Maternal Betaine Supplementation throughout Gestation and Lactation Modifies Hepatic Cholesterol Metabolic Genes in Weaning Piglets via AMPK/LXR-Mediated Pathway and Histone Modification

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Sampling
2.2. Measurement of Cholesterol and Bile Acid Concentrations
2.3. Determination of Serum Betaine Content and Hepatic S-Adenosyl Methionine and S-Adenosyl Homocysteine Concentrations
2.4. Serum Hormones and Amino Acids Profile
2.5. Real-Time PCR for mRNA Quantification
2.6. Western Blotting for Protein Quantification
2.7. Chromatin Immunoprecipitation
2.8. Statistical Analysis
3. Results
3.1. Serum Concentrations of Metabolites in Weaning Piglets
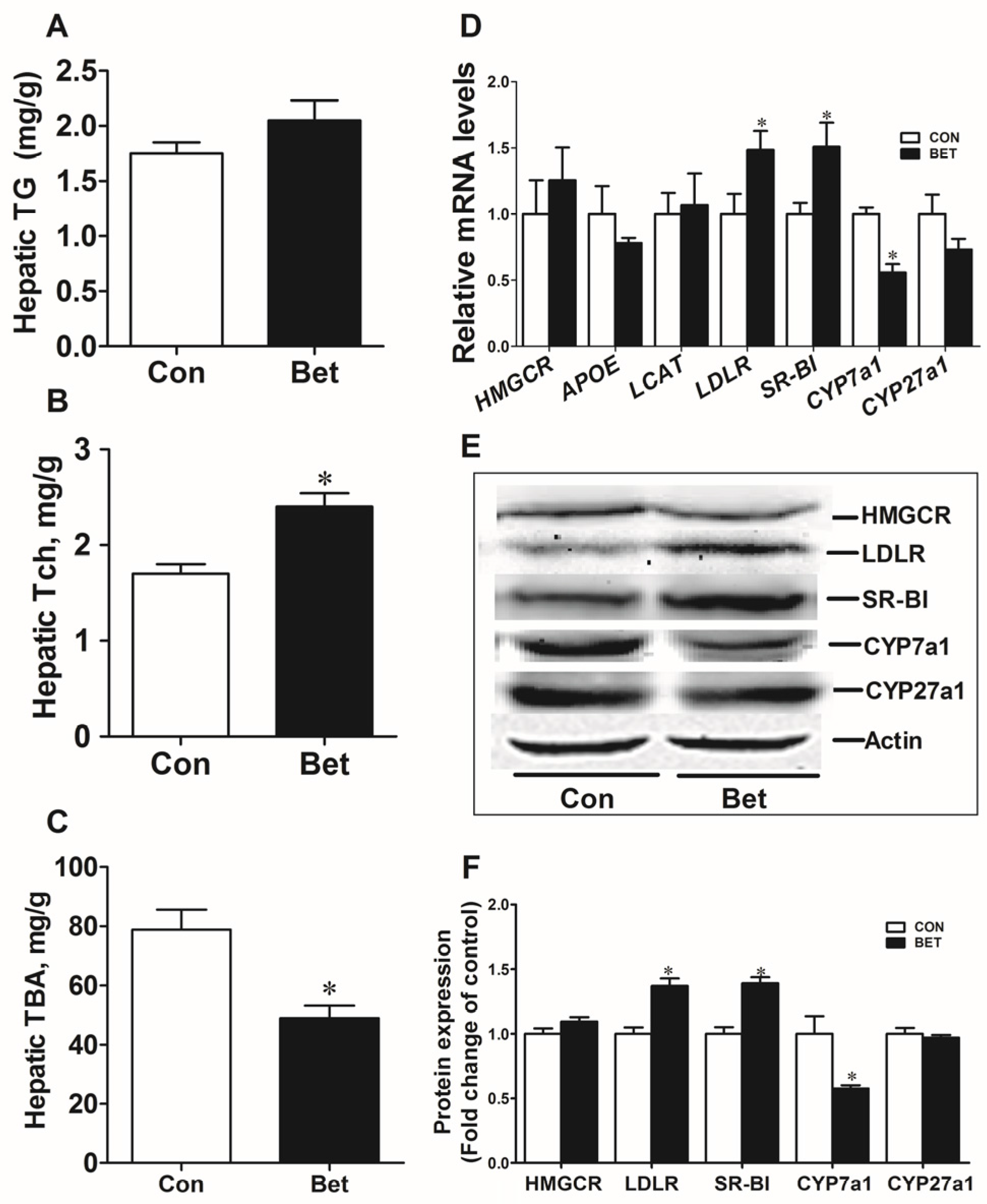
3.2. Cholesterol Metabolism in Weaning Piglets
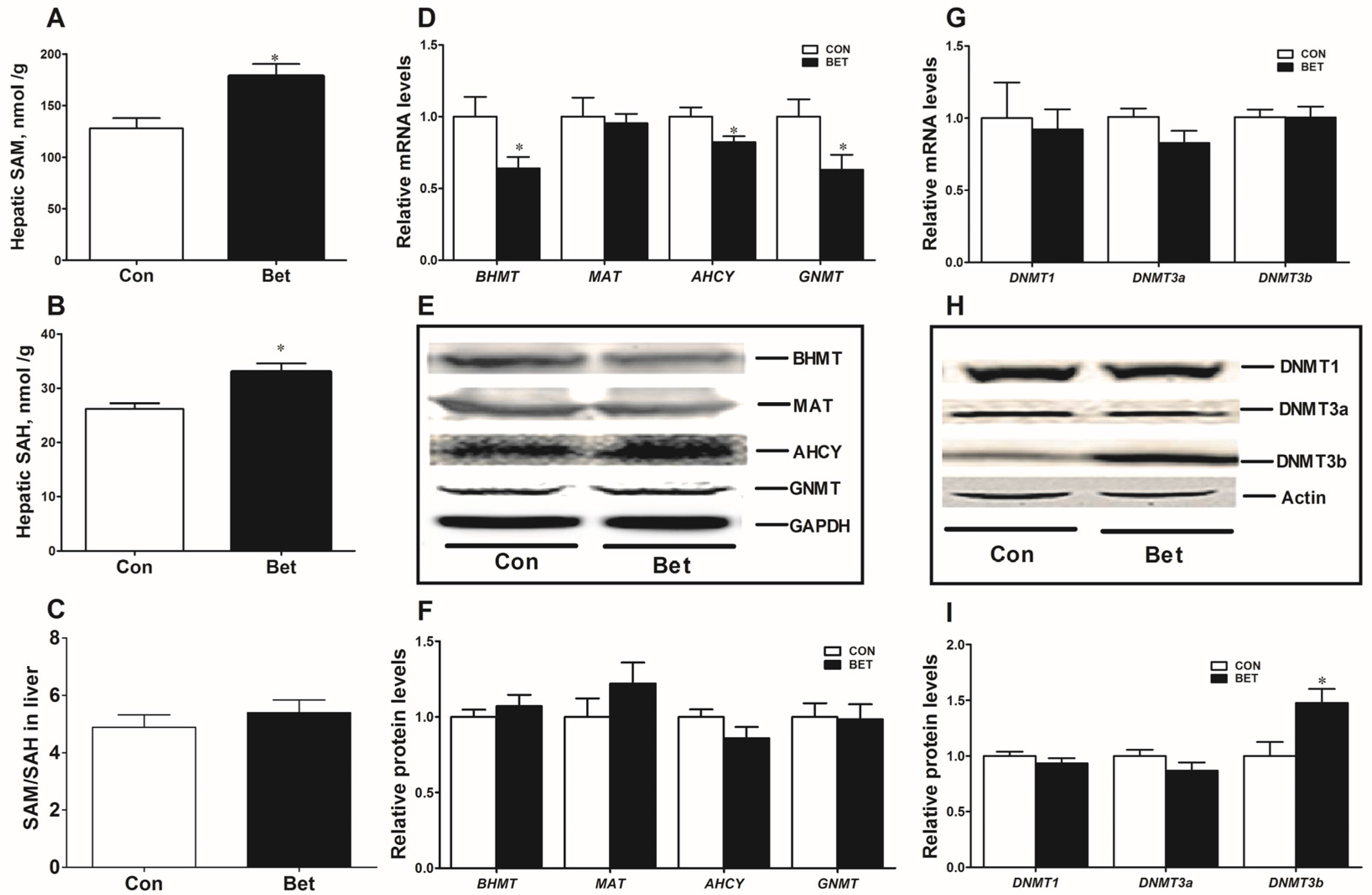
3.3. Methionine Metabolism in Weaning Piglets
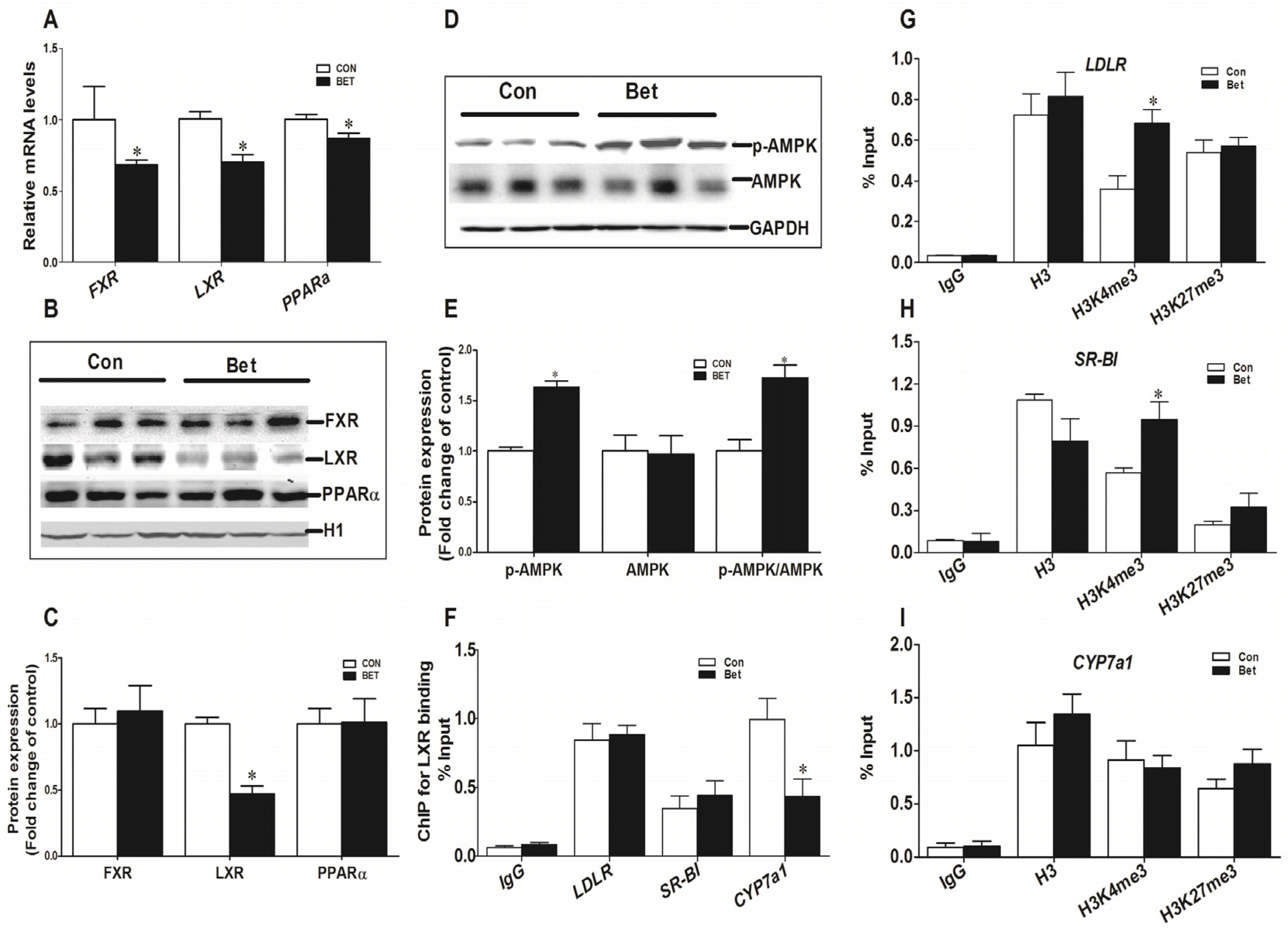
3.4. Transcriptional Regulation of Cholesterol Metabolic Genes
4. Discussion
Acknowledgments
Author Contributions
Conflicts of interest
Abbreviations
| AHCY | S-adenosylhomocysteine hydrolase |
| AMPK | AMP-activated protein kinase |
| Bet | piglets born to sows supplemented with betaine diets |
| BHMT | betaine-homocysteine S-methyltransferase |
| BW | body weight |
| Con | piglets born to sows supplemented with normal diets |
| CYP7α1 | cholesterol-7alpha-hydroxylase |
| CYP27α1 | cholesterol-27alpha-hydroxylase |
| DNMTs | DNA methyltransferases |
| FXR | farnesoid X receptor |
| GNMT | glycine N-methyltransferase |
| H1 | histone H1 |
| H3 | histone H3 |
| H3K4me3 | histone H3 lysine 4 trimethylation |
| H3K27me3 | histone H3 lysine 27 trimethylation |
| HDLR | high-density lipoprotein receptor |
| HMGCR | 3-hydroxy-3-methylglutaryl coenzyme A reductase |
| LDLR | low density lipoprotein receptor |
| LXR | liver X receptor |
| Met | Methionine |
| NR | Nuclear receptors |
| PPARα | peroxisome proliferator-activated receptor alpha |
| SAM | S-adenosylmethionine |
| SAH | S-adenosylhomocysteine |
| SETD7 | SET Domain Containing Protein 7 |
| SR-BI | scavenger receptor class B type I |
| Tch | total cholesterol |
| TBA | total bile acids |
| TMG | trimethylglycine |
References
- Lever, M.; Slow, S. The clinical significance of betaine, an osmolyte with a key role in methyl group metabolism. Clin. Biochem. 2010, 43, 732–744. [Google Scholar] [CrossRef] [PubMed]
- Barak, A.J.; Beckenhauer, H.C.; Badakhsh, S.; Tuma, D.J. The effect of betaine in reversing alcoholic steatosis. Alcohol. Clin. Exp. Res. 1997, 21, 1100–1102. [Google Scholar] [CrossRef] [PubMed]
- Graf, D.; Kurz, A.K.; Reinehr, R.; Fischer, R.; Kircheis, G.; Haussinger, D. Prevention of bile acid-induced apoptosis by betaine in rat liver. Hepatology 2002, 36, 829–839. [Google Scholar] [CrossRef] [PubMed]
- Ji, C.; Kaplowitz, N. Betaine decreases hyperhomocysteinemia, endoplasmic reticulum stress, and liver injury in alcohol-fed mice. Gastroenterology 2003, 124, 1488–1499. [Google Scholar] [CrossRef]
- Li, M.; Reynolds, C.M.; Segovia, S.A.; Gray, C.; Vickers, M.H. Developmental programming of nonalcoholic fatty liver disease: The effect of early life nutrition on susceptibility and disease severity in later life. BioMed Res. Int. 2015, 2015, 437107. [Google Scholar] [CrossRef] [PubMed]
- Trapani, L.; Segatto, M.; Pallottini, V. Regulation and deregulation of cholesterol homeostasis: The liver as a metabolic “power station”. World J. Hepatol. 2012, 4, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Ness, G.C. Physiological feedback regulation of cholesterol biosynthesis: Role of translational control of hepatic HMG-CoA reductase and possible involvement of oxylanosterols. Biochim. Biophys. Acta 2015, 1851, 667–673. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Liu, T.; Chang, L.; Rui, C.; Xiao, Y.; Li, S.; Hogenesch, J.B.; Chen, Y.E.; Lin, J.D. The liver clock controls cholesterol homeostasis through Trib1 protein-mediated regulation of PCSK9/Low density lipoprotein receptor (LDLR) axis. J. Biol. Chem. 2015, 290, 31003–31012. [Google Scholar] [CrossRef] [PubMed]
- Calabresi, L.; Gomaraschi, M.; Simonelli, S.; Bernini, F.; Franceschini, G. HDL and atherosclerosis: Insights from inherited HDL disorders. Biochim. Biophys. Acta 2015, 1851, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Dias, V.; Ribeiro, V. Ethnic differences in the prevalence of polymorphisms in CYP7A1, CYP7B1 and CYP27A1 enzymes involved in cholesterol metabolism. J. Pharm. Bioallied Sci. 2011, 3, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Liang, G.; Ou, J.; Goldstein, J.L.; Brown, M.S. Central role for liver X receptor in insulin-mediated activation of Srebp-1c transcription and stimulation of fatty acid synthesis in liver. Proc. Natl. Acad. Sci. USA 2004, 101, 11245–11250. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.Y.; Aleksunes, L.M.; Tanaka, Y.; Fu, Z.D.; Guo, Y.; Guo, G.L.; Lu, H.; Zhong, X.B.; Klaassen, C.D. Bile acids via FXR initiate the expression of major transporters involved in the enterohepatic circulation of bile acids in newborn mice. Am. J. Phys. Gastrointest. Liver Phys. 2012, 302, G979–G996. [Google Scholar] [CrossRef] [PubMed]
- Musso, G.; Gambino, R.; Cassader, M. Recent insights into hepatic lipid metabolism in non-alcoholic fatty liver disease (NAFLD). Prog. Lipid Res. 2009, 48, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Reue, K.; Fong, L.G.; Young, S.G.; Tontonoz, P. Feedback regulation of cholesterol uptake by the LXR-IDOL-LDLR axis. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 2541–2546. [Google Scholar] [CrossRef] [PubMed]
- Grefhorst, A.; Oosterveer, M.H.; Brufau, G.; Boesjes, M.; Kuipers, F.; Groen, A.K. Pharmacological LXR activation reduces presence of SR-B1 in liver membranes contributing to LXR-mediated induction of HDL-cholesterol. Atherosclerosis 2012, 222, 382–389. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, J.M.; Kliewer, S.A.; Moore, L.B.; Smith-Oliver, T.A.; Oliver, B.B.; Su, J.L.; Sundseth, S.S.; Winegar, D.A.; Blanchard, D.E.; Spencer, T.A.; et al. Activation of the nuclear receptor LXR by oxysterols defines a new hormone response pathway. J. Biol. Chem. 1997, 272, 3137–3140. [Google Scholar] [CrossRef] [PubMed]
- Tobin, K.A.; Steineger, H.H.; Alberti, S.; Spydevold, O.; Auwerx, J.; Gustafsson, J.A.; Nebb, H.I. Cross-talk between fatty acid and cholesterol metabolism mediated by liver X receptor-alpha. Mol. Endocrinol. 2000, 14, 741–752. [Google Scholar] [PubMed]
- Milona, A.; Owen, B.M.; Van Mil, S.; Dormann, D.; Mataki, C.; Boudjelal, M.; Cairns, W.; Schoonjans, K.; Milligan, S.; Parker, M.; et al. The normal mechanisms of pregnancy-induced liver growth are not maintained in mice lacking the bile acid sensor FXR. Am. J. Phys. Gastrointest. Liver Phys. 2010, 298, G151–G158. [Google Scholar] [CrossRef] [PubMed]
- Gadaleta, R.M.; Cariello, M.; Sabba, C.; Moschetta, A. Tissue-specific actions of FXR in metabolism and cancer. Biochim. Biophys. Acta 2015, 1851, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Williams, K.T.; Schalinske, K.L. Homocysteine metabolism and its relation to health and disease. BioFactors 2010, 36, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Finkelstein, J.D. Metabolic regulatory properties of S-adenosylmethionine and S-adenosylhomocysteine. Clin. Chem. Lab. Med. 2007, 45, 1694–1699. [Google Scholar] [CrossRef] [PubMed]
- Caudill, M.A.; Wang, J.C.; Melnyk, S.; Pogribny, I.P.; Jernigan, S.; Collins, M.D.; Santos-Guzman, J.; Swendseid, M.E.; Cogger, E.A.; James, S.J. Intracellular S-adenosylhomocysteine concentrations predict global DNA hypomethylation in tissues of methyl-deficient cystathionine beta-synthase heterozygous mice. J. Nutr. 2001, 131, 2811–2818. [Google Scholar] [PubMed]
- Yang, J.; Craddock, L.; Hong, S.; Liu, Z.M. Amp-activated protein kinase suppresses LXR-dependent sterol regulatory element-binding protein-1c transcription in rat hepatoma MCA-RH7777 cells. J. Cell. Biochem. 2009, 106, 414–426. [Google Scholar] [CrossRef] [PubMed]
- Dahlhoff, C.; Worsch, S.; Sailer, M.; Hummel, B.A.; Fiamoncini, J.; Uebel, K.; Obeid, R.; Scherling, C.; Geisel, J.; Bader, B.L.; et al. Methyl-donor supplementation in obese mice prevents the progression of NAFLD, activates AMPK and decreases acyl-carnitine levels. Mol. Metab. 2014, 3, 565–580. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, J.; Li, R.; Yang, X.; Sun, Q.; Albrecht, E.; Zhao, R. Maternal dietary protein affects transcriptional regulation of myostatin gene distinctively at weaning and finishing stages in skeletal muscle of meishan pigs. Epigenetics 2011, 6, 899–907. [Google Scholar] [CrossRef] [PubMed]
- Molloy, A.M.; Mills, J.L.; Cox, C.; Daly, S.F.; Conley, M.; Brody, L.C.; Kirke, P.N.; Scott, J.M.; Ueland, P.M. Choline and homocysteine interrelations in umbilical cord and maternal plasma at delivery. Am. J. Clin. Nutr. 2005, 82, 836–842. [Google Scholar] [PubMed]
- Ramis, G.; Evangelista, J.N.B.; Quereda, J.J.; Pallares, F.J.; de la Fuente, J.M.; Munoz, A. Use of betaine in gilts and sows during lactation: Effects on milk quality, reproductive parameters, and piglet performance. J. Swine Health Prod. 2011, 19, 226–232. [Google Scholar]
- Dominguez-Salas, P.; Moore, S.E.; Cole, D.; da Costa, K.A.; Cox, S.E.; Dyer, R.A.; Fulford, A.J.; Innis, S.M.; Waterland, R.A.; Zeisel, S.H.; et al. DNA methylation potential: Dietary intake and blood concentrations of one-carbon metabolites and cofactors in rural african women. Am. J. Clin. Nutr. 2013, 97, 1217–1227. [Google Scholar] [CrossRef] [PubMed]
- Storch, K.J.; Wagner, D.A.; Young, V.R. Methionine kinetics in adult men: Effects of dietary betaine on l-[2h3-methyl-1–13c]methionine. Am. J. Clin. Nutr. 1991, 54, 386–394. [Google Scholar] [PubMed]
- McGregor, D.O.; Dellow, W.J.; Robson, R.A.; Lever, M.; George, P.M.; Chambers, S.T. Betaine supplementation decreases post-methionine hyperhomocysteinemia in chronic renal failure. Kidney Int. 2002, 61, 1040–1046. [Google Scholar] [CrossRef] [PubMed]
- Schwab, U.; Torronen, A.; Toppinen, L.; Alfthan, G.; Saarinen, M.; Aro, A.; Uusitupa, M. Betaine supplementation decreases plasma homocysteine concentrations but does not affect body weight, body composition, or resting energy expenditure in human subjects. Am. J. Clin. Nutr. 2002, 76, 961–967. [Google Scholar] [PubMed]
- Yang, H.S.; Lee, J.I.; Joo, S.T.; Park, G.B. Effects of dietary glycine betaine on growth and pork quality of finishing pigs. Asian Australas. J. Anim. Sci. 2009, 22, 706–711. [Google Scholar] [CrossRef]
- Ross, A.B.; Bruce, S.J.; Blondel-Lubrano, A.; Oguey-Araymon, S.; Beaumont, M.; Bourgeois, A.; Nielsen-Moennoz, C.; Vigo, M.; Fay, L.B.; Kochhar, S.; et al. A whole-grain cereal-rich diet increases plasma betaine, and tends to decrease total and ldl-cholesterol compared with a refined-grain diet in healthy subjects. Br. J. Nutr. 2011, 105, 1492–1502. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Sun, Q.; Li, X.; Wang, M.; Cai, D.; Zhao, R. In ovo injection of betaine affects hepatic cholesterol metabolism through epigenetic gene regulation in newly hatched chicks. PLoS ONE 2015, 10, e0122643. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Guo, G.L. Farnesoid X receptor, the bile acid sensing nuclear receptor, in liver regeneration. Acta Pharm. Sin. B 2015, 5, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Pandak, W.M.; Hylemon, P.B. LXR alpha is the dominant regulator of CYP7A1 transcription. Biochem. Biophys. Res. Commun. 2002, 293, 338–343. [Google Scholar] [CrossRef]
- Deminice, R.; da Silva, R.P.; Lamarre, S.G.; Kelly, K.B.; Jacobs, R.L.; Brosnan, M.E.; Brosnan, J.T. Betaine supplementation prevents fatty liver induced by a high-fat diet: Effects on one-carbon metabolism. Amino Acids 2015, 47, 839–846. [Google Scholar] [CrossRef] [PubMed]
- Meier, J.L. Metabolic mechanisms of epigenetic regulation. ACS Chem. Biol. 2013, 8, 2607–2621. [Google Scholar] [CrossRef] [PubMed]
- Medici, V.; Shibata, N.M.; Kharbanda, K.K.; LaSalle, J.M.; Woods, R.; Liu, S.; Engelberg, J.A.; Devaraj, S.; Torok, N.J.; Jiang, J.X.; et al. Wilson’s disease: Changes in methionine metabolism and inflammation affect global DNA methylation in early liver disease. Hepatology 2013, 57, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Wagner, T.; Jung, M. New lysine methyltransferase drug targets in cancer. Nat. Biotechnol. 2012, 30, 622–623. [Google Scholar] [CrossRef] [PubMed]
- Cai, D.; Jia, Y.; Song, H.; Sui, S.; Lu, J.; Jiang, Z.; Zhao, R. Betaine supplementation in maternal diet modulates the epigenetic regulation of hepatic gluconeogenic genes in neonatal piglets. PLoS ONE 2014, 9, e105504. [Google Scholar] [CrossRef] [PubMed]
- Obeid, R. The metabolic burden of methyl donor deficiency with focus on the betaine homocysteine methyltransferase pathway. Nutrients 2013, 5, 3481–3495. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, R.L.; Stead, L.M.; Devlin, C.; Tabas, I.; Brosnan, M.E.; Brosnan, J.T.; Vance, D.E. Physiological regulation of phospholipid methylation alters plasma homocysteine in mice. J. Biol. Chem. 2005, 280, 28299–28305. [Google Scholar] [CrossRef] [PubMed]
- Park, E.I.; Garrow, T.A. Interaction between dietary methionine and methyl donor intake on rat liver betaine-homocysteine methyltransferase gene expression and organization of the human gene. J. Biol. Chem. 1999, 274, 7816–7824. [Google Scholar] [CrossRef] [PubMed]
- Purohit, V.; Abdelmalek, M.F.; Barve, S.; Benevenga, N.J.; Halsted, C.H.; Kaplowitz, N.; Kharbanda, K.K.; Liu, Q.Y.; Lu, S.C.; McClain, C.J.; et al. Role of S-adenosylmethionine, folate, and betaine in the treatment of alcoholic liver disease: Summary of a symposium. Am. J. Clin. Nutr. 2007, 86, 14–24. [Google Scholar] [PubMed]
- Finkelstein, J.D.; Harris, B.J.; Kyle, W.E. Methionine metabolism in mammals: Kinetic study of betaine-homocysteine methyltransferase. Arch. Biochem. Biophys. 1972, 153, 320–324. [Google Scholar] [CrossRef]
- Laker, R.C.; Lillard, T.S.; Okutsu, M.; Zhang, M.; Hoehn, K.L.; Connelly, J.J.; Yan, Z. Exercise prevents maternal high-fat diet-induced hypermethylation of the Pgc-1alpha gene and age-dependent metabolic dysfunction in the offspring. Diabetes 2014, 63, 1605–1611. [Google Scholar] [CrossRef] [PubMed]
- Gugusheff, J.R.; Bae, S.E.; Rao, A.; Clarke, I.J.; Poston, L.; Taylor, P.D.; Coen, C.W.; Muhlhausler, B.S. Sex and age-dependent effects of a maternal junk food diet on the mu-opioid receptor in rat offspring. Behav. Brain Res. 2016, 301, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Kwekel, J.C.; Desai, V.G.; Moland, C.L.; Branham, W.S.; Fuscoe, J.C. Age and sex dependent changes in liver gene expression during the life cycle of the rat. BMC Genom. 2010, 11. [Google Scholar] [CrossRef] [PubMed]
- Cai, D.; Jia, Y.; Lu, J.; Yuan, M.; Sui, S.; Song, H.; Zhao, R. Maternal dietary betaine supplementation modifies hepatic expression of cholesterol metabolic genes via epigenetic mechanisms in newborn piglets. Br. J. Nutr. 2014, 112, 1459–1468. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Gestation | Lactation | |||
|---|---|---|---|---|
| Con | Bet | Con | Bet | |
| Ingredient, g/kg | ||||
| Corn | 370 | 370 | 332.5 | 332.5 |
| Wheat | 300 | 300 | 100 | 100 |
| Bran | 80 | 80 | 50 | 50 |
| Soybean meal | 170 | 170 | 253 | 253 |
| Maize starch | 0 | 0 | 150 | 150 |
| Lignocelluloses | 30 | 30 | 0 | 0 |
| CaHPO4 | 20 | 20 | 20 | 20 |
| Soybean oil | 8 | 8 | 34.5 | 34.5 |
| Fish meal | 0 | 0 | 40 | 40 |
| Premix * | 20 | 20 | 20 | 20 |
| Betaine | 0 | 3 | 0 | 3 |
| Digestible energy, MJ/kg | 13.1 | 13.1 | 14.39 | 14.39 |
| Calculated composition % | ||||
| Crude protein, % | 15 | 15 | 18 | 18 |
| Crude fiber, % | 4.5 | 4.5 | 2.3 | 2.3 |
| Calcium, % | 0.84 | 0.84 | 0.9 | 0.9 |
| Phosphorous, % | 0.65 | 0.65 | 0.7 | 0.7 |
| Target Genes | Sequences (5′ to 3′) | GenBank No. | |
|---|---|---|---|
| mRNA expression | |||
| AHCY | F: gtggtggtgtgtggctacgg | R: gcagagcacagatggggtca | NM_001201381.1 |
| APOE | F: gtgcgcaaccgcttggtgctct | R: gacgagccgcttgcgcacgtt | NM_214308.1 |
| CYP7a1 | F: tagcaggcttcccgattc | R: ctgaccagttccgagatgtg | AK230868.1 |
| CYP27a1 | F: tgtggctcgcatcgttc | R: tcacctggcagctcctt | EF625352.1 |
| GNMT | F: acaaagatgtgccca | R: gtgctgaggatgtggtcgta | NM_001110419.2 |
| MAT | F:ctgacagtcctgtcttgggagc | R: gccagagtgattctttgatgcc | NC_010458.3 |
| BHMT | F:gaggctgtgtgggcagttgaag | R: acaatggatgctcctgcctttacc | NM_001200042.1 |
| DNMT1 | F: tcagggaccacactgtaag | R: gctgcagccattcttcttgt | DQ060156.1 |
| DNMT3a | F: ggctcttctttgagttctaccg | R: gcgagatgtccctcttgtca | DQ785811.1 |
| DNMT3b | F: tgaagagtccatcgctgttg | R: caatcaccaggtcaaaggg | NM_001162404.1 |
| FXR | F: cggagaagcattacca | R: aagcattcagccaaca | XM 001928800.2 |
| HMGCR | F: caggctgaagtaagggaga | R: cacgaagtaggtggcga | DQ432054.1 |
| LDLR | F: actgctcatcctcc tctt | R: ttccgtggtcttctggta | AF065990.1 |
| LXR | F: atttccaggagtgccgtctt | R: cttgccgcttcagtttctt | AB254406.1 |
| LCAT | F: ggctggtggaagaaatgc | R: gggttggcgtagtaagaaata | NM_001164856.1 |
| PPARα | F:actgaacatcgaatgtagaatct | R: tctgaatcttacagctccgatc | NM_001044526.1 |
| PPIA | F: gactgagtggttggatgg | R: tgatcttcttgctggtctt | NC_010460.3 |
| SR-BI | F:tcaagcagcaggtcctcaag | R: cttgtgcctgaactccctgta | NM_213967.1 |
| ChIP assay | |||
| LDLR fragment | F: tcagaggagaggaagtggct | R: atccagcgctcagatgaat | |
| SR-BI fragment | F: gttgcatgaatgagcctact | R: cgtgaattccataggtaaca | |
| CYP7a1 fragment | F: tgtctccacgggcgtaccaga | R: gtggcaatatacagacatct | |
| Antibodies | MW | Species | Source | Catalogue No. |
|---|---|---|---|---|
| Western Blotting | ||||
| GNMT | 33 kd | Rabbit | proteintechTM | 18790-1-AP |
| BHMT | 50 kd | Rabbit | proteintechTM | 15965-1-AP |
| MAT | 38 kd | Rabbit | proteintechTM | 15952-1-AP |
| AHCY | 60 kd | Rabbit | proteintechTM | 10658-3-AP |
| DNMT1 | 184 kd | Rabbit | Santa Cruz | sc-20701 |
| DNMT3a | 102 kd | Rabbit | Bioworld | BS6587 |
| DNMT3b | 96 kd | Rabbit | Bioworld | BS2572 |
| HMGCR | 97 kd | Rabbit | Bioworld | BS6625 |
| LDLR | 160 kd | Rabbit | proteintechTM | 10785-1-AP |
| SR-BI | 82 kd | Rabbit | Abcam | ab137829 |
| CYP7a1 | 57 kd | Rabbit | Abcam | ab79847 |
| CYP27a1 | 60 kd | Rabbit | proteintechTM | 14739-1-AP |
| AMPK | 65 kd | Mouse | santa cruz | sc-25792 |
| P-AMPKa1/2 | 65 kd | Rabbit | santa cruz | sc-33524 |
| FXR | 69 kd | Goat | santa cruz | sc-1205 |
| LXR α/β | 49 kd | Rabbit | santa cruz | sc-13068 |
| PPARα | 55 kd | Rabbit | santa cruz | sc-9000 |
| GAPDH | 36 kd | Mouse | KangChen Bio-tech | KC-5G4 |
| β-actin | 42 kd | Mouse | KangChen Bio-tech | KC-5A08 |
| H1 | 30kd | Rabbit | Abcam | ab17584 |
| Variables | Control (n = 8) | Betaine (n = 8) |
|---|---|---|
| Body weight, kg | 7.27 ± 0.31 | 7.37 ± 0.40 |
| Liver weight, g | 183.7 ± 14.9 | 178.0 ± 10.5 |
| Biochemical metabolites | ||
| Serum betaine, μmol/L | 1.49 ± 0.11 | 3.55 ± 0.31 * |
| TG, mmol/L | 1.40 ± 0.15 | 1.11 ± 0.14 |
| TCH, mmol/L | 4.43 ± 0.38 | 4.88 ± 0.34 |
| TBA, μmol/L | 58.4 ± 4.12 | 4.88 ± 0.34 |
| LDLC, mmol/L | 1.94 ± 0.19 | 2.55 ± 0.20 |
| HDLC, mmol/L | 2.00 ± 0.14 | 1.81± 0.11 |
| LDLC/ HDLC | 1.00 ± 0.09 | 1.47 ± 0.10 * |
| Amino acids | ||
| Isoleucine (μmol/L) | 139 ± 13.5 | 148 ± 15.2 |
| Leucine (μmol/L) | 279 ± 23.8 | 309 ± 18.3 |
| Lysine (μmol/L) | 235 ± 10.2 | 259 ± 41.9 |
| Methionine (μmol/L) | 46.1 ± 6.34 | 82.3 ± 7.16 * |
| Phenylalanine (μmol/L) | 90.4 ± 5.88 | 128 ± 10.9 * |
| Tyrosine (μmol/L) | 299 ± 35.0 | 296 ± 44.5 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, D.; Yuan, M.; Liu, H.; Pan, S.; Ma, W.; Hong, J.; Zhao, R. Maternal Betaine Supplementation throughout Gestation and Lactation Modifies Hepatic Cholesterol Metabolic Genes in Weaning Piglets via AMPK/LXR-Mediated Pathway and Histone Modification. Nutrients 2016, 8, 646. https://0-doi-org.brum.beds.ac.uk/10.3390/nu8100646
Cai D, Yuan M, Liu H, Pan S, Ma W, Hong J, Zhao R. Maternal Betaine Supplementation throughout Gestation and Lactation Modifies Hepatic Cholesterol Metabolic Genes in Weaning Piglets via AMPK/LXR-Mediated Pathway and Histone Modification. Nutrients. 2016; 8(10):646. https://0-doi-org.brum.beds.ac.uk/10.3390/nu8100646
Chicago/Turabian StyleCai, Demin, Mengjie Yuan, Haoyu Liu, Shifeng Pan, Wenqiang Ma, Jian Hong, and Ruqian Zhao. 2016. "Maternal Betaine Supplementation throughout Gestation and Lactation Modifies Hepatic Cholesterol Metabolic Genes in Weaning Piglets via AMPK/LXR-Mediated Pathway and Histone Modification" Nutrients 8, no. 10: 646. https://0-doi-org.brum.beds.ac.uk/10.3390/nu8100646