
Intraperitoneal Administration of Short-Chain Fatty Acids Improves Lipid Metabolism of Long–Evans Rats in a Sex-Specific Manner

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experiment Design
2.2. Blood and Tissue Sampling
2.3. Analysis of Biochemical Parameters
2.4. Gene Expression Analysis
2.5. Lipidomics Analysis
2.6. Data Analysis
3. Results
3.1. Effect of SCFAs on Body Weight, Food Intake, Glucose, and NEFA Levels
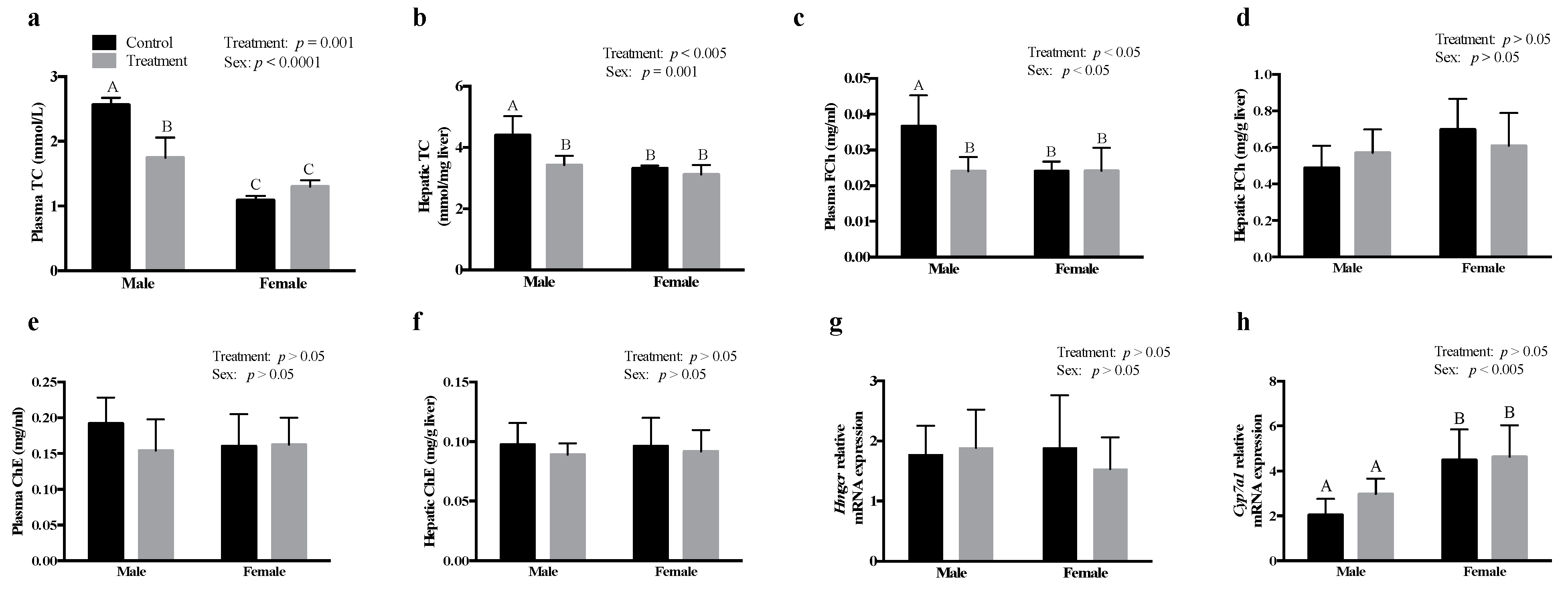
3.2. SCFAs Had Sex-Specific Effects on Cholesterol Metabolism
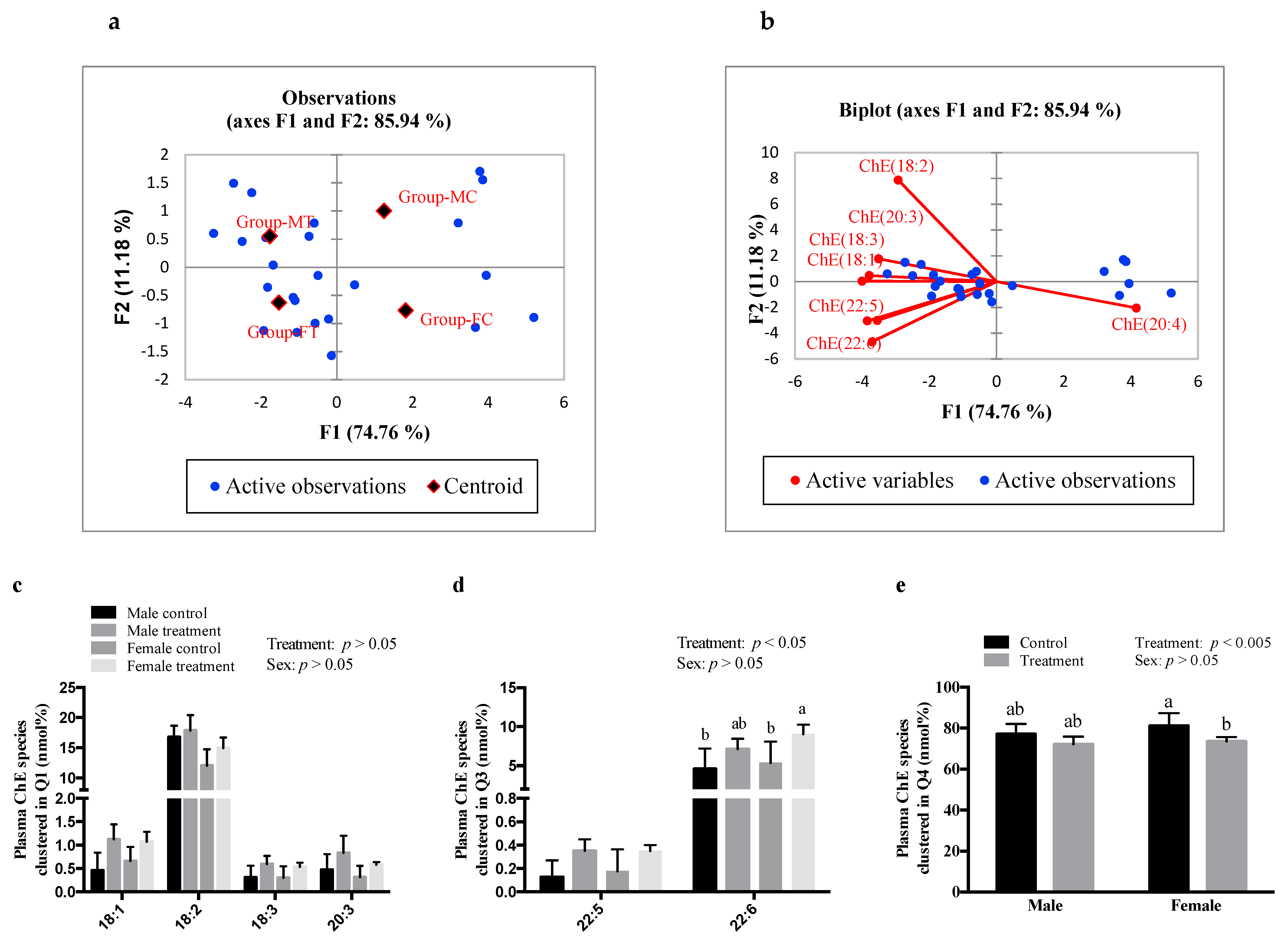
3.3. SCFAs Altered Plasma and Hepatic ChE Fatty Acid Composition
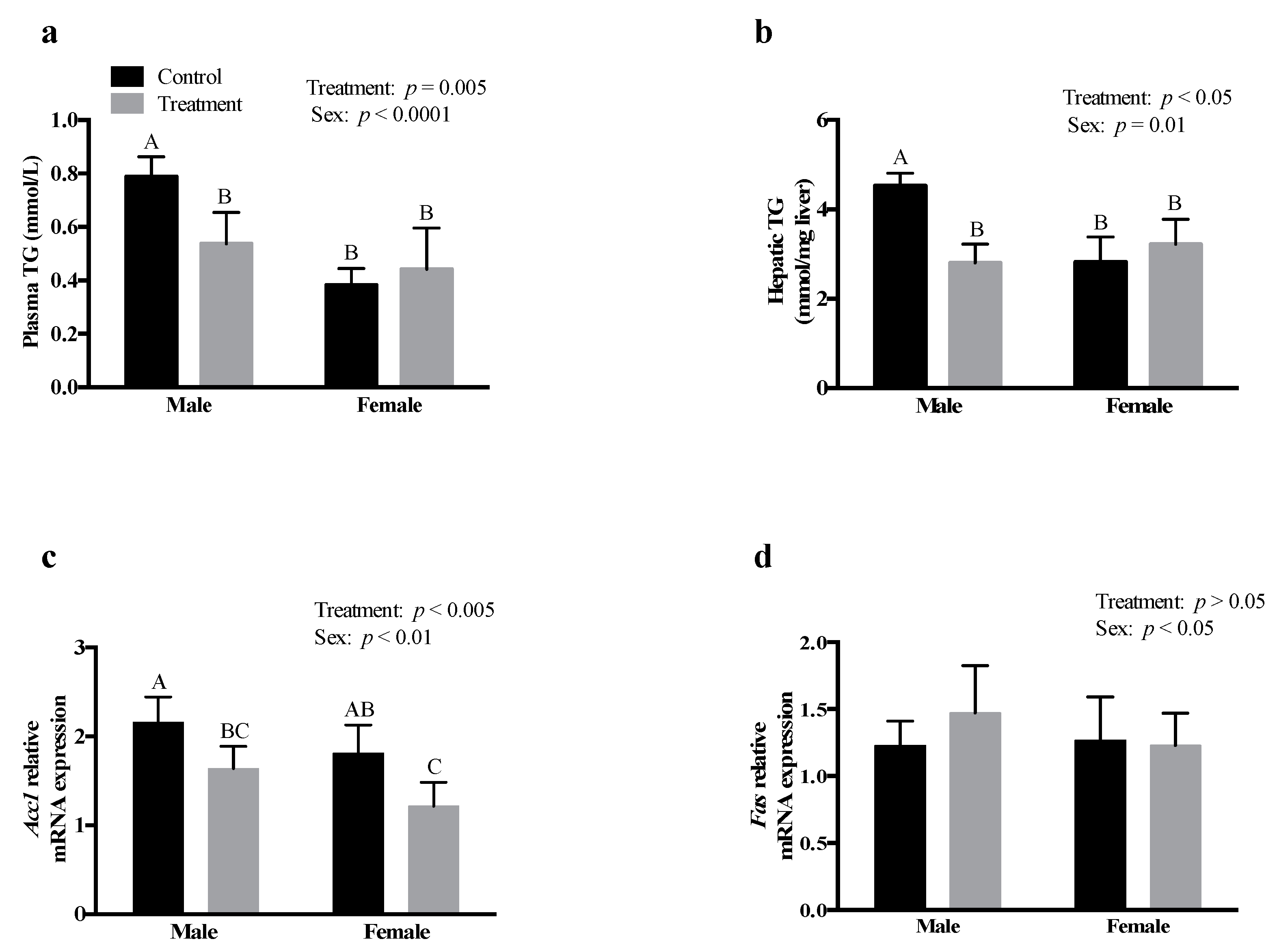
3.4. SCFAs Had Sex-Specific Effects on TG Metabolism
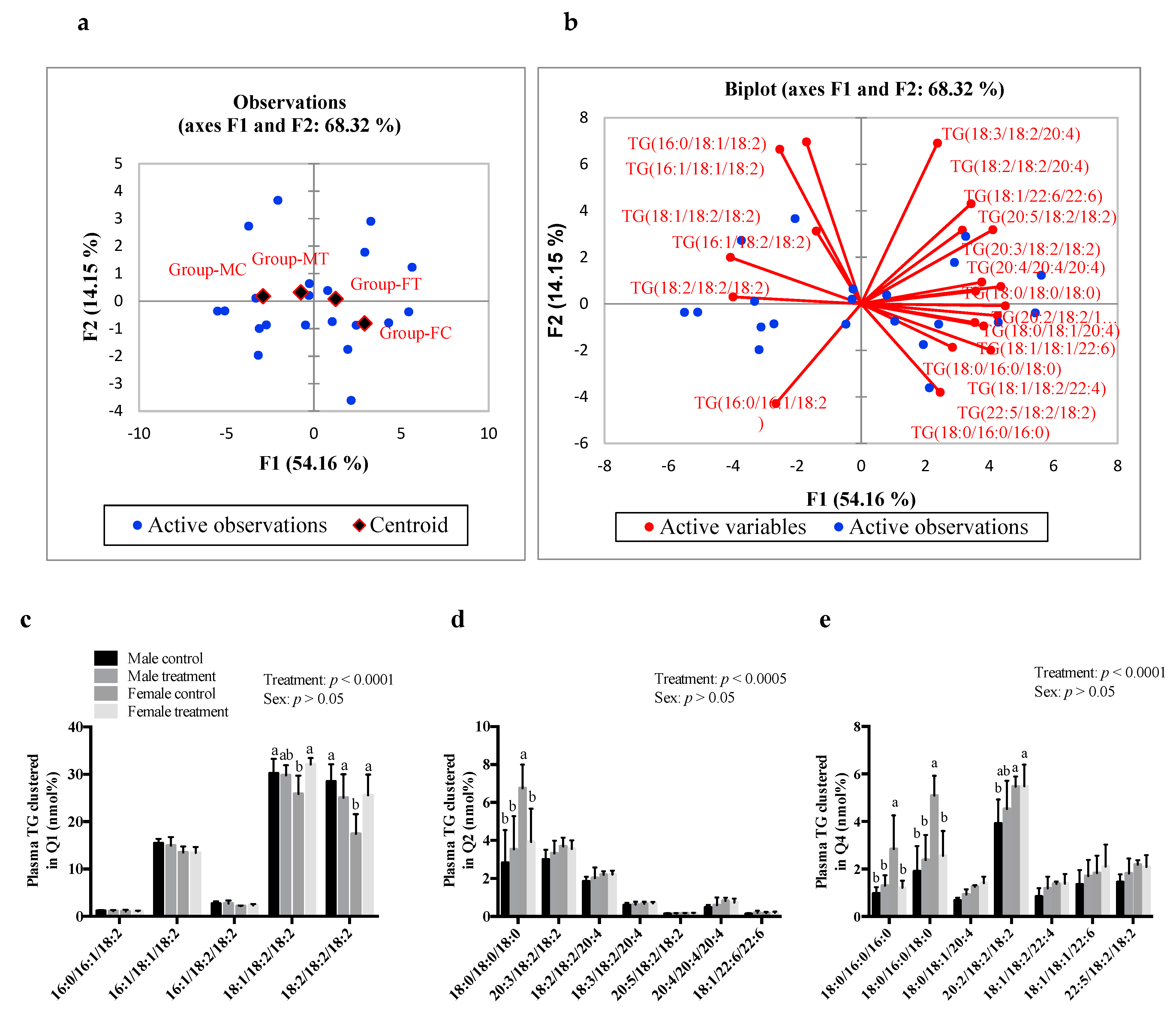
3.5. SCFAs Had Sex-Specific Effects on TG Fatty Acid Composition
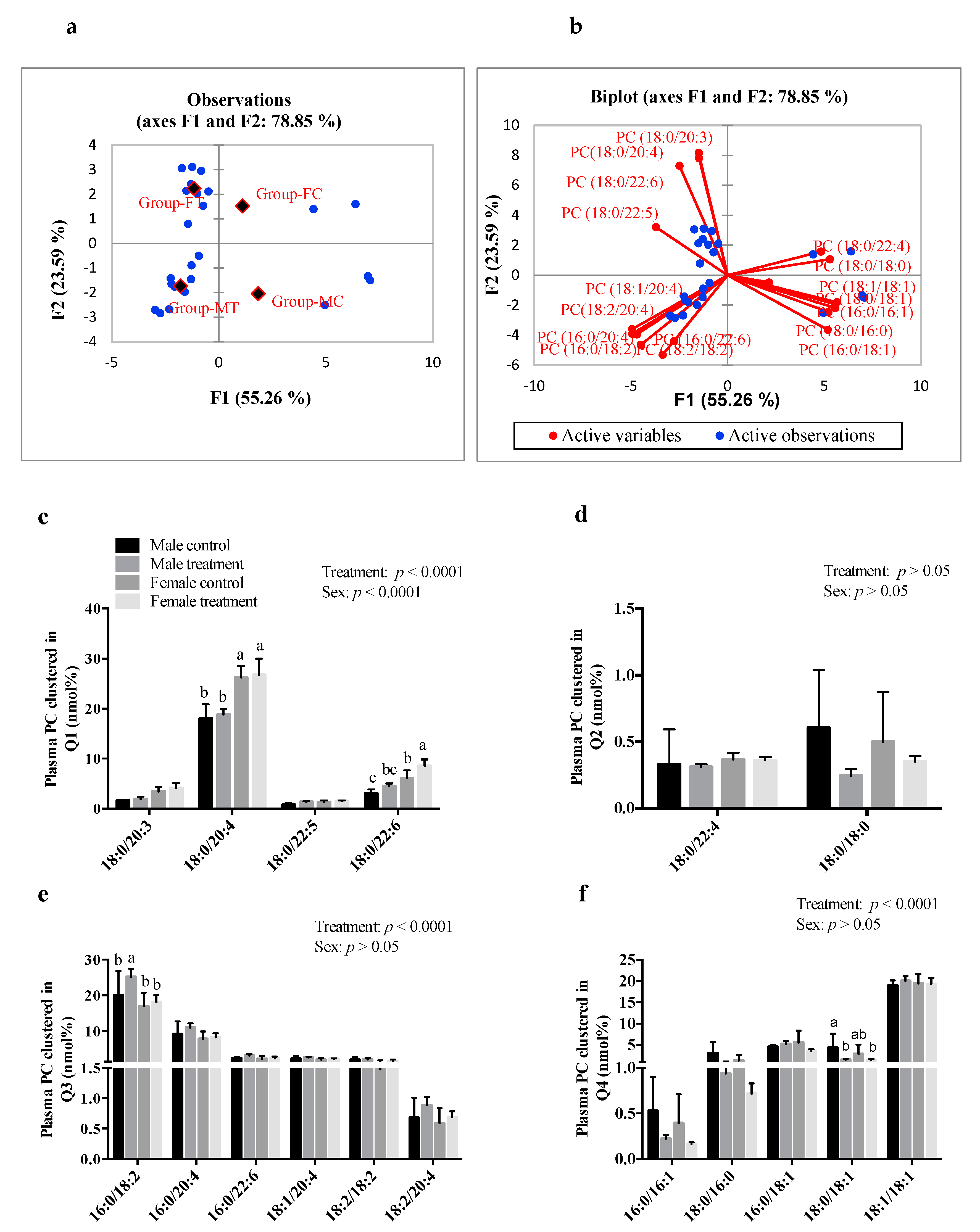
3.6. SCFAs Had Sex-Specific Effects on Plasma and Hepatic PC Fatty Acid Composition
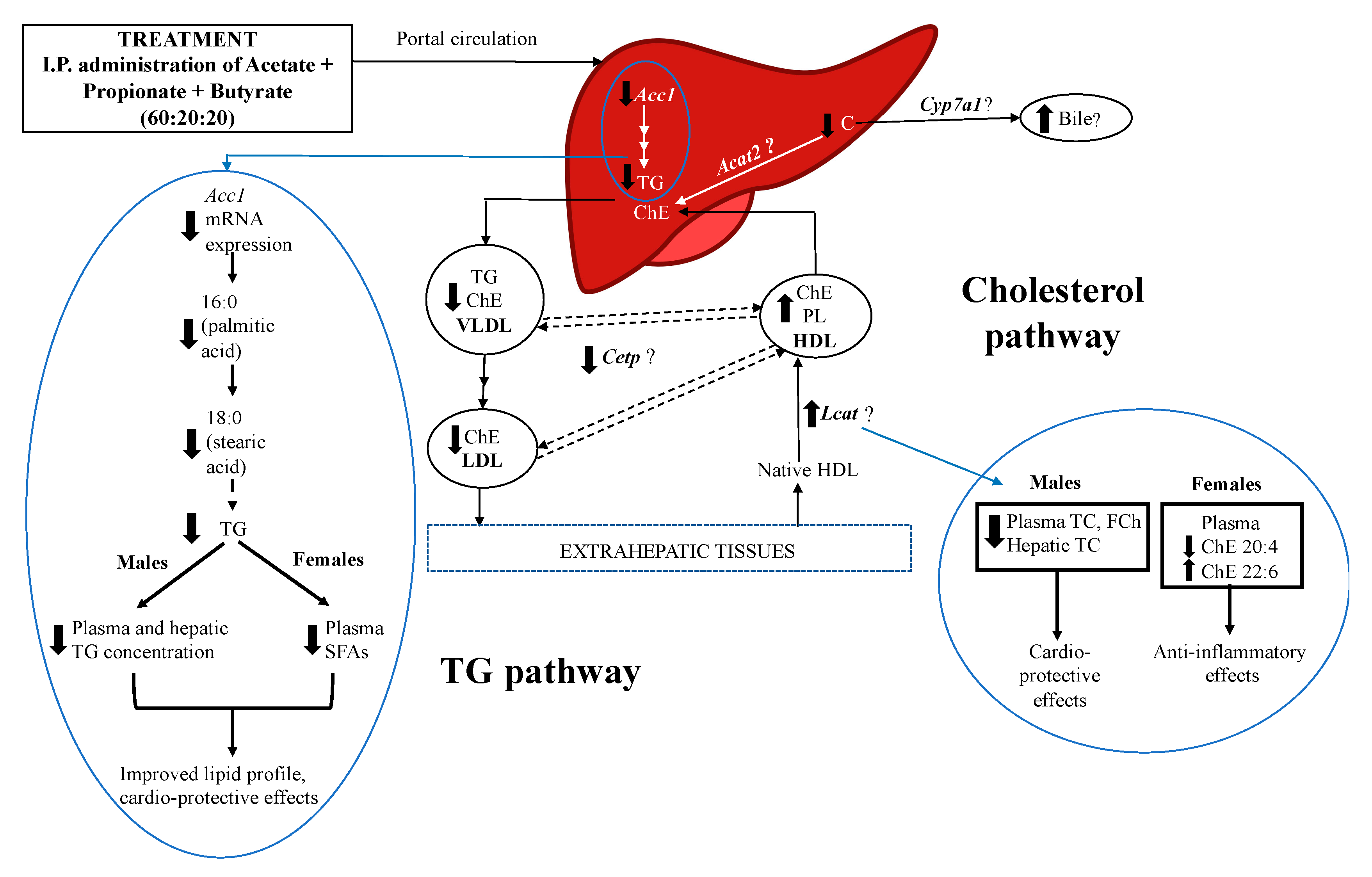
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Byrne, C.S.; Chambers, E.S.; Morrison, D.J.; Frost, G. The role of short chain fatty acids in appetite regulation and energy homeostasis. Int. J. Obes. 2015, 39, 1331–1338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohira, H.; Tsutsui, W.; Fujioka, Y. Are Short Chain Fatty Acids in Gut Microbiota Defensive Players for Inflammation and Atherosclerosis? J. Atheroscler. Thromb. 2017, 24, 660–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Topping, D.L.; Clifton, P.M. Short-Chain Fatty Acids and Human Colonic Function: Roles of Resistant Starch and Nonstarch Polysaccharides. Physiol. Rev. 2001, 81, 1031–1064. [Google Scholar] [CrossRef]
- Den Besten, G.; Bleeker, A.; Gerding, A.; van Eunen, K.; Havinga, R.; van Dijk, T.H.; Oosterveer, M.H.; Jonker, J.W.; Groen, A.K.; Reijngoud, D.J.; et al. Short-chain fatty acids protect against high-fat diet-induced obesity via a pparg-dependent switch from lipogenesis to fat oxidation. Diabetes 2015, 64, 2398–2408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.-J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hijova, E.; Chmelarova, A. Short chain fatty acids and colonic health. Bratisl. Lekaárske List. 2007, 108, 354–358. [Google Scholar]
- Binder, H.J. Role of colonic short-chain fatty acid transport in diarrhea. Annu. Rev. Physiol. 2010, 72, 297–313. [Google Scholar] [CrossRef] [PubMed]
- Levrat, M.-A.; Rémésy, C.; Demigné, C. High Propionic Acid Fermentations and Mineral Accumulation in the Cecum of Rats Adapted to Different Levels of Inulin. J. Nutr. 1991, 121, 1730–1737. [Google Scholar] [CrossRef] [PubMed]
- Topping, D.L.; Illman, R.J.; Clarke, J.M.; Trimble, R.P.; Jackson, K.A.; Marsono, Y. Dietary Fat and Fiber Alter Large Bowel and Portal Venous Volatile Fatty Acids and Plasma Cholesterol but Not Biliary Steroids in Pigs. J. Nutr. 1993, 123, 133–143. [Google Scholar] [CrossRef]
- Cummings, J.H.; Pomare, E.W.; Branch, W.J.; Naylor, C.P.; Macfarlane, G.T. Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut 1987, 28, 1221–1227. [Google Scholar] [CrossRef] [Green Version]
- Remely, M.; Aumueller, E.; Merold, C.; Dworzak, S.; Hippe, B.; Zanner, J.; Pointner, A.; Brath, H.; Haslberger, A.G. Effects of short chain fatty acid producing bacteria on epigenetic regulation of FFAR3 in type 2 diabetes and obesity. Gene 2014, 537, 85–92. [Google Scholar] [CrossRef]
- Canfora, E.E.; Jocken, J.W.; Blaak, E.E. Short-chain fatty acids in control of body weight and insulin sensitivity. Nat. Rev. Endocrinol. 2015, 11, 577–591. [Google Scholar] [CrossRef]
- den Besten, G.; Lange, K.; Havinga, R.; Van Dijk, T.H.; Gerding, A.; Van Eunen, K.; Müller, M.; Groen, A.K.; Hooiveld, G.J.; Bakker, B.M.; et al. Gut-derived short-chain fatty acids are vividly assimilated into host carbohydrates and lipids. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 305, G900–G910. [Google Scholar] [CrossRef]
- Jocken, J.W.E.; Hernández, M.A.G.; Hoebers, N.T.H.; Morris, M. Short-Chain Fatty Acids Differentially Affect Intracellular Lipolysis in a Human White Adipocyte Model. Front. Endocrinol. 2018, 8, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Fushimi, T.; Suruga, K.; Oshima, Y.; Fukiharu, M.; Tsukamoto, Y.; Goda, T. Dietary acetic acid reduces serum cholesterol and triacylglycerols in rats fed a cholesterol-rich diet. Br. J. Nutr. 2006, 95, 916–924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakakibara, S.; Yamauchi, T.; Oshima, Y.; Tsukamoto, Y.; Kadowaki, T. Acetic acid activates hepatic AMPK and reduces hyperglycemia in diabetic KK-A(y) mice. Biochem. Biophys. Res. Commun. 2006, 344, 597–604. [Google Scholar] [CrossRef] [PubMed]
- Zambell, K.L.; Fitch, M.D.; Fleming, S.E. Acetate and Butyrate Are the Major Substrates for De Novo Lipogenesis in Rat Colonic Epithelial Cells. J. Nutr. 2003, 133, 3509–3515. [Google Scholar] [CrossRef]
- Zhao, Y.; Liu, J.; Hao, W.; Zhu, H.; Liang, N.; Zhen-Yu, C.; Ma, K.Y.; Chen, Z.-Y. Structure-Specific Effects of Short-Chain Fatty Acids on Plasma Cholesterol Concentration in Male Syrian Hamsters. J. Agric. Food Chem. 2017. [Google Scholar] [CrossRef]
- Varlamov, O.; Bethea, C.L. Sex-specific differences in lipid and glucose metabolism. Front. Endocrinol. 2015, 5, 241. [Google Scholar] [CrossRef] [Green Version]
- Christ, E.J.V.J. Fatty acid synthesis in mitochondria elongation of short-chain fatty acids and formation of unsaturated long-chain fatty acids. Biochim. Biophys. Acta (BBA)/Lipids Lipid Metab. 1968, 152, 50–62. [Google Scholar] [CrossRef]
- Rosqvist, F.; Bjermo, H.; Kullberg, J.; Johansson, L.; Michaëlsson, K.; Ahlström, H.; Lind, L.; Risérus, U. Fatty acid composition in serum cholesterol esters and phospholipids is linked to visceral and subcutaneous adipose tissue content in elderly individuals: A cross-sectional study. Lipids Health Dis. 2017, 16, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Frost, G.; Sleeth, M.L.; Sahuri-Arisoylu, M.; Lizarbe, B.; Cerdan, S.; Brody, L.P.; Anastasovska, J.; Ghourab, S.; Hankir, M.K.; Zhang, S.; et al. The short-chain fatty acid acetate reduces appetite via a central homeostatic mechanism. Nat. Commun. 2014, 5, 3611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipids from animal animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Chechi, K.; Herzberg, G.R.; Cheema, S.K. Prostaglandins, Leukotrienes and Essential Fatty Acids Maternal dietary fat intake during gestation and lactation alters tissue fatty acid composition in the adult offspring of C57Bl / 6 mice. Prostaglandins Leukot. Essent. Fat. Acids 2010, 83, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S. Quantitation of Lipid Classes by Thin-Layer Chromatography with Flame Ionization. Curr. Protoc. Food Anal. Chem. 2003, 7, 1–14. [Google Scholar] [CrossRef]
- Marshall, K.E.; Thomas, R.H.; Roxin, A.; Chen, E.K.Y.; Brown, J.C.L.; Gillies, E.R.; Sinclair, B.J. Seasonal accumulation of acetylated triacylglycerols by a freeze-tolerant insect. J. Exp. Biol. 2014, 217, 1580–1587. [Google Scholar] [CrossRef] [Green Version]
- Chomczynski, P. Single-Step Method of RNA Isolation by Acid Guanidinium Extraction. Anal. Biochem. 1987, 159, 156–159. [Google Scholar] [CrossRef]
- Balogun, K.A.; Cheema, S.K. Neurochemistry International The expression of neurotrophins is differentially regulated by omega-3 polyunsaturated fatty acids at weaning and postweaning in C57BL/6 mice cerebral cortex. Neurochem. Int. 2014, 66, 33–42. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-∆∆CT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A Rapid Method of Total Lipid Extraction and Purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [Green Version]
- Pham, T.H.; Zaeem, M.; Fillier, T.A.; Nadeem, M.; Vidal, N.P.; Manful, C.; Cheema, S.; Cheema, M.; Thomas, R.H. Targeting Modified Lipids during Routine Lipidomics Analysis using HILIC and C30 Reverse Phase Liquid Chromatography coupled to Mass Spectrometry. Sci. Rep. 2019, 9, 5048. [Google Scholar] [CrossRef] [Green Version]
- Vidal, N.P.; Pham, H.T.; Manful, C.; Pumphrey, R.; Nadeem, M.; Cheema, M.; Galagedara, L.; Leke-Aladekoba, A.; Abbey, L.; Thomas, R. The use of natural media amendments to produce kale enhanced with functional lipids in controlled environment production system. Sci. Rep. 2018, 8, 14771. [Google Scholar] [CrossRef]
- Morrison, D.J.; Preston, T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes 2016, 7, 189–200. [Google Scholar] [CrossRef] [Green Version]
- Gao, Z.; Yin, J.; Zhang, J.; Ward, R.E.; Martin, R.J.; Lefevre, M.; Cefalu, W.T.; Ye, J. Butyrate improves insulin sensitivity and increases energy expenditure in mice. Diabetes 2009, 58, 1509–1517. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.R.; Barrick, C.; Kim, K.; Lindner, J.; Blondeau, B.; Fujimoto, Y.; Shiota, M.; Kesterson, R.A.; Kahn, B.B.; Magnuson, M.A. Deletion of PPARgamma in adipose tissues of mice protects against high fat diet-induced obesity and insulin resistance. Proc. Natl. Acad. Sci. USA 2005, 102, 6207–6212. [Google Scholar] [CrossRef] [Green Version]
- King, M.W. Cholesterol: Synthesis, Metabolism, Regulation. The Medical Biochemistry Page. 2016. Available online: http://themedicalbiochemistrypage.org/cholesterol.php (accessed on 15 August 2018).
- Phelps, T.; Snyder, E.; Rodriguez, E.; Child, H.; Harvey, P. The influence of biological sex and sex hormones on bile acid synthesis and cholesterol homeostasis. Biol. Sex Differ. 2019, 52, 1–12. [Google Scholar] [CrossRef]
- Sorci-Thomas, M.G.; Bhat, S.; Thomas, M.J. Activation of lecithin: Cholesterol acyltransferase by HDL ApoA-I central helices. Future Lipidol. 2009. [Google Scholar] [CrossRef] [Green Version]
- Viollet, B.; Guigas, B.; Leclerc, J.; Hébrard, S.; Lantier, L.; Mounier, R.; Andreelli, F.; Foretz, M. AMP-activated protein kinase in the regulation of hepatic energy metabolism: From physiology to therapeutic perspectives. Acta Physiol. 2009. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.; Bryzgalova, G.; Hedman, E.; Khan, A.; Efendic, S.; Gustafsson, J.-A.; Dahlman-Wright, K. Long-Term Administration of Estradiol Decreases Expression of Hepatic Lipogenic Genes and Improves Insulin Sensitivity in ob/ob Mice: A Possible Mechanism Is through Direct Regulation of Signal Transducer and Activator of Transcription 3. Mol. Endocrinol. 2006, 20, 1287–1299. [Google Scholar] [CrossRef] [Green Version]
- Kimura, I.; Ozawa, K.; Inoue, D.; Imamura, T.; Kimura, K.; Maeda, T.; Terasawa, K.; Kashihara, D.; Hirano, K.; Tani, T.; et al. The gut microbiota suppresses insulin-mediated fat accumulation via the short-chain fatty acid receptor GPR43. Nat. Commun. 2013, 4, 1829. [Google Scholar] [CrossRef] [Green Version]
- Barber, M.C.; Price, N.T.; Travers, M.T. Structure and regulation of acetyl-CoA carboxylase genes of metazoa. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2005. [Google Scholar] [CrossRef]
- Lewis-Barned, N.J.; Sutherland, W.H.; Walker, R.J.; De Jong, A.S.; Walker, H.L.; Edwards, A.E.; Markham, V.; Goulding, A. Plasma cholesteryl ester fatty acid composition, insulin sensitivity, the menopause and hormone replacement therapy. J. Endocrinol. 2000. [Google Scholar] [CrossRef]
- Salomaa, V.; Ahola, I.; Tuomilehto, J.; Aro, A.; Pietinen, P.; Korhonen, H.J.; Penttilä, I. Fatty acid composition of serum cholesterol esters in different degrees of glucose intolerance: A population-based study. Metabolism 1990, 39, 1285–1291. [Google Scholar] [CrossRef]
- Harkewicz, R.; Hartvigsen, K.; Almazan, F.; Dennis, E.A.; Witztum, J.L.; Miller, Y.I. Cholesteryl ester hydroperoxides are biologically active components of minimally oxidized low density lipoprotein. J. Biol. Chem. 2008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lands, B. Omega-3 PUFAs lower the propensity for arachidonic acid cascade overreactions. BioMed Res. Int. 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; He, G.; Peng, Y.; Zhong, W.; Wang, Y.; Zhang, B. Sodium butyrate alleviates adipocyte inflammation by inhibiting NLRP3 pathway. Nat. Publ. Gr. 2015, 5, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratnayake, W.M.N.; Galli, C. Fat and fatty acid terminology, methods of analysis and fat digestion and metabolism: A background review paper. Ann. Nutr. Metab. 2009, 55, 8–43. [Google Scholar] [CrossRef] [PubMed]
- Saito, K.; Goda, K.; Kobayashi, A.; Yamada, N.; Maekawa, K.; Saito, Y.; Sugai, S. Arachidonic acid-containing phosphatidylcholine characterized by consolidated plasma and liver lipidomics as an early onset marker for tamoxifen-induced hepatic phospholipidosis. J. Appl. Toxicol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Chalil, A.; Kitson, A.P.; Henao, J.J.A.; Marks, K.A.; Elzinga, J.L.; Lamontagne-Kam, D.M.E.; Chalil, D.; Badoud, F.; Mutch, D.M.; Stark, K.D. PEMT, Δ6 desaturase, and palmitoyldocosahexaenoyl phosphatidylcholine are increased in rats during pregnancy. J. Lipid Res. 2018, 59, 123–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grimm, M.O.W.; Grösgen, S.; Rothhaar, T.L.; Burg, V.K.; Hundsdörfer, B.; Haupenthal, V.J.; Friess, P.; Müller, U.; Fassbender, K.; Riemenschneider, M.; et al. Intracellular APP Domain Regulates Serine-Palmitoyl-CoA Transferase Expression and Is Affected in Alzheimer’s Disease. Int. J. Alzheimers Dis. 2011, 2011, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Orešič, M.; Seppänen-Laakso, T.; Sun, D.; Tang, J.; Therman, S.; Viehman, R.; Mustonen, U.; van Erp, T.G.; Hyötyläinen, T.; Thompson, P.; et al. Phospholipids and insulin resistance in psychosis: A lipidomics study of twin pairs discordant for schizophrenia. Genome Med. 2012, 4, 1. [Google Scholar] [CrossRef]
- Li, Z.; Jiang, H.; Ding, T.; Lou, C.; Bui, H.H.; Kuo, M.S.; Jiang, X.C. Deficiency in lysophosphatidylcholine acyltransferase 3 reduces plasma levels of lipids by reducing lipid absorption in mice. Gastroenterology 2015, 149, 1519–1529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rong, X.; Wang, B.; Dunham, M.M.; Hedde, P.N.; Wong, J.S.; Gratton, E.; Young, S.G.; Ford, A.D.; Tontonoz, P. Lpcat3-dependent production of arachidonoyl phospholipids is a key determinant of triglyceride secretion. Elife 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Christie, W. Plasma Lipoproteins: Composition, structure and biochemistry. AOSC Lipid Libr. 2014. [Google Scholar] [CrossRef]
- Rui, L. Energy metabolism in the liver. Compr. Physiol. 2014. [Google Scholar] [CrossRef] [Green Version]
- Mauvais-Jarvis, F.; Arnold, A.P.; Reue, K. A Guide for the Design of Pre-clinical Studies on Sex Differences in Metabolism. Cell Metab. 2017, 25, 1216–1230. [Google Scholar] [CrossRef] [Green Version]
- Innis, S.M.; Dyer, R. Dietary triacylglycerols with palmitic acid (16:0) in the 2-position increase 16:0 in the 2-position of plasma and chylomicron triacylglycerols, but reduce phospholipid arachidonic and docosahexaenoic acids, and alter cholesteryl ester metabolism in formula-Fed piglets. J. Nutr. 1997, 127, 1311–1319. [Google Scholar]
- Link, J.C.; Chen, X.; Arnold, A.P.; Reue, K. Metabolic impact of sex chromosomes. Mini-Rev. Adipocyte 2013, 2, 74–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugiyama, M.G.; Agellon, L.B. Sex differences in lipid metabolism and metabolic disease risk. Biochem. Cell Biol. 2012, 90, 124–141. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | MC | MT | FC | FT |
|---|---|---|---|---|
| Weight (g) | ||||
| Body weight (day 0) | 271.3 ± 17.58 a | 273.2 ± 10.65 a | 198.78 ± 6.02 b | 198.26 ± 7.73 b |
| Body weight (day 7) | 300.33 ± 18.79 a | 301.83 ± 8.57 a | 214 ± 7.72 b | 203.66 ± 5.58 b |
| Food intake | 23.00 ± 1.75 a | 22.33 ± 1.87 a | 14.60 ± 1.07 b | 14.80 ± 0.88 b |
| Liver | 12.58 ± 1.24 a | 12.72 ± 1.24 a | 8.49 ± 0.45 b | 8.97 ± 0.44 b |
| Abdominal fat | 2.03 ± 0.56 ab | 2.34 ± 0.43 a | 1.74 ± 0.28 ab | 1.43 ± 0.26 b |
| Concentration (mmol/L) | ||||
| Glucose | 6.38 a | 4.76 b | 5.52 b | 6.01 ab |
| NEFA | 0.42 | 0.39 | 0.37 | 0.34 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shah, S.; Fillier, T.; Pham, T.H.; Thomas, R.; Cheema, S.K. Intraperitoneal Administration of Short-Chain Fatty Acids Improves Lipid Metabolism of Long–Evans Rats in a Sex-Specific Manner. Nutrients 2021, 13, 892. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13030892
Shah S, Fillier T, Pham TH, Thomas R, Cheema SK. Intraperitoneal Administration of Short-Chain Fatty Acids Improves Lipid Metabolism of Long–Evans Rats in a Sex-Specific Manner. Nutrients. 2021; 13(3):892. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13030892
Chicago/Turabian StyleShah, Shrushti, Tiffany Fillier, Thu Huong Pham, Raymond Thomas, and Sukhinder Kaur Cheema. 2021. "Intraperitoneal Administration of Short-Chain Fatty Acids Improves Lipid Metabolism of Long–Evans Rats in a Sex-Specific Manner" Nutrients 13, no. 3: 892. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13030892