
Exercise: A Possibly Effective Way to Improve Vitamin D Nutritional Status



1
Shanghai Frontiers Science Research Base of Exercise and Metabolic Health, Shanghai University of Sport, Shanghai 200438, China
2
School of Kinesiology, Shanghai University of Sport, Shanghai 200438, China
*
Author to whom correspondence should be addressed.
Nutrients 2022, 14(13), 2652; https://0-doi-org.brum.beds.ac.uk/10.3390/nu14132652
Submission received: 26 May 2022
/
Revised: 20 June 2022
/
Accepted: 23 June 2022
/
Published: 27 June 2022
(This article belongs to the Special Issue Personalized Exercise and Sports Nutrition)
Abstract
:Vitamin D deficiency has become a widespread public health problem owing to its potential adverse health effects. Generally, the nutritional status of vitamin D depends on sunlight exposure and dietary or supplementary intake. However, recent studies have found that exercise can influence circulating 25(OH)D levels; although, the results have been inconclusive. In this review, we focused on the effect of exercise on circulating vitamin D metabolites and their possible mechanisms. We found that endurance exercise can significantly increase serum 25(OH)D levels in vitamin D-deficient people but has no significant effect on vitamin D-sufficient people. This benefit has not been observed with resistance training. Only chronic endurance exercise training can significantly increase serum 1,25(OH)2D, and the effect may be sex-dependent. Exercise may influence 25(OH)D levels in the circulation by regulating either the vitamin D metabolites stored in tissues or the utilization by target tissues. The effects of exercise on 25(OH)D levels in the circulation may be dependent on many factors, such as the vitamin D nutritional status, exercise type and intensity, and sex. Therefore, further research on the effects and mechanisms of exercise on vitamin D metabolites is required.
1. Introduction
The recent increase in vitamin D-related research has led to the discovery of the vitamin D receptor (VDR) in many tissues. A growing body of literature has shown that the biological role of vitamin D goes beyond the traditionally understood duties dealing with muscles and bones and is important for energy metabolism, oxidative stress, maintenance, and improvement of physical fitness [1,2,3,4]. A study found that vitamin D3 supplementation increased serum 25(OH)D levels; additionally, the expression of 291 genes, involving as many as 160 metabolic pathways, was significantly upregulated or downregulated [5]. This finding suggests that vitamin D plays an important role in health. Alarmingly, a survey found that vitamin D deficiency has become a global public health problem [6,7]. Vitamin D deficiency is closely associated with various chronic non-communicable diseases and functional disorders [1,8,9,10]. Therefore, maintaining adequate vitamin D levels is significant for promoting health.
Vitamin D is mainly synthesized by the skin, and its sources in food are scarce [11,12]. In the epidermis, 7-dehydrocholesterol can be transformed into vitamin D3 upon exposure to sunlight, while vitamin D2/3 in foods/supplements is absorbed into the circulation through the intestines. Both skin-synthesized vitamin D3 and food/supplement-derived D2/3 are catalyzed by 25-hydroxylase [mainly cytochrome P450 family 27 subfamily A member 1 (CYP27A1) and cytochrome P450 family 2 subfamily R member 1 (CYP2R1)] to 25(OH)D in the liver. Due to its long half-life [13] and strong vitamin D binding protein (VDBP) binding ability [14], serum 25(OH)D is the most abundant and stable vitamin D metabolite in the circulation; hence, its serum concentration is used to evaluate the nutritional status of vitamin D [15]. Subsequently, 25(OH)D is catalyzed by 25(OH)D-1α hydroxylase, cytochrome P450 family 27 subfamily B member 1 (CYP27B1), to 1,25(OH)2D in the kidney, which can bind to the VDR in the target tissue and regulate physiological processes. Vitamin D 24-hydroxylase, cytochrome P450 family 24 subfamily A member 1 (CYP24A1), an important degrading enzyme, can degrade 25(OH)D and 1,25(OH)2D in the kidney, which is then excreted through bile. Therefore, it is generally believed that skin synthesis and dietary intake/supplement are the main factors determining the nutritional status of vitamin D. Although scholars have suggested that vitamin D nutritional status can be improved by increasing the duration of sun exposure, the proportion of participants in one study with sufficient sunlight exposure was only 56.12% [16]. Moreover, prolonged UV exposure leads to hyperpigmentation and possible skin cancer [17]. Additionally, vitamin D is a photolabile; thus, prolonged UV exposure does not significantly increase vitamin D levels [18,19]. Therefore, support for prolonged sun exposure has lessened [20]. Vitamin D supplementation is a generally accepted method for improving its nutritional status; however, the recommended standards vary widely among the WHO, IOM, UK, EU, and China [21,22,23]. Moreover, while low doses are considered ineffective, high doses may lead to adverse effects, such as severe hypercalcemia and hyperphosphatemia [24].
Recent studies have found that physical inactivity is an important risk factor for morbidity and mortality from chronic non-communicable diseases [25] and for vitamin D deficiency [26]. Many observational studies have shown that the maintenance of vitamin D nutritional status is related to physical activity/exercise habits [27,28], and physical activity levels are significantly positively correlated with 25(OH)D levels [29,30]. Our meta-analysis of these studies revealed that physical activity was significantly positively correlated with circulating 25(OH)D levels [31]. However, results from experimental studies are inconsistent and may depend on the exercise type [31]. In this review, we focus on endurance exercise and resistance exercise to examine the effect of exercise on 25(OH)D and 1,25(OH)2D and its possible mechanisms.
2. Endurance Exercise and Vitamin D
2.1. The Effect of Acute Endurance Exercise
2.1.1. Human Studies
Five human studies have investigated the effects of acute endurance exercise on 25(OH)D and 1,25(OH)2D, with three focusing on professional athletes and two on non-athletes (Table 1). Mieszkowski et al. found that serum 25(OH)D levels in male runners [baseline serum 25(OH)D level >20 ng/mL] with and without vitamin D supplementation were significantly increased immediately and 24 h after an ultra-marathon race compared with those before the race [32]. Dzik also reported that serum 25(OH)D3 levels in male soccer players (10–14 years old) significantly increased at 15 min and 1 h after a VO2max test [25(OH)D >70 nmol/L]. In an analysis of both pre-pubertal and pubertal boys, the concentration of 25(OH)D3 increased 15 min after the VO2max test and dropped one hour after exercise, but these changes were not significantly different at specific time points [33]. However, Maimoun found that intensity exercise [47% Wmax, baseline serum 25(OH)D level: 79.4 ± 13.7 nmol/L; 64% Wmax, baseline serum 25(OH)D level: 83.4 ± 16 nmol/L] did not alter the concentration of 25(OH)D in male competitive road cyclists during cycling exercise or after 15 min of recovery [34]. Conversely, two studies involving non-athletes demonstrated that acute endurance exercise may increase circulating 25(OH)D levels. Maimoun et al. found that maximal incremental exercise can significantly increase the level of 25(OH)D in physically highly active elderly participants but not in moderately active elderly and young physically active adults [35]. Sun et al. found that serum 25(OH)D concentration significantly increased immediately and 1, 3, and 24 h after 30 min of cycling exercise at 70% VO2peak [36]. However, in the subgroup analysis, the 25(OH)D level increase of women [baseline serum 25(OH)D level:55.1 ± 15.6 nmol/L] was significant only at 24 h after exercise. The acute effect of exercise on 25(OH)D levels may be affected by intensity [33], sex [36], and age [33,35]. As for serum 1,25(OH)2D levels, no significant variation was observed in response to acute endurance exercise [34,35,36].
2.1.2. Animal Studies
Results from animal studies investigating the effect of acute endurance exercise on serum 25(OH)D have been inconsistent compared to those from human studies (Table 1). Makanae et al. found that acute endurance exercise (anaerobic threshold intensity) did not alter serum 25(OH)D levels in adult male Sprague-Dawley rats [37]. Moreover, serum 25(OH)D levels in horses were significantly reduced at 30 min, 1 week, and 3 weeks after high-intensity exercise [38]. However, only two experimental animal studies have been conducted to investigate the effect of acute endurance exercise on serum 25(OH)D levels.
2.2. The Effect of Chronic Endurance Exercise Training
2.2.1. Human Studies
Twelve human studies investigated the effect of endurance exercise training on serum 25(OH)D levels and did not yield consistent results (Table 1). Some studies found that chronic endurance exercise training can significantly increase serum 25(OH)D levels [39,40,41,42,43,44,45], but other studies have reported contradicting results [46,47,48,49,50]. However, when we sorted these studies, we found that in people with vitamin D deficiency {25(OH)D < 20 ng/mL or 50 nmol/L [57]}, endurance training can significantly improve serum 25(OH)D levels [39,40,42,43,45], and even severe vitamin D deficiency status (<10 ng/mL) improved to vitamin D deficiency status (10–20 ng/mL) in postmenopausal women [42]. However, endurance exercises had no significant effects on serum 25(OH)D levels in overweight and obese subjects, regardless of vitamin D nutritional status [48]. For participants with sufficient vitamin D levels {25(OH)D ≥ 20 ng/mL or 50 nmol/L [57]}, endurance training combined with vitamin D supplementation significantly increased serum 25(OH)D levels [40,44], while endurance training alone did not [40,46,49,50]. While Pilch et al. found that serum 25(OH)D levels were significantly reduced in postmenopausal obese women with sufficient vitamin D levels after endurance exercise intervention, the study was conducted in late autumn and had no control group; hence, it was impossible to determine whether the decrease in 25(OH)D was due to endurance exercise training or a seasonal decline [47]. When considering sun exposure, we found that studies providing relevant sun exposure information were all conducted in the morning [39,40,41,47] or evening [39] or autumn and winter [46,50]. During this time, sun exposure is weaker and has less effect on vitamin D. Taken together, the effect of chronic endurance exercise training on 25(OH)D levels in the circulation may be affected by the vitamin D status.
2.2.2. Animal Studies
Six animal studies investigated the effect of chronic endurance exercise training on 25(OH)D and 1,25(OH)2D (Table 1). Aly et al. found that a 4-week swimming regimen significantly increased serum 25(OH)D levels in diabetic mice. No significant change in serum 25(OH)D levels was observed in healthy mice; however, their serum 25(OH)D levels were significantly higher [51]. Buskermolen et al. found that although 6 weeks of endurance training increased serum 25(OH)D levels in female Wistar rats, the change was not significant [52]. The female Wistar rats in Buskermolen’s study were fed 1.5 IU/g (>1000 IU/kg [58,59,60]) vitamin D3, which is sufficient to maintain an adequate vitamin D status. Some animal studies have shown that chronic endurance exercise training can significantly increase serum 1,25(OH)2D levels in healthy female rats [53,54]. However, while Wang et al. found that 12 weeks of treadmill endurance exercise training slightly increased serum 1,25(OH)2D3 levels in aged male rats, the results were not significant [55]. Conversely, Xu et al. found that 8 weeks of swimming and downhill running significantly reduced serum 1,25(OH)2D3 level in 5-week old male mice [56]. In all animal studies which reported increased 1,25(OH)2D levels [53,54], the mice were all female, while those that were unchanged or decreased were male [55,56]. The effect of exercise training on 1,25(OH)2D may therefore depend on sex.
2.3. Mechanism
Endurance exercise induces greater improvements in aerobic capacity and its associated cardiopulmonary and metabolic variables [61]. In terms of energy metabolism, endurance exercise can activate several secondary signal molecules, such as AMPK, CaMKII, and p38, which promote an increase in PGC-1α. Subsequently, PGC-1α promotes mitochondrial biogenesis, exercise-induced fast-to-slow fiber-type transformation, and exercise-induced expression of important muscle antioxidant enzymes. Therefore, endurance exercise, especially submaximal endurance exercise, effectively increases fat metabolism [62]. Adipose tissue is one main storage depot for vitamin D [63]. Hengist et al. suggested that release of vitamin D stored in adipose tissue is a byproduct of lipolysis [64]. In other words, in the process of releasing triglycerides from adipocytes through the action of lipolytic enzymes, the stored vitamin D metabolites were also released. Lipolysis is regulated by various factors, such as atrial natriuretic peptides (ANPs), brain natriuretic peptides (BNPs), insulin, and beta adrenergic hormones [65]. Endurance exercise can promote the release of these hormones [66], promoting lipolytic processes and releasing vitamin D metabolites from the adipose tissue. Moreover, a systematic review showed that all exercise protocols (high-intensity interval exercise, moderate-intensity continuous exercise, and sprint interval exercise) can generate elevated energy expenditure through excessive post-exercise oxygen consumption (EPOC) [67]. Exercise-induced energy deficit has the most potent effect on endogenous lipid metabolism, elevating plasma triacylglycerol concentration and increasing plasma fatty acid mobilization and oxidation the day after performing endurance exercises [68]. The reason that endurance training can increase serum 25(OH)D levels may be attributed to lipolytic processes during exercise and EPOC.
Abboud et al. found that serum 25(OH)D levels in pasture sheep at the end of winter were significantly lower than those during the summer, but intramuscular 25(OH)D content at the end of winter was significantly higher [69]. After 25(OH)D3 supplementation, intramuscular 25(OH)D3 levels decreased as serum 25(OH)D3 levels rose, returning serum and skeletal muscle 25(OH)D concentrations to the more adequate summer levels [25(OH)D > 50 nmol/L] [69]. Abboud et al. found that when the vitamin D nutritional status improves, skeletal muscle cells may lose their ability to accumulate large amounts of 25(OH)D [69]. In addition, vitamin D nutritional status is regulated by a variety of factors such as serum Ca2+, Pi, parathyroid hormone (PTH), and FGF23 (fibroblast growth factor-23 (FGF23)) [70,71]. PTH stimulates the expression of CYP27B1 in the kidney, while FGF23, high Ca2+ or Pi levels, and 1,25(OH)2D downregulate it. In contrast, 1,25(OH)2D and FGF23 strongly induce the expression of CYP24A1, while PTH reduces its expression by stimulating its mRNA [72]. Moreover, PTH enhances the production of 1,25(OH)2D, which in turn activates an inhibitory loop regulating PTH production. Similarly, FGF23 regulates the production of 1,25(OH)2D, an inducer of FGF23 synthesis in the bones [70,73]. These factors work together to maintain vitamin D nutritional homeostasis, explaining why exercise cannot adequately elevate the level of 25(OH)D. Interestingly, serum calcium and PTH levels were significantly increased in the three groups [35]. Changes in PTH and calcium levels may therefore be responsible for the transient changes in 25(OH)D levels when there is no deficiency.
Endurance exercise can increase VDR mRNA levels [38,51,55]. In the target tissue, 1,25(OH)2D can bind to VDR and exert physiological functions, which may explain why exercise promotes health. Because 1,25(OH)2D utilization in the target tissue increases, so does conversion of 25(OH)D to 1,25(OH)2D3, resulting in reduced serum 25(OH)D levels [38].
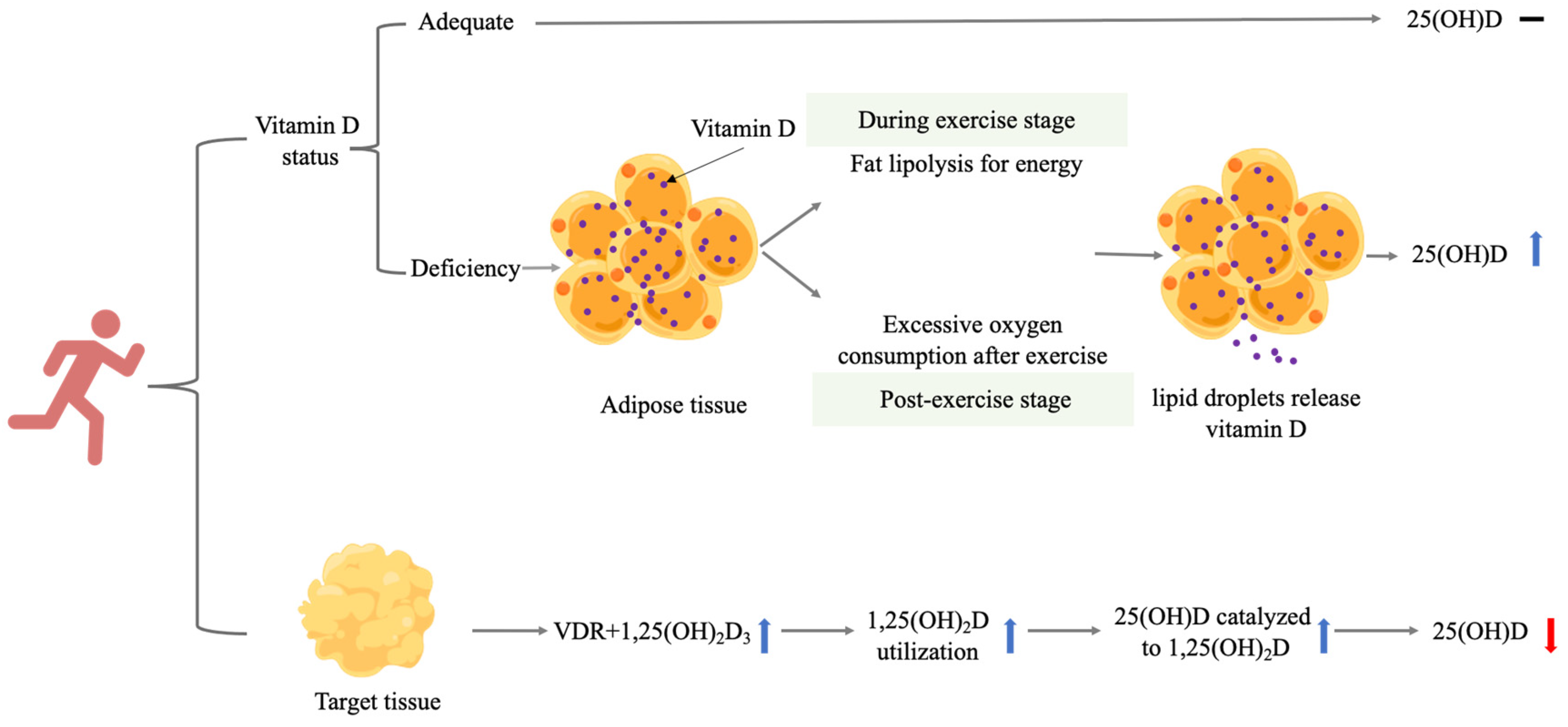
A summary of how endurance training may exert its effects on 25(OH)D in several ways can be seen in Figure 1.
3. Resistance Exercise
3.1. The Effect of Acute Resistance Exercise
Human and Animal Studies
One human study and one animal study have investigated the effect of acute resistance exercise intervention on 25(OH)D (Table 2). Barker et al. found that serum 25(OH)D concentrations significantly increased immediately after acute resistance exercise in 14 recreationally active adults; however, the levels subsequently decreased after 24, 48, 72, and 168 h [74]. Conversely, Makanae et al. reported no significant change in serum 25(OH)D concentrations in adult male Sprague-Dawley rats in response to acute resistance exercise [37]. In the human study, subjects performed an intense-stretch shortening contraction (10 sets of 10 repetitive jumps), whereas rats were put through isometric exercise (5 sets of 10 contractions). The inconsistency in outcome between the two studies may partially be explained by the differences in resistance exercise intensity and volume.
3.2. The Effect of Chronic Resistance Exercise Training
3.2.1. Human Studies
Five human studies have investigated the effects of resistance exercise training on 25(OH)D levels (Table 2). Resistance exercise training significantly increased circulating 25(OH)D levels in vitamin D-deficient post-stroke hemiplegia patients [75] and healthy participants [76]. Conversely, resistance exercise training had no effect on 25(OH)D levels in healthy vitamin D-deficient young men [77] and older adults without vitamin D supplementation [78]. However, Agergaard et al. found that resistance exercise training significantly reduced serum 25(OH)D levels in young and elderly participants without vitamin D supplementation [79]. Factors such as vitamin D supplementation, season, and experimental design should be considered when interpreting these findings. We found that all groups received vitamin D supplementation in Zhang’s study [75]; however, serum 25(OH)D levels were higher at 3 months and 1 year following resistance exercise training combined with vitamin D, compared to only vitamin D supplementation. In Bass’s study, there was a lack of control groups and seasonal information; hence, it is unclear whether the increase in serum 25(OH)D levels is due to resistance training or seasonal factors [76]. Detailed seasonal information was provided in Aschauer’s study (from mid-February to mid-July) [78], Sun’s study (from March to July) [77], and Agergaard’s study (from November to December) [79]. We found a clear seasonal trend in mean serum 25(OH)D concentrations, suggesting that the change in 25(OH)D concentrations induced by resistance training may have been caused by large seasonal fluctuations [77,78].
3.2.2. Animal Studies
Two animal studies have investigated the effect of resistance exercise training on 25(OH)D and 1,25(OH)2D3 (Table 2). Buskermolen et al. found that 6 weeks of peak power training did not alter serum 25(OH)D levels in rats [52]. Conversely, Xu et al. found that 8 weeks of jumping training significantly reduced serum 1,25(OH)2D3 levels in male mice [56].
3.3. Mechanisms
The predominant adaptation of resistance exercises is in the musculoskeletal system, including increases in muscle mass, muscle strength, and bone density [61]. Muscle mass is increased when resistance exercise triggers muscle signaling events that activate mTOR, leading to increased protein synthesis [62]. Therefore, resistance exercise can be effective in increasing muscle weight and hypertrophy. Mason et al. reported that circulating VDBP can be internalized into skeletal muscle cells to provide high-affinity intracellular binding sites for 25(OH)D [80]. The authors postulate that this intracellular VDBP enables 25(OH)D to diffuse into muscle cells where it is bound and retained until VDBP undergoes proteolysis [80]. The released 25(OH)D then diffuses from the skeletal muscle cells into the circulation and is immediately bound by VDBP in the circulation [80]. Thus, muscle tissue may be an important target tissue and extravascular storage pool for vitamin D. In Sun’s study, fat-free mass and muscle mass were significantly increased [77]. Similarly, in Agergaard’s study, the cross-sectional area of the quadriceps muscle had significant gains in the group who did not receive vitamin D supplements [79]. This result suggests that increased muscle mass from resistance training provides a reservoir of vitamin D, leading to reduced or unchanged serum 25(OH)D levels.
Moreover, 25(OH)D can be released from skeletal muscle [69,81,82,83,84]. This release is regulated by the VDR, PTH, VDBP, and vitamin D nutritional status [69,83,84]. PTH reduces the net uptake of 25(OH)D3 in C2 myotubes and mouse muscle fibers and reduces its retention in myotubes [69]. In Barker’s study, PTH levels significantly increased after acute resistance exercise [74]. In Zhang’s study, there was a significant increase in PTH levels at 3 months and 1 year following chronic resistance exercise combined with vitamin D supplements, compared to only receiving vitamin D supplements [75]. The increase in circulating 25(OH)D levels may be due to the effect of PTH on its uptake and retention in skeletal muscle cells [74,75]. However, chronic resistance training alone did not significantly alter the PTH levels [52,77]. This may also be the reason why vitamin D supplementation combined with resistance training, and not resistance exercise training alone, increases 25(OH)D levels.
Resistance training can increase the level of CYP27B1 [37], which can catalyze the conversion of 25(OH)D to 1,25(OH)2D3. Moreover, resistance training can increase target tissue VDR levels [37], increasing 1,25(OH)2D3 utilization, which may explain why serum 25(OH)D levels are not altered in response to resistance training [37]. In addition, resistance training increases CYP24A1 levels [56], which can degrade 25(OH)D and 1,25(OH)2D; hence, the decrease in 1,25(OH)2D may be caused by increased degradation, while its synthesis remains unchanged [56]. These factors may individually or together contribute to reduction/unchanged 25(OH)D levels in the circulation in response to resistance training.
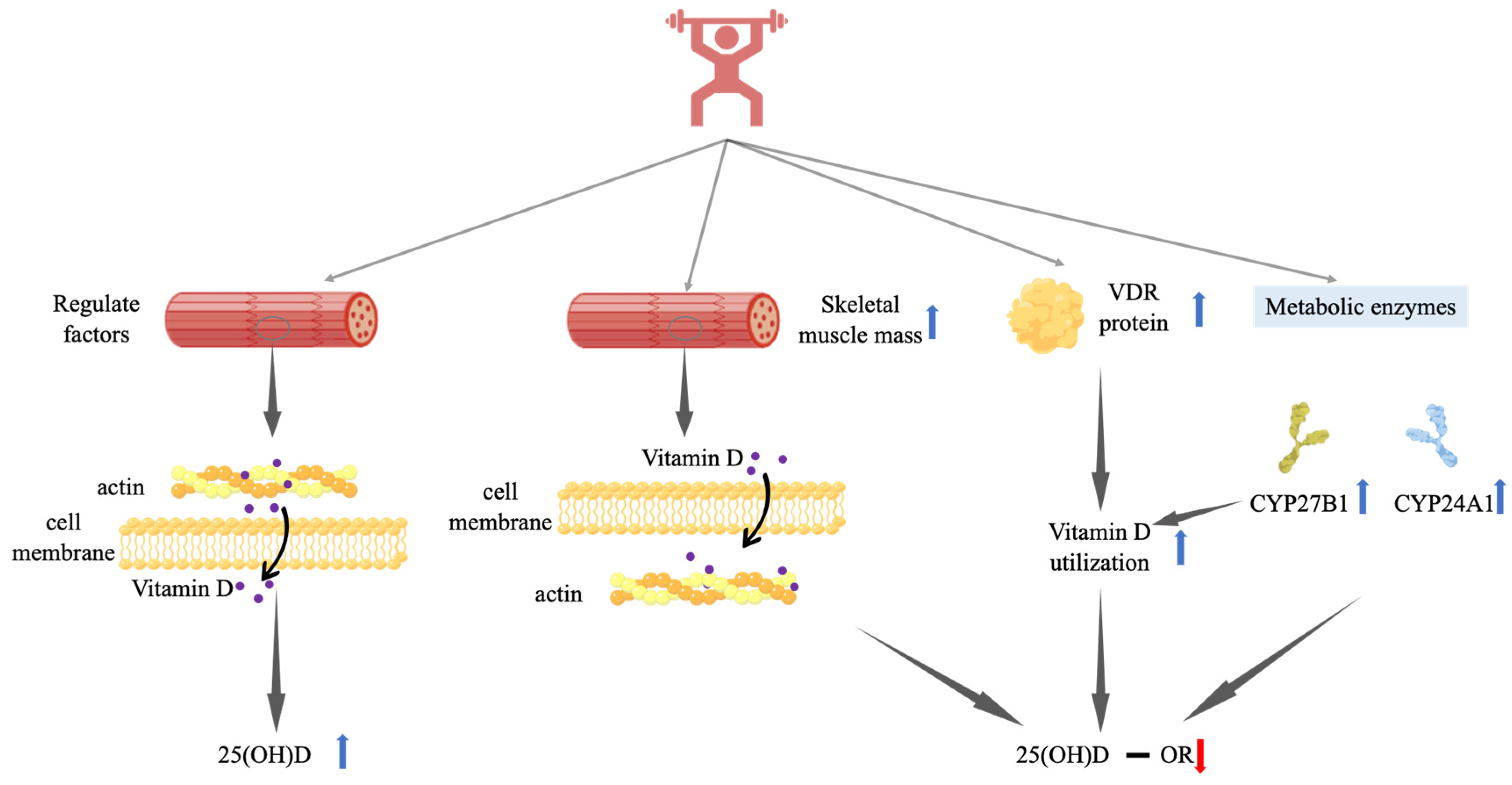
How resistance training may exert its effect on 25(OH)D in various ways is briefly illustrated in Figure 2.
4. Others
The effect of endurance combined with resistance exercise training intervention on 25(OH)D was investigated in three human studies and one animal study, which did not yield consistent results (Table 3). In the human studies, chronic endurance combined with resistance exercise training intervention significantly increased serum 25(OH)D levels [85,86]. Evans et al. found that 4 months of recruit training significantly reduced serum 25(OH)D levels in healthy men with adequate vitamin D {25(OH)D ≥ 20 ng/mL or 50 nmol/L [57]}, while no significant change was observed in healthy women with adequate vitamin D levels [87]. Conversely, Buskermolen et al. reported that 6 weeks of peak power combined with endurance training did not alter serum 25(OH)D levels in Wistar rats [52].
5. Limitations and Perspectives
Aside from the small number of relevant studies, there are many limitations. First, while mass spectrometry, enzyme-linked immunosorbent assays, and other methods can detect 25(OH)D levels, their accuracies vary greatly. Moreover, 25(OH)D2 levels are difficult to detect [24]. Second, the vitamin D nutritional status is affected by exposure to season/sunlight. Except for the Sun study and the Li study, which clearly stated that chronic exercise intervention was conducted indoors [77] or outdoors [86], the vast majority of studies did not provide relevant information. Some studies did not provide seasonal information or information on sunlight exposure. Third, some studies did not include a blank control group. These limitations should be addressed in future research. In this paper, there are also some strengths. First, we relatively comprehensively summarize the relevant research in recent years. Second, due to the different effects of different exercise types on health, we focus on the analysis of the effects of endurance training and resistance training exercise. Third, from the perspective of the two major extra-circulating depots and the regulatory factors of vitamin D, this review comprehensively explained the possible mechanism of exercise on vitamin D.
Because the nutritional status of vitamin D is influenced by various factors, we recommend incorporating the following considerations in future studies. First, due to their lipid solubility, vitamin D metabolites are sequestered in adipose tissue, leading to decreased bioavailability in obese subjects [88]. Moreover, Drincic believed that because of volumetric dilution, obese individuals have lower 25(OH)D concentrations [89]. Therefore, body fat is significantly negatively correlated with serum 25(OH)D levels [90] and obese individuals have a higher risk of vitamin D deficiency [91,92]. In addition, lipolysis may be impaired in obese individuals [93], and obesity affects the regulation of vitamin D metabolism enzymes [94], which may explain why 25(OH)D levels in overweight and obese adults were not altered in the Lithgow study [48]. Second, exercise in the fed and fasted states differed in terms of energy metabolism substrates. A study found that exercise performed in the fasted state induces higher fat oxidation than exercise performed in the fed state [95]. Moreover, fasting increases post-exercise circulating FFAs [96]. Therefore, the effects of exercise on serum 25(OH)D or 1,25(OH)2D levels may be influenced by whether it is performed under fed or fasted states. Third, vitamin D metabolites are primarily found in circulation, adipose tissue, and skeletal muscle [63]. Thus, adipose and muscle tissues are two major extra circulatory depots for vitamin D metabolites, which are not reflected in serum 25(OH)D levels. Therefore, when studying the effect of exercise on vitamin D, extravascular storage tissues should be included in the analysis.
6. Conclusions
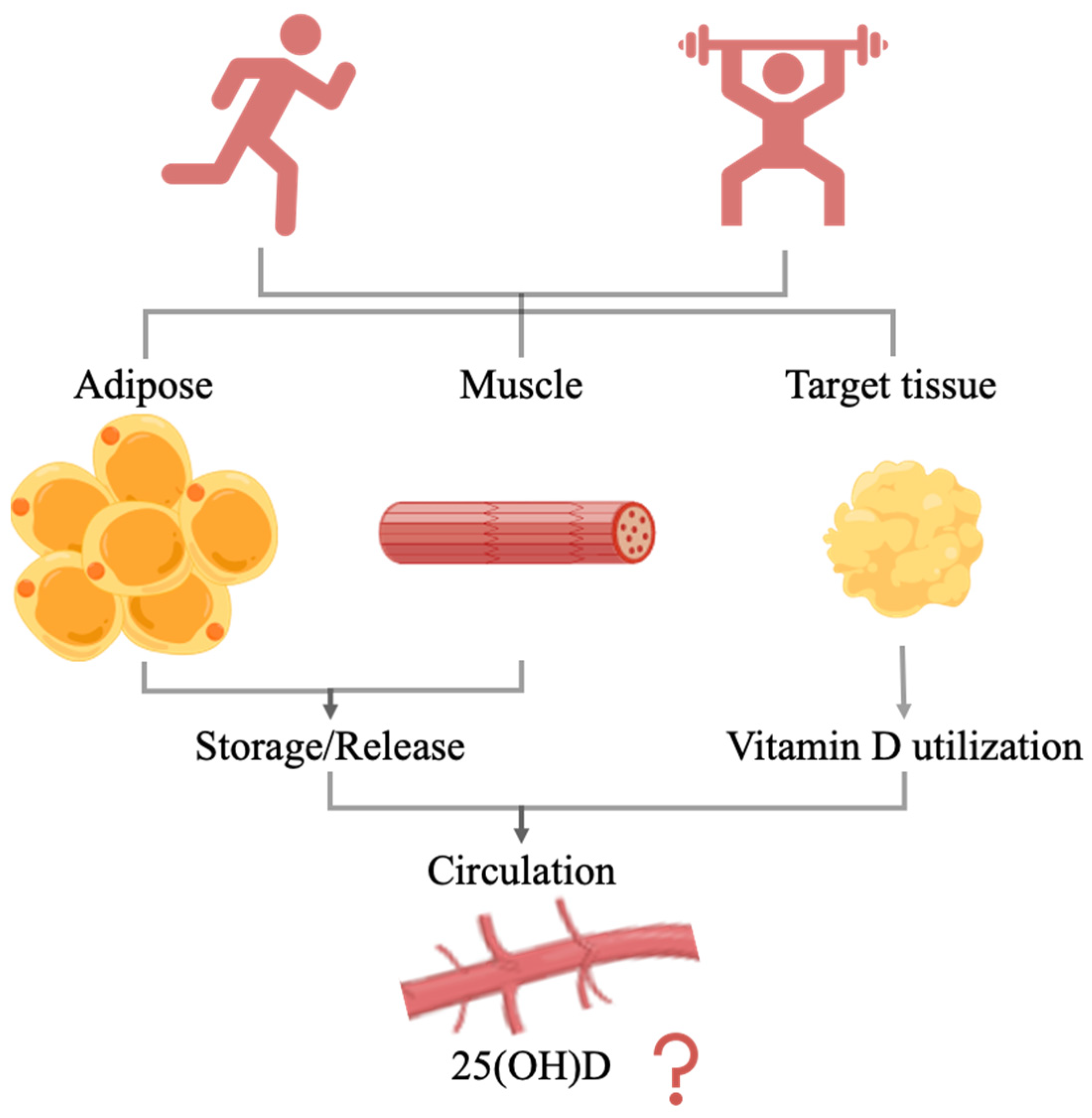
In conclusion, endurance exercise can significantly increase serum 25(OH)D levels in vitamin D-deficient subjects but has no significant effect on vitamin D-sufficient subjects. Moreover, resistance training did not significantly increase 25(OH)D concentrations. Only chronic endurance exercise intervention significantly increased serum 1,25(OH)2D levels, and this effect may be sex-dependent. Exercise may influence 25(OH)D levels in circulation by regulating either the release of vitamin D metabolites from storage tissues or the utilization of target tissue (Figure 3). The effects of exercise on 25(OH)D levels may depend on the vitamin D nutritional status, exercise type, exercise intensity, and sex. The organism is a complex entity, and vitamin D is tightly regulated by a variety of factors. Exercise elicits various bodily responses, and the effects of exercise on vitamin D nutritional levels may be the result of a combination of these. Therefore, further research on the effects and mechanisms of exercise on 25(OH)D levels is needed.
Author Contributions
Conceptualization, J.Z. and Z.-B.C.; methodology, Jinghua Zhang; software, J.Z.; validation, J.Z. and Z.-B.C.; formal analysis, J.Z. and Z.-B.C.; investigation, J.Z. and Z.-B.C.; resources, J.Z. and Z.-B.C.; data curation, J.Z. and Z.-B.C.; writing—original draft preparation, J.Z.; writing—review and editing, J.Z. and Z.-B.C.; visualization, J.Z. and Z.-B.C.; supervision, Z.-B.C.; project administration, J.Z. and Z.-B.C.; funding acquisition, Z.-B.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Natural Science Foundation of Shanghai (No. 20ZR1454400).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Maddaloni, E.; Cavallari, I.; Napoli, N.; Conte, C. Vitamin D and diabetes mellitus. Front. Horm. Res. 2018, 50, 161–176. [Google Scholar]
- Sun, X.; Cao, Z.B.; Tanisawa, K.; Ito, T.; Oshima, S.; Higuchi, M. Vitamin D supplementation reduces insulin resistance in Japanese adults: A secondary analysis of a double-blind, randomized, placebo-controlled trial. Nutr. Res. 2016, 36, 1121–1129. [Google Scholar] [CrossRef]
- Yan, C.L. Molecular mechanism of vitamin D in the cardiovascular system. J. Investig. Med. 2015, 59, 868–871. [Google Scholar]
- Artaza-Artabe, I.; Sáez-López, P.; Sánchez-Hernández, N.; Fernández-Gutierrez, N.; Malafarina, V. The relationship between nutrition and frailty: Effects of protein intake, nutritional supplementation, vitamin D and exercise on muscle metabolism in the elderly. A systematic review. Maturitas 2016, 93, 89–99. [Google Scholar] [CrossRef]
- Hossein-nezhad, A.; Spira, A.; Holick, M.F. Influence of vitamin D status and vitamin D3 supplementation on genome wide expression of white blood cells: A randomized double-blind clinical trial. PLoS ONE 2013, 8, e58725. [Google Scholar] [CrossRef] [Green Version]
- Rosas-Peralta, M.; Holick, M.F.; Borrayo-Sanchez, G.; Madrid-Miller, A.; Ramirez-Arias, E.; Arizmendi-Uribe, E. Dysfunctional immunometabolic effects of vitamin D deficiency, increased cardiometabolic risk. Potential epidemiological alert in America? Endocrinol. Diabetes Nutr. 2017, 64, 162–173. [Google Scholar] [CrossRef]
- Palacios, C.; Gonzalez, L. Is vitamin D deficiency a major global public health problem? J. Steroid. Biochem. Mol. Biol. 2014, 144PA, 138–145. [Google Scholar] [CrossRef] [Green Version]
- Dzik, K.P.; Kaczor, J.J. Mechanisms of vitamin D on skeletal muscle function: Oxidative stress, energy metabolism and anabolic state. Eur. J. Appl. Physiol. 2019, 119, 825–839. [Google Scholar] [CrossRef] [Green Version]
- Han, B.; Wang, X.; Wang, N.; Li, Q.; Chen, Y.; Zhu, C.; Chen, Y.; Xia, F.; Pu, X.; Cang, Z.; et al. Investigation of vitamin D status and its correlation with insulin resistance in a Chinese population. Public Health Nutr. 2017, 20, 1602–1608. [Google Scholar] [CrossRef] [Green Version]
- Prasad, P.; Kochhar, A. Interplay of vitamin D and metabolic syndrome: A review. Diabetes Metab. Syndr. 2016, 10, 105–112. [Google Scholar] [CrossRef]
- Poskitt, E.M.; Cole, T.J.; Lawson, D.E. Diet, sunlight, and 25-hydroxy vitamin D in healthy children and adults. Br. Med. J. 1979, 1, 221–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawson, D.E.; Paul, A.A.; Black, A.E.; Cole, T.J.; Mandal, A.R.; Davie, M. Relative contributions of diet and sunlight to vitamin D state in the elderly. Br. Med. J. 1979, 2, 303–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Datta, P.; Philipsen, P.A.; Olsen, P.; Bogh, M.K.; Johansen, P.; Schmedes, A.V.; Morling, N.; Wulf, H.C. The half-life of 25(OH)D after UVB exposure depends on gender and vitamin D receptor polymorphism but mainly on the start level. Photochem. Photobiol. Sci. 2017, 16, 985–995. [Google Scholar] [CrossRef]
- Bikle, D.D.; Gee, E.; Halloran, B.; Kowalski, M.A.; Ryzen, E.; Haddad, J.G. Assessment of the free fraction of 25-hydroxyvitamin D in serum and its regulation by albumin and the vitamin D-binding protein. J. Clin. Endocrinol. Metab. 1986, 63, 954–959. [Google Scholar] [CrossRef] [PubMed]
- Kiourtzidis, M.; Kühn, J.; Brandsch, C.; Baur, A.C.; Wensch-Dorendorf, M.; Stangl, G.I. Markers indicating body vitamin D stores and responses of liver and adipose tissues to changes in vitamin D intake in male mice. Nutrients 2020, 12, 1391. [Google Scholar] [CrossRef]
- Zhang, R.H.; He, D.H.; Zhou, B.; Zhu, Y.B.; Zhao, D.; Huang, L.C.; Ding, G.Q. Analysis of vitamin D status in men highly exposed to sunlight. Biomed. Environ. Sci. 2015, 28, 913–916. [Google Scholar]
- Leffell, D.J.; Brash, D.E. Sunlight and skin cancer. Sci. Am. 1996, 275, 52–53, 56–59. [Google Scholar] [CrossRef]
- Webb, A.R.; DeCosta, B.R.; Holick, M.F. Sunlight regulates the cutaneous production of vitamin D3 by causing its photodegradation. J. Clin. Endocrinol. Metab. 1989, 68, 882–887. [Google Scholar] [CrossRef]
- Holick, M.F. Sunlight and vitamin D for bone health and prevention of autoimmune diseases, cancers, and cardiovascular disease. Am. J. Clin. Nutr. 2004, 80, 1678S–1688S. [Google Scholar] [CrossRef] [Green Version]
- Webb, A.R. Who, what, where and when-influences on cutaneous vitamin D synthesis. Prog. Biophys Mol. Biol. 2006, 92, 17–25. [Google Scholar] [CrossRef]
- Roth, D.E.; Abrams, S.A.; Aloia, J.; Bergeron, G.; Bourassa, M.W.; Brown, K.H.; Calvo, M.S.; Cashman, K.D.; Combs, G.; De-Regil, L.M.; et al. Global prevalence and disease burden of vitamin D deficiency: A roadmap for action in low- and middle-income countries. Ann. N. Y. Acad. Sci. 2018, 1430, 1–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cashman, K.D.; Ritz, C.; Kiely, M.; Odin, C. Improved dietary guidelines for vitamin D: Application of individual participant aata (IPD)-level meta-regression analyses. Nutrients 2017, 9, 469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chinese Nutrition Society. Dietary Guidelines for Chinese Residents 2016; People’s Medical Publishing House: Beijing, China, 2016; pp. 201–202. (In Chinese) [Google Scholar]
- Razzaque, M.S. Can adverse effects of excessive vitamin D supplementation occur without developing hypervitaminosis D? J. Steroid. Biochem. Mol. Biol. 2018, 180, 81–86. [Google Scholar] [CrossRef]
- Lee, I.M.; Shiroma, E.J.; Lobelo, F.; Puska, P.; Blair, S.N.; Katzmarzyk, P.T. Effect of physical inactivity on major non-communicable diseases worldwide: An analysis of burden of disease and life expectancy. Lancet 2012, 380, 219–229. [Google Scholar] [CrossRef] [Green Version]
- Palaniswamy, S.; Hyppönen, E.; Williams, D.M.; Jokelainen, J.; Lowry, E.; Keinänen-Kiukaanniemi, S.; Herzig, K.H.; Järvelin, M.R.; Sebert, S. Potential determinants of vitamin D in Finnish adults: A cross-sectional study from the Northern Finland birth cohort 1966. BMJ Open 2017, 7, e013161. [Google Scholar] [CrossRef] [PubMed]
- Wanner, M.; Richard, A.; Martin, B.; Linseisen, J.; Rohrmann, S. Associations between objective and self-reported physical activity and vitamin D serum levels in the US population. Cancer Causes Control 2015, 26, 881–891. [Google Scholar] [CrossRef] [Green Version]
- Touvier, M.; Deschasaux, M.; Montourcy, M.; Sutton, A.; Charnaux, N.; Kesse-Guyot, E.; Assmann, K.E.; Fezeu, L.; Latino-Martel, P.; Druesne-Pecollo, N.; et al. Determinants of vitamin D status in Caucasian adults: Influence of sun exposure, dietary intake, sociodemographic, lifestyle, anthropometric, and genetic factors. J. Investig. Derm. 2015, 135, 378–388. [Google Scholar] [CrossRef] [Green Version]
- Scragg, R.; Holdaway, I.; Jackson, R.; Lim, T. Plasma 25-hydroxyvitamin D3 and its relation to physical activity and other heart disease risk factors in the general population. Ann. Epidemiol. 1992, 2, 697–703. [Google Scholar] [CrossRef]
- Scragg, R.; Holdaway, I.; Singh, V.; Metcalf, P.; Baker, J.; Dryson, E. Serum 25-hydroxyvitamin D3 is related to physical activity and ethnicity but not obesity in a multicultural workforce. Aust. N. Z. J. Med. 1995, 25, 218–223. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, K.; Cao, Z. Effects of physical activity on vitamin D: A systematic review and meta-analysis of observational and experimental studies. J. Shanghai Univ. Sport 2021, 45, 81–96. (In Chinese) [Google Scholar]
- Mieszkowski, J.; Stankiewicz, B.; Kochanowicz, A.; Niespodziński, B.; Kowalik, T.; Żmijewski, M.; Kowalski, K.; Rola, R.; Bieńkowski, T.; Antosiewicz, J. Ultra-marathon-induced increase in serum levels of vitamin D metabolites: A double-blind randomized controlled trial. Nutrients 2020, 12, 3629. [Google Scholar] [CrossRef]
- Dzik, K.P.; Grzywacz, T.; Łuszczyk, M.; Kujach, S.; Flis, D.J.; Kaczor, J.J. Single bout of exercise triggers the increase of vitamin D blood concentration in adolescent trained boys: A pilot study. Sci. Rep. 2022, 12, 1825. [Google Scholar] [CrossRef]
- Maimoun, L.; Manetta, J.; Couret, I.; Dupuy, A.M.; Mariano-Goulart, D.; Micallef, J.P.; Peruchon, E.; Rossi, M. The intensity level of physical exercise and the bone metabolism response. Int. J. Sports Med. 2006, 27, 105–111. [Google Scholar] [CrossRef]
- Maimoun, L.; Simar, D.; Caillaud, C.; Coste, O.; Barbotte, E.; Peruchon, E.; Rossi, M.; Mariano-Goulart, D. Response of calciotropic hormones and bone turnover to brisk walking according to age and fitness level. J. Sci. Med. Sport 2009, 12, 463–467. [Google Scholar] [CrossRef]
- Sun, X.; Cao, Z.B.; Taniguchi, H.; Tanisawa, K.; Higuchi, M. Effect of an acute bout of endurance exercise on serum 25(OH)D concentrations in young adults. J. Clin. Endocrinol. Metab. 2017, 102, 3937–3944. [Google Scholar] [CrossRef] [Green Version]
- Makanae, Y.; Ogasawara, R.; Sato, K.; Takamura, Y.; Matsutani, K.; Kido, K.; Shiozawa, N.; Nakazato, K.; Fujita, S. Acute bout of resistance exercise increases vitamin D receptor protein expression in rat skeletal muscle. Exp. Physiol. 2015, 100, 1168–1176. [Google Scholar] [CrossRef]
- Puangthong, C.; Sukhong, P.; Saengnual, P.; Srikuea, R.; Chanda, M. A single bout of high-intensity exercise modulates the expression of vitamin D receptor and vitamin D-metabolising enzymes in horse skeletal muscle. Equine Vet. J. 2021, 53, 796–805. [Google Scholar] [CrossRef]
- Farag, H.A.M.; Hosseinzadeh-Attar, M.J.; Muhammad, B.A.; Esmaillzadeh, A.; el Bilbeisi, A.H. Effects of vitamin D supplementation along with endurance physical activity on lipid profile in metabolic syndrome patients: A randomized controlled trial. Diabetes Metab. Syndr. Clin. Res. Rev. 2019, 13, 1093–1098. [Google Scholar] [CrossRef]
- Mieszkowski, J.; Niespodzinski, B.; Kochanowicz, A.; Gmiat, A.; Prusik, K.; Prusik, K.; Kortas, J.; Ziemann, E.; Antosiewicz, J. The effect of nordic walking training combined with vitamin D supplementation on postural control and muscle strength in elderly people-a randomized controlled trial. Int. J. Env. Res. Public Health 2018, 15, 1951. [Google Scholar] [CrossRef] [Green Version]
- Prusik, K.; Kortas, J.; Prusik, K.; Mieszkowski, J.; Jaworska, J.; Skrobot, W.; Lipinski, M.; Ziemann, E.; Antosiewicz, J. Nordic walking training causes a decrease in blood cholesterol in elderly women supplemented with vitamin D. Front. Endocrinol. 2018, 9, 42. [Google Scholar] [CrossRef] [Green Version]
- Malandish, A.; Tartibian, B.; Sheikhlou, Z.; Afsargharehbagh, R.; Rahmati, M. The effects of short-term moderate intensity aerobic exercise and long-term detraining on electrocardiogram indices and cardiac biomarkers in postmenopausal women. J. Electrocardiol. 2020, 60, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, R.; Xu, l.; Chen, X. Effect of high intensity interval training on rehabilitation of elderly chronic obstructive pulmonary disease patients with osteoporosis. Chin. J. Front. Med. Sci. 2019, 11, 30–34. (In Chinese) [Google Scholar]
- Song, L.; Xuan, Y.; Yang, J.; Xuan, M.; Wang, Y.; Song, L.; Wang, W.; Zhang, X. The association among the management of diet and sport, glucose metabolism, bone metabolism, and bone mineral density in postmenopausal women with type II diabetes and osteoporosis: A clinical study. Chin. J. Front. Med. Sci. 2014, 20, 156–160. (In Chinese) [Google Scholar]
- Shi, D.; Shi, X.; Li, F.; Ren, J.; Gu, L. Clinical effect of exercise therapy on the patients with osteoporosis. Chin. J. Geriatr. 2013, 32, 872–874. (In Chinese) [Google Scholar]
- Klausen, T.; Breum, L.; Sørensen, H.A.; Schifter, S.; Sonne, B. Plasma levels of parathyroid hormone, vitamin D, calcitonin, and calcium in association with endurance exercise. Calcif. Tissue Int. 1993, 52, 205–208. [Google Scholar] [CrossRef] [PubMed]
- Pilch, W.; Tyka, A.; Cebula, A.; Sliwicka, E.; Pilaczynska-Szczesniak, L.; Tyka, A. Effects of a 6-week Nordic walking training on changes in 25(OH)D blood concentration in women aged over 55. J. Sports Med. Phys. Fit. 2017, 57, 124–129. [Google Scholar] [CrossRef]
- Lithgow, H.M.; Florida-James, G.; Leggate, M. The combined effect of high-intensity intermittent training and vitamin D supplementation on glycemic control in overweight and obese adults. Physiol. Rep. 2018, 6, e13684. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.J.; Levinson, A.; George, D.; Canas, J.; Kumar, S.; Balagopal, P.B. Vitamin D status and cardiovascular risk in obesity: Effect of physical activity in nonvitamin D supplemented adolescents. Metab. Syndr. Relat. Disord. 2018, 16, 197–203. [Google Scholar] [CrossRef]
- Sun, X.; Cao, Z.B.; Tanisawa, K.; Taniguchi, H.; Kubo, T.; Higuchi, M. Effects of chronic endurance exercise training on serum 25(OH)D concentrations in elderly Japanese men. Endocrine 2018, 59, 330–337. [Google Scholar] [CrossRef]
- Aly, Y.E.; Abdou, A.S.; Rashad, M.M.; Nassef, M.M. Effect of exercise on serum vitamin D and tissue vitamin D receptors in experimentally induced type 2 Diabetes Mellitus. J. Adv. Res. 2016, 7, 671–679. [Google Scholar] [CrossRef] [Green Version]
- Buskermolen, J.; van der Meijden, K.; Furrer, R.; Mons, D.J.; van Essen, H.W.; Heijboer, A.C.; Lips, P.; Jaspers, R.T.; Bravenboer, N. Effects of different training modalities on phosphate homeostasis and local vitamin D metabolism in rat bone. PeerJ 2019, 24, e6184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, J.K.; Aloia, J.F.; Yasumura, S. Effect of physical activity on calcium and phosphorus metabolism in the rat. Am. J. Physiol. 1989, 256, E1–E6. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, J.; Shimamura, C.; Takeda, T.; Abe, H.; Ichimura, S.; Sato, Y.; Toyama, Y. Effects of treadmill exercise on bone mass, bone metabolism, and calciotropic hormones in young growing rats. J. Bone Min. Metab. 2004, 22, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Hou, J. Effects of chronic aerobic exercise training on expression of vitamin D receptor in bone tissue of aged rats. J. Xi’An Technol. Univ. 2018, 38, 318–323. (In Chinese) [Google Scholar]
- Xu, S.; Li, S.; Chen, X. The effect of exercise on the expression of FGF23—klotho/FGFR1 axis and related factors of male mice. Chin. J. Sports Med. 2019, 38, 882–889. (In Chinese) [Google Scholar]
- Ross, A.C.; Manson, J.E.; Abrams, S.A.; Aloia, J.F.; Brannon, P.M.; Clinton, S.K.; Durazo-Arvizu, R.A.; Gallagher, J.C.; Gallo, R.L.; Jones, G.; et al. The 2011 report on dietary reference intakes for calcium and vitamin D from the Institute of Medicine: What clinicians need to know. J. Clin. Endocrinol. Metab. 2011, 96, 53–58. [Google Scholar] [CrossRef]
- Fleet, J.C.; Gliniak, C.; Zhang, Z.; Xue, Y.; Smith, K.B.; McCreedy, R.; Adedokun, S.A. Serum metabolite profiles and target tissue gene expression define the effect of cholecalciferol intake on calcium metabolism in rats and mice. J. Nutr. 2008, 138, 1114–1120. [Google Scholar] [CrossRef] [Green Version]
- Lee, G.Y.; Park, C.Y.; Cha, K.S.; Lee, S.E.; Pae, M.; Han, S.N. Differential effect of dietary vitamin D supplementation on natural killer cell activity in lean and obese mice. J. Nutr. Biochem. 2018, 55, 178–184. [Google Scholar] [CrossRef]
- Maia-Ceciliano, T.C.; Dutra, R.R.; Aguila, M.B.; Mandarim-De-Lacerda, C.A. The deficiency and the supplementation of vitamin D and liver: Lessons of chronic fructose-rich diet in mice. J. Steroid. Biochem. Mol. Biol. 2019, 192, 105399. [Google Scholar] [CrossRef]
- Evans, P.L.; McMillin, S.L.; Weyrauch, L.A.; Witczak, C.A. Regulation of skeletal muscle glucose transport and glucose metabolism by exercise training. Nutrients 2019, 11, 2432. [Google Scholar] [CrossRef] [Green Version]
- Scott, K.; Edward, T. Exercise Physiology: Theory and Application to Fitness and Performance Tenth Edition; McGraw-Hill Education: New York, NY, USA, 2019; pp. 302–320. [Google Scholar]
- Heaney, R.P.; Horst, R.L.; Cullen, D.M.; Armas, L.A. Vitamin D3 distribution and status in the body. J. Am. Coll. Nutr. 2009, 28, 252–256. [Google Scholar] [CrossRef] [PubMed]
- Hengist, A.; Perkin, O.; Gonzalez, J.T.; Betts, J.A.; Hewison, M.; Manolopoulos, K.N.; Jones, K.S.; Koulman, A.; Thompson, D. Mobilising vitamin D from adipose tissue: The potential impact of exercise. Nutr. Bull. 2019, 44, 25–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lafontan, M.; Moro, C.; Berlan, M.; Crampes, F.; Sengenes, C.; Galitzky, J. Control of lipolysis by natriuretic peptides and cyclic GMP. Trends Endocrinol. Metab. 2008, 19, 130–137. [Google Scholar] [CrossRef] [PubMed]
- de Glisezinski, I.; Larrouy, D.; Bajzova, M.; Koppo, K.; Polak, J.; Berlan, M.; Bulow, J.; Langin, D.; Marques, M.A.; Crampes, F.; et al. Adrenaline but not noradrenaline is a determinant of exercise-induced lipid mobilization in human subcutaneous adipose tissue. J. Physiol. 2009, 587, 3393–3404. [Google Scholar] [CrossRef]
- Panissa, V.L.G.; Fukuda, D.H.; Staibano, V.; Marques, M.; Franchini, E. Magnitude and duration of excess of post-exercise oxygen consumption between high-intensity interval and moderate-intensity continuous exercise: A systematic review. Obes. Rev. 2021, 22, e13099. [Google Scholar] [CrossRef]
- Newsom, S.A.; Schenk, S.; Thomas, K.M.; Harber, M.P.; Knuth, N.D.; Goldenberg, N.; Horowitz, J.F. Energy deficit after exercise augments lipid mobilization but does not contribute to the exercise-induced increase in insulin sensitivity. J. Appl. Physiol. 1985 2010, 108, 554–560. [Google Scholar] [CrossRef] [Green Version]
- Abboud, M.; Rybchyn, M.S.; Liu, J.; Ning, Y.; Gordon-Thomson, C.; Brennan-Speranza, T.C.; Cole, L.; Greenfield, H.; Fraser, D.R.; Mason, R.S. The effect of parathyroid hormone on the uptake and retention of 25-hydroxyvitamin D in skeletal muscle cells. J. Steroid. Biochem. Mol. Biol. 2017, 173, 173–179. [Google Scholar] [CrossRef]
- Blau, J.E.; Collins, M.T. The PTH-vitamin D-FGF23 axis. Rev. Endocr. Metab. Disord. 2015, 16, 165–174. [Google Scholar] [CrossRef]
- Kägi, L.; Bettoni, C.; Pastor-Arroyo, E.M.; Schnitzbauer, U.; Hernando, N.; Wagner, C.A. Regulation of vitamin D metabolizing enzymes in murine renal and extrarenal tissues by dietary phosphate, FGF23, and 1,25(OH)2D3. PLoS ONE 2018, 13, e0195427. [Google Scholar] [CrossRef]
- Bikle, D.D. Vitamin D metabolism, mechanism of action, and clinical applications. Chem. Biol. 2014, 21, 319–329. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Tang, W.; Zhou, J.; Stubbs, J.R.; Luo, Q.; Pi, M.; Quarles, L.D. Fibroblast growth factor 23 is a counter-regulatory phosphaturic hormone for vitamin D. J. Am. Soc. Nephrol. 2006, 17, 1305–1315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barker, T.; Henriksen, V.T.; Martins, T.B.; Hill, H.R.; Kjeldsberg, C.R.; Schneider, E.D.; Dixon, B.M.; Weaver, L.K. Higher serum 25-hydroxyvitamin D concentrations associate with a faster recovery of skeletal muscle strength after muscular injury. Nutrients 2013, 5, 1253–1275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Li, Y.; Mao, H.; Song, H.; Chen, Y. Effects of lower extremity intensive weight-bearing exercise training on osteoporosis in patients with post-stroke hemiplegia. Chin. J. Gerontol. 2017, 37, 5382–5383. (In Chinese) [Google Scholar]
- Bass, J.J.; Nakhuda, A.; Deane, C.S.; Brook, M.S.; Wilkinson, D.J.; Phillips, B.E.; Philp, A.; Tarum, J.; Kadi, F.; Andersen, D.; et al. Overexpression of the vitamin D receptor (VDR) induces skeletal muscle hypertrophy. Mol. Metab. 2020, 42, 101059. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Ma, X.-K.; Zhang, L.; Cao, Z.-B. Effects of resistance training on serum 25(OH)D concentrations in young men: A randomized controlled trial. Nutr. Metab. 2020, 17, 1–7. [Google Scholar] [CrossRef]
- Aschauer, R.; Unterberger, S.; Zöhrer, P.A.; Draxler, A.; Franzke, B.; Strasser, E.M.; Wagner, K.H.; Wessner, B. Effects of vitamin D3 supplementation and resistance training on 25-hydroxyvitamin D status and functional performance of older adults: A randomized placebo-controlled trial. Nutrients 2021, 14, 86. [Google Scholar] [CrossRef]
- Agergaard, J.; Trostrup, J.; Uth, J.; Iversen, J.V.; Boesen, A.; Andersen, J.L.; Schjerling, P.; Langberg, H. Does vitamin-D intake during resistance training improve the skeletal muscle hypertrophic and strength response in young and elderly men?—A randomized controlled trial. Nutr. Metab. 2015, 12, 32. [Google Scholar] [CrossRef] [Green Version]
- Mason, R.S.; Rybchyn, M.S.; Abboud, M.; Brennan-Speranza, T.C.; Fraser, D.R. The role of skeletal muscle in maintaining vitamin D status in winter. Curr. Dev. Nutr. 2019, 3, nzz087. [Google Scholar] [CrossRef]
- Abboud, M.; Puglisi, D.A.; Davies, B.N.; Rybchyn, M.; Whitehead, N.P.; Brock, K.E.; Cole, L.; Gordon-Thomson, C.; Fraser, D.R.; Mason, R.S. Evidence for a specific uptake and retention mechanism for 25-hydroxyvitamin D (25OHD) in skeletal muscle cells. Endocrinology 2013, 154, 3022–3030. [Google Scholar] [CrossRef] [Green Version]
- Abboud, M.; Gordon-Thomson, C.; Hoy, A.J.; Balaban, S.; Rybchyn, M.S.; Cole, L.; Su, Y.; Brennan-Speranza, T.C.; Fraser, D.R.; Mason, R.S. Uptake of 25-hydroxyvitamin D by muscle and fat cells. J. Steroid. Biochem. Mol. Biol. 2013, 144 Pt A, 232–236. [Google Scholar] [CrossRef]
- Abboud, M.; Rybchyn, M.S.; Ning, Y.J.; Brennan-Speranza, T.C.; Girgis, C.M.; Gunton, J.E.; Fraser, D.R.; Mason, R.S. 1,25-Dihydroxycholecalciferol (calcitriol) modifies uptake and release of 25-hydroxycholecalciferol in skeletal muscle cells in culture. J. Steroid. Biochem. Mol. Biol. 2018, 177, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Girgis, C.M.; Mokbel, N.; Cha, K.M.; Houweling, P.J.; Abboud, M.; Fraser, D.R.; Mason, R.S.; Clifton-Bligh, R.J.; Gunton, J.E. The vitamin D receptor (VDR) is expressed in skeletal muscle of male mice and modulates 25-hydroxyvitamin D (25OHD) uptake in myofibers. Endocrinology 2014, 155, 3227–3237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gustafsson, M.K.; Romundstad, P.R.; Stafne, S.N.; Helvik, A.S.; Stunes, A.K.; Morkved, S.; Salvesen, K.A.; Thorsby, P.M.; Mosti, M.P.; Syversen, U. The effect of an exercise program in pregnancy on vitamin D status among healthy, pregnant Norwegian women: A randomized controlled trial. BMC Pregnancy Childbirth 2019, 19, 76. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Yang, Z.; Han, W.; Tian, X.; Li, Q.; Shuai, S.; Liu, Y. Effects of combined exercise therapy on bone metabolism in patients with postmenopausal osteoporosis. Int. J. Orthop. 2019, 40, 52–62. (In Chinese) [Google Scholar]
- Evans, R.K.; Antczak, A.J.; Lester, M.; Yanovich, R.; Israeli, E.; Moran, D.S. Effects of a 4-month recruit training program on markers of bone metabolism. Med. Sci. Sports Exerc. 2008, 40, 660–670. [Google Scholar] [CrossRef] [PubMed]
- Wortsman, J.; Matsuoka, L.Y.; Chen, T.C.; Lu, Z.; Holick, M.F. Decreased bioavailability of vitamin D in obesity. Am. J. Clin. Nutr. 2000, 72, 690–693. [Google Scholar] [CrossRef]
- Drincic, A.T.; Armas, L.A.; Van Diest, E.E.; Heaney, R.P. Volumetric dilution, rather than sequestration best explains the low vitamin D status of obesity. Obesity (Silver Spring) 2012, 20, 1444–1448. [Google Scholar] [CrossRef]
- Jungert, A.; Neuhäuser-Berthold, M. Sex-specific determinants of serum 25-hydroxyvitamin D3 concentrations in an elderly German cohort: A cross-sectional study. Nutr. Metab. 2015, 12, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Fiamenghi, V.I.; Mello, E.D. Vitamin D deficiency in children and adolescents with obesity: A meta-analysis. J. Pediatr. 2021, 97, 273–279. [Google Scholar] [CrossRef]
- Vranić, L.; Mikolašević, I.; Milić, S. Vitamin D Deficiency: Consequence or Cause of Obesity? Medicina 2019, 55, 541. [Google Scholar] [CrossRef] [Green Version]
- Di Nisio, A.; De Toni, L.; Sabovic, I.; Rocca, M.S.; De Filippis, V.; Opocher, G.; Azzena, B.; Vettor, R.; Plebani, M.; Foresta, C. Impaired release of vitamin D in dysfunctional adipose tissue: New cues on vitamin D supplementation in obesity. J. Clin. Endocrinol. Metab. 2017, 102, 2564–2574. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, L.; Hachemi, M.A.; Karkeni, E.; Couturier, C.; Astier, J.; Defoort, C.; Svilar, L.; Martin, J.C.; Tourniaire, F.; Landrier, J.F. Diet induced obesity modifies vitamin D metabolism and adipose tissue storage in mice. J. Steroid. Biochem. Mol. Biol. 2019, 185, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Vieira, A.F.; Costa, R.R.; Macedo, R.C.; Coconcelli, L.; Kruel, L.F. Effects of aerobic exercise performed in fasted v. fed state on fat and carbohydrate metabolism in adults: A systematic review and meta-analysis. Br. J. Nutr. 2016, 116, 1153–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aird, T.P.; Davies, R.W.; Carson, B.P. Effects of fasted vs fed-state exercise on performance and post-exercise metabolism: A systematic review and meta-analysis. Scand. J. Med. Sci. Sports 2018, 28, 1476–1493. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Possible mechanisms of the effect of endurance training on serum 25(OH)D. Vitamin D indicates serum vitamin D metabolites; − indicates no significant change; ↑ indicates significant increase; and ↓ indicates significantly reduction. Vitamin D status: 25(OH)D ≥ 20 ng/mL or 50 nmol/L; vitamin D deficiency: 25(OH)D < 20 ng/mL or 50 nmol/L. Abbreviations: VDR, vitamin D receptor.
Figure 1.
Possible mechanisms of the effect of endurance training on serum 25(OH)D. Vitamin D indicates serum vitamin D metabolites; − indicates no significant change; ↑ indicates significant increase; and ↓ indicates significantly reduction. Vitamin D status: 25(OH)D ≥ 20 ng/mL or 50 nmol/L; vitamin D deficiency: 25(OH)D < 20 ng/mL or 50 nmol/L. Abbreviations: VDR, vitamin D receptor.

Figure 2.
Possible mechanisms of the effect of resistance training on serum 25(OH)D. Vitamin D indicates serum vitamin D metabolites; − indicates no significant change; ↑ indicates significant increase; and ↓ indicates significantly reduction. Regulation factors include VDR, PTH, DBP, and vitamin D nutritional status. 25(OH)D-1α hydroxylase, CYP27B1, can convert 25(OH)D to 1,25(OH)2D. Vitamin D 24-hydroxylase, CYP24A1, is an important degrading enzyme of vitamin D. Abbreviations: VDR, vitamin D receptor.
Figure 2.
Possible mechanisms of the effect of resistance training on serum 25(OH)D. Vitamin D indicates serum vitamin D metabolites; − indicates no significant change; ↑ indicates significant increase; and ↓ indicates significantly reduction. Regulation factors include VDR, PTH, DBP, and vitamin D nutritional status. 25(OH)D-1α hydroxylase, CYP27B1, can convert 25(OH)D to 1,25(OH)2D. Vitamin D 24-hydroxylase, CYP24A1, is an important degrading enzyme of vitamin D. Abbreviations: VDR, vitamin D receptor.

Figure 3.
The effect of exercise on 25(OH)D in circulation.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary of the effect of acute endurance exercise intervention.
| Study | Participants/Animal, n | Endurance Exercise Intervention | Sunlight Exposure | Main Findings |
|---|---|---|---|---|
| Acute endurance exercise intervention-human studies | ||||
| Mieszkowski (2020) [32] | Experimental, n = 13, 42.00 ± 8.44 years old, Ultra-Marathon Race, 150,000 vitamin D3; Control, n = 14, 40.00 ± 8.11 years old, Ultra-Marathon Race, placebo solution | Ultra-Marathon Race | 18:00 h, 19 July; most of the time, the sky was overcast | 25(OH)D3: significantly increased immediately and 24 h after the ultra-marathon vs. 24 h before the ultra-marathon in both groups |
| Dzik (2022) [33] | Male soccer players, n = 12 (pre-pubertal, n = 5; pubertal, n = 7) | VO2max test | - | 25(OH)D3: significantly increased at 15 min and 1 h after exercise vs. before; increased 15 min after the VO2max test and dropped one hour after exercise, but not significantly different. |
| Maimoun (2006) [34] | Male competitive road cyclists, n = 7, 20–30 years old | 47% Wmax; 64% Wmax | - | 25(OH)D: no change 1,25(OH)2D: no change |
| Maimoun (2009) [35] | Elderly moderately active (ModEl, n = 18), 71.9 ± 7.3 years old; Elderly active (HAcEl; n = 18), 71.7 ± 8.6 years old; Young active (AcYo; n = 9), 25.8 ± 2.3 years old | maximal incremental exercise | - | 25(OH)D: significantly increased in HAcEl, but not in ModEl and AcYo 1,25(OH)2D: no change |
| Sun (2017) [36] | Healthy young men, n = 10, 18–22 years old; Healthy young women, n = 10, 19–22 years old | cycling exercise for 30 min at 70% VO2max | at the laboratory | 25(OH)D: significantly greater at 0 h, 1 h, 3 h and 24 h after exercise vs. before exercise; subgroup analysis: significantly increased at 24 h after exercise in women only 1,25(OH)2D: no change |
| Acute endurance exercise intervention: animal studies | ||||
| Makanae (2015) [37] | Adult male Sprague–Dawley rats, 10 weeks age | 60 min, 25 m/min | at the laboratory | 25(OH)D3: no change |
| Puangthong (2021) [38] | Healthy ponies, n = 6 (5 geldings, 1 mare), 6.3 ± 2.2 years age | 77–93% of HRmax, 16.5 ± 1 min, 5.2 ± 0.3 km | at the laboratory | 25(OH)D2: significantly reduced at 30 min, 1 week, and 3 weeks after high-intensity exercise |
| Chronic endurance exercise intervention-human studies | ||||
| Farag (2019) [39] | Vitamin D plus PA group: n = 21, 40.42 ± 5.89 years old, 2000IU/day, endurance PA | Endurance PA: 12 weeks, daily endurance PA, 30 min/day | Either at morning, 7:30 a.m. or afternoon after 3:00 p.m. | 25(OH)D: significantly increased |
| Mieszkowski (2018) [40] | High-intensity interval training group (HI-NW): LD (n = 8, 67.37 ± 6.30 years old, 800 IU/day vitamin D3), and HD (n = 8, 67.63 ± 7.29 years old, 4000 IU/day vitamin D3); Moderate-inteensity continuous training group (MI-NW): LD (n = 13, 69.08 ± 4.87 years old, 800 IU/day vitamin D3) and HD (n = 13, 70.85 ± 4.61 years old, 4000 IU/day vitamin D3) | Nordic walking training: 12 weeks, two hours, three times a week. HI-NW: 30 s acceleration going uphill,60 s release going downhill for eight time; 70% HRmax for 28 min. MI-NW: 60–70 HRmax for 40 min | morning hours | 25(OH)D3: significantly increased in HI-NW with LD and HD group and MI-NW with HD group; no change in MI-NW with LD group. |
| Prusik (2018) [41] | Experimental group (EG), n = 35, 68.4 ± 5.0 years old | EG: Nordic walking training, 12 weeks, three times a week, 60–70% HRmax for 45–55 min; 4000 IU/day vitamin D supplement | 1 h after breakfast | 25(OH)D3: significantly increased after 12 weeks of Nordic walking training with vitamin D supplementation; no change after 6 months without training and vitamin D supplementation |
| Malandish (2020) [42] | Postmenopausal women Exercise group (EX), n = 13, 53.36 ± 3.98 years old; Control group (C), n = 13, 53.00 ± 3.26 years old | EX: 12 weeks training, 3 sessions per week, 55–60 min per session, 40 min of walking or jogging aerobic exercise on treadmill C: no intervention | - | 25(OH)D: significantly increased after exercise vs. before exercise in EX group and compared to C group; no change in C group |
| Li (2019) [43] | elderly chronic obstructive pulmonary disease patients with osteoporosis, 65–82 years old Experimental group, n = 31; Control group, n = 31 | Experimental group: 12 weeks, 4 times/week, 5 set/session, 5 min/set, 5 min between sets, 75% CPET, 25 min/session. Control group: 12 weeks, 4 times/week, 5 set/session, 5 min/set, 5 min between sets, 50% CPET, 25 min/session. | - | 25(OH)D: significantly increased after exercise in experimental group and control group; significantly increased after exercise in experimental group vs. control group after exercise intervention |
| Song (2014) [44] | postmenopausal women with type II diabetes and osteoporosis Experimental group: n = 278, 52.82 ± 5.12 years old; Control group: n = 284, 53.26 ± 5.12 years old | Experimental group: 48 weeks, moderate intensity, 20–30 min/time, two times/day, 0.25 ug/day Calcitriol and 600 mg vitamin D supplementation Control group: 0.25 ug/day Calcitriol and 600 mg vitamin D supplementation | - | 25(OH)D: significantly increased 24 weeks and 48 weeks after exercise vs. before exercise in experimental group and higher than control group at same time points |
| Shi (2013) [45] | Patients with osteoporosis, 50–89 years old, n = 82 exercise group (n = 40); control group (n = 42) | exercise group: Wu xing Bone gymnastics, 90 days, 30–45 min/time, two times/day control group: calcium and Calcitriol supplementation | - | 25(OH)D: significantly increased after exercise intervention vs. before exercise intervention in exercise group; no change in control group |
| Klausen (1993) [46] | Male marathon runners, n = 9, 41–50 years old | Endurance training: median running distance was 61 km per week, 4 weeks | the months of December and January | 25(OH)D3: no change at 2 week and 4 week retraining. 1,25(OH)2D3: significantly reduced at 4 week retraining vs. before retraining |
| Pilch (2017) [47] | Women, n = 17, 57 ± 4.20 years old | Nordic walking training, 6 weeks, three times a week, 90 min/time, 60–70% HRmax. | morning hours | 25(OH)D: significantly reduced after exercise intervention |
| Lithgow (2018) [48] | Overweight and obese adults Placebo group: n = 10, 34 ± 10 years old; Vitamin D group: n = 10, 34 ± 9 years old | Placebo group: HIIT intervention, 6 weeks, 3 sessions/week, 10 repetitions of 1 min intervals interspersed with 1 min active recovery at a power output of 50 W. placebo tablets Vitamin D group: HIIT with 4000 IU/day vitamin D3 | - | 25(OH)D3: significantly increased in vitamin D group than placebo group; no change between before and after exercise in placebo group |
| Hossain (2018) [49] | Intervention group: n = 7, 14–18 years old; Control group: n = 7, 14–18 years old | Intervention group: brisk walking, 12 weeks, 45 min/time, three times a week Control group: no change routine lifestyle | - | 25(OH)D: no change in both groups |
| Sun (2018) [50] | The 5-week endurance exercise training group (ET group), n = 10, 66.5–75.3 years old; Sedentary control group (SC group), n = 10, 63.8–73.0 years old | ET group: aerobic exercise, 5 weeks, three times per week, 60% VO2max during week 1, 70% during weeks 2 and 3, and 75% during weeks 4 and 5, 30 min for weeks 1 and 2, and 45 min for weeks 3–5 SC group: no intervention | From October to November | 25(OH)D: significantly reduced after exercise in SC group; no change in ET group |
| Chronic endurance exercise intervention: animal studies | ||||
| Aly (2016) [51] | Adult male albino, Group I(a): control sedentary, n = 15; Group I(b): control exercised, n = 15; Group II(a): diabetic sedentary, n = 15; Group II(b): diabetic exercised, n = 15 | Group I(b) and Group II(b): swimming moderate exercise, 4 weeks, 60 min/time, 5 time per week Group I(a) and Group II(a): no intervention | at the laboratory | 25(OH)D: significantly increased in Group II(b) vs. Group II(a); no change between Group I(a) and Group I(b) |
| Buskermolen (2019) [52] | Female wistar rat, 13 weeks old Control group, n = 8; Endurance training group (ET), n = 10 | ET: treadmill running, 6 weeks, 10 min at a speed of 16 m/min without a slope, increased up to 45 min with a speed of 26 m/min on a 10% slope Control group: no intervention | at the laboratory | 25(OH)D: no change between ET and control group |
| Yeh (1989) [53] | Female Sprague-Dawley rats, 75 ± 5 g Exercise group; Pair-fed exercise group; control group; | Exercise group and Pair-fed exercise group: flat-bed treadmill running, 13 weeks, 60 min/time, 5 times per week, 18–25 m/min Control group: no intervention | at the laboratory | 25(OH)D: no change in the three groups 1,25(OH)2D3: significantly increased in Exercise group and Pair-fed exercise group vs. control |
| Iwamoto (2004) [54] | Female Wistar rats, 6 weeks old, n = 20 7 weeks of exercise (7EX), n = 5; 7 weeks of sedentary control (7CON), n = 5; 11 weeks of exercise (11EX), n = 5; 11 weeks of sedentary control (11CON), n = 5 | 7EX and 11EX: running on flat-bed treadmill, 7 weeks or 11 weeks, 60 min/time, 5 time a week 7CON and11CON: no intervention | at the laboratory | 1,25(OH)2D3: significantly increased in 7EX vs. 7CON; significantly increased in 11EX than 11CON |
| Wang (2018) [55] | Male F344 rats Sedentary young rats (Young), n = 9; Sedentary aged rats (Aged), n = 9; Aged rats with aerobic exercise training (Aged + EX), n = 9 | Aged + EX: running treadmill, 12 weeks, 7 times per week, 1 h/time, 10% slope, 8–20 m/min Young and Aged group: no intervention | at the laboratory | 1,25(OH)2D3: slightly increased, not significant |
| Xu (2019) [56] | C57BL/6 male mice, 5 weeks old Swimming group (group S), n = 7; Downhill running group (group R), n = 7; Control (group C), n = 7 | group S: swimming training, 8 weeks, 6 times per week, 50 min/time, 65–70%VO2max group R: downhill running, 8 weeks, 6 times per week, 50 min/time, −9% slope, 0.8 km/h group C: no intervention | at the laboratory | 1,25(OH)2D3: significantly reduced in group S and group R vs. group C |
- Indicates no relevant information. Wmax indicates maximal workload; VO2max indicates maximal oxygen uptake. HRmax indicates maximal heart rate. Abbreviations: PA indicates physical activity; HR indicates heart rate; NW indicates Nordic walking training; CPET indicates cardiopulmonary exercise test; and HIIT indicates high-intensity intermittent training.
Table 2.
Summary of the effect of resistance exercise intervention (human study and animal study).
| Study | Participants/Animal, n | Resistance Exercise Intervention | Sunlight Exposure | Main Findings |
|---|---|---|---|---|
| Acute resistance exercise intervention: human study | ||||
| Barker (2013) [74] | Recreationally active subjects Intense-stretch shortening contraction leg (SSC); Control leg (CON) | SSC: 10 sets of 10 jumps with a 20-s rest between each set at 75% of body mass on one leg only CON: no intervention | December to March; at the laboratory | 25(OH)D: significantly increased immediately after acute resistance exercise; decreased after 24, 48, 72, and 168 h |
| Acute resistance exercise intervention: animal study | ||||
| Makanae (2015) [37] | Male Sprague-Dawley, 10 weeks old | Isometrically exercise, five sets of ten 3 s contractions, with a 7 s interval between contractions and 3 min rest intervals between sets | at the laboratory | 25(OH)D3: no change |
| Chronic resistance exercise intervention: human study | ||||
| Zhang (2017) [75] | patients with post-stroke hemiplegia, 59.58 ± 4.39 years old Experimental group, n = 25; Control group, n = 25 | Experimental group: weight-bearing exercise training, one year, 40 min/time, two times/day. Routine rehabilitation. Calcium and calciferol supplement Control group: Routine rehabilitation. Calcium and calciferol supplement | - | 25(OH)D: significantly increased at 3 months and 1 year of intervention in Experimental group vs. before intervention and vs. control group at same time points. |
| Bass (2020) [76] | Male and female healthy participants, n = 37, 48.4 ± 2.6 years old | 20 weeks, three times a week, 70% 1 repetition max, single sets of 12 repetitions with 2-min rests between sets of seated chest press, lat pull down, seated lever row, leg extension, seated leg curl, seated leg press, back extension and abdominal curls | - | 25(OH)D: significantly increased after exercise intervention |
| Sun (2020) [77] | healthy men, n = 18, 19–39 years old resistance training group (RT), n = 9, 24.2 ± 3.1 years old; non-exercise control group (CON), n = 9, 26.7 ± 6.2 years old | RT: progressive resistance training, 12 weeks, 2–3 times per week, resistance workload gradually changed from light to heavy CON: no intervention | From March to July, Between 16:30 h and 20:00 h in a gymnasium | 25(OH)D: significantly increased after 12 weeks of exercise intervention vs. baseline in both groups; significantly higher at 6 weeks compared with the values at baseline in the CON group, whereas no notable differences were found in the RT group |
| Aschauer (2021) [78] | Older adults, n = 85, 65–85 years old Control group (CON), Placebo, 400 mg calcium/day; Vitamin D3 daily group (VDD), 800 IU vitamin D3/day, 400 mg calcium/day; Vitamin D3 monthly group (VDM), 50,000 IU vitamin D3/month, 400 mg calcium/day | Three groups have conducted Resistance training: 10 weeks, twice a week, 60–90 min/session | From mid-February to mid-July | 25(OH)D: no change in CON; significantly increased in both VDD and VDM |
| Agergaard (2015) [79] | Healthy sedentary young and elderly men Young vitamin D group, n = 7, 23.3 ± 2.0 years old; Young placebo group, n = 10, 22.4 ± 1.8 years old; elderly vitamin D group, n = 7, 67.1 ± 2.9 years old ; elderly placebo group, n = 10, 66.6 ± 4.2 years old | Four groups have conducted resistance training exercise: 12 weeks, 3 sessions/week, Progressive loading levels | From November to April | 25(OH)D: significantly reduced at 0, 2, 6, and 12 weeks in young placebo group vs. at −4 weeks; significantly reduced at 0, 6, and 12 weeks in young placebo group vs. at −4 weeks; significantly increased at 0, 2, 6, and 12 weeks in young vitamin D group and elderly vitamin D group vs. at −4 weeks |
| Acute resistance exercise intervention: animal studies | ||||
| Buskermolen (2019) [52] | Female wistar rat, 13 weeks old peak power training (PT), n = 10; Control group, n = 8 | PT: peak power training, 10 sprints of 15 s in gallop at a maximal attainable velocity on a progressively increasing slope starting at 10% reaching up to 40% by the end Control group: no intervention | at the laboratory | 25(OH)D: no change |
| Xu (2019) [56] | C57BL/6 male mice, 5 weeks old Jumping group (group J), n = 7; Control group (group C), n = 7 | Group J: jumping training, 8 weeks, 6 times per week, 6–7 sets/min, 50 min/time Group C: no intervention | at the laboratory | 1,25(OH)2D3: significantly reduced in group J vs. group C |
- Indicates no relevant information.
Table 3.
Summary of the effect of endurance combined with resistance exercise intervention (human studies and animal study).
Table 3.
Summary of the effect of endurance combined with resistance exercise intervention (human studies and animal study).
| Study | Participants/Animal, n | Endurance Exercise Intervention | Sunlight Exposure | Main Findings |
|---|---|---|---|---|
| Endurance combined with resistance exercise intervention: human studies | ||||
| Gustafsson (2019) [85] | healthy, pregnant Norwegian women Intervention group: n = 429, 30.5 ± 4.4 years old; Control group: n = 426, 30.4 ± 4.3 years old | Intervention group: aerobic and strength training, 12 weeks, 3 times per week, 60 min/time | - | 25(OH)D: no significant effect of the exercise program on levels of total, free, or bioavailable 25(OH)D in only baseline level adjust model; additionally adjusted for study site and sampling month, revealed a significant between-group difference in levels of total, free, and bioavailable 25(OH)D. |
| Li (2019) [86] | Patients with postmenopausal osteoporosis Training group: n = 26, 55.46 ± 4.12 years old; Control group: n = 26, 56.25 ± 3.75 years old | Training group: 12 weeks, (a) endurance exercise training, brisk walk outdoors, 4 times per week, 30 min/ time, 50%VO2max; (b) progressive resistance training. calcium and Calcitriol supplementation Control group: calcium and Calcitriol supplementation | brisk walk outdoors | 25(OH)D: significantly increased after intervention in both groups; significantly increased in Training group vs. control group |
| Evans [87] | Healthy men, n = 41, 19.3 ± 1.2 years old; Healthy women, n = 153, 19.0 ± 1,0 years old | Marching under load, running and jumping, battle drills, and walking and standing for prolonged periods of time | - | 25(OH)D: significantly reduced at 4 months in male participants; no change in female participants |
| Endurance combined with resistance exercise intervention-animal study | ||||
| Buskermolen [52] | Female wistar rat, 13 weeks old peak power training and endurance training group, n = 10; Control group, n = 8 | Peak power training: 10 sprints of 15 s in gallop at a maximal attainable velocity on a progressively increasing slope starting at 10% reaching up to 40% by the end endurance training: treadmill running, 6 weeks, 10 min at a speed of 16 m/min without a slope, increased up to 45 min with a speed of 26 m/min on a 10% slope Control group: no intervention | - | 25(OH)D: no change |
- Indicates no relevant information.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhang, J.; Cao, Z.-B. Exercise: A Possibly Effective Way to Improve Vitamin D Nutritional Status. Nutrients 2022, 14, 2652. https://0-doi-org.brum.beds.ac.uk/10.3390/nu14132652
AMA Style
Zhang J, Cao Z-B. Exercise: A Possibly Effective Way to Improve Vitamin D Nutritional Status. Nutrients. 2022; 14(13):2652. https://0-doi-org.brum.beds.ac.uk/10.3390/nu14132652
Chicago/Turabian StyleZhang, Jinghua, and Zhen-Bo Cao. 2022. "Exercise: A Possibly Effective Way to Improve Vitamin D Nutritional Status" Nutrients 14, no. 13: 2652. https://0-doi-org.brum.beds.ac.uk/10.3390/nu14132652
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.