0.917 ** | 293.0 ± 2.5 292.7 ± 4.4 0.819 * | 292.8 ± 2.8 292.7 ± 4.7 0.953 * | 293.5 ± 2.0 292.8 ± 4.1 0.116 ** |
| Indicator | Diet Type | Pre-ICT Exercise | Post-ICT Exercise | ||||
|---|---|---|---|---|---|---|---|
| All | Females | Males | All | Females | Males | ||
| Testosterone (ng·mL−1) | Customary diet Ketogenic diet p | 2.64 ± 2.15 3.02 ± 2.43 0.569 ** | 1.31 ± 0.37 1.74 ± 1.66 0.594 ** | 5.59 ± 1.09 5.84 ± 0.88 0.500 ** | 2.72 ± 2.27 3.16 ± 2.63 0.982 ** | 1.27 ± 0.37 1.75 ± 1.79 0.534 ** | 5.90 ± 0.75 6.25 ± 0.67 0.500 ** |
| Cortisol (ng·mL−1) | Customary diet Ketogenic diet p | 306.7 ± 108.9 293.7 ± 97.5 0.756 ** | 314.1 ± 127.7 297.3 ± 106.5 0.639 * | 290.4 ± 56.8 285.7 ± 84.7 0.893 ** | 362.1 ± 126.3 326.5 ± 109.8 0.183 * | 359.4 ± 133.4 337.0 ± 123.0 0.537 * | 368.1 ± 123.7 303.3 ± 80.2 0.080 ** |
| T/C ratio | Customary diet Ketogenic diet p | 0.0096 ± 0.0077 0.0084 ± 0.0081 0.079 ** | 0.0049 ± 0.0026 0.0039 ± 0.0015 0.101 * | 0.0197 ± 0.0045 0.0181 ± 0.0083 0.500 ** | 0.0111 ± 0.0099 0.0108 ± 0.0102 0.501 ** | 0.0059 ± 0.0040 0.0054 ± 0.0044 0.534 ** | 0.0225 ± 0.0094 0.0226 ± 0.0094 0.893 ** |
| Creatine kinase (U·L−1) | Customary diet Ketogenic diet p | 318.0 ± 242.6 250.2 ± 223.5 0.469 ** | 280.7 ± 260.4 144.2 ± 76.0 0.110 ** | 400.2 ± 197.3 483.3 ± 272.1 0.345 ** | 362.6 ± 310.8 277.3 ± 245.3 0.717 ** | 313.8 ± 334.6 163.5 ± 84.1 0.248 ** | 469.9 ± 247.8 527.7 ± 306.6 0.345 ** |
| LDH (U·L−1) | Customary diet Ketogenic diet p | 425 ± 78 402 ± 55 0.293 * | 443 ± 66 402 ± 55 0.075 ** | 386 ± 95 403 ± 63 0.893 ** | 470 ± 95 448 ± 68 0.416 * | 478 ± 79 454 ± 64 0.594 ** | 454 ± 133 436 ± 83 0.686 ** |
| Glucose (mg·dL−1) | Customary diet Ketogenic diet p | 89 ± 9 87 ± 9 0.592 * | 86 ± 7 85 ± 9 0.699 * | 93 ± 11 91 ± 9 0.753 ** | 118 ± 19 121 ± 20 0.541 * | 121 ± 21 123 ± 17 0.728 * | 111 ± 13 116 ± 25 0.043 ** |
| Urea (mmol·L−1) | Customary diet Ketogenic diet p | 6.5 ± 1.5 7.3 ± 2.1 0.108 * | 7.0 ± 1.5 7.4 ± 2.4 0.482 * | 5.5 ± 0.8 7.3 ± 1.8 0.080 ** | 6.4 ± 1.2 7.1 ± 1.9 0.118 * | 6.7 ± 1.3 7.2 ± 2.3 0.452 * | 5.8 ± 0.9 7.1 ± 1.2 0.080 ** |
| AlAT (U·L−1) | Customary diet Ketogenic diet p | 18.0 ± 4.3 19.8 ± 6.9 0.279 * | 17.9 ± 4.5 17.1 ± 4.5 0.663 * | 18.4 ± 4.4 25.8 ± 7.9 0.043 ** | 23.1 ± 6.5 24.7 ± 5.9 0.134 ** | 22.4 ± 7.1 22.4 ± 4.8 0.534 ** | 24.7 ± 5.3 29.9 ± 5.2 0.043 ** |
| AspAT (U·L−1) | Customary diet Ketogenic diet p | 35.3 ± 11.1 33.2 ± 7.4 0.569 ** | 34.9 ± 13.0 30.4 ± 5.6 0.374 ** | 36.1 ± 6.6 39.2 ± 7.7 0.893 ** | 38.4 ± 13.9 36.1 ± 7.3 0.642 ** | 37.7 ± 16.8 33.3 ± 6.2 1.000 ** | 39.9 ± 3.0 42.1 ± 6.4 0.345 ** |
| Bilirubin (μmol·L−1) | Customary diet Ketogenic diet p | 10 ± 5 14 ± 9 0.042 * | 12 ± 4 15 ± 9 0.248 * | 6 ± 4 11 ± 8 0.059 ** | 19 ± 8 20 ± 8 0.709 * | 20 ± 9 19 ± 9 0.786 * | 20 ± 9 19 ± 9 0.043 ** |
| Indicator | Diet Type | Pre-ICT Exercise | Post-ICT Exercise | ||||
|---|---|---|---|---|---|---|---|
| All | Females | Males | All | Females | Males | ||
| WBC (109·L−1) | Customary diet Ketogenic diet p | 7.2 ± 1.7 7.7 ± 2.1 0.359 * | 7.5 ± 1.8 8.1 ± 2.3 0.409 * | 6.5 ± 1.5 6.7 ± 1.3 0.686 ** | 11.5 ± 2.3 11.9 ± 3.4 0.540 * | 12.2 ± 2.1 12.8 ± 3.7 0.556 * | 9.9 ± 2.3 10.0 ± 1.2 0.893 ** |
| LYM (109·L−1) | Customary diet Ketogenic diet p | 2.7 ± 0.7 2.7 ± 0.5 0.920 * | 2.8 ± 0.7 2.7 ± 0.6 0.821 * | 2.5 ± 0.7 2.6 ± 0.3 0.893 ** | 5.2 ± 1.3 5.1 ± 1.0 0.889 * | 5.4 ± 1.2 5.3 ± 1.1 0.878 ** | 4.6 ± 1.4 4.7 ± 0.8 0.893 ** |
| MON (109·L−1) | Customary diet Ketogenic diet p | 0.5 ± 0.2 0.6 ± 0.2 0.005 ** | 0.5 ± 0.2 0.6 ± 0.2 0.036 * | 0.4 ± 0.1 0.5 ± 0.1 0.080 ** | 0.7 ± 0.2 0.9 ± 0.3 0.017 * | 0.8 ± 0.2 1.0 ± 0.3 0.056 * | 0.6 ± 0.2 0.7 ± 0.1 0.043 ** |
| GRA (109·L−1) | Customary diet Ketogenic diet p | 4.1 ± 1.2 4.4 ± 1.7 0.423 * | 4.3 ± 1.2 4.8 ± 1.9 0.412 * | 3.6 ± 1.3 3.6 ± 1.2 0.893 ** | 5.6 ± 1.4 5.9 ± 2.7 0.733 ** | 6.0 ± 1.2 6.5 ± 2.9 0.575 ** | 4.7 ± 1.7 4.6 ± 1.3 0.893 ** |
| LYM (%) | Customary diet Ketogenic diet p | 37.7 ± 8.1 36.4 ± 8.7 0.537 * | 36.8 ± 5.7 35.1 ± 8.7 0.500 * | 39.5 ± 12.5 39.3 ± 8.8 0.686 ** | 44.9 ± 8.4 44.3 ± 9.3 0.750 * | 44.2 ± 6.4 42.9 ± 8.8 0.523 * | 46.5 ± 12.6 47.3 ± 10.7 0.893 ** |
| MON (%) | Customary diet Ketogenic diet p | 6.2 ± 1.3 7.6 ± 1.2 0.013 * | 6.3 ± 1.5 7.5 ± 1.4 0.087 * | 6.2 ± 0.5 7.7 ± 0.8 0.080 ** | 6.4 ± 1.0 7.5 ± 0.9 0.019 * | 6.2 ± 1.2 7.5 ± 0.9 0.016 * | 6.6 ± 0.7 7.5 ± 1.1 0.345 ** |
| GRA (%) | Customary diet Ketogenic diet p | 56.1 ± 8.3 56.0 ± 9.1 0.973 * | 56.9 ± 6.1 57.4 ± 9.1 0.856 * | 54.3 ± 12.5 53.0 ± 9.4 0.893 ** | 48.7 ± 8.6 48.2 ± 9.6 0.808 * | 49.6 ± 6.5 49.6 ± 9.2 0.989 * | 46.9 ± 12.9 45.2 ± 10.8 0.686 ** |
| RBC (1012·L−1) | Customary diet Ketogenic diet p | 5.87 ± 0.26 5.85 ± 0.27 0.156 * | 5.84 ± 0.28 5.82 ± 0.30 0.221 * | 5.92 ± 0.21 5.90 ± 0.18 0.500 ** | 5.79 ± 0.24 5.80 ± 0.27 0.715 * | 5.76 ± 0.26 5.77 ± 0.29 0.798 * | 5.86 ± 0.19 5.87 ± 0.21 0.500 ** |
| HGB (mmol·L−1) | Customary diet Ketogenic diet p | 11.15 ± 0.22 10.76 ± 0.32 0.002 * | 11.19 ± 0.19 10.71 ± 0.36 0.003 * | 11.04 ± 0.26 10.88 ± 0.21 0.345 ** | 11.01 ± 0.21 10.68 ± 0.35 0.004 * | 11.01 ± 0.22 10.58 ± 0.34 0.013 ** | 11.03 ± 0.22 10.89 ± 0.32 0.345 ** |
| HCT (L·L−1) | Customary diet Ketogenic diet p | 0.398 ± 0.047 0.403 ± 0.024 0.577 * | 0.376 ± 0.028 0.392 ± 0.016 0.113 * | 0.447 ± 0.044 0.428 ± 0.018 0.225 ** | 0.413 ± 0.039 0.418 ± 0.038 0.506 * | 0.405 ± 0.038 0.405 ± 0.037 0.997 * | 0.429 ± 0.038 0.447 ± 0.022 0.500 ** |
| MCV (fL) | Customary diet Ketogenic diet p | 85.4 ± 3.7 85.7 ± 3.9 0.196 * | 85.8 ± 4.1 86.1 ± 4.4 0.237 * | 84.6 ± 3.0 84.8 ± 2.5 0.500 ** | 86.2 ± 3.8 86.3 ± 3.9 0.340 * | 86.5 ± 4.2 86.8 ± 4.3 0.169 * | 85.4 ± 2.8 85.3 ± 3.0 0.500 ** |
| MCH (fmol) | Customary diet Ketogenic diet p | 1.90 ± 0.10 1.84 ± 0.09 0.001 * | 1.92 ± 0.11 1.84 ± 0.10 0.012 * | 1.87 ± 0.06 1.84 ± 0.05 0.285 ** | 1.90 ± 0.10 1.84 ± 0.10 0.003 * | 1.91 ± 0.11 1.84 ± 0.11 0.004 * | 1.88 ± 0.07 1.86 ± 0.08 0.418 ** |
| MCHC (mmol·L−1) | Customary diet Ketogenic diet p | 22.25 ± 0.43 21.51 ± 0.65 0.002 * | 22.33 ± 0.38 21.40 ± 0.72 0.004 * | 22.08 ± 0.54 21.76 ± 0.41 0.944 ** | 22.08 ± 0.43 21.36 ± 0.71 0.004 * | 22.09 ± 0.45 21.17 ± 0.68 0.005 * | 22.06 ± 0.42 21.77 ± 0.63 0.345 ** |
| RDW (%) | Customary diet Ketogenic diet p | 12.9 ± 1.0 13.0 ± 0.8 0.386 * | 12.7 ± 1.1 12.8 ± 0.8 0.407 * | 13.2 ± 0.5 13.3 ± 0.9 0.787 ** | 13.0 ± 1.0 13.2 ± 0.9 0.196 * | 12.9 ± 1.1 13.1 ± 1.0 0.294 * | 13.1 ± 0.8 13.3 ± 0.5 0.418 ** |
| PLT (109·L−1) | Customary diet Ketogenic diet p | 248 ± 76 266 ± 81 0.289 * | 259 ± 86 277 ± 95 0.451 * | 225 ± 47 243 ± 31 0.345 ** | 265 ± 77 305 ± 95 0.070 * | 270 ± 90 324 ± 100 0.059 * | 253 ± 43 264 ± 77 0.893 ** |
| MPV (fL) | Customary diet Ketogenic diet p | 8.7 ± 0.8 8.9 ± 0.9 0.157 * | 8.7 ± 0.9 8.9 ± 1.0 0.212 * | 8.7 ± 0.6 8.9 ± 0.9 0.787 ** | 8.9 ± 0.8 9.1 ± 0.9 0.179 * | 8.9 ± 0.9 9.1 ± 1.0 0.378 * | 8.7 ± 0.8 9.0 ± 0.9 0.273 ** |
| PCT (cL·L−1) | Customary diet Ketogenic diet p | 0.166 ± 0.035 0.186 ± 0.040 0.035 * | 0.163 ± 0.041 0.186 ± 0.044 0.085 * | 0.172 ± 0.020 0.186 ± 0.032 0.225 ** | 0.189 ± 0.047 0.225 ± 0.053 0.019 * | 0.190 ± 0.056 0.232 ± 0.054 0.059 * | 0.186 ± 0.021 0.210 ± 0.053 0.138 ** |
| PDW (%) | Customary diet Ketogenic diet p | 12.6 ± 1.2 12.8 ± 1.3 0.561 * | 12.3 ± 1.3 13.0 ± 1.4 0.216 * | 13.3 ± 0.8 12.5 ± 0.9 0.043 ** | 12.8 ± 1.2 12.8 ± 1.1 0.845 * | 12.7 ± 1.2 12.7 ± 1.2 0.967 * | 13.1 ± 1.5 12.8 ± 1.2 0.787 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Durkalec-Michalski, K.; Nowaczyk, P.M.; Główka, N.; Ziobrowska, A.; Podgórski, T. Is a Four-Week Ketogenic Diet an Effective Nutritional Strategy in CrossFit-Trained Female and Male Athletes? Nutrients 2021, 13, 864. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13030864
Durkalec-Michalski K, Nowaczyk PM, Główka N, Ziobrowska A, Podgórski T. Is a Four-Week Ketogenic Diet an Effective Nutritional Strategy in CrossFit-Trained Female and Male Athletes? Nutrients. 2021; 13(3):864. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13030864
Chicago/Turabian StyleDurkalec-Michalski, Krzysztof, Paulina M. Nowaczyk, Natalia Główka, Anna Ziobrowska, and Tomasz Podgórski. 2021. "Is a Four-Week Ketogenic Diet an Effective Nutritional Strategy in CrossFit-Trained Female and Male Athletes?" Nutrients 13, no. 3: 864. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13030864