
The Impact of Milk Fat Globule Membrane with Exercise on Age-Related Degeneration of Neuromuscular Junctions


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Materials and Experimental Diets
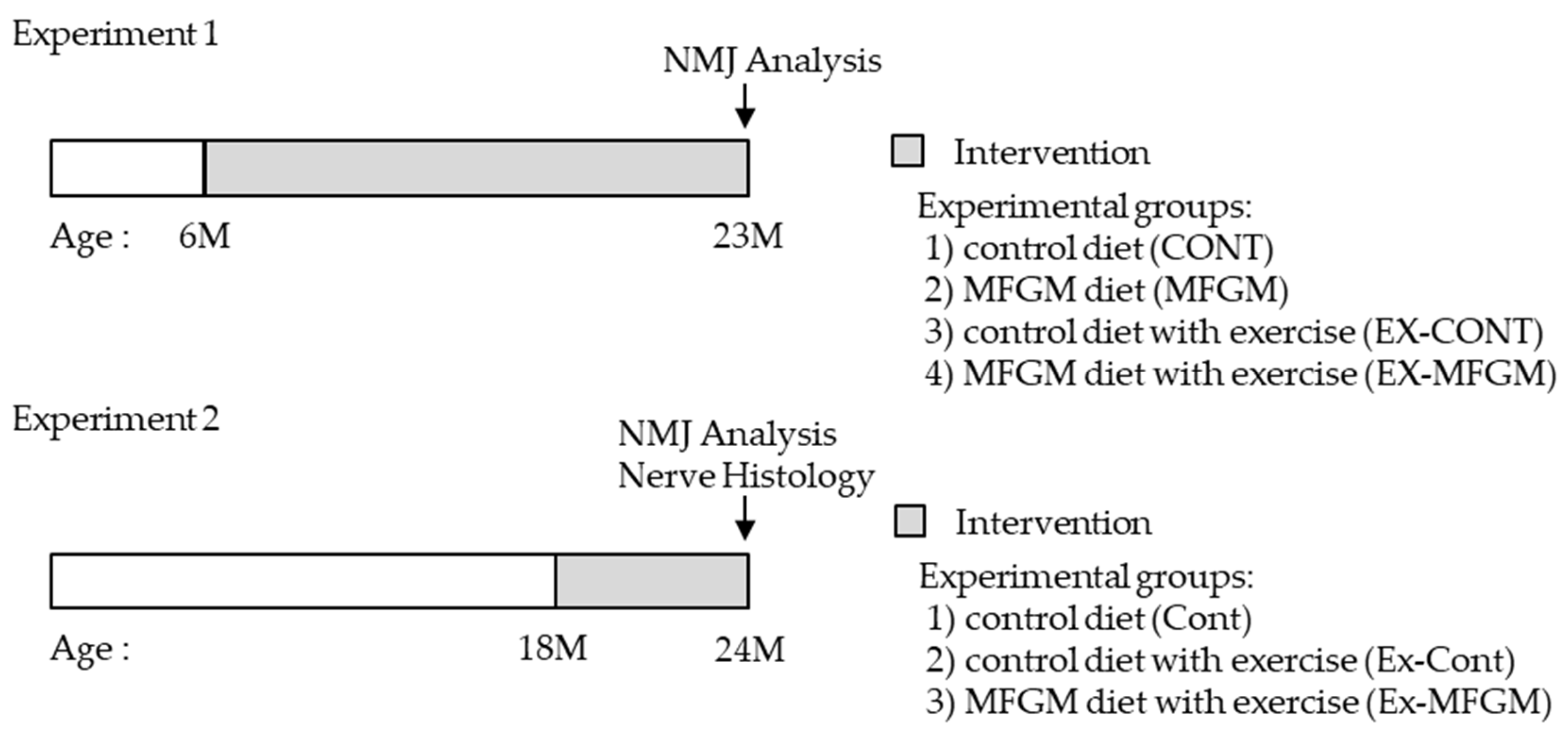
2.3. Experimental Design
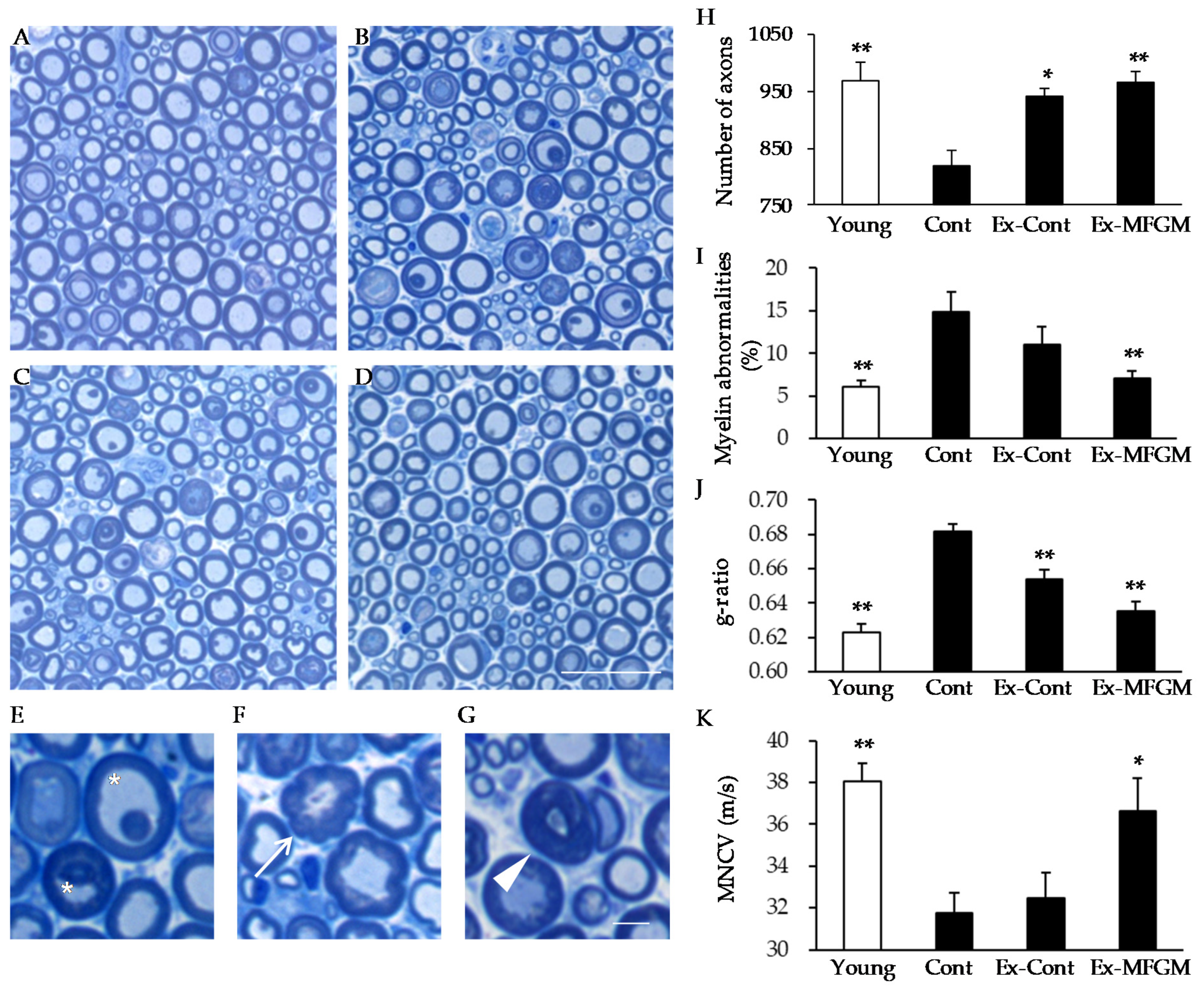
2.3.1. Experiment 1
2.3.2. Experiment 2
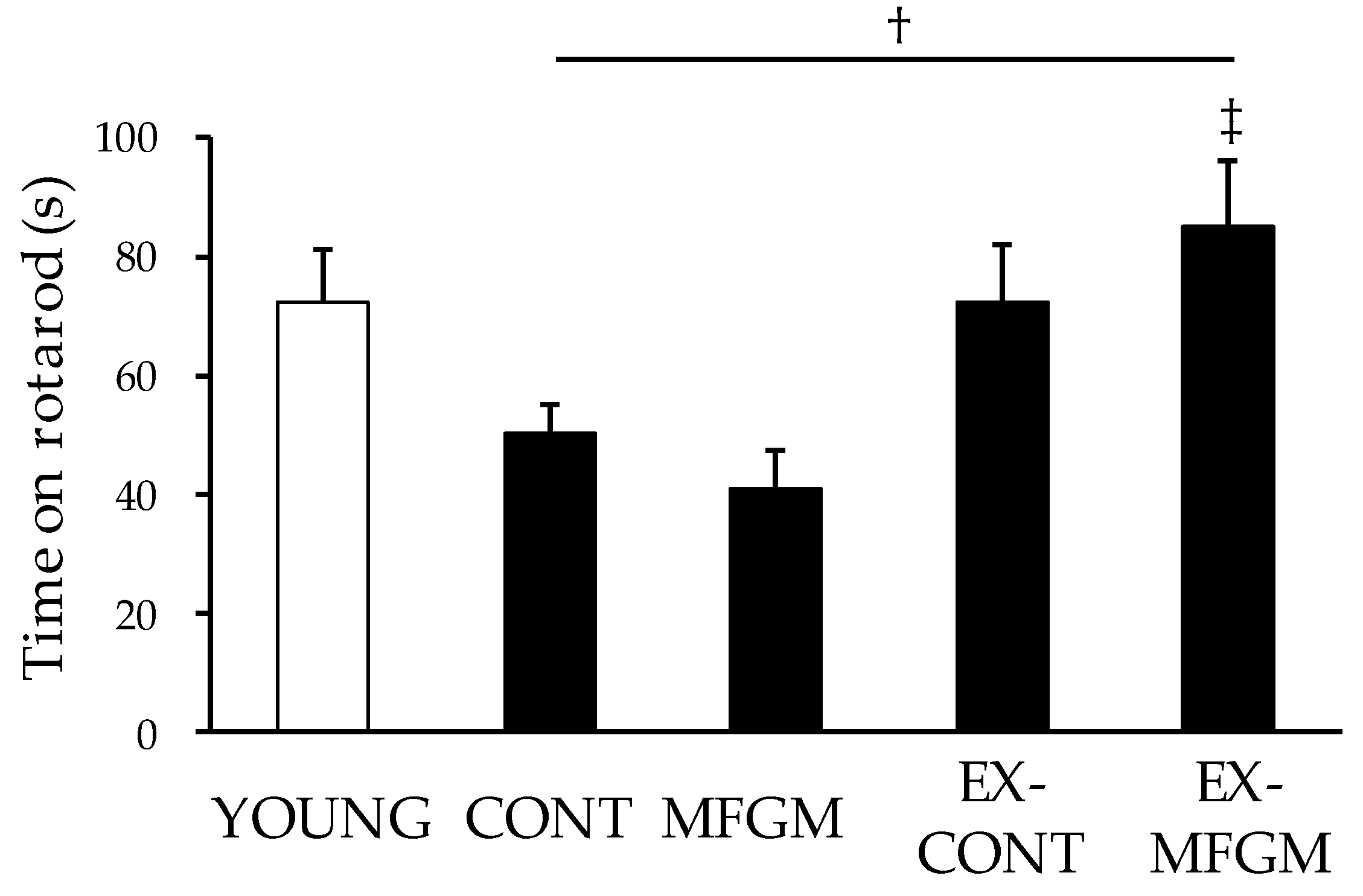
2.4. Rotarod Test
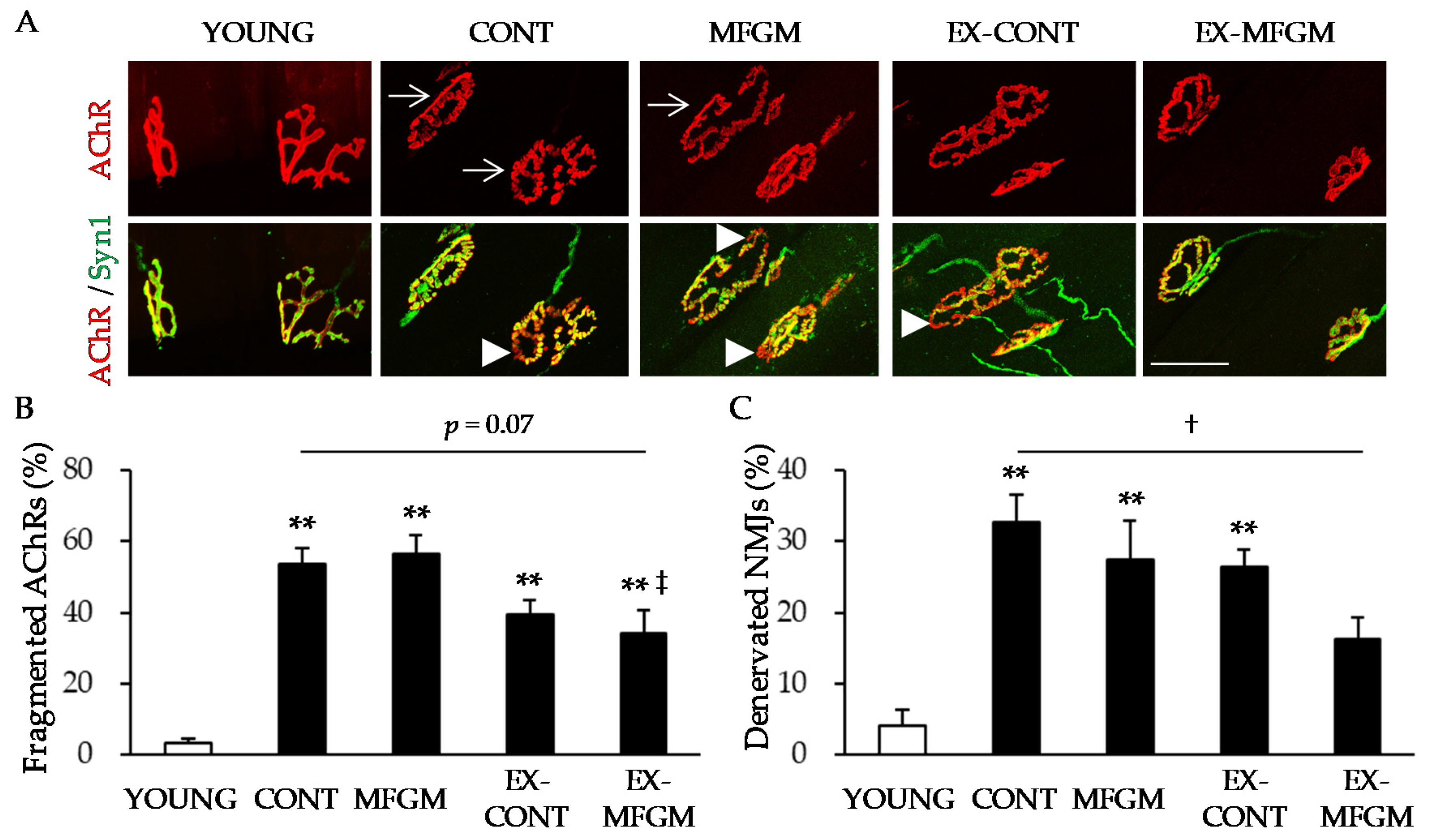
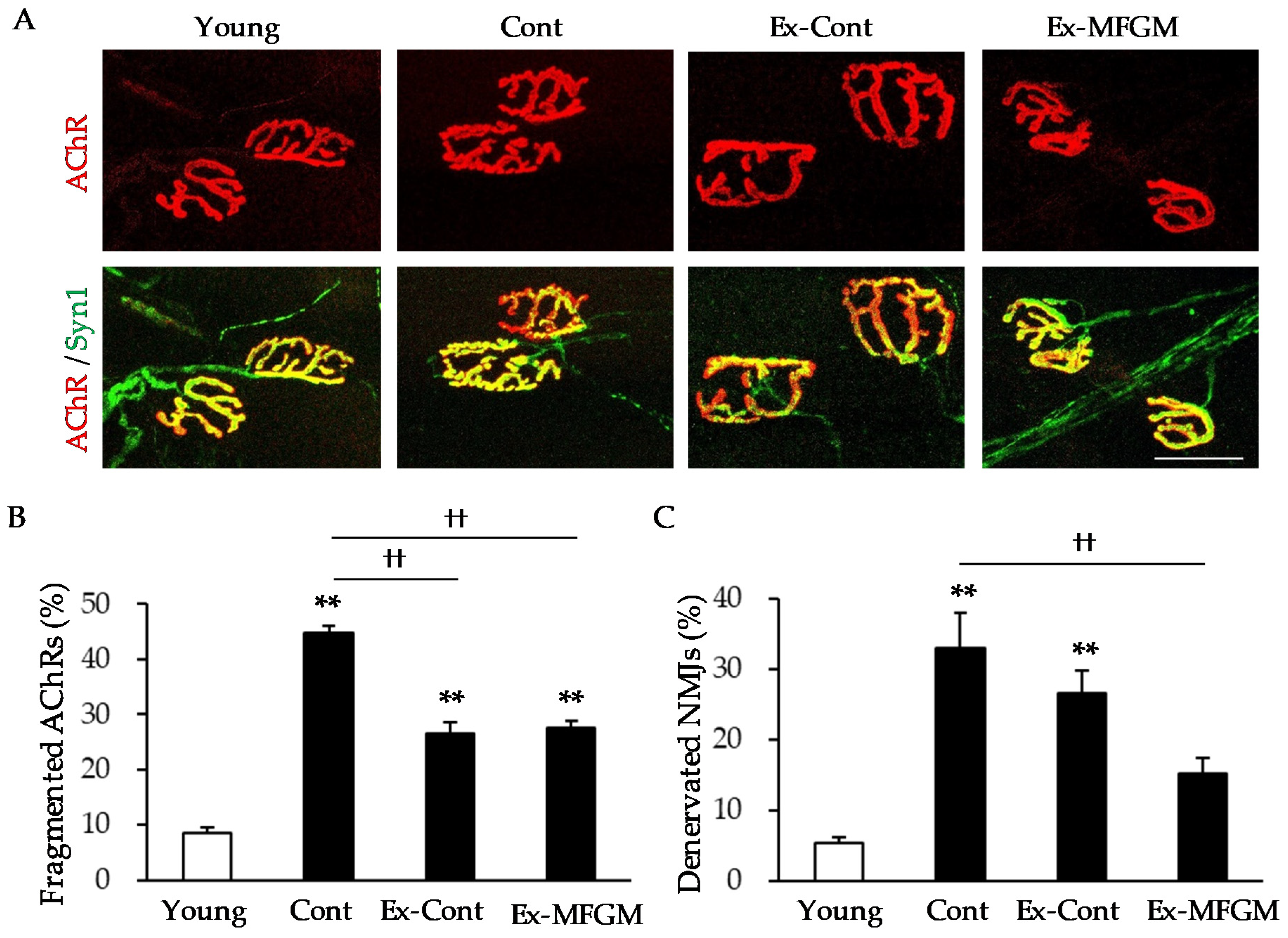
2.5. NMJ Analysis
2.6. Western Blot Analysis
2.7. Nerve Histology
2.8. Spinal Cord Histology
2.9. Motor Nerve Conduction Velocity
2.10. Statistical Analyses
3. Results
3.1. Experiment 1
3.1.1. Effects of MFGM with Exercise from a Young Age on Body and Muscle Weight
3.1.2. Effects of MFGM with Exercise from a Young Age on Motor Function
3.1.3. Effect of MFGM with Exercise from a Young Age on Age-Related NMJ Alterations
3.2. Experiment 2
3.2.1. Effect of MFGM with Exercise from an Old Age on Age-Related NMJ Alterations
3.2.2. Effect of MFGM with Exercise from an Old Age on Nerve Structure and Function
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hunter, S.K.; Pereira, H.M.; Keenan, K.G. The aging neuromuscular system and motor performance. J. Appl. Physiol. 2016, 121, 982–995. [Google Scholar] [CrossRef] [PubMed]
- Newman, A.B.; Simonsick, E.M.; Naydeck, B.L.; Boudreau, R.; Kritchevsky, S.B.; Nevitt, M.C.; Pahor, M.; Satterfield, S.; Brach, J.S.; Studenski, S.A.; et al. Association of Long-Distance Corridor Walk Performance With Mortality, Cardiovascular Disease, Mobility Limitation, and Disability. JAMA 2006, 295, 2018–2026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doherty, T.J. Invited Review: Aging and sarcopenia. J. Appl. Physiol. 2003, 95, 1717–1727. [Google Scholar] [CrossRef] [Green Version]
- Roubenoff, R.; Hughes, V.A. Sarcopenia: Current Concepts. J. Gerontol. Ser. A Boil. Sci. Med Sci. 2000, 55, M716–M724. [Google Scholar] [CrossRef] [Green Version]
- Newman, A.B.; Haggerty, C.L.; Goodpaster, B.; Harris, T.; Kritchevsky, S.; Nevitt, M.; Miles, T.; Visser, M. The Health Aging and Body Compositi Strength and Muscle Quality in a Well-Functioning Cohort of Older Adults: The Health, Aging and Body Composition Study. J. Am. Geriatr. Soc. 2003, 51, 323–330. [Google Scholar] [CrossRef]
- Goodpaster, B.H.; Park, S.W.; Harris, T.B.; Kritchevsky, S.B.; Nevitt, M.; Schwartz, A.V.; Simonsick, E.M.; Tylavsky, F.A.; Visser, M.; Newman, A.B.; et al. The Loss of Skeletal Muscle Strength, Mass, and Quality in Older Adults: The Health, Aging and Body Composition Study. J. Gerontol. Ser. A Boil. Sci. Med Sci. 2006, 61, 1059–1064. [Google Scholar] [CrossRef]
- Chai, R.J.; Vukovic, J.; Dunlop, S.; Grounds, M.; Shavlakadze, T. Striking Denervation of Neuromuscular Junctions without Lumbar Motoneuron Loss in Geriatric Mouse Muscle. PLoS ONE 2011, 6, e28090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valdez, G.; Tapia, J.C.; Kang, H.; Clemenson, G.D.; Gage, F.H.; Lichtman, J.W.; Sanes, J.R. Attenuation of age-related changes in mouse neuromuscular synapses by caloric restriction and exercise. Proc. Natl. Acad. Sci. USA 2010, 107, 14863–14868. [Google Scholar] [CrossRef] [Green Version]
- Oda, K. Age changes of motor innervation and acetylcholine receptor distribution on human skeletal muscle fibres. J. Neurol. Sci. 1984, 66, 327–338. [Google Scholar] [CrossRef]
- Wokke, J.; Jennekens, F.; Oord, C.V.D.; Veldman, H.; Smit, L.; Leppink, G. Morphological changes in the human end plate with age. J. Neurol. Sci. 1990, 95, 291–310. [Google Scholar] [CrossRef]
- Chung, T.; Park, J.S.; Kim, S.; Montes, N.; Walston, J.; Höke, A. Evidence for dying-back axonal degeneration in age-associated skeletal muscle decline. Muscle Nerve 2017, 55, 894–901. [Google Scholar] [CrossRef]
- Kurokawa, K.; Mimori, Y.; Tanaka, E.; Kohriyama, T.; Nakamura, S. Age-Related Change in Peripheral Nerve Conduction: Compound Muscle Action Potential Duration and Dispersion. Gerontology 1999, 45, 168–173. [Google Scholar] [CrossRef]
- Kougias, D.G.; Das, T.; Perez, A.B.; Pereira, S.L. A role for nutritional intervention in addressing the aging neuromuscular junction. Nutr. Res. 2018, 53, 1–14. [Google Scholar] [CrossRef]
- Stockinger, J.; Maxwell, N.; Shapiro, D.; Decabo, R.; Valdez, G. Caloric Restriction Mimetics Slow Aging of Neuromuscular Synapses and Muscle Fibers. J. Gerontol. Ser. A Boil. Sci. Med Sci. 2017, 73, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Cavaletto, M.; Giuffrida, M.G.; Conti, A. Milk Fat Globule Membrane Components–A Proteomic Approach. Adv. Exp. Med. Biol. 2008, 606, 129–141. [Google Scholar] [CrossRef]
- Kim, H.; Suzuki, T.; Kim, M.; Kojima, N.; Ota, N.; Shimotoyodome, A.; Hase, T.; Hosoi, E.; Yoshida, H. Effects of Exercise and Milk Fat Globule Membrane (MFGM) Supplementation on Body Composition, Physical Function, and Hematological Parameters in Community-Dwelling Frail Japanese Women: A Randomized Double Blind, Placebo-Controlled, Follow-Up Trial. PLoS ONE 2015, 10, e0116256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minegishi, Y.; Ota, N.; Soga, S.; Shimotoyodome, A. Effects of Nutritional Supplementation with Milk Fat Globule Membrane on Physical and Muscle Function in Healthy Adults Aged 60 and Over with Semiweekly Light Exercise: A Randomized Double-Blind, Placebo-Controlled Pilot Trial. J. Nutr. Sci. Vitaminol. 2016, 62, 409–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshinaka, Y.; Soga, S.; Ota, N.; Yokoyama, K.; Yamada, Y.; Kimura, M. Light rhythmic exercise with dietary milk fat globule membrane improves physical fitness in an elderly Japanese population: A double-blind randomized placebo-controlled trial. Biosci. Biotechnol. Biochem. 2018, 82, 677–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ota, N.; Soga, S.; Hase, T.; Shimotoyodome, A. Daily consumption of milk fat globule membrane plus habitual exercise improves physical performance in healthy middle-aged adults. SpringerPlus 2015, 4, 120. [Google Scholar] [CrossRef] [Green Version]
- Haramizu, S.; Mori, T.; Yano, M.; Ota, N.; Hashizume, K.; Otsuka, A.; Hase, T.; Shimotoyodome, A. Habitual exercise plus dietary supplementation with milk fat globule membrane improves muscle function deficits via neuromuscular development in senescence-accelerated mice. SpringerPlus 2014, 3, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Haramizu, S.; Ota, N.; Otsuka, A.; Hashizume, K.; Sugita, S.; Hase, T.; Murase, T.; Shimotoyodome, A. Dietary milk fat globule membrane improves endurance capacity in mice. Am. J. Physiol. Integr. Comp. Physiol. 2014, 307, R1009–R1017. [Google Scholar] [CrossRef] [Green Version]
- Yano, M.; Haramizu, S.; Ota, N.; Minegishi, Y.; Shimotoyodome, A. Continuous Supplementation of Milk Fat Globule Membrane with Habitual Exercise from a Young Age Improves Motor Coordination and Skeletal Muscle Function in Aged Mice. J. Nutr. Sci. Vitaminol. 2019, 65, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Inoue, A.; Setoguchi, K.; Matsubara, Y.; Okada, K.; Sato, N.; Iwakura, Y.; Higuchi, O.; Yamanashi, Y. Dok-7 Activates the Muscle Receptor Kinase MuSK and Shapes Synapse Formation. Sci. Signal. 2009, 2, ra7. [Google Scholar] [CrossRef]
- Okada, K.; Inoue, A.; Okada, M.; Murata, Y.; Kakuta, S.; Jigami, T.; Kubo, S.; Shiraishi, H.; Eguchi, K.; Motomura, M.; et al. The Muscle Protein Dok-7 Is Essential for Neuromuscular Synaptogenesis. Science 2006, 312, 1802–1805. [Google Scholar] [CrossRef] [PubMed]
- Yano, M.; Minegishi, Y.; Sugita, S.; Ota, N. Milk fat globule membrane supplementation with voluntary running exercise attenuates age-related motor dysfunction by suppressing neuromuscular junction abnormalities in mice. Exp. Gerontol. 2017, 97, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Sugita, S.; Fleming, L.L.; Wood, C.; Vaughan, S.; Gomes, M.P.S.M.; Camargo, W.; Naves, L.A.; Prado, V.; Prado, M.A.M.; Guatimosim, C.; et al. VAChT overexpression increases acetylcholine at the synaptic cleft and accelerates aging of neuromuscular junctions. Skelet. Muscle 2016, 6, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Yuan, X.; Klein, D.; Kerscher, S.; West, B.L.; Weis, J.; Katona, I.; Martini, R. Macrophage Depletion Ameliorates Peripheral Neuropathy in Aging Mice. J. Neurosci. 2018, 38, 4610–4620. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Sugita, S.; Tokunaga, T.; Minegishi, Y.; Ota, N. TRPM8-mediated cutaneous stimulation modulates motor neuron activity during treadmill stepping in mice. J. Physiol. Sci. 2019, 69, 931–938. [Google Scholar] [CrossRef]
- Mizukami, H.; Ogasawara, S.; Yamagishi, S.-I.; Takahashi, K.; Yagihashi, S. Methylcobalamin effects on diabetic neuropathy and nerve protein kinase C in rats. Eur. J. Clin. Investig. 2010, 41, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Tsuboi, K.; Mizukami, H.; Inaba, W.; Baba, M.; Yagihashi, S. The dipeptidyl peptidase IV inhibitor vildagliptin suppresses development of neuropathy in diabetic rodents: Effects on peripheral sensory nerve function, structure and molecular changes. J. Neurochem. 2016, 136, 859–870. [Google Scholar] [CrossRef] [Green Version]
- Yamagishi, S.-I.; Ogasawara, S.; Mizukami, H.; Yajima, N.; Wada, R.-I.; Sugawara, A.; Yagihashi, S. Correction of protein kinase C activity and macrophage migration in peripheral nerve by pioglitazone, peroxisome proliferator activated-γ-ligand, in insulin-deficient diabetic rats. J. Neurochem. 2007, 104, 491–499. [Google Scholar] [CrossRef]
- Ueta, R.; Sugita, S.; Minegishi, Y.; Shimotoyodome, A.; Ota, N.; Ogiso, N.; Eguchi, T.; Yamanashi, Y. DOK7 Gene Therapy Enhances Neuromuscular Junction Innervation and Motor Function in Aged Mice. iScience 2020, 23, 101385. [Google Scholar] [CrossRef] [PubMed]
- Arimura, S.; Okada, T.; Tezuka, T.; Chiyo, T.; Kasahara, Y.; Yoshimura, T.; Motomura, M.; Yoshida, N.; Beeson, D.; Takeda, S.; et al. DOK7 gene therapy benefits mouse models of diseases characterized by defects in the neuromuscular junction. Science 2014, 345, 1505–1508. [Google Scholar] [CrossRef] [PubMed]
- Flurkey, K.; Currer, J.; Harrison, D. The Mouse in Aging Research; Elsevier: Amsterdam, The Netherlands, 2007; pp. 637–672. [Google Scholar]
- Cheng, A.; Morsch, M.; Murata, Y.; Ghazanfari, N.; Reddel, S.; Phillips, W. Sequence of Age-Associated Changes to the Mouse Neuromuscular Junction and the Protective Effects of Voluntary Exercise. PLoS ONE 2013, 8, e67970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slater, C.R. ‘Fragmentation’ of NMJs: A sign of degeneration or regeneration? A long journey with many junctions. Neuroscience 2020, 439, 28–40. [Google Scholar] [CrossRef]
- Willadt, S.; Nash, M.; Slater, C. Age-related changes in the structure and function of mammalian neuromuscular junctions. Ann. N. Y. Acad. Sci. 2017, 1412, 41–53. [Google Scholar] [CrossRef]
- Willadt, S.; Nash, M.; Slater, C.R. Age-related fragmentation of the motor endplate is not associated with impaired neuromuscular transmission in the mouse diaphragm. Sci. Rep. 2016, 6, 24849. [Google Scholar] [CrossRef]
- Valdez, G.; Tapia, J.C.; Lichtman, J.W.; Fox, M.A.; Sanes, J.R. Shared Resistance to Aging and ALS in Neuromuscular Junctions of Specific Muscles. PLoS ONE 2012, 7, e34640. [Google Scholar] [CrossRef] [Green Version]
- Pandya, A.V.; Patani, R. Decoding the relationship between ageing and amyotrophic lateral sclerosis: A cellular perspective. Brain 2020, 143, 1057–1072. [Google Scholar] [CrossRef]
- Badawi, Y.; Nishimune, H. Impairment Mechanisms and Intervention Approaches for Aged Human Neuromuscular Junctions. Front. Mol. Neurosci. 2020, 13, 568426. [Google Scholar] [CrossRef]
- Doherty, T.J.; Vandervoort, A.A.; Taylor, A.W.; Brown, W.F. Effects of motor unit losses on strength in older men and women. J. Appl. Physiol. 1993, 74, 868–874. [Google Scholar] [CrossRef] [PubMed]
- Tomlinson, B.; Irving, D. The numbers of limb motor neurons in the human lumbosacral cord throughout life. J. Neurol. Sci. 1977, 34, 213–219. [Google Scholar] [CrossRef]
- Jacob, J.M. Lumbar motor neuron size and number is affected by age in male F344 rats. Mech. Ageing Dev. 1998, 106, 205–216. [Google Scholar] [CrossRef]
- Hashizume, K.; Kanda, K. Differential effects of aging on motoneurons and peripheral nerves innervating the hindlimb and forelimb muscles of rats. Neurosci. Res. 1995, 22, 189–196. [Google Scholar] [CrossRef]
- LaFratta, C.W.; Canestrari, R. A comparison of sensory and motor nerve conduction velocities as related to age. Arch. Phys. Med. Rehabil. 1966, 47, 286–290. [Google Scholar] [PubMed]
- LaFratta, C.W.; Smith, O.H. A study of the relationship of motor nerve conduction velocity in the adult to age, sex, and handedness. Arch. Phys. Med. Rehabil. 1964, 45, 407–412. [Google Scholar]
- Rivner, M.H.; Swift, T.R.; Malik, K. Influence of age and height on nerve conduction. Muscle Nerve 2001, 24, 1134–1141. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, R.; Walsh, M.; Singh, R.; Rodriguez, K.; Gao, X.; Rahman, M.; Chaudhuri, A.; Bhattacharya, A. Oxidative damage to myelin proteins accompanies peripheral nerve motor dysfunction in aging C57BL/6 male mice. J. Neurol. Sci. 2016, 370, 47–52. [Google Scholar] [CrossRef]
- Walsh, M.E.; Sloane, L.B.; Fischer, K.E.; Austad, S.N.; Richardson, A.; Van Remmen, H. Use of Nerve Conduction Velocity to Assess Peripheral Nerve Health in Aging Mice. J. Gerontol. Ser. A Boil. Sci. Med Sci. 2014, 70, 1312–1319. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, V.S.; White, Z.; Terrill, J.R.; Hodgetts, S.; Fitzgerald, M.; Shavlakadze, T.; Harvey, A.R.; Grounds, M.D. Resistance wheel exercise from mid-life has minimal effect on sciatic nerves from old mice in which sarcopenia was prevented. Biogerontology 2017, 18, 769–790. [Google Scholar] [CrossRef]
- Waworuntu, R.V.; Hanania, T.; Boikess, S.R.; Rex, C.S.; Berg, B.M. Early life diet containing prebiotics and bioactive whey protein fractions increased dendritic spine density of rat hippocampal neurons. Int. J. Dev. Neurosci. 2016, 55, 28–33. [Google Scholar] [CrossRef] [Green Version]
- Timby, N.; Domellöf, E.; Hernell, O.; Lönnerdal, B.; Domellöf, M. Neurodevelopment, nutrition, and growth until 12 mo of age in infants fed a low-energy, low-protein formula supplemented with bovine milk fat globule membranes: A randomized controlled trial. Am. J. Clin. Nutr. 2014, 99, 860–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oshida, K.; Shimizu, T.; Takase, M.; Tamura, Y.; Shimizu, T.; Yamashiro, Y. Effects of Dietary Sphingomyelin on Central Nervous System Myelination in Developing Rats. Pediatr. Res. 2003, 53, 589–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Lee, Y.I.; Thompson, W.J. Changes in Aging Mouse Neuromuscular Junctions Are Explained by Degeneration and Regeneration of Muscle Fiber Segments at the Synapse. J. Neurosci. 2011, 31, 14910–14919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brink, L.R.; Gueniot, J.P.; Lönnerdal, B. Effects of milk fat globule membrane and its various components on neurologic development in a postnatal growth restriction rat model. J. Nutr. Biochem. 2019, 69, 163–171. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | YOUNG | CONT | MFGM | EX-CONT | EX-MFGM |
|---|---|---|---|---|---|
| Body weight (g) | 30.6 ± 0.6 | 34.8 ± 1.1 | 36.5 ± 1.3 * | 33.9 ± 0.9 | 36.0 ± 1.7 * |
| EDL muscle (mg) | 14.6 ± 0.5 | 12.1 ± 0.4 * | 11.7 ± 0.7 ** | 12.1 ± 0.5 * | 12.7 ± 0.2 |
| Soleus muscle (mg) | 10.6 ± 1.3 | 10.1 ± 0.6 | 10.0 ± 0.4 | 13.0 ± 0.7 | 12.7 ± 0.9 |
| TA muscle (mg) | 53.3 ± 2.5 | 53.3 ± 2.6 | 50.2 ± 4.2 | 51.4 ± 3.4 | 54.0 ± 1.5 |
| Gastro muscle (mg) | 143.3 ± 4.7 | 127.3 ± 5.5 | 127.3 ± 11.4 | 134.7 ± 1.5 | 141.2 ± 5.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sugita, S.; Tamura, K.; Yano, M.; Minegishi, Y.; Ota, N. The Impact of Milk Fat Globule Membrane with Exercise on Age-Related Degeneration of Neuromuscular Junctions. Nutrients 2021, 13, 2310. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13072310
Sugita S, Tamura K, Yano M, Minegishi Y, Ota N. The Impact of Milk Fat Globule Membrane with Exercise on Age-Related Degeneration of Neuromuscular Junctions. Nutrients. 2021; 13(7):2310. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13072310
Chicago/Turabian StyleSugita, Satoshi, Kotaro Tamura, Michiko Yano, Yoshihiko Minegishi, and Noriyasu Ota. 2021. "The Impact of Milk Fat Globule Membrane with Exercise on Age-Related Degeneration of Neuromuscular Junctions" Nutrients 13, no. 7: 2310. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13072310