
Gut Microbial Signatures of Distinct Trimethylamine N-Oxide Response to Raspberry Consumption

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Study Design and Data Collection
2.2. TMAO Profiling and Patient Stratification
2.3. Fecal Metagenomics
2.4. Statistical Analyses
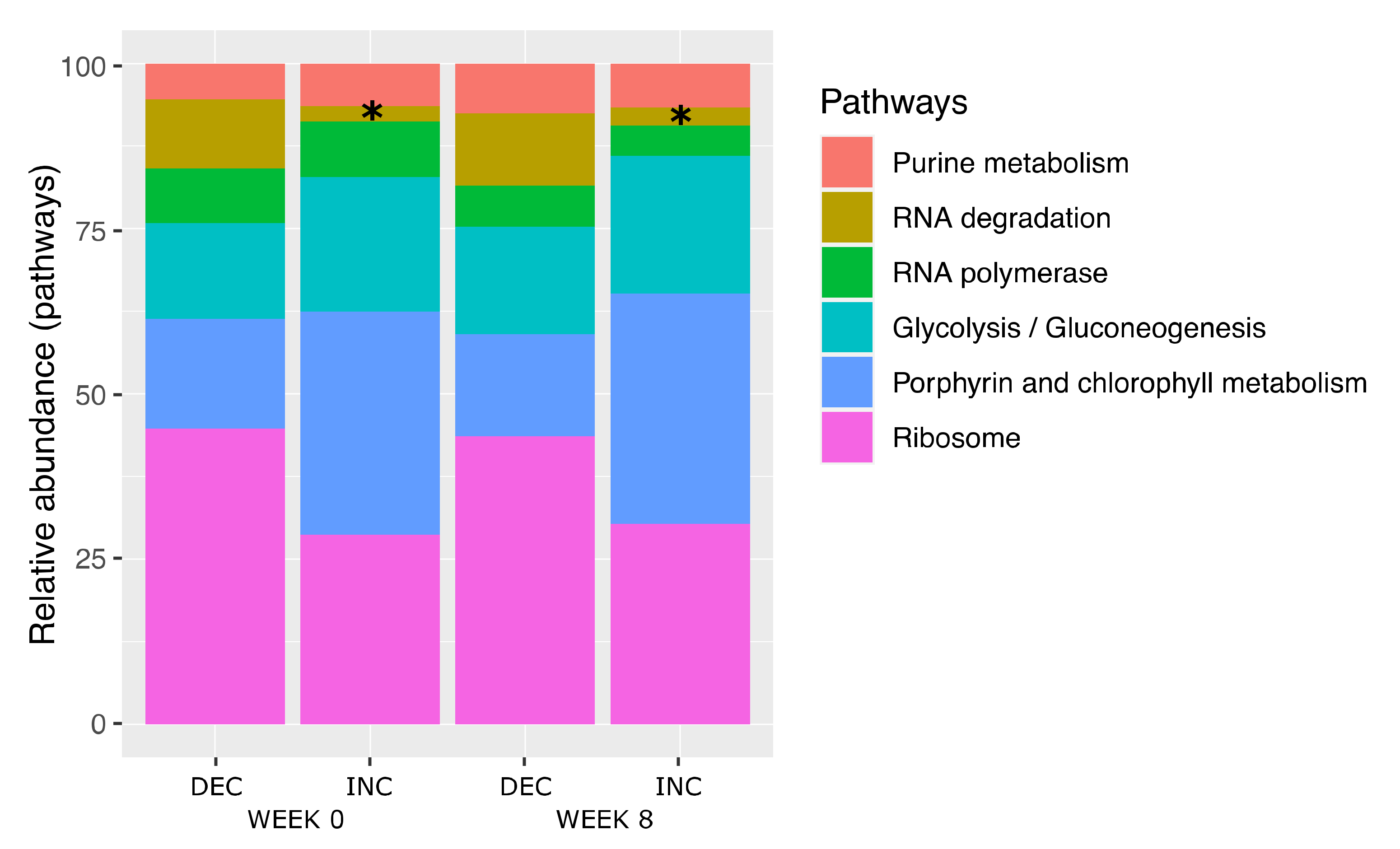
3. Results
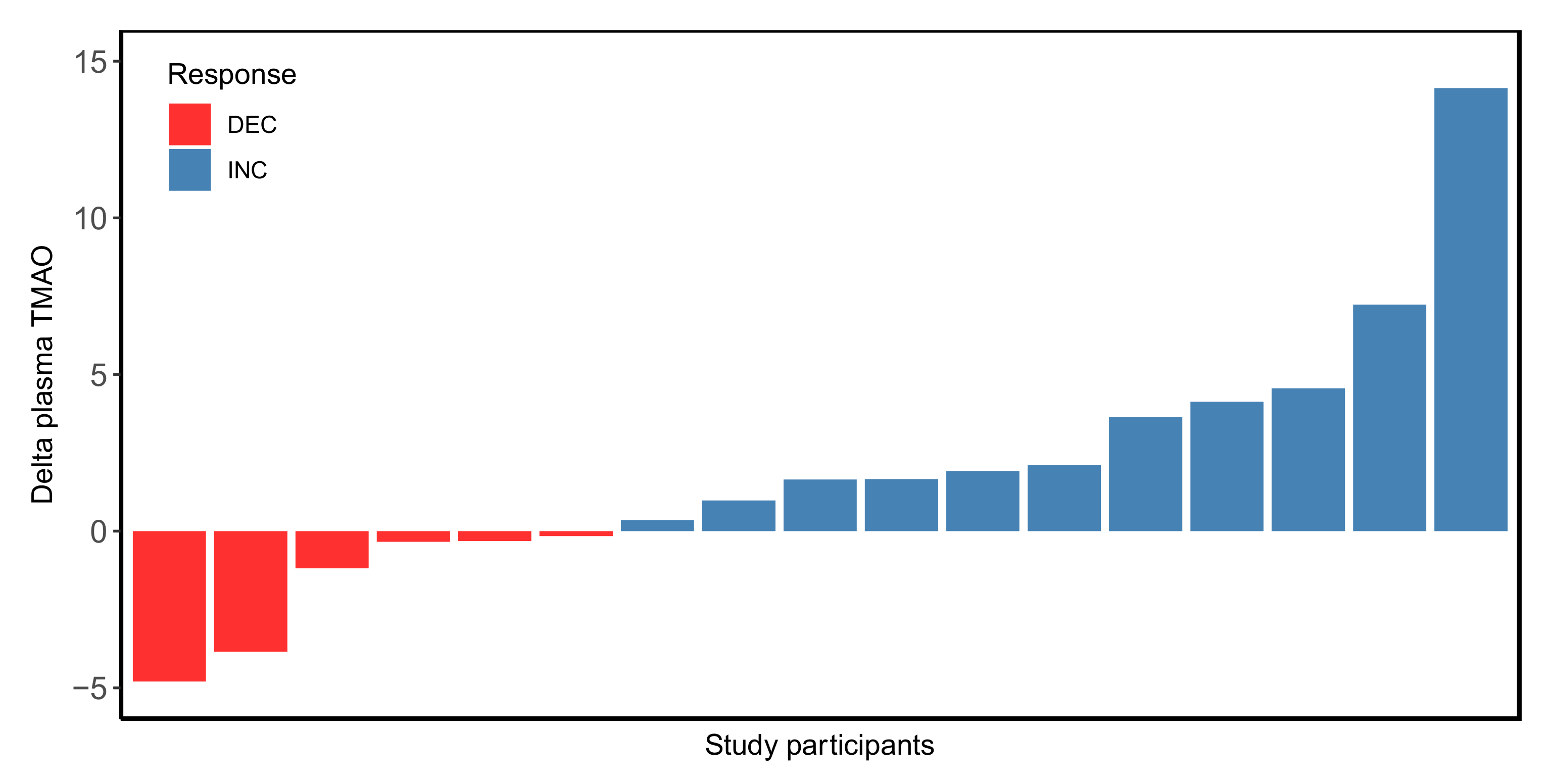
3.1. Inter-Individual Variability in Plasma TMAO Levels in Pre- versus Post-Rb Consumption
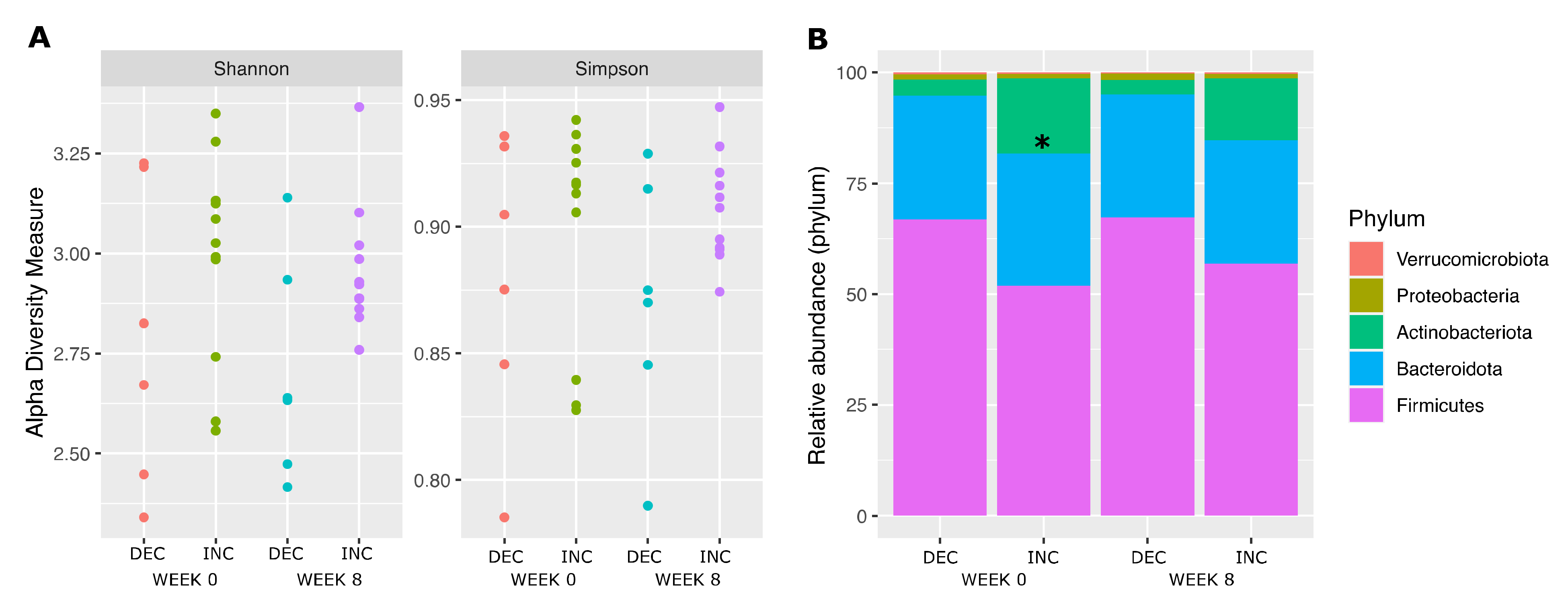
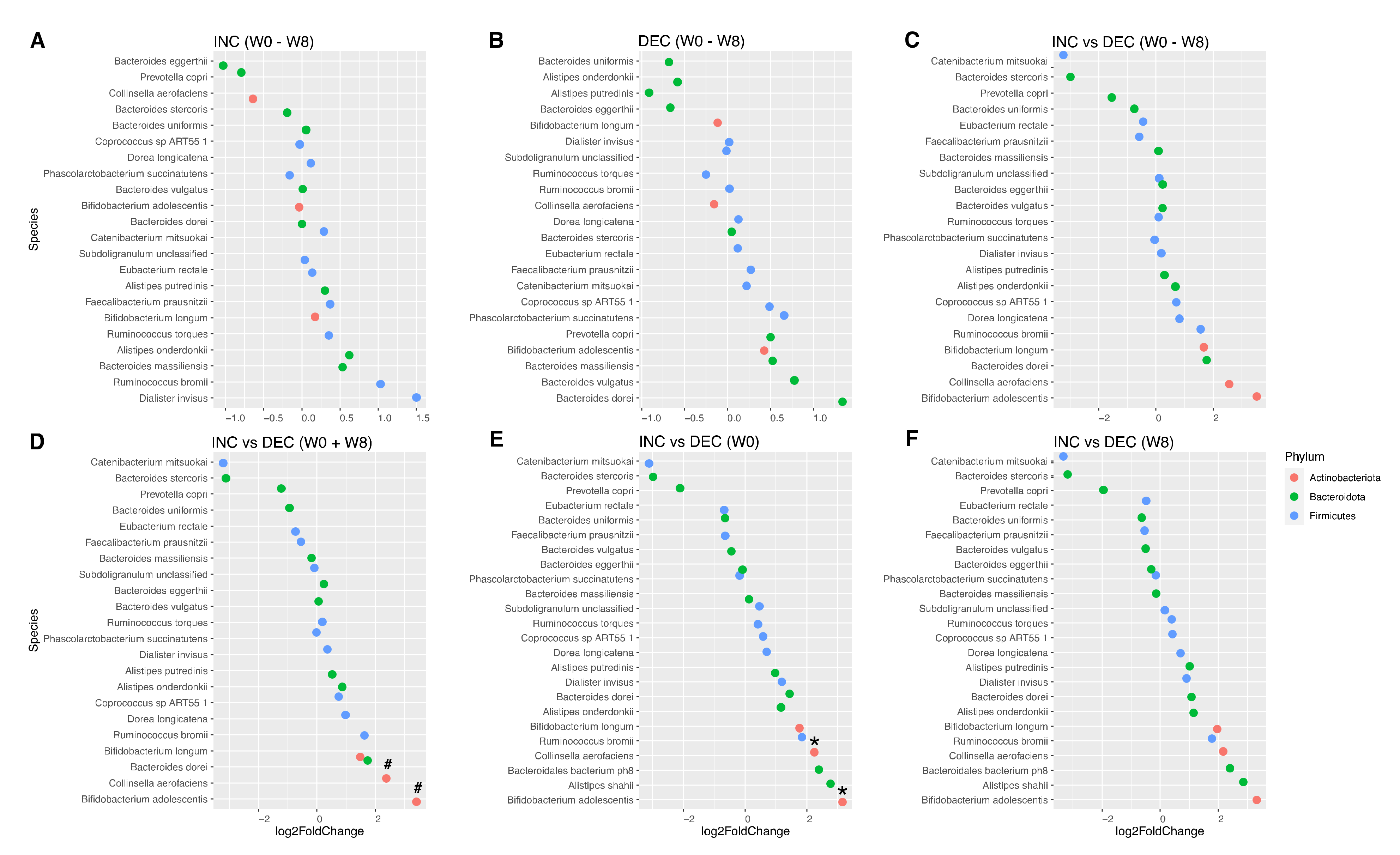
3.2. Gut Microbiota Composition According to the Plasma TMAO Response to the Rb Consumption
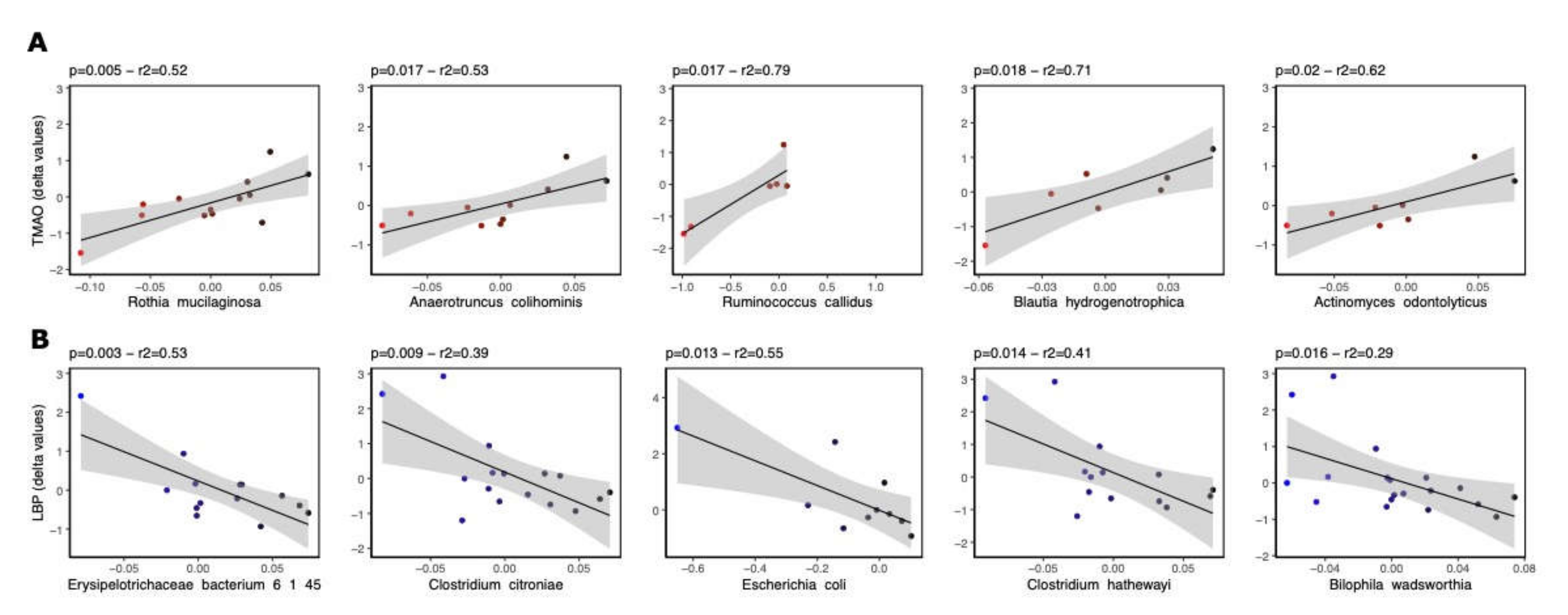
3.3. Bacterial Species Associated with LBP and TMAO following Rb Consumption
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stremmel, W.; Schmidt, K.V.; Schuhmann, V.; Kratzer, F.; Garbade, S.F.; Langhans, C.D.; Fricker, G.; Okun, J.G. Blood Trimethylamine-N-Oxide Originates from Microbiota Mediated Breakdown of Phosphatidylcholine and Absorption from Small Intestine. PLoS ONE 2017, 12, e0170742. [Google Scholar] [CrossRef] [PubMed]
- Koeth, R.A.; Wang, Z.; Levison, B.S.; Buffa, J.A.; Org, E.; Sheehy, B.T.; Britt, E.B.; Fu, X.; Wu, Y.; Li, L.; et al. Intestinal microbiota metabolism of L-carnitine, a nutrient in red meat, promotes atherosclerosis. Nat. Med. 2013, 19, 576–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.J.; Shu, X.O.; Herrington, D.M.; Moore, S.C.; Meyer, K.A.; Ose, J.; Menni, C.; Palmer, N.D.; Eliassen, H.; Harada, S.; et al. Circulating trimethylamine N-oxide in association with diet and cardiometabolic biomarkers: An international pooled analysis. Am. J. Clin. Nutr. 2021, 113, 1145–1156. [Google Scholar] [CrossRef] [PubMed]
- Franck, M.; de Toro-Martin, J.; Garneau, V.; Guay, V.; Kearney, M.; Pilon, G.; Roy, D.; Couture, P.; Couillard, C.; Marette, A.; et al. Effects of Daily Raspberry Consumption on Immune-Metabolic Health in Subjects at Risk of Metabolic Syndrome: A Randomized Controlled Trial. Nutrients 2020, 12, 3858. [Google Scholar] [CrossRef] [PubMed]
- Scarsella, C.; Alméras, N.; Mauriège, P.; Blanchet, C.; Sauvé, L.; Dewailly, E.; Bergeron, J.; Després, J.P. Prevalence of metabolic alterations predictive of cardiovascular disease risk in the Québec population. Can. J. Cardiol. 2003, 19, 51–57. [Google Scholar]
- Chong, J.; Xia, J. MetaboAnalystR: An R package for flexible and reproducible analysis of metabolomics data. Bioinformatics 2018, 34, 4313–4314. [Google Scholar] [CrossRef] [Green Version]
- Franck, M.; de Toro-Martin, J.; Varin, T.V.; Garneau, V.; Pilon, G.; Roy, D.; Couture, P.; Couillard, C.; Marette, A.; Vohl, M.C. Raspberry consumption: Identification of distinct immune-metabolic response profiles by whole blood transcriptome profiling. J. Nutr. Biochem. 2022, 101, 108946. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Dadi, T.H.; Renard, B.Y.; Wieler, L.H.; Semmler, T.; Reinert, K. SLIMM: Species level identification of microorganisms from metagenomes. PeerJ 2017, 5, e3138. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franzosa, E.A.; McIver, L.J.; Rahnavard, G.; Thompson, L.R.; Schirmer, M.; Weingart, G.; Lipson, K.S.; Knight, R.; Caporaso, J.G.; Segata, N.; et al. Species-level functional profiling of metagenomes and metatranscriptomes. Nat. Methods 2018, 15, 962–968. [Google Scholar] [CrossRef] [PubMed]
- The R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 11 December 2021).
- Istas, G.; Feliciano, R.P.; Weber, T.; Garcia-Villalba, R.; Tomas-Barberan, F.; Heiss, C.; Rodriguez-Mateos, A. Plasma urolithin metabolites correlate with improvements in endothelial function after red raspberry consumption: A double-blind randomized controlled trial. Arch. Biochem. Biophys. 2018, 651, 43–51. [Google Scholar] [CrossRef] [PubMed]
- O’Donovan, A.N.; Herisson, F.M.; Fouhy, F.; Ryan, P.M.; Whelan, D.; Johnson, C.N.; Cluzel, G.; Ross, R.P.; Stanton, C.; Caplice, N.M. Gut microbiome of a porcine model of metabolic syndrome and HF-pEF. Am. J. Physiol. Heart Circ. Physiol. 2020, 318, H590–H603. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.; Mu, C.; He, X.; Zheng, K.; Guo, H.; Zhu, W. Characteristics of gut microbiota and its response to a Chinese Herbal Formula in elder patients with metabolic syndrome. Drug Discov. Ther. 2018, 12, 161–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nirmalkar, K.; Murugesan, S.; Pizano-Zarate, M.L.; Villalobos-Flores, L.E.; Garcia-Gonzalez, C.; Morales-Hernandez, R.M.; Nunez-Hernandez, J.A.; Hernandez-Quiroz, F.; Romero-Figueroa, M.D.S.; Hernandez-Guerrero, C.; et al. Gut Microbiota and Endothelial Dysfunction Markers in Obese Mexican Children and Adolescents. Nutrients 2018, 10, 2009. [Google Scholar] [CrossRef] [Green Version]
- Karlsson, F.H.; Fak, F.; Nookaew, I.; Tremaroli, V.; Fagerberg, B.; Petranovic, D.; Backhed, F.; Nielsen, J. Symptomatic atherosclerosis is associated with an altered gut metagenome. Nat. Commun. 2012, 3, 1245. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Zhao, W.; Liu, X.; Cheng, L. Metagenomic analysis of the gut microbiome in atherosclerosis patients identify cross-cohort microbial signatures and potential therapeutic target. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2020, 34, 14166–14181. [Google Scholar] [CrossRef]
- Fu, B.C.; Hullar, M.A.J.; Randolph, T.W.; Franke, A.A.; Monroe, K.R.; Cheng, I.; Wilkens, L.R.; Shepherd, J.A.; Madeleine, M.M.; Le Marchand, L.; et al. Associations of plasma trimethylamine N-oxide, choline, carnitine, and betaine with inflammatory and cardiometabolic risk biomarkers and the fecal microbiome in the Multiethnic Cohort Adiposity Phenotype Study. Am. J. Clin. Nutr. 2020, 111, 1226–1234. [Google Scholar] [CrossRef]
- Devkota, S.; Wang, Y.; Musch, M.W.; Leone, V.; Fehlner-Peach, H.; Nadimpalli, A.; Antonopoulos, D.A.; Jabri, B.; Chang, E.B. Dietary-fat-induced taurocholic acid promotes pathobiont expansion and colitis in Il10-/- mice. Nature 2012, 487, 104–108. [Google Scholar] [CrossRef] [Green Version]
- Mollar, A.; Marrachelli, V.G.; Nunez, E.; Monleon, D.; Bodi, V.; Sanchis, J.; Navarro, D.; Nunez, J. Bacterial metabolites trimethylamine N-oxide and butyrate as surrogates of small intestinal bacterial overgrowth in patients with a recent decompensated heart failure. Sci. Rep. 2021, 11, 6110. [Google Scholar] [CrossRef] [PubMed]
- Mikolasevic, I.; Delija, B.; Mijic, A.; Stevanovic, T.; Skenderevic, N.; Sosa, I.; Krznaric-Zrnic, I.; Abram, M.; Krznaric, Z.; Domislovic, V.; et al. Small intestinal bacterial overgrowth and non-alcoholic fatty liver disease diagnosed by transient elastography and liver biopsy. Int. J. Clin. Pract. 2021, 75, e13947. [Google Scholar] [CrossRef] [PubMed]
- Leite, G.; Morales, W.; Weitsman, S.; Celly, S.; Parodi, G.; Mathur, R.; Barlow, G.M.; Sedighi, R.; Millan, M.J.V.; Rezaie, A.; et al. The duodenal microbiome is altered in small intestinal bacterial overgrowth. PLoS ONE 2020, 15, e0234906. [Google Scholar] [CrossRef] [PubMed]
- Michail, S.; Lin, M.; Frey, M.R.; Fanter, R.; Paliy, O.; Hilbush, B.; Reo, N.V. Altered gut microbial energy and metabolism in children with non-alcoholic fatty liver disease. FEMS Microbiol. Ecol. 2015, 91, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Spencer, M.D.; Hamp, T.J.; Reid, R.W.; Fischer, L.M.; Zeisel, S.H.; Fodor, A.A. Association between composition of the human gastrointestinal microbiome and development of fatty liver with choline deficiency. Gastroenterology 2011, 140, 976–986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turnbaugh, P.J.; Ridaura, V.K.; Faith, J.J.; Rey, F.E.; Knight, R.; Gordon, J.I. The effect of diet on the human gut microbiome: A metagenomic analysis in humanized gnotobiotic mice. Sci. Transl. Med. 2009, 1, 6ra14. [Google Scholar] [CrossRef] [Green Version]
- Dumas, M.E.; Barton, R.H.; Toye, A.; Cloarec, O.; Blancher, C.; Rothwell, A.; Fearnside, J.; Tatoud, R.; Blanc, V.; Lindon, J.C.; et al. Metabolic profiling reveals a contribution of gut microbiota to fatty liver phenotype in insulin-resistant mice. Proc. Natl. Acad. Sci. USA 2006, 103, 12511–12516. [Google Scholar] [CrossRef] [Green Version]
- Yu, B.; Hailman, E.; Wright, S.D. Lipopolysaccharide binding protein and soluble CD14 catalyze exchange of phospholipids. J. Clin. Investig. 1997, 99, 315–324. [Google Scholar] [CrossRef]
- Nier, A.; Engstler, A.J.; Maier, I.B.; Bergheim, I. Markers of intestinal permeability are already altered in early stages of non-alcoholic fatty liver disease: Studies in children. PLoS ONE 2017, 12, e0183282. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | N | DEC | N | INC | p-Value | ||||
|---|---|---|---|---|---|---|---|---|---|
| Sex (men/women) | 6 | 3/3 | 11 | 4/7 | 0.58 | ||||
| Age (years) | 6 | 29.3 | ± | 8.1 | 11 | 34.3 | ± | 11.5 | 0.78 |
| BMI (kg/m2) | 6 | 30.5 | ± | 3.7 | 11 | 30.0 | ± | 5.9 | 0.89 |
| Waist circumference (cm) | 6 | 98.2 | ± | 17.5 | 11 | 98.7 | ± | 12.7 | 0.53 |
| SBP (mmHg) | 6 | 118.4 | ± | 4.6 | 11 | 112.5 | ± | 13.8 | 0.92 |
| DBP (mmHg) | 6 | 75.9 | ± | 8.8 | 11 | 72.1 | ± | 9.5 | 0.89 |
| Total-C (mmol/L) | 6 | 5.11 | ± | 0.93 | 11 | 4.43 | ± | 0.84 | 0.21 |
| HDL-C (mmol/L) | 6 | 1.23 | ± | 0.59 | 11 | 1.34 | ± | 0.36 | 0.88 |
| LDL-C (mmol/L) | 6 | 3.13 | ± | 0.93 | 11 | 2.38 | ± | 0.81 | 0.19 |
| TG (mmol/L) | 6 | 1.63 | ± | 0.86 | 11 | 1.56 | ± | 0.96 | 0.92 |
| Fasting glucose (mmol/L) | 5 | 5.54 | ± | 0.83 | 11 | 4.96 | ± | 0.43 | 0.21 |
| Fasting insulin (pmol/L) | 6 | 84.33 | ± | 55.52 | 9 | 90.67 | ± | 40.61 | 0.42 |
| HbA1C (%) | 6 | 5.05 | ± | 0.33 | 11 | 5.05 | ± | 0.31 | 0.77 |
| LBP (μg/mL) | 6 | 4.86 | ± | 0.67 | 11 | 5.49 | ± | 0.92 | 0.24 |
| LPS (pg/mL) | 6 | 76.02 | ± | 19.87 | 11 | 87.09 | ± | 19.63 | 0.33 |
| CRP (mg/L) | 6 | 2.30 | ± | 2.07 | 10 | 1.81 | ± | 1.10 | 0.57 |
| DEC | INC | p-Values | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Variable | N | Week 0 | N | Week 8 | N | Week 0 | N | Week 8 | Group | Visit | Int | ||||||||
| BMI (kg/m2) | 6 | 30.5 | ± | 3.7 | 6 | 30.4 | ± | 3.9 | 11 | 30.0 | ± | 5.9 | 11 | 30.2 | ± | 6.1 | 0.56 | 0.76 | 0.51 |
| Waist circ (cm) | 6 | 98.2 | ± | 17.5 | 6 | 98.7 | ± | 12.7 | 11 | 98.7 | ± | 12.7 | 11 | 99.1 | ± | 13.2 | 0.66 | 0.78 | 0.48 |
| SBP (mmHg) | 6 | 118.4 a | ± | 4.6 | 6 | 116.1 b | ± | 6.9 | 11 | 112.5 a | ± | 13.8 | 11 | 109.8 b | ± | 13.7 | 0.89 | 0.03 | 0.84 |
| DBP (mmHg) | 6 | 75.9 | ± | 8.8 | 6 | 72.6 | ± | 6.1 | 11 | 72.1 | ± | 9.5 | 11 | 70.1 | ± | 11.3 | 0.36 | 0.08 | 0.48 |
| Total-C (mmol/L) | 6 | 5.11 | ± | 0.93 | 6 | 4.99 | ± | 0.95 | 11 | 4.43 | ± | 0.84 | 11 | 4.20 | ± | 0.69 | 0.31 | 0.21 | 0.69 |
| HDL-C (mmol/L) | 6 | 1.23 | ± | 0.59 | 6 | 1.20 | ± | 0.52 | 11 | 1.34 | ± | 0.36 | 11 | 1.31 | ± | 0.24 | 0.92 | 0.57 | 0.97 |
| LDL-C (mmol/L) | 6 | 3.13 | ± | 0.93 | 6 | 3.08 | ± | 0.90 | 11 | 2.38 | ± | 0.81 | 11 | 2.33 | ± | 0.66 | 0.45 | 0.68 | 0.99 |
| TG (mmol/L) | 6 | 1.63 | ± | 0.86 | 6 | 1.54 | ± | 0.93 | 11 | 1.56 | ± | 0.96 | 11 | 1.21 | ± | 0.49 | 0.30 | 0.30 | 0.54 |
| Glucose (mmol/L) | 5 | 5.54 a | ± | 0.83 | 6 | 5.10 b | ± | 0.72 | 11 | 4.96 | ± | 0.43 | 11 | 4.95 | ± | 0.55 | 0.80 | 0.002 | 0.15 |
| Insulin (pmol/L) | 6 | 84.33 | ± | 55.52 | 5 | 71.60 | ± | 26.99 | 9 | 90.67 | ± | 40.61 | 10 | 103.70 | ± | 64.42 | 0.63 | 0.49 | 0.79 |
| HbA1C (%) | 6 | 5.05 | ± | 0.33 | 6 | 5.09 | ± | 0.29 | 11 | 5.05 | ± | 0.31 | 11 | 5.12 | ± | 0.28 | 0.76 | 0.71 | 0.71 |
| CRP (mg/L) | 6 | 2.30 | ± | 2.07 | 6 | 3.40 | ± | 2.57 | 10 | 1.81 | ± | 1.10 | 10 | 2.30 | ± | 1.90 | 0.38 | 0.05 | 0.42 |
| LBP (μg/mL) | 6 | 4.86 | ± | 0.67 | 6 | 5.53 | ± | 1.47 | 11 | 5.49 | ± | 0.92 | 11 | 5.27 | ± | 0.89 | 0.44 | 0.40 | 0.11 |
| LPS (pg/mL) | 6 | 76.02 | ± | 19.87 | 6 | 71.63 | ± | 29.83 | 11 | 87.09 | ± | 19.63 | 11 | 85.10 | ± | 21.37 | 0.95 | 0.39 | 0.74 |
| Phylum | Class | Order | Family | Genus | Species | β-Value | p-Value | r2 |
|---|---|---|---|---|---|---|---|---|
| Actinobacteriota | Actinobacteria | Actinomycetales | Actinomycetaceae | Actinomyces | Actinomyces odontolyticus | 9.5 | 0.02 | 0.62 |
| Actinobacteriota | Actinobacteria | Coriobacteriales | Coriobacteriaceae | Collinsella | Collinsella aerofaciens | −0.3 | 0.03 | 0.42 |
| Actinobacteriota | Actinobacteria | Actinomycetales | Micrococcaceae | Rothia | Rothia mucilaginosa | 9.5 | 0.01 | 0.52 |
| Bacteroidota | Bacteroidia | Bacteroidales | Bacteroidaceae | Bacteroides | Bacteroides nordii | 5.2 | 0.04 | 0.82 |
| Firmicutes | Clostridia | Clostridiales | Ruminococcaceae | Anaerotruncus | Anaerotruncus colihominis | 9.1 | 0.02 | 0.53 |
| Firmicutes | Clostridia | Clostridiales | Lachnospiraceae | Blautia | Blautia hydrogenotrophica | 20.0 | 0.02 | 0.71 |
| Firmicutes | Erysipelotrichia | Erysipelotrichales | Erysipelotrichaceae | Erysipelotrichaceae | Erysipelotrichaceae bacterium 21 3 | 12.6 | 0.03 | 0.45 |
| Firmicutes | Clostridia | Clostridiales | Clostridiales | Flavonifractor | Flavonifractor plautii | 9.7 | 0.03 | 0.36 |
| Firmicutes | Bacilli | Lactobacillales | Carnobacteriaceae | Granulicatella | Granulicatella unclassified | 13.0 | 0.03 | 0.48 |
| Firmicutes | Clostridia | Clostridiales | Ruminococcaceae | Ruminococcus | Ruminococcus callidus | 1.8 | 0.02 | 0.79 |
| Firmicutes | Bacilli | Lactobacillales | Streptococcaceae | Streptococcus | Streptococcus infantis | 9.7 | 0.02 | 0.55 |
| Proteobacteria | Deltaproteobacteria | Desulfovibrionales | Desulfovibrionaceae | Bilophila | Bilophila wadsworthia | 17.9 | 0.02 | 0.37 |
| Phylum | Class | Order | Family | Genus | Species | β-Value | p-Value | r2 |
|---|---|---|---|---|---|---|---|---|
| Actinobacteriota | Actinobacteria | Coriobacteriales | Coriobacteriaceae | Adlercreutzia | Adlercreutzia equolifaciens | −11.0 | 0.02 | 0.25 |
| Bacteroidota | Bacteroidia | Bacteroidales | Rikenellaceae | Alistipes | Alistipes finegoldii | −2.4 | 0.04 | 0.22 |
| Firmicutes | Bacilli | Lactobacillales | Carnobacteriaceae | Granulicatella | Granulicatella unclassified | −14.4 | 0.04 | 0.32 |
| Firmicutes | Bacilli | Lactobacillales | Streptococcaceae | Streptococcus | Streptococcus australis | −19.8 | 0.04 | 0.31 |
| Firmicutes | Bacilli | Lactobacillales | Streptococcaceae | Streptococcus | Streptococcus infantis | −17.6 | 0.05 | 0.34 |
| Firmicutes | Bacilli | Lactobacillales | Streptococcaceae | Streptococcus | Streptococcus mitis oralis pneumoniae | −17.6 | 0.05 | 0.31 |
| Firmicutes | Clostridia | Clostridiales | Clostridiaceae | Clostridium | Clostridium citroniae | −15.0 | 0.01 | 0.39 |
| Firmicutes | Clostridia | Clostridiales | Clostridiaceae | Clostridium | Clostridium hathewayi | −10.6 | 0.01 | 0.41 |
| Firmicutes | Clostridia | Clostridiales | Clostridiales | Clostridiales | Clostridiales bacterium 1 7 47FAA | −14.9 | 0.03 | 0.41 |
| Firmicutes | Clostridia | Clostridiales | Clostridiales | Flavonifractor | Flavonifractor plautii | −9.0 | 0.04 | 0.28 |
| Firmicutes | Clostridia | Clostridiales | Lachnospiraceae | Lachnospiraceae | Lachnospiraceae bacterium 7 1 58FAA | −8.1 | 0.03 | 0.21 |
| Firmicutes | Clostridia | Clostridiales | Ruminococcaceae | Anaerotruncus | Anaerotruncus colihominis | −16.4 | 0.05 | 0.32 |
| Firmicutes | Erysipelotrichia | Erysipelotrichales | Erysipelotrichaceae | Erysipelotrichaceae | Erysipelotrichaceae bacterium 6 1 45 | −17.5 | 0.00 | 0.53 |
| Firmicutes | Erysipelotrichia | Erysipelotrichales | Erysipelotrichaceae | Holdemania | Holdemania filiformis | −14.1 | 0.04 | 0.27 |
| Proteobacteria | Deltaproteobacteria | Desulfovibrionales | Desulfovibrionaceae | Bilophila | Bilophila wadsworthia | −14.0 | 0.02 | 0.29 |
| Proteobacteria | Gammaproteobacteria | Enterobacteriales | Enterobacteriaceae | Escherichia | Escherichia coli | −4.4 | 0.01 | 0.55 |
| Proteobacteria | Gammaproteobacteria | Enterobacteriales | Enterobacteriaceae | Escherichia | Escherichia unclassified | −7.8 | 0.03 | 0.53 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Franck, M.; de Toro-Martín, J.; V. Varin, T.; Garneau, V.; Pilon, G.; Roy, D.; Couture, P.; Couillard, C.; Marette, A.; Vohl, M.-C. Gut Microbial Signatures of Distinct Trimethylamine N-Oxide Response to Raspberry Consumption. Nutrients 2022, 14, 1656. https://0-doi-org.brum.beds.ac.uk/10.3390/nu14081656
Franck M, de Toro-Martín J, V. Varin T, Garneau V, Pilon G, Roy D, Couture P, Couillard C, Marette A, Vohl M-C. Gut Microbial Signatures of Distinct Trimethylamine N-Oxide Response to Raspberry Consumption. Nutrients. 2022; 14(8):1656. https://0-doi-org.brum.beds.ac.uk/10.3390/nu14081656
Chicago/Turabian StyleFranck, Maximilien, Juan de Toro-Martín, Thibault V. Varin, Véronique Garneau, Geneviève Pilon, Denis Roy, Patrick Couture, Charles Couillard, André Marette, and Marie-Claude Vohl. 2022. "Gut Microbial Signatures of Distinct Trimethylamine N-Oxide Response to Raspberry Consumption" Nutrients 14, no. 8: 1656. https://0-doi-org.brum.beds.ac.uk/10.3390/nu14081656