
Current Applications of Digital PCR in Veterinary Parasitology: An Overview
 , ,
, ,  ,
,  and
and 
Abstract
:1. Introduction
2. Protozoa
2.1. Eimeria spp.
2.2. Haemosporidian Parasites
2.3. Cytauxzoon felis
2.4. Cryptosporidium spp.
2.5. Leishmania infantum
2.6. Theileria spp.
2.7. Toxoplasma gondii
3. Trematodes
Schistosoma japonicum
4. Cestodes
Echinococcus multilocularis
5. Nematodes
5.1. Gastrointestinal Nematodes
5.1.1. Haemonchus contortus
5.1.2. Trichuris spp.
5.1.3. Mixed Infections with Gastrointestinal Nematodes in Ruminants
5.1.4. Dual Infections with Ascaridia galli and Heterakis gallinarum in Poultry
5.2. Dirofilaria immitis
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Kumar, S.; Gupta, S.; Mohmad, A.; Fular, A.; Parthasarathi, B.C.; Chaubey, A.K. Molecular Tools-Advances, Opportunities and Prospects for the Control of Parasites of Veterinary Importance. Int. J. Trop. Insect Sci. 2021, 41, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Baltrušis, P.; Höglund, J. Digital PCR: Modern Solution to Parasite Diagnostics and Population Trait Genetics. Parasites Vectors 2023, 16, 143. [Google Scholar] [CrossRef]
- Elmahalawy, S.T.; Halvarsson, P.; Skarin, M.; Höglund, J. Droplet Digital Polymerase Chain Reaction (DdPCR) as a Novel Method for Absolute Quantification of Major Gastrointestinal Nematodes in Sheep. Vet. Parasitol. 2018, 261, 1–8. [Google Scholar] [CrossRef]
- Snyder, R.P.; Guerin, M.T.; Hargis, B.M.; Imai, R.; Kruth, P.S.; Page, G.; Rejman, E.; Barta, J.R. Exploiting Digital Droplet PCR and Next Generation Sequencing Technologies to Determine the Relative Abundance of Individual Eimeria Species in a DNA Sample. Vet. Parasitol. 2021, 296, 109443. [Google Scholar] [CrossRef] [PubMed]
- Tarbiat, B.; Enweji, N.; Baltrusis, P.; Halvarsson, P.; Osterman-Lind, E.; Jansson, D.S.; Höglund, J. A Novel Duplex DdPCR Assay for Detection and Differential Diagnosis of Ascaridia galli and Heterakis gallinarum Eggs from Chickens Feces. Vet. Parasitol. 2021, 296, 109499. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Zhao, Z.; Chen, L.; Li, J.; Ju, X. Development of a Droplet Digital PCR for Detection of Trichuriasis in Sheep. J. Parasitol. 2020, 106, 603–610. [Google Scholar] [CrossRef]
- Baltrušis, P.; Halvarsson, P.; Höglund, J. Exploring Benzimidazole Resistance in Haemonchus contortus by next Generation Sequencing and Droplet Digital PCR. Int. J. Parasitol. Drugs Drug Resist. 2018, 8, 411–419. [Google Scholar] [CrossRef]
- Shang Kuan, T.-C.; Prichard, R.K. Developmental Regulation of Dirofilaria immitis Microfilariae and Evaluation of Ecdysone Signaling Pathway Transcript Level Using Droplet Digital PCR. Parasites Vectors 2020, 13, 614. [Google Scholar] [CrossRef]
- Van Dorssen, C.F.; Gordon, C.A.; Li, Y.; Williams, G.M.; Wang, Y.; Luo, Z.; Gobert, G.N.; You, H.; McManus, D.P.; Gray, D.J. Rodents, Goats and Dogs—Their Potential Roles in the Transmission of Schistosomiasis in China. Parasitology 2017, 144, 1633–1642. [Google Scholar] [CrossRef]
- Chen, B.; Jiang, Y.; Cao, X.; Liu, C.; Zhang, N.; Shi, D. Droplet Digital PCR as an Emerging Tool in Detecting Pathogens Nucleic Acids in Infectious Diseases. Clin. Chim. Acta 2021, 517, 156–161. [Google Scholar] [CrossRef]
- Wichianchot, S.; Hongsrichan, N.; Maneeruttanarungroj, C.; Pinlaor, S.; Iamrod, K.; Purisarn, A.; Donthaisong, P.; Karanis, P.; Nimsuphan, B.; Rucksaken, R. A Newly Developed Droplet Digital PCR for Ehrlichia Canis Detection: Comparisons to Conventional PCR and Blood Smear Techniques. J. Vet. Med. Sci. 2022, 84, 831–840. [Google Scholar] [CrossRef] [PubMed]
- Massolo, A.; Gerber, A.; Umhang, G.; Nicholas, C.; Liccioli, S.; Mori, K.; Klein, C. Droplet Digital PCR as a Sensitive Tool to Assess Exposure Pressure from Echinococcus multilocularis in Intermediate Hosts. Acta Trop. 2021, 223, 106078. [Google Scholar] [CrossRef] [PubMed]
- Powell, E.A.; Babady, N.E. Digital PCR in the Clinical Microbiology Laboratory: Another Tool on the Molecular Horizon. Clin. Microbiol. Newsl. 2018, 40, 27–32. [Google Scholar] [CrossRef]
- Salipante, S.J.; Jerome, K.R. Digital PCR—An Emerging Technology with Broad Applications in Microbiology. Clin. Chem. 2020, 66, 117–123. [Google Scholar] [CrossRef]
- Fatoba, A.J.; Adeleke, M.A. Diagnosis and Control of Chicken Coccidiosis: A Recent Update. J. Parasit. Dis. 2018, 42, 483–493. [Google Scholar] [CrossRef]
- Long, P.L.; Joyner, L.P. Problems in the Identification of Species of Eimeria. J. Protozool. 1984, 31, 535–541. [Google Scholar] [CrossRef]
- Kawahara, F.; Taira, K.; Nagai, S.; Onaga, H.; Onuma, M.; Nunoya, T. Detection of Five Avian Eimeria Species by Species-Specific Real-Time Polymerase Chain Reaction Assay. Avian Dis. 2008, 52, 652–656. [Google Scholar] [CrossRef]
- Kirkpatrick, N.C.; Blacker, H.P.; Woods, W.G.; Gasser, R.B.; Noormohammadi, A.H. A Polymerase Chain Reaction-Coupled High-Resolution Melting Curve Analytical Approach for the Monitoring of Monospecificity of Avian Eimeria Species. Avian Pathol. 2009, 38, 13–19. [Google Scholar] [CrossRef]
- Lalonde, L.F.; Gajadhar, A.A. Detection and Differentiation of Coccidian Oocysts by Real-Time PCR and Melting Curve Analysis. J. Parasitol. 2011, 97, 725–730. [Google Scholar] [CrossRef]
- Perkins, S.L. Malaria’s Many Mates: Past, Present, and Future of the Systematics of the Order Haemosporida. J. Parasitol. 2014, 100, 11–25. [Google Scholar] [CrossRef]
- Valkiūnas, G.; Iezhova, T.A. Keys to the Avian Haemoproteus Parasites (Haemosporida, Haemoproteidae). Malar. J. 2022, 21, 269. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, C.T.; Thomas, N.J.; Hunter, D.B. (Eds.) Parasitic Diseases of Wild Birds; Wiley-Blackwell: Ames, IA, USA, 2008; ISBN 978-0-8138-2081-1. [Google Scholar]
- Bernotienė, R.; Palinauskas, V.; Iezhova, T.; Murauskaitė, D.; Valkiūnas, G. Avian Haemosporidian Parasites (Haemosporida): A Comparative Analysis of Different Polymerase Chain Reaction Assays in Detection of Mixed Infections. Exp. Parasitol. 2016, 163, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Valkiŭnas, G.; Bensch, S.; Iezhova, T.A.; Krizanauskiené, A.; Hellgren, O.; Bolshakov, C.V. Nested Cytochrome b Polymerase Chain Reaction Diagnostics Underestimate Mixed Infections of Avian Blood Haemosporidian Parasites: Microscopy Is Still Essential. J. Parasitol. 2006, 92, 418–422. [Google Scholar] [CrossRef] [PubMed]
- Bensch, S.; Hellgren, O. The Use of Molecular Methods in Studies of Avian Haemosporidians. In Avian Malaria and Related Parasites in the Tropics; Santiago-Alarcon, D., Marzal, A., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 113–135. ISBN 978-3-030-51632-1. [Google Scholar]
- Huang, X. Assessment of Associations between Malaria Parasites and Avian Hosts-A Combination of Classic System and Modern Molecular Approach. Biology 2021, 10, 636. [Google Scholar] [CrossRef]
- Jiang, Y.; Brice, B.; Nguyen, M.; Loh, R.; Greay, T.; Adlard, R.; Ryan, U.; Yang, R. Further Characterisation of Leucocytozoon Podargii in Wild Tawny Frogmouths (Podargus strigoides) in Western Australia. Parasitol. Res. 2019, 118, 1833–1840. [Google Scholar] [CrossRef]
- Kao, Y.-F.; Peake, B.; Madden, R.; Cowan, S.R.; Scimeca, R.C.; Thomas, J.E.; Reichard, M.V.; Ramachandran, A.; Miller, C.A. A Probe-Based Droplet Digital Polymerase Chain Reaction Assay for Early Detection of Feline Acute Cytauxzoonosis. Vet. Parasitol. 2021, 292, 109413. [Google Scholar] [CrossRef]
- Lloret, A.; Addie, D.D.; Boucraut-Baralon, C.; Egberink, H.; Frymus, T.; Gruffydd-Jones, T.; Hartmann, K.; Horzinek, M.C.; Hosie, M.J.; Lutz, H.; et al. Cytauxzoonosis in Cats: ABCD Guidelines on Prevention and Management. J. Feline Med. Surg. 2015, 17, 637–641. [Google Scholar] [CrossRef]
- O’Leary, J.K.; Sleator, R.D.; Lucey, B. Cryptosporidium spp. Diagnosis and Research in the 21st Century. Food Waterborne Parasitol. 2021, 24, e00131. [Google Scholar] [CrossRef]
- Fayer, R. Cryptosporidium: A Water-Borne Zoonotic Parasite. Vet. Parasitol. 2004, 126, 37–56. [Google Scholar] [CrossRef]
- Khurana, S.; Chaudhary, P. Laboratory Diagnosis of Cryptosporidiosis. Trop. Parasitol. 2018, 8, 2. [Google Scholar] [CrossRef]
- Yang, R.; Paparini, A.; Monis, P.; Ryan, U. Comparison of Next-Generation Droplet Digital PCR (DdPCR) with Quantitative PCR (QPCR) for Enumeration of Cryptosporidium Oocysts in Faecal Samples. Int. J. Parasitol. 2014, 44, 1105–1113. [Google Scholar] [CrossRef] [PubMed]
- Zahedi, A.; Ryan, U.; Rawlings, V.; Greay, T.; Hancock, S.; Bruce, M.; Jacobson, C. Cryptosporidium and Giardia in Dam Water on Sheep Farms—An Important Source of Transmission? Vet. Parasitol. 2020, 288, 109281. [Google Scholar] [CrossRef]
- Mthethwa, N.P.; Amoah, I.D.; Reddy, P.; Bux, F.; Kumari, S. Development and Evaluation of a Molecular Based Protocol for Detection and Quantification of Cryptosporidium spp. in Wastewater. Exp. Parasitol. 2022, 234, 108216. [Google Scholar] [CrossRef] [PubMed]
- Hawash, Y. DNA Extraction from Protozoan Oocysts/Cysts in Feces for Diagnostic PCR. Korean J. Parasitol. 2014, 52, 263–271. [Google Scholar] [CrossRef]
- Schrader, C.; Schielke, A.; Ellerbroek, L.; Johne, R. PCR Inhibitors—Occurrence, Properties and Removal. J. Appl. Microbiol. 2012, 113, 1014–1026. [Google Scholar] [CrossRef] [PubMed]
- Zacharia, A.; Ahmada, W.; Outwater, A.H.; Ngasala, B.; Van Deun, R. Evaluation of Occurrence, Concentration, and Removal of Pathogenic Parasites and Fecal Coliforms in Three Waste Stabilization Pond Systems in Tanzania. Sci. World J. 2019, 2019, 3415617. [Google Scholar] [CrossRef]
- Bruno, B.; Romano, A.; Zanatta, R.; Spina, S.; Mignone, W.; Ingravalle, F.; Barzanti, P.; Ceccarelli, L.; Goria, M. Serum Indirect Immunofluorescence Assay and Real-Time PCR Results in Dogs Affected by Leishmania infantum: Evaluation before and after Treatment at Different Clinical Stages. J. Vet. Diagn. Investig. 2019, 31, 222–227. [Google Scholar] [CrossRef]
- Francino, O.; Altet, L.; Sánchez-Robert, E.; Rodriguez, A.; Solano-Gallego, L.; Alberola, J.; Ferrer, L.; Sánchez, A.; Roura, X. Advantages of Real-Time PCR Assay for Diagnosis and Monitoring of Canine Leishmaniosis. Vet. Parasitol. 2006, 137, 214–221. [Google Scholar] [CrossRef]
- Akhoundi, M.; Downing, T.; Votýpka, J.; Kuhls, K.; Lukeš, J.; Cannet, A.; Ravel, C.; Marty, P.; Delaunay, P.; Kasbari, M.; et al. Leishmania Infections: Molecular Targets and Diagnosis. Mol. Asp. Med. 2017, 57, 1–29. [Google Scholar] [CrossRef]
- Tsokana, C.N.; Athanasiou, L.V.; Valiakos, G.; Spyrou, V.; Manolakou, K.; Billinis, C. Molecular Diagnosis of Leishmaniasis, Species Identification and Phylogenetic Analysis. In Leishmaniasis—Trends in Epidemiology, Diagnosis and Treatment; Claborn, D., Ed.; InTech: London, UK, 2014; ISBN 978-953-51-1232-7. [Google Scholar]
- Pereira, D.C.A.; Teixeira-Neto, R.G.; Lopes, V.V.; Pena, H.P.; Paz, G.F.; Custodio, C.H.X.; Belo, V.S.; Da Fonseca Júnior, A.A.; Da Silva, E.S. Development of Quantitative PCR and Digital PCR for the Quantification of Leishmania infantum in Dogs. Mol. Cell Biochem. 2023. [Google Scholar] [CrossRef]
- De Vasconcelos, T.C.B.; Bruno, S.F.; de Miranda, L.H.M.; Conceição-Silva, F.; Belo, V.S.; Figueiredo, F.B. Parasite Load, INOS and Cytokine Profiles, and Histopathological Aspects of Leishmania infantum Infection in Dogs with Different Clinical Presentations. Cienc. Rural. 2019, 49, e20180984. [Google Scholar] [CrossRef]
- Murthy, S.; Suresh, A.; Dandasena, D.; Singh, S.; Subudhi, M.; Bhandari, V.; Bhanot, V.; Arora, J.S.; Sharma, P. Multiplex DdPCR: A Promising Diagnostic Assay for Early Detection and Drug Monitoring in Bovine Theileriosis. Pathogens 2023, 12, 296. [Google Scholar] [CrossRef]
- Mans, B.J.; Pienaar, R.; Latif, A.A. A Review of Theileria Diagnostics and Epidemiology. Int. J. Parasitol. Parasites Wildl. 2015, 4, 104–118. [Google Scholar] [CrossRef] [PubMed]
- Mhadhbi, M.; Naouach, A.; Boumiza, A.; Chaabani, M.F.; BenAbderazzak, S.; Darghouth, M.A. In Vivo Evidence for the Resistance of Theileria annulata to Buparvaquone. Vet. Parasitol. 2010, 169, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Mancusi, A.; Giordano, A.; Bosco, A.; Girardi, S.; Proroga, Y.T.R.; Morena, L.; Pinto, R.; Sarnelli, P.; Cringoli, G.; Rinaldi, L.; et al. Development of a Droplet Digital Polymerase Chain Reaction Tool for the Detection of Toxoplasma Gondii in Meat Samples. Parasitol. Res. 2022, 121, 1467–1473. [Google Scholar] [CrossRef] [PubMed]
- Opsteegh, M.; Langelaar, M.; Sprong, H.; Den Hartog, L.; De Craeye, S.; Bokken, G.; Ajzenberg, D.; Kijlstra, A.; Der Giessen, J.V. Direct Detection and Genotyping of Toxoplasma Gondii in Meat Samples Using Magnetic Capture and PCR. Int. J. Food Microbiol. 2010, 139, 193–201. [Google Scholar] [CrossRef]
- Marino, A.M.F.; Giunta, R.P.; Salvaggio, A.; Castello, A.; Alfonzetti, T.; Barbagallo, A.; Aparo, A.; Scalzo, F.; Reale, S.; Buffolano, W.; et al. Toxoplasma Gondii in Edible Fishes Captured in the Mediterranean Basin. Zoonoses Public. Health 2019, 66, 826–834. [Google Scholar] [CrossRef]
- Gray, D.J.; Li, Y.-S.; Williams, G.M.; Zhao, Z.-Y.; Harn, D.A.; Li, S.-M.; Ren, M.-Y.; Feng, Z.; Guo, F.-Y.; Guo, J.-G.; et al. A Multi-Component Integrated Approach for the Elimination of Schistosomiasis in the People’s Republic of China: Design and Baseline Results of a 4-Year Cluster-Randomised Intervention Trial. Int. J. Parasitol. 2014, 44, 659–668. [Google Scholar] [CrossRef]
- Wang, T.-P.; Vang Johansen, M.; Zhang, S.-Q.; Wang, F.-F.; Wu, W.-D.; Zhang, G.-H.; Pan, X.-P.; Ju, Y.; Ørnbjerg, N. Transmission of Schistosoma japonicum by Humans and Domestic Animals in the Yangtze River Valley, Anhui Province, China. Acta Trop. 2005, 96, 198–204. [Google Scholar] [CrossRef]
- Liccioli, S.; Duignan, P.J.; Lejeune, M.; Deunk, J.; Majid, S.; Massolo, A. A New Intermediate Host for Echinococcus multilocularis: The Southern Red-Backed Vole (Myodes gapperi) in Urban Landscape in Calgary, Canada. Parasitol. Int. 2013, 62, 355–357. [Google Scholar] [CrossRef]
- Besier, R.B.; Kahn, L.P.; Sargison, N.D.; Van Wyk, J.A. The Pathophysiology, Ecology and Epidemiology of Haemonchus contortus Infection in Small Ruminants. In Advances in Parasitology; Elsevier: Amsterdam, The Netherlands, 2016; Volume 93, pp. 95–143. ISBN 978-0-12-810395-1. [Google Scholar]
- Sargison, N.D. Pharmaceutical Control of Endoparasitic Helminth Infections in Sheep. Vet. Clin. North. Am. Food Anim. Pract. 2011, 27, 139–156. [Google Scholar] [CrossRef]
- Playford, M.; Smith, A.; Love, S.; Besier, R.; Kluver, P.; Bailey, J. Prevalence and Severity of Anthelmintic Resistance in Ovine Gastrointestinal Nematodes in Australia (2009–2012). Aust. Vet. J. 2014, 92, 464–471. [Google Scholar] [CrossRef] [PubMed]
- Rose, H.; Rinaldi, L.; Bosco, A.; Mavrot, F.; De Waal, T.; Skuce, P.; Charlier, J.; Torgerson, P.R.; Hertzberg, H.; Hendrickx, G.; et al. Widespread Anthelmintic Resistance in European Farmed Ruminants: A Systematic Review. Vet. Rec. 2015, 176, 546-546. [Google Scholar] [CrossRef]
- Tsotetsi, A.M.; Njiro, S.; Katsande, T.C.; Moyo, G.; Baloyi, F.; Mpofu, J. Prevalence of Gastrointestinal Helminths and Anthelmintic Resistance on Small-Scale Farms in Gauteng Province, South Africa. Trop. Anim. Health Prod. 2013, 45, 751–761. [Google Scholar] [CrossRef]
- Várady, M.; Papadopoulos, E.; Dolinská, M.; Königová, A. Anthelmintic Resistance in Parasites of Small Ruminants: Sheep versus Goats. Helminthologia 2011, 48, 137–144. [Google Scholar] [CrossRef]
- Höglund, J.; Gustafsson, K.; Ljungström, B.-L.; Engström, A.; Donnan, A.; Skuce, P. Anthelmintic Resistance in Swedish Sheep Flocks Based on a Comparison of the Results from the Faecal Egg Count Reduction Test and Resistant Allele Frequencies of the β-Tubulin Gene. Vet. Parasitol. 2009, 161, 60–68. [Google Scholar] [CrossRef]
- Von Samson-Himmelstjerna, G.; Walsh, T.K.; Donnan, A.A.; Carrière, S.; Jackson, F.; Skuce, P.J.; Rohn, K.; Wolstenholme, A.J. Molecular Detection of Benzimidazole Resistance in Haemonchus contortus Using Real-Time PCR and Pyrosequencing. Parasitology 2009, 136, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Baltrušis, P.; Halvarsson, P.; Höglund, J. Utilization of Droplet Digital PCR to Survey Resistance Associated Polymorphisms in the β Tubulin Gene of Haemonchus contortus in Sheep Flocks in Sweden. Vet. Parasitol. 2020, 288, 109278. [Google Scholar] [CrossRef] [PubMed]
- Baltrušis, P.; Komáromyová, M.; Várady, M.; Von Samson-Himmelstjerna, G.; Höglund, J. Assessment of the F200Y Mutation Frequency in the β Tubulin Gene of Haemonchus contortus Following the Exposure to a Discriminating Concentration of Thiabendazole in the Egg Hatch Test. Exp. Parasitol. 2020, 217, 107957. [Google Scholar] [CrossRef]
- Elmahalawy, S.T.; Halvarsson, P.; Skarin, M.; Höglund, J. Genetic Variants in Dyf-7 Validated by Droplet Digital PCR Are Not Drivers for Ivermectin Resistance in Haemonchus contortus. Int. J. Parasitol. Drugs Drug Resist. 2018, 8, 278–286. [Google Scholar] [CrossRef]
- Baltrušis, P.; Charvet, C.L.; Halvarsson, P.; Mikko, S.; Höglund, J. Using Droplet Digital PCR for the Detection of Hco-Acr-8b Levamisole Resistance Marker in H. contortus. Int. J. Parasitol. Drugs Drug Resist. 2021, 15, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Baltrušis, P.; Halvarsson, P.; Charvet, C.L.; Höglund, J. The Presence and Relative Frequency Detection of the Levamisole-Resistance-Associated S168T Substitution in Hco-Acr-8 in Haemonchus contortus. Int. J. Parasitol. Drugs Drug Resist. 2023, 21, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Waller, P.J.; Chandrawathani, P. Haemonchus Contortus: Parasite Problem No. 1 from Tropics—Polar Circle. Problems and Prospects for Control Based on Epidemiology. Trop. Biomed. 2005, 22, 131–137. [Google Scholar]
- Ljungström, S.; Melville, L.; Skuce, P.J.; Höglund, J. Comparison of Four Diagnostic Methods for Detection and Relative Quantification of Haemonchus contortus Eggs in Feces Samples. Front. Vet. Sci. 2018, 4, 239. [Google Scholar] [CrossRef]
- Zarlenga, D.S.; Hoberg, E.P.; Tuo, W. The Identification of Haemonchus Species and Diagnosis of Haemonchosis. In Advances in Parasitology; Elsevier: Amsterdam, The Netherlands, 2016; Volume 93, pp. 145–180. ISBN 978-0-12-810395-1. [Google Scholar]
- Högberg, N.; Hessle, A.; Lidfors, L.; Enweji, N.; Höglund, J. Nematode Parasitism Affects Lying Time and Overall Activity Patterns in Lambs Following Pasture Exposure around Weaning. Vet. Parasitol. 2021, 296, 109500. [Google Scholar] [CrossRef] [PubMed]
- Höglund, J.; Baltrušis, P.; Enweji, N.; Gustafsson, K. Signs of Multiple Anthelmintic Resistance in Sheep Gastrointestinal Nematodes in Sweden. Vet. Parasitol. Reg. Stud. Rep. 2022, 36, 100789. [Google Scholar] [CrossRef]
- Högberg, N.; Hessle, A.; Lidfors, L.; Höglund, J. The Effect of Weaning Age on Animal Performance in Lambs Exposed to Naturally Acquired Nematode Infections. Vet. Parasitol. 2023, 316, 109900. [Google Scholar] [CrossRef]
- Baltrušis, P.; Halvarsson, P.; Höglund, J. Molecular Detection of Two Major Gastrointestinal Parasite Genera in Cattle Using a Novel Droplet Digital PCR Approach. Parasitol. Res. 2019, 118, 2901–2907. [Google Scholar] [CrossRef]
- Daş, G.; Abel, H.; Savaş, T.; Sohnrey, B.; Gauly, M. Egg Production Dynamics and Fecundity of Heterakis gallinarum Residing in Different Caecal Environments of Chickens Induced by Fibre-Rich Diets. Vet. Parasitol. 2014, 205, 606–618. [Google Scholar] [CrossRef]
- Wongrak, K.; Gauly, M.; Daş, G. Diurnal Fluctuations in Nematode Egg Excretion in Naturally and in Experimentally Infected Chickens. Vet. Parasitol. 2015, 208, 195–203. [Google Scholar] [CrossRef]
- Luna-Olivares, L.A.; Ferdushy, T.; Kyvsgaard, N.C.; Nejsum, P.; Thamsborg, S.M.; Roepstorff, A.; Iburg, T.M. Localization of Ascaridia Galli Larvae in the Jejunum of Chickens 3 Days Post Infection. Vet. Parasitol. 2012, 185, 186–193. [Google Scholar] [CrossRef]
- McDougald, L.R. Blackhead Disease (Histomoniasis) in Poultry: A Critical Review. Avian Dis. 2005, 49, 462–476. [Google Scholar] [CrossRef]
- Curry, E.; Prichard, R.; Lespine, A. Genetic Polymorphism, Constitutive Expression and Tissue Localization of Dirofilaria immitis P-Glycoprotein 11: A Putative Marker of Macrocyclic Lactone Resistance. Parasit. Vectors 2022, 15, 482. [Google Scholar] [CrossRef]


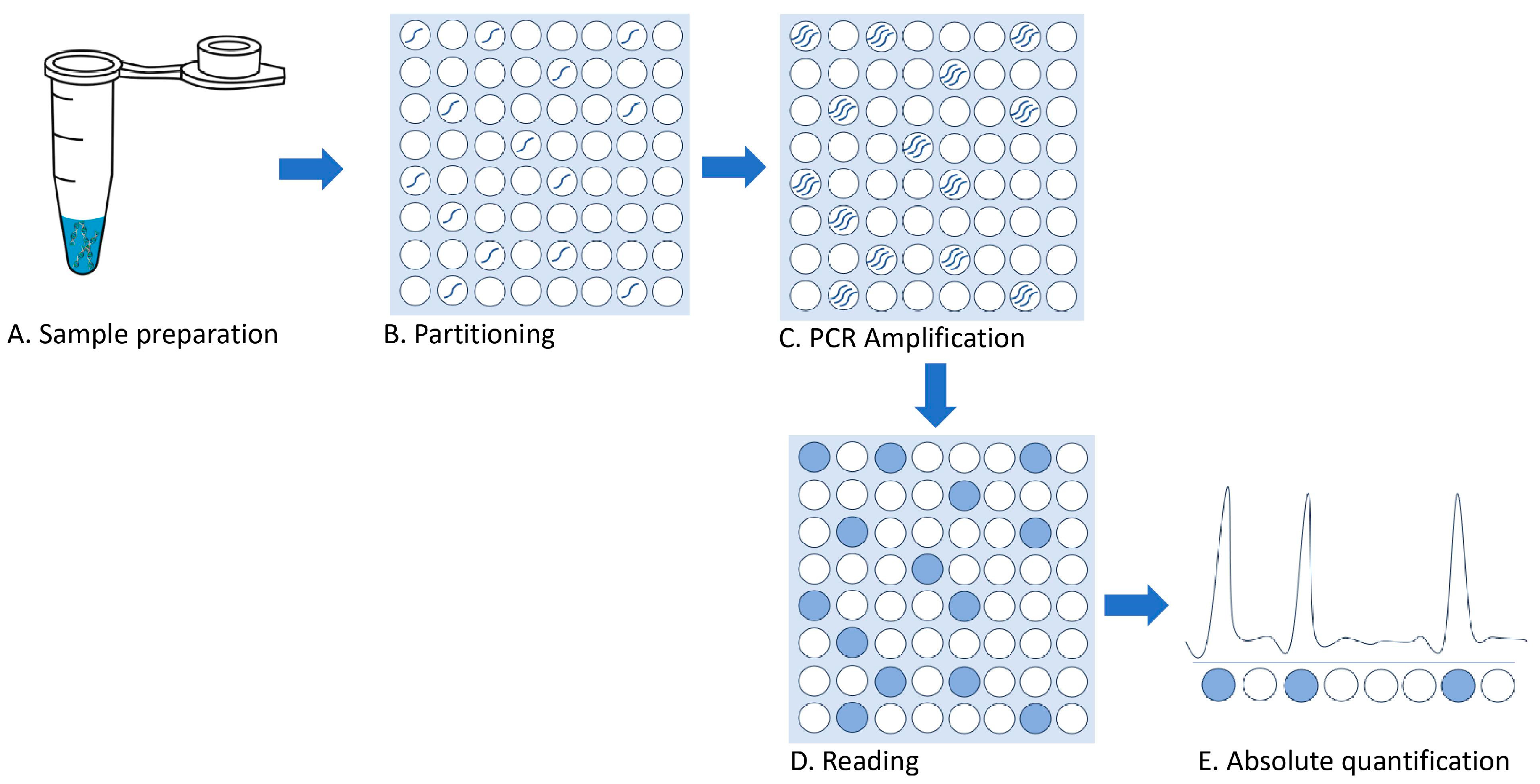{kind=link}
| Advantages | Disadvantages |
|---|---|
| Absolute quantification | High cost |
| No reference samples required | Inability to sequence the PCR amplicons |
| No standard curve required | |
| Better performance in the presence of inhibitors | |
| High sensitivity |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsokana, C.N.; Symeonidou, I.; Sioutas, G.; Gelasakis, A.I.; Papadopoulos, E. Current Applications of Digital PCR in Veterinary Parasitology: An Overview. Parasitologia 2023, 3, 269-283. https://0-doi-org.brum.beds.ac.uk/10.3390/parasitologia3030028
Tsokana CN, Symeonidou I, Sioutas G, Gelasakis AI, Papadopoulos E. Current Applications of Digital PCR in Veterinary Parasitology: An Overview. Parasitologia. 2023; 3(3):269-283. https://0-doi-org.brum.beds.ac.uk/10.3390/parasitologia3030028
Chicago/Turabian StyleTsokana, Constantina N., Isaia Symeonidou, Georgios Sioutas, Athanasios I. Gelasakis, and Elias Papadopoulos. 2023. "Current Applications of Digital PCR in Veterinary Parasitology: An Overview" Parasitologia 3, no. 3: 269-283. https://0-doi-org.brum.beds.ac.uk/10.3390/parasitologia3030028