
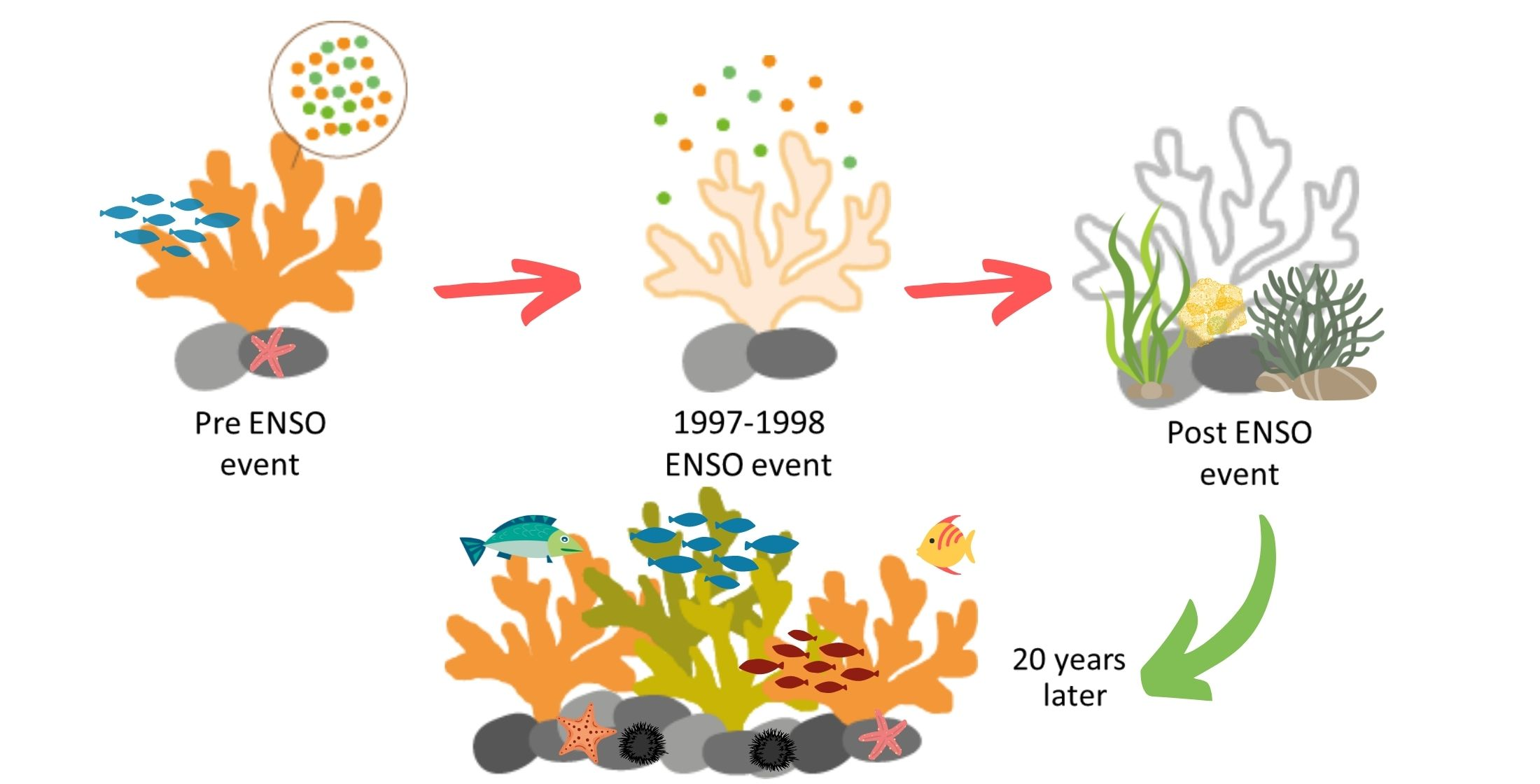
Coral Recovery in the Central Mexican Pacific 20 Years after the 1997–1998 El Niño Event

 ,
,  ,
,  ,
,  and
and
Abstract
:
1. Introduction
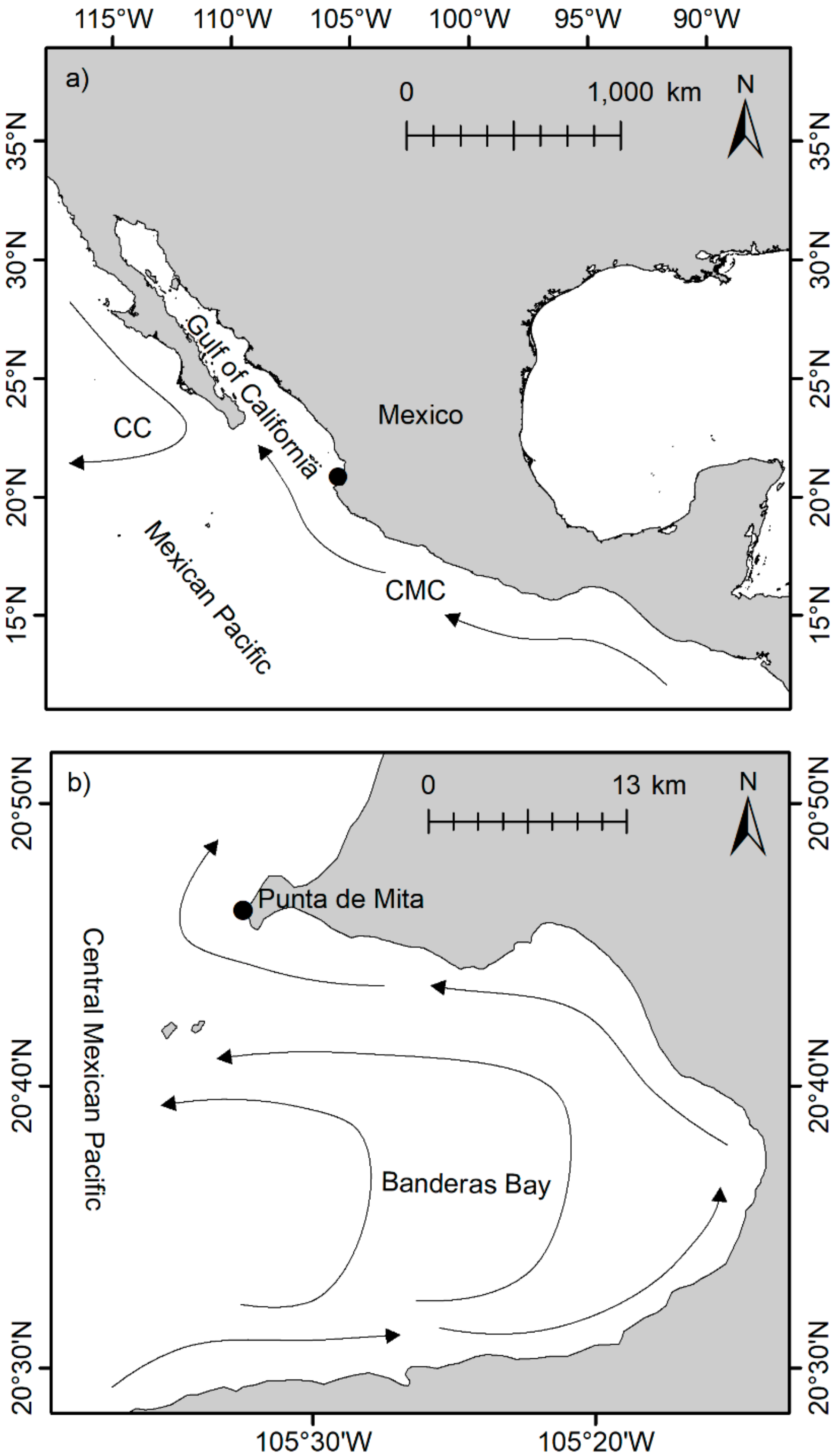
2. Materials and Methods
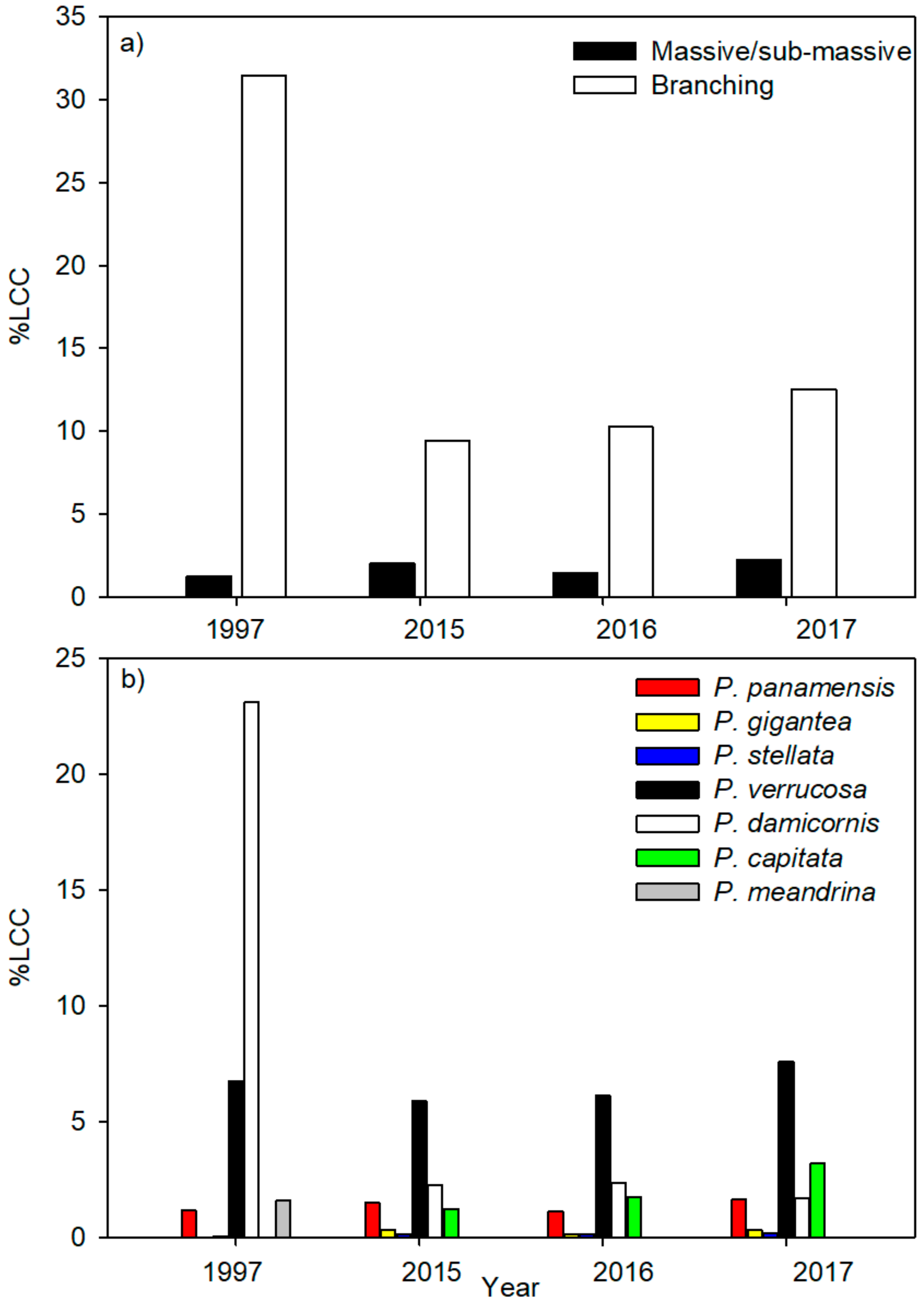
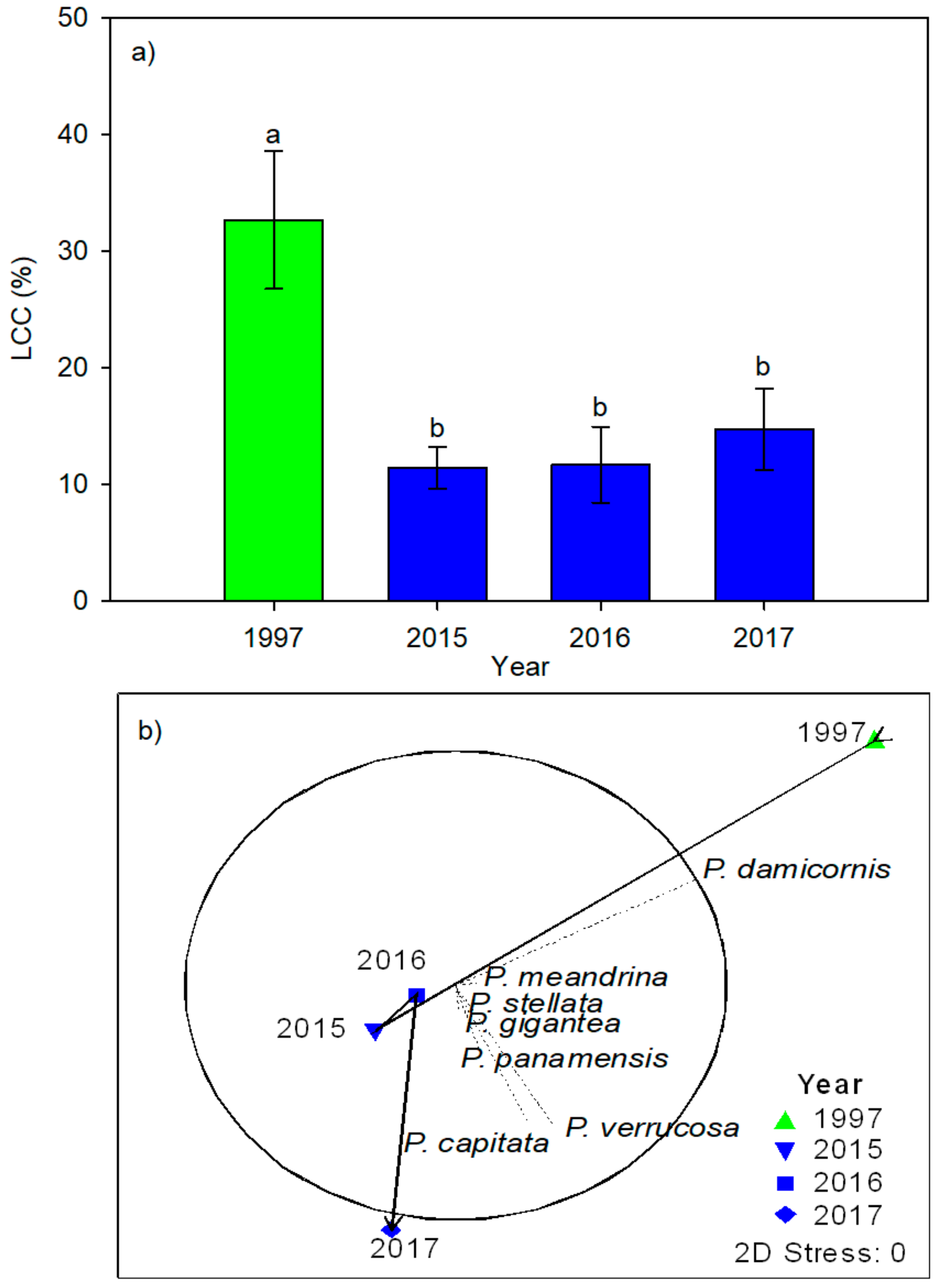
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Carpenter, K.E.; Abrar, M.; Abey, G.; Aronson, R.B.; Banks, S.; Bruckner, A.; Chiriboga, A.; Cortés, J.; Delbeek, J.C.; DeVantier, L.; et al. One third of reef-building corals face elevated extinction risk from climate change and local impacts. Science 2008, 321, 560–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burke, L.; Reytar, K.; Spalding, M.; Perry, A. Reefs at Risk Revisited; World Resources Institute: Washington, DC, USA, 2011. [Google Scholar]
- Hoegh-Guldberg, O. Coral reef sustainability through adaptation: Glimmer of hope or persistent mirage? Curr. Opin. Environ. Sustain. 2014, 7, 127–133. [Google Scholar] [CrossRef] [Green Version]
- Hughes, T.P.; Anderson, K.D.; Connolly, S.R.; Heron, S.F.; Kerry, J.T.; Lough, J.M.; Baird, A.H.; Baum, J.K.; Berumen, M.L.; Bridge, T.C.; et al. Spatial and temporal patterns of mass bleaching of corals in the Anthropocene. Science 2018, 359, 80–83. [Google Scholar] [CrossRef] [Green Version]
- Skirving, W.J.; Heron, S.F.; Marsh, B.L.; Liu, G.; De La Cour, J.L.; Geiger, E.F.; Eakin, C.M. The relentless march of mass coral bleaching: A global perspective of changing heat stress. Coral Reefs 2019, 38, 547–557. [Google Scholar] [CrossRef] [Green Version]
- Souter, D.; Planes, S.; Wicquart, J.; Logan, M.; Obura, D.; Staub, F. Status of coral reefs of the world: 2020. In Global Coral Reef Monitoring Network; International Coral Reef Initiative, Australian Institute of Marine Science: Townsville, Australia, 2020. [Google Scholar]
- Hoegh-Guldberg, O. Coral reef ecosystems and anthropogenic climate change. Reg. Environ. Chang. 2011, 11, S215–S227. [Google Scholar] [CrossRef]
- Brown, B.E. Coral bleaching: Causes and consequences. Coral Reefs 1997, 16, S129–S138. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O.; Mumby, P.J.; Hooten, A.J.; Steneck, R.S.; Greenfield, P.; Gomez, E.; Harvell, C.D.; Sale, P.F.; Edwards, A.J.; Caldeira, K.; et al. Coral reefs under rapid climate change and ocean acidification. Science 2007, 318, 1737–1742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzmán, H.M.; Cortés, J. Reef recovery 20 years after the 1982–1983 El Niño massive mortality. Mar. Biol. 2007, 151, 401–411. [Google Scholar] [CrossRef]
- Glynn, P.W.; Riegl, B.; Purkis, S.; Kerr, J.M.; Smith, T.B. Coral reef recovery in the Galapagos Islands: The northernmost islands (Darwin and Wenman). Coral Reefs 2015, 34, 421–436. [Google Scholar] [CrossRef]
- Alvarado, J.J.; Beita-Jiménez, A.; Mena, S.; Fernández-García, C.; Cortés, J.; Sánchez-Noguera, C.; Jiménez, C.; Guzmán-Mora, A.G. Cuando la conservación no puede seguir el ritmo del desarrollo: Estado de salud de los ecosistemas coralinos del Pacífico Norte de Costa Rica. Rev. Biol. Trop. 2018, 66, S280–S308. [Google Scholar] [CrossRef] [Green Version]
- Baker, A.C.; Glynn, P.W.; Riegl, B. Climate change and coral reef bleaching: An ecological assessment of long-term impacts, recovery trends and future outlook. Estuar. Coast. Shelf Sci. 2008, 80, 435–471. [Google Scholar] [CrossRef]
- Rodríguez-Troncoso, A.P.; Carpizo-Ituarte, E.; Cupul-Magaña, A.L. Physiological response to high temperature in the Eastern Tropical Pacific coral Pocillopora verrucosa. Mar. Ecol. 2016, 37, 1168–1175. [Google Scholar] [CrossRef]
- Cruz-García, R.; Rodríguez-Troncoso, A.P.; Rodríguez-Zaragoza, F.A.; Mayfield, A.; Cupul-Magaña, A.L. Ephemeral effects of El Niño southern oscillation events on an eastern tropical Pacific coral community. Mar. Freshw. Res. 2020, 71, 1259–1268. [Google Scholar] [CrossRef]
- Graham, N.A.J.; Nash, K.L.; Kool, J.T. Coral reef recovery dynamics in a changing world. Coral Reefs 2011, 30, 283–294. [Google Scholar] [CrossRef]
- Romero-Torres, M.; Acosta, A.; Palacio-Castro, A.M.; Treml, E.A.; Capata, F.A.; Paz-García, D.A.; Porter, J.W. Coral reef resilience to thermal stress in the Eastern Tropical Pacific. Glob. Chang. Biol. 2020, 26, 3880–3890. [Google Scholar] [CrossRef] [PubMed]
- Carriquiry, J.D.; Cupul-Magaña, A.L.; Rodríguez-Zaragoza, F.; Medina-Rosas, P. Coral bleaching and mortality in the Mexican Pacific during the 1997–1998 El Niño and prediction from a remote sensing approach. B Mar. Sci. 2001, 69, 237–249. [Google Scholar]
- Tortolero-Langarica, J.J.A.; Rodríguez-Troncoso, A.P.; Cupul-Magaña, A.L.; Carricart-Ganivet, J.P. Calcification and growth rate recovery of the reef-building Pocillopora species in the northeast tropical Pacific following an ENSO disturbance. PeerJ. 2017, 5, e3191. [Google Scholar] [CrossRef] [Green Version]
- Reyes-Bonilla, H.; Carriquiry, J.D.; Leyte-Morales, G.E.; Cupul-Magaña, A.L. Effects of the El Niño-Southern Oscillation and the anti-El Niño event (1997–1999) on coral reefs of the western coast of Mexico. Coral Reefs 2002, 21, 368–372. [Google Scholar] [CrossRef]
- Pantoja, D.A.; Marinone, S.G.; Parés-Sierra, A.; Gómez-Valdivia, F. Numerical modeling of seasonal and mesoscale hydrography and circulation in the Mexican Central Pacific. Cienc. Mar. 2012, 38, 363–379. [Google Scholar] [CrossRef]
- Portela, E.; Beier, E.; Barton, E.D.; Castro, R.; Godínez, V.; Palacios-Hernández, E.; Fiedler, P.C.; Sánchez-Velasco, L.; Trasviña, A. Water masses and circulation in the Tropical Pacific off Central Mexico and surrounding areas. J. Phys. Oceanogr. 2016, 46, 3069–3081. [Google Scholar] [CrossRef] [Green Version]
- Plata, L.; Filonov, A.; Tereshchenko, I.; Nelly, L.; Monzón, C.; Avalos, D.; Vargas, C. Geostrophic currents in the presence of an internal waves field in Bahia de Banderas, Mexico. E-Gnosis 2006, 4, 18. [Google Scholar]
- Santiago-Valentín, J.D.; Colley, S.B.; Glynn, P.W.; Cupul-Magaña, A.L.; López-Pérez, R.A.; Rodríguez-Zaragoza, F.A.; Benítez-Villalobos, F.; Bautista-Guerrero, E.; Zavala-Casas, D.A.; Rodríguez-Troncoso, A.P. Regional and species sexual reproductive patterns of three zooxanthellate scleractinian corals across the Eastern Tropical Pacific. Mar. Ecol. 2018, e12497. [Google Scholar] [CrossRef]
- Plata, L.; Filonov, A. Marea interna en la parte noroeste de la Bahía de Banderas, México. Cienc. Mar. 2007, 33, 197–215. [Google Scholar]
- Veron, J.E.N.; Stafford-Smith, M.G.; Turak, E.; DeVantier, L.M. Corals of the world. 2016 V. 0.01. Available online: http://www.coralsoftheworld.org/page/home/?version=0.01 (accessed on 9 March 2021).
- Clarke, K.R.; Gorley, R.N. Primer6: User Manual/Tutorial; Primer-E: Plymouth, UK, 2006. [Google Scholar]
- Zar, J.H. Biostatistical Analysis; Prentice Hall: Hoboken, NJ, USA, 2010. [Google Scholar]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; Primer-E Press: Plymouth, UK, 2008. [Google Scholar]
- Systat Software, Inc. SigmaPlot, version 11.0; Systat Software, Inc.: Chicago, IL, USA, 2008.
- Reyes-Bonilla, H. 1987 coral reef bleaching at Cabo Pulmo reef, Gulf of California, Mexico. Bull. Mar. Sci. 1993, 52, 832–837. [Google Scholar]
- Iglesias-Prieto, R.; Reyes-Bonilla, H.; Riosmena-Rodríguez, R. Effects of 1997-1998 ENSO on coral reef communities in the Gulf of California, Mexico. Geofis. Int. 2003, 42, 467–471. [Google Scholar]
- LaJeunesse, T.C.; Reyes-Bonilla, H.; Warner, M.E. Spring “bleaching” among Pocillopora in the Sea of Cortez, Eastern Pacific. Coral Reefs 2007, 26, 265–270. [Google Scholar] [CrossRef]
- López-Pérez, R.A.; Calderón-Aguilera, L.E.; Reyes-Bonilla, H.; Carriquiry, J.D.; Medina-Rosas, P.; Cupul-Magaña, A.L.; Herrero-Pérezrul, M.D.; Hernández-Ramírez, H.A.; Ahumada-Sempoal, Á.; Luna-Salguero, B.M. Coral communities from Guerrero, Southern Mexican Pacific. Mar. Ecol. 2012, 33, 407–416. [Google Scholar] [CrossRef]
- López-Pérez, R.A.; Guendulain-García, S.; Granja-Fernández, R.; Hernpandez-Urraca, V.; Galván-Rowland, L.; Zepeta-Vilchis, R.; López-López, D. Reef community changes associated with the 2009–2010 El Niño in the Southern Mexican Pacific. Pac. Sci. 2016, 70, 175–190. [Google Scholar] [CrossRef]
- Glynn, P.W. Coral reef bleaching in the 1980s and possible connections with global warming. Trends. Ecol. Evol. 1991, 6, 175–179. [Google Scholar] [CrossRef]
- Guzmán, H.M.; Cortés, J.; Richmond, R.H.; Glynn, P.W. Efectos del fenómeno de “El Niño Oscilación Sureña” 1982/1983 en los arrecifes de la Isla del Caño, Costa Rica. Rev. Biol. Trop. 1987, 35, 325–332. [Google Scholar]
- Guzmán, H.M.; Cortés, J. Coral reef community structure at Caño Island, Pacific Costa Rica. Mar. Ecol. 1989, 10, 23–41. [Google Scholar] [CrossRef]
- Guzmán, H.M.; Cortés, J. Changes in reef community structure after fifteen years of natural disturbances in the Eastern Pacific (Costa Rica). Bull. Mar. Sci. 2001, 69, 133–149. [Google Scholar]
- Guzmán, H.M.; Cortés, J. Cocos Island (Pacific of Costa Rica) coral reefs after the 1982–1983 El Niño disturbance. Rev. Biol. Trop. 1992, 40, 309–324. [Google Scholar]
- Jiménez, C.; Cortés, J.; León, A.; Ruíz, E. Coral bleaching and mortality associated with the 1997–1998 El Niño in an upwelling environment in the Eastern Pacific (Gulf of Papagayo, Costa Rica). Bull. Mar. Sci. 2001, 69, 151–169. [Google Scholar]
- Cortés, J. The coral reefs of Golfo Dulce, Costa Rica: Distribution and community structure. Atoll. Res. Bull. 1990, 344, 1–37. [Google Scholar] [CrossRef]
- Glynn, P.W.; Maté, J.L.; Baker, A.C.; Calderón, M.O. Coral bleaching and mortality in Panama and Ecuador during the 1997–1998 El Niño-Southern Oscillation event: Spatial/temporal patterns and comparisons with the 1982–1983 event. Bull. Mar. Sci. 2001, 69, 79–109. [Google Scholar]
- Glynn, P.W. Coral mortality and disturbances to coral reefs in the Tropical Eastern Pacific. In Global Ecological Consequences of the 1982~1983 El Niño-Southern Oscillation; Glynn, P.W., Ed.; Elsevier: Amsterdam, The Netherlands, 1990; pp. 55–126. [Google Scholar]
- Randall, C.J.; Toth, L.T.; Leichter, J.J.; Maté, J.L.; Aronson., R.B. Upwelling buffers climate change impacts on coral reefs of the eastern tropical Pacific. Ecology 2020, 101, e02918. [Google Scholar] [CrossRef]
- Tortolero-Langarica, J.J.A.; Carricart-Ganivet, J.P.; Cupul-Magaña, A.L.; Rodríguez-Troncoso, A.P. Historical insights on growth rates of the reef building corals Pavona gigantea and Porites panamensis from the Northeastern tropical Pacific. Mar. Environ. Res. 2017, 132, 23–32. [Google Scholar] [CrossRef]
- Santiago-Valentín, J.D.; Rodríguez-Troncoso, A.P.; Bautista-Guerrero, E.; López-Pérez, A.; Cupul-Magaña, A.L. Successful sexual reproduction of the scleractinian coral Porites panamensis: Evidence of planktonic larvae and recruitment. Invertebr. Biol. 2019, 138, 29–39. [Google Scholar] [CrossRef]
- Martínez-Castillo, V.; Rodríguez-Troncoso, A.P.; Santiago-Valentín, J.D.; Cupul-Magaña, A.L. The influence of urban pressures on coral physiology on marginal coral reefs of the Mexican Pacific. Coral Reefs 2020, 39, 625–637. [Google Scholar] [CrossRef]
- Saavedra-Sotelo, N.C.; Calderon-Aguilera, L.E.; Reyes-Bonilla, H.; López-Pérez, R.A.; Medina Rosas, P.; Rocha-Olivares, A. Limited genetic connectivity of Pavona gigantea in the Mexican Pacific. Coral Reefs 2011, 30, 677–686. [Google Scholar] [CrossRef]
- Paz-García, D.A.; Chávez-Romo, H.E.; Correa-Sandoval, F.; Reyes-Bonilla, H.; López-Pérez, R.A.; Medina-Rosas, P.; Hernández-Cortés, M.P. Genetic connectivity patterns of corals Pocillopora damicornis and Porites panamensis (Anthozoa: Scleractinia) along the west coast of Mexico. Pac. Sci. 2012, 66, 43–61. [Google Scholar] [CrossRef]
- Burt, J.; Al-Harthi, S.; Al-Cibahy, A. Long-term impacts of coral bleaching events on the world’s warmest Reefs. Mar. Environ. Res. 2011, 72, 225–229. [Google Scholar]
- van Woesik, R.; Sakai, K.; Ganase, A.; Loya, A. Revisiting the winners and the losers a decade after coral bleaching. Mar. Ecol. Prog. Ser. 2011, 434, 67–76. [Google Scholar] [CrossRef] [Green Version]
- Loya, Y.; Sakai, K.; Yamazato, K.; Nakano, Y.; Sambali, H.; van Woesik, R. Coral bleaching: The winners and the losers. Ecol. Lett. 2001, 4, 122–131. [Google Scholar] [CrossRef]
- Paz-García, D.A.; Balart, E.F.; García-de León, F.J. Cold water bleaching of Pocillopora in the Gulf of California. In Proceedings of the 12th International Coral Reef Symposium, Cairns, Australia, 9–13 July 2012. [Google Scholar]
- Al-Sofyani, A.A.; Floos, Y.A.M. Effect of temperature on two reef-building corals Pocillopora damicornis and P. verrucosa in the Red Sea. Oceanologia 2013, 55, 917–935. [Google Scholar] [CrossRef] [Green Version]
- Tsounis, G.; Edmunds, P.J. The potential for self-seeding by the coral Pocillopora spp. in Moorea, French Polynesia. PeerJ. 2016, 4, e2544. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Troncoso, A.P.; Carpizo-Ituarte, E.; Pettay, D.T.; Warner, M.E.; Cupul-Magaña, A.L. The effects of abnormal decrease in temperature on the Eastern Pacific reef-building coral Pocillopora verrucosa. Mar. Biol. 2014, 161, 131–139. [Google Scholar] [CrossRef]
- Alvarado, J.J.; Sánchez-Noguera, C.; Arias-Godínez, G.; Araya, T.; Fernández-García, C.; Guzmán, A.G. Impact of El Niño 2015–2016 on the coral reefs of the Pacific of Costa Rica: The potential role of marine protection. Rev. Biol. Trop. 2020, 68, S271–S282. [Google Scholar] [CrossRef]
- Pratchett, M.S.; McWilliam, M.J.; Riegl, B. Contrasting shifts in coral assemblages with increasing disturbances. Coral Reefs 2020, 39, 783–793. [Google Scholar] [CrossRef]
- Cabral-Tena, R.A.; López-Pérez, A.; Reyes-Bonilla, H.; Calderon-Aguilera, L.E.; Norzagaray-López, C.O.; Rodríguez-Zaragoza, F.A.; Cupul-Magaña, A.; Rodríguez-Troncoso, A.P.; Ayala-Bocos, A. Calcification of coral assemblages in the Eastern Pacific: Rrshuffling calcification scenarios under climate change. Ecol. Indic. 2018, 95, 726–734. [Google Scholar] [CrossRef]
- Sweatman, H.; Delean, S.; Syms, C. Assessing loss of coral cover on Australia’s Great Barrier Reef over two decades, with implications for longer-term trends. Coral Reefs 2011, 30, 521–531. [Google Scholar] [CrossRef]
- Norzagaray-López, C.O.; Calderon-Aguilera, L.E.; Hernández-Ayón, J.M.; Reyes-Bonilla, H.; Carricart-Ganivet, J.P.; Cabral-Tena, R.A.; Balart, E.F. Low calcification rates and calcium carbonate production in Porites panamensis at its northernmost geographic distribution. Mar. Ecol. 2015, 36, 1244–1255. [Google Scholar] [CrossRef]
- LaJeunesse, T.C.; Parkinson, J.E.; Gabrielson, P.W.; Jeong, H.J.; Reimer, J.D.; Voolstra, C.R.; Santos, S.R. Systematic revision of Symbiodiniaceae highlights the antiquity and diversity of coral endosymbionts. Curr. Biol. 2018, 28, 2570–2580. [Google Scholar] [CrossRef] [Green Version]
- LaJeunesse, T.C.; Smith, R.; Walther, M.; Pinzón, J.; Pettay, D.T.; McGinley, M.; Aschaffenburg, M.; Medina-Rosas, P.; Cupul-Magaña, A.L.; López-Pérez, A.; et al. Host-symbiont recombination versus natural selection in the response of coral-dinoflagellate symbioses to environmental disturbance. Proc. R. Soc. B 2010, 277, 2925–2934. [Google Scholar] [CrossRef] [Green Version]
- Prouty, N.G.; Cohen, A.; Yates, K.K.; Storlazzi, C.D.; Swarzenski, P.W.; White, D. Vulnerability of coral reefs to bioerosion from land-based sources of pollution. J. Geophys. Res. Oceans 2017, 122, 9319–9331. [Google Scholar] [CrossRef]
- Fabricius, K.E. Effects of terrestrial runoff on the ecology of corals and coral reefs: Review and synthesis. Mar. Pollut. Bull. 2005, 50, 125–146. [Google Scholar] [CrossRef]
- Fabricius, K.E. Factors determining the resilience of coral reefs to eutrophication: A review and conceptual model. In Coral Reefs: An Ecosysztem in Transition; Dubinsky, Z., Stambler, N., Eds.; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2011; pp. 493–505. [Google Scholar]
- Merchand-Rojas, M.A. The inter-state development of Puerto Vallarta and Bahia de Banderas: México. Prob. Des. 2012, 43, 147–173. [Google Scholar]
- Glynn, P.W.; Ault, J.S. A biogeographic analysis and review of the far eastern Pacific coral reef region. Coral Reefs 2000, 19, 1–23. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
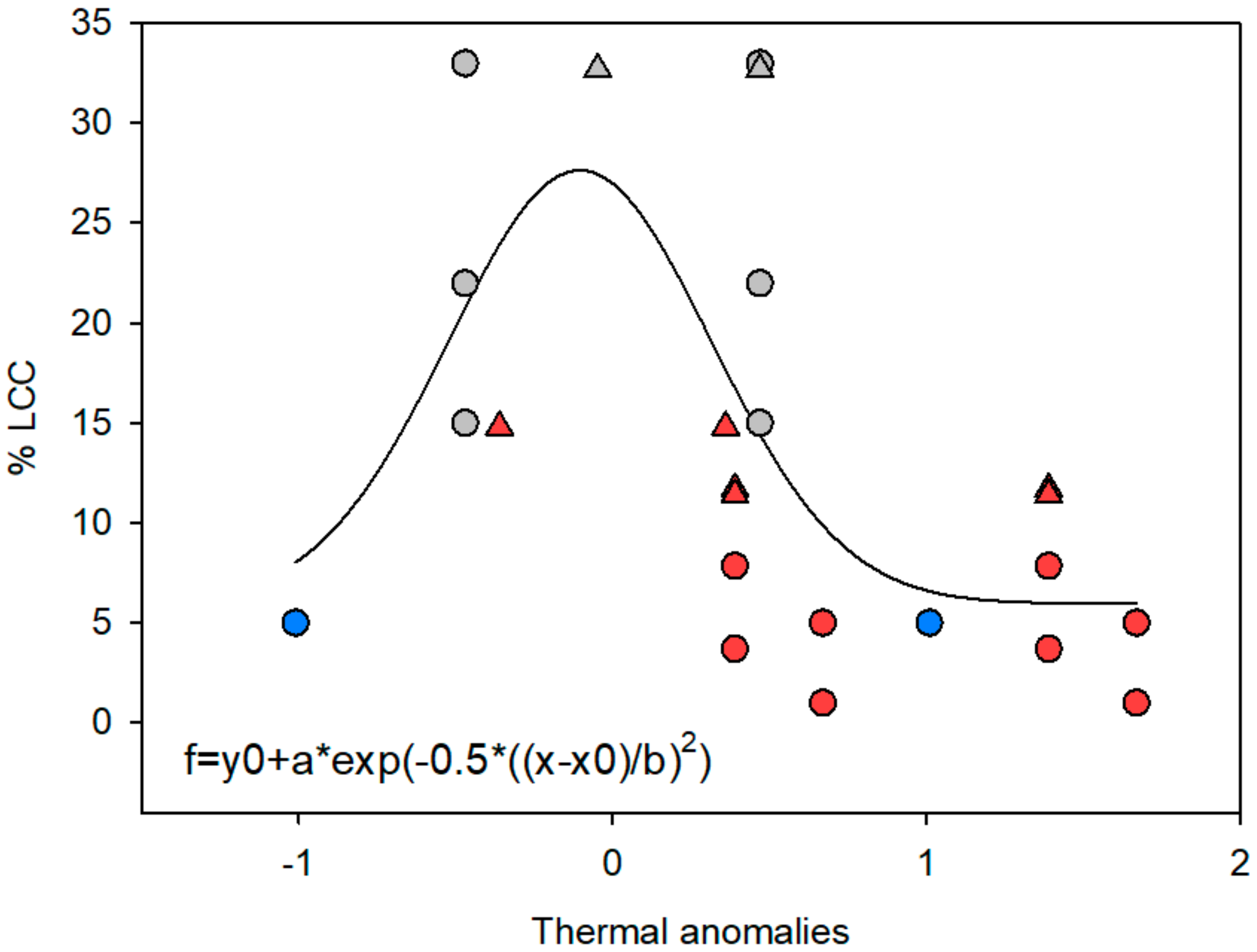{kind=link}
| ETP Site | Years Surveyed | ENSO Event * | Pre-ENSO % LCC | Post-ENSO % LCC (% Mortality) | % LCC After Recovery | Reference |
|---|---|---|---|---|---|---|
| Cabo Pulmo (GC) | 1987–1988 | Strong | ~30 | (10) | NA | [31] |
| GC | 1997–1999 | Very Strong | ~28 | 25 (18) | NA | [20] |
| GC | 1996–1998 | Very strong | NA | 59.3 (13.6–21.9) | NA | [32] |
| GC | 2006 | Weak | NA | No mortality | - | [33] |
| Careyeros (CMP) | 1997–1998 | Very strong | 22 | 1 (96) | NA | [18] |
| Punta de Mita (CMP) | 1997–1998 | Very strong | 33 | <5 (96) | NA | [18] |
| CMP | 1997–1999 | Very strong | ~15 | <5 (>60) | NA | [20] |
| Punta de Mita (CMP) | 1997–2017 | Very strong | 32.7 | NA | 14.8 | This study |
| Oaxaca (SMP) | 1997–1999 | Very strong | ~57 | <5 | NA | [20] |
| SMP | 2009 | Moderate | 18–65 | NA | NA | [34] |
| El Chato (SMP) | 2009 | Moderate | 18 | NA | NA | [35] |
| Manzanillo (SMP) | 2009 | Moderate | 59 | NA | NA | [35] |
| Zacatoso (SMP) | 2009 | Moderate | 65 | NA | NA | [35] |
| CR | 1982–1983 | Very strong | NA | (50) | NA | [36] |
| Isla del Caño (CR) | 1980–1984 | Very strong | 17.8 | 8.55 (46.16) | NA | [37] |
| Isla del Caño (CR) | 1985 | Weak | 5.4 | NA | 6.94–18.76 | [38] |
| Isla del Caño (CR) | 1984–1999 | Several events | ~32 | ~5 (5–52) | ~ 10 | [39] |
| Cocos Island (CR) | 1978 and 1987 | Very strong | NA | (90) | 2.99 | [40] |
| Cocos Islands (CR) | 1987, 2002 | Several events | 2.99 | NA | 14.87 | [9] |
| Gulf of Papagayo (CR) | 1996–1998 | Very strong | NA | (5.7) | NA | [41] |
| Punta El Bajo (CR) | 1985 | Weak | 45.9 | NA | NA | [42] |
| PN | 1982–1983 | Very strong | NA | (75–85) | NA | [36] |
| PN | 1997–1999 | Very strong | NA | (13.1) | NA | [43] |
| Gulf of Chiriqui (PN) | Pre-1983 and 1984 | Very strong | 60–90 | 3.3–63.4 | NA | [44] |
| Gulf of Chiriquí (PN) | 2016–2018 | Very strong | 50 | 40.8 (9.2) | NA | [45] |
| Gulf of Panama (PN) | 2016–2018 | Very strong | 78 | 71.2 (6.8) | NA | [45] |
| GI | 1982–1983 | Very strong | NA | (97–100) | NA | [36,44] |
| GI | 1997–1999 | Very strong | NA | (26.2) | NA | [43] |
| GI | 2007, 2012 | Several events | 19.4 | NA | 32.3 | [11] |
| Shark Bay, Wenman (GI) | 1975, 2012 | Several events | 14.4 | NA | 36.94 | [11] |
| EC | 1975, 1986, 1998 | Several events | NA | (7) | NA | [40] |
| Overall ETP | 1982–1997 | Several events | 32.7 | 7.1 | 33.9 | [17] |
| Index | Pseudo-F | P (Perm) |
|---|---|---|
| Number of species | 1.0187 | 0.4242 |
| Diversity (H’) | 1.9903 | 0.1438 |
| Evenness (J’) | 2.761 | 0.0666 |
| Dominance (λ) | 2.1264 | 0.1238 |
| Live coral cover (LCC) | 6.5954 | 0.0026 * |
| Test | t | P (Perm) |
|---|---|---|
| 1997 vs. 2015 | 4.7504 | 0.004 * |
| 1997 vs. 2016 | 3.4194 | 0.0121 * |
| 1997 vs. 2017 | 2.8015 | 0.037 * |
| 2015 vs. 2016 | 0.077855 | 0.9424 |
| 2015 vs. 2017 | 0.92631 | 0.3741 |
| 2016 vs. 2017 | 0.64205 | 0.5309 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-Castillo, V.; Rodríguez-Troncoso, A.P.; Mayfield, A.B.; Rodríguez-Zaragoza, F.A.; Cupul-Magaña, A.L. Coral Recovery in the Central Mexican Pacific 20 Years after the 1997–1998 El Niño Event. Oceans 2022, 3, 48-59. https://0-doi-org.brum.beds.ac.uk/10.3390/oceans3010005
Martínez-Castillo V, Rodríguez-Troncoso AP, Mayfield AB, Rodríguez-Zaragoza FA, Cupul-Magaña AL. Coral Recovery in the Central Mexican Pacific 20 Years after the 1997–1998 El Niño Event. Oceans. 2022; 3(1):48-59. https://0-doi-org.brum.beds.ac.uk/10.3390/oceans3010005
Chicago/Turabian StyleMartínez-Castillo, Violeta, Alma Paola Rodríguez-Troncoso, Anderson B. Mayfield, Fabián Alejandro Rodríguez-Zaragoza, and Amílcar Leví Cupul-Magaña. 2022. "Coral Recovery in the Central Mexican Pacific 20 Years after the 1997–1998 El Niño Event" Oceans 3, no. 1: 48-59. https://0-doi-org.brum.beds.ac.uk/10.3390/oceans3010005