
Sarcocystis Species (Apicomplexa, Eucoccidiorida) Parasitizing Snakes
1
Departamento de Anatomia, Patologia e Clínicas Veterinárias, Escola de Medicina Veterinária e Zootecnia, Universidade Federal Bahia, Salvador 40170-110, Brazil
2
Departamento de Medicina Veterinária Preventiva e Saúde Animal, Faculdade de Medicina Veterinária e Zootecnia, Universidade de São Paulo, São Paulo 05508-270, Brazil
*
Author to whom correspondence should be addressed.
Parasitologia 2023, 3(4), 327-347; https://0-doi-org.brum.beds.ac.uk/10.3390/parasitologia3040032
Submission received: 15 September 2023
/
Revised: 30 October 2023
/
Accepted: 3 November 2023
/
Published: 10 November 2023
(This article belongs to the Special Issue Sarcocystis in Domestic and Wildlife Animals)
Abstract
:The genus Sarcocystis has approximately 200 species that are able to infect homeothermic and poikilothermic animals. So far, 23 Sarcocystis spp. have snakes as definitive hosts (DHs) and mammals and/or reptiles as intermediate hosts (IHs). Some of these species cause disease or even death in their IH. At least two Sarcocystis spp. from snakes have relevance in public health. Sarcocystis nesbitti causes serious myopathy in humans, and S. singaporensis is lethal for rats and was successfully tested in the biological control of these rodents. Molecular identification was not reported for half of Sarcocystis spp. shed by snakes. For some snake species, their role as DHs for Sarcocystis spp. is totally unknown, including certain snakes which are bred as companion animals. The aim of this review is to discuss the role of snakes as DHs or IHs of Sarcocystis spp. and the future directions for the identification of the parasites and the elucidation of their life cycles.
Keywords:
Sarcocystidae; reptile; sporocyst; bradyzoite; sarcocyst; small mammal; lizard; definitive host; intermediate host1. Introduction
Sarcocystis spp. are cyst-forming parasites of the phylum Apicomplexa, class Sporozoasida, subclass Coccidiasina, order Eucoccidiorida and family Sarcocystidae. Parasites of the genus Sarcocystis infect a large spectrum of vertebrate homeothermic and poikilothermic animals, including mammals, birds and reptiles [1]. The family Sarcocystidae encompasses the genera Sarcocystis, Toxoplasma, Besnoitia, Hammondia, Neospora and Cystoisospora [2]; the genus Frenkelia used to belong to the family Sarcocystidae, but phylogenetic studies led to its placement in the genus Sarcocystis [3,4,5]. The family Sarcocystidae can be divided in two categories: parasites with an obligate two-host life-cycle, including Hammondia spp. and Sarcocystis spp. [6,7], and those with a facultative two-host life-cycle, such as Toxoplasma gondii and Neospora caninum [8]. A unique characteristic of Sarcocystis spp. is the sporulation of oocysts in the intestinal tract of their DHs; as their oocysts possess a fragile and delicate wall, fully sporulated sporocysts are released from oocysts in the intestinal lumen of the DHs and represent the predominant parasitic stage shed in their feces. Oocysts of other Sarcocystidae genera are shed in the nonsporulated form in the feces of the DHs [8].
The DHs of Sarcocystis spp. are carnivores or omnivores who are capable of preying on and ingesting encysted bradyzoites in tissues of the preyed-on animals. Bradyzoites ingested by a DH undergo other parasitic stages that culminate with the formation of infective oocysts and sporocysts in their intestinal tracts. Approximately one fourth of the over 200 described species of Sarcocystis sp. have domestic or wild canids and felids as DHs; until 2016, complete life cycles were known for 26 Sarcocystis spp. [1]. In wildlife, an increasing number of predator reptiles have been identified as DHs of this parasite, including snakes. The aim of this review is to discuss the role of snakes as DHs and IHs of Sarcocystis spp. and their potential impact on animal and public health.
2. Materials and Methods
The literature search for any paper related to Sarcocystis spp. in snakes was carried out using the following databases: Scopus “https://0-www-scopus-com.brum.beds.ac.uk/” (accessed on 2 May 2023), Pubmed “https://pubmed.ncbi.nlm.nih.gov/ (accessed on 2 May 2023) and Google Scholar “https://0-scholar-google-com.brum.beds.ac.uk/” (accessed on 2 May 2023). A broad number of keywords was used and included Sarcocystis, Sarcocystidae, sarcocyst, sarcosporidiosis, sarcocystosis, and cyst-forming, which were combined with snake and reptile. Articles in English and in other languages were retrieved from these databases, as well as textbooks that were related to the topic of the current review.
3. Snakes as Definitive Hosts of Sarcocystis spp.
In the early 1970s, the role of snakes as DHs of Sarcocystis sp. was demonstrated in Australia [9]; sporocysts of the parasite were shown in the feces of five Australian carpet pythons (Morelia spilotes variegata). Intestinal scrapings of the snakes had fully sporulated oocysts, which contained delicate walls and sporulated sporocysts. The bush rat (Rattus fuscipes) was suspected to be the IH of the parasite, as a high percentage of these rats had previously been confirmed to have high rates of sarcocysts in their skeletal and subcutaneous musculature [9]. Therefore, it has been hypothesized that reptiles could also participate in the life cycle of certain species of Sarcocystis. These findings were connected to the discovery of cysts that seemed to be Sarcocystis sp. in bush rats (Rattus fuscipes) from the same region. Both rat and snake had formerly been associated in the food chain, as they were shown to participate in a nematode life cycle as IHs and DHs of the parasite, respectively. The initial hypothesis was validated through a successful laboratory experiment using Norway rat (Rattus norvegicus) [9].
Also in the 1970s, Sarcocystis singaporensis was identified in snakes, whose original species name was S. orientalis [10,11]. Sarcocystis singaporensis, which has reticulated python (Python reticulatus) as its main DH, was further re-described, and two additional species were identified: S. zamani and S. villivillosi [12]. Up to now, S. singaporensis has been one of the most studied species in snakes due to its endemic situation in Southeast Asia, and for causing death in certain species of rats [13,14,15]. Through experimental infections, Timor python (Python timorensis), black headed python (Aspidites melanocephalus) and African rook python (Python sebae) were identified as additional DHs of S. villivillosi and S. singaporensis [16].
Sarcocystis murinotechis was classified in tiger snakes (Notechis ater) from Australia and was shown to have different species of murids to IHs [17]; it was differentiated from S. singaporensis and Rzepchzyk’s Sarcocystis sp., which also have rats as the IH. Since the DH of S. singaporensis did not occur in Australia, experimental infections were performed to identify the hosts of S. murinothecheis. The DH of S. murinotechis, the tiger snake Notechis ater, is the sole representative of the Family Elapidae of Sarcocystis spp. infecting snakes, and was proved to be a different species from S. singaporensis [17].
One of the most important species that cycles in snakes is Sarcocystis nesbitti, which is zoonotic and apparently endemic in Southeast Asia; it causes muscular sarcocystosis in humans [18,19,20]. Based on phylogenetic evidence suggesting that snakes could be the DH of this parasite [21], studies were conducted to confirm this hypothesis. Molecular evidence of S. nesbitti in snakes’ stools reinforced this assumption. The reticulated python (Python reticulatus) and the monocled cobra (Naja kaouthia) were considered putative DHs for this species, but no sporocysts were observed in their feces [22,23]. Potential sporocysts of S. nesbitti were detected in scrub python (Simalia amethistina) in Australia [13]. This was an unexpected finding due to the geographical barrier and the lack of the stated IHs of this species in the Australian territory. Thus far, the DH of S. nesbitti in Southeast Asia remains unknown.
Viperid snakes are only hosts for Sarcocystis spp. in the snake–rodent life cycle. Most of these species were detected in the African continent. The Viperidae family is divided into two subfamilies: Viperinae and Crotalinae. Specimens of the former are found in Africa, Asia and Europe, and the latter in Asia and in the Americas [24]. Sarcocystis crotali was identified in Mojave rattlesnake (Crotalus scutulatus scutulatus) in the United States [25]. Little is known about Sarcocystis spp. parasitizing members of the subfamily Crotalinae, but some studies were reported in the United States and Brazil [26,27,28,29].
Snakes of the genus Bittis serve as DHs for Sarcocystis dirumpens and Sarcocystis hoarensis [30,31]. Sarcocystis hoarensis had previously been believed to be Besnoitia sp., due to the localization of the cysts and to its macroscopic aspect in the skin [32]; ultrastructural studies of its sarcocyst showed typical characteristics of Sarcocystis sp. and it was reclassified in this genus [30]. Out of the few biological experiments performed with this family group, S. atheridis and S. gerbilliechis exhibited certain host specificity [33,34]. The latter showed a narrow specificity with the species level, and S. atheridis to the genus. Nevertheless, regarding the employed experimental host range, it would be necessary to enlarge the spectrum of tested animals to confirm this observation. Sarcocystis muriviperae was identified in two animals of distinct families, and seems to be the only named species less specific to the DH [35,36,37], as infection by Sarcocystis nesbitti in Naja kaouthia was not truly confirmed [22,23].
Colubrid snakes act as DHs of a great number of Sarcocystis species. Up to now, for all Sarcocystis spp. that share a snake–reptile life cycle, the DH has been found as a snake of the family Colubridae. Seven species of this family also cycle between mammals [1,38,39]. This fact may be related to the wide distribution of Colubridae, since it is recognized as the most voluminous taxon in the order Squamata, with a great number of genera and species [40,41]. Most of the Sarcocystis spp. of colubrids were detected in Africa, Europe and Asia. A few reports were from the Americas. In the snake–rodent cycle, S. idahoensis and S. pantherophisi are reported. Sarcocystis idahoensis has the Great Basin gopher snake (Pituophis melanoleucus deserticola) as DH [42,43], whereas S. pantherophisi has the eastern rat snake (Pantherophis alleghaniensis) as DH [38]. Their major importance relies in the induction of clinical signs in their IH (Section 3).
Sarcocystis spp. life cycles in colubrids, in the snake–rodent or in the snake–reptile cycles, was demonstrated to be more specific to the DH in the level of genus or even species than its IH [44,45]. Some snake species are hosts for more than one species of Sarcocystis, a fact that had already been observed in other Sarcocystis spp. in different hosts [12,16,44].
Based on current knowledge, Sarcocystis zuoi was the only species with strict host specificity; the king rat snake (Elaphe carinata) is its only known DH [45]. This might be related to the lack of experimental infections in other species such as S. pantherophisi [38] and S. attenuati [39], for example. In an experiment performed by Hu et al. [45], towards the identification of the DH for S. zuoi, the beauty snake (E. taeniura) was ruled out of its host range. Later, this snake species was identified as the natural DH of S. attenuati in Asia. Although they had been classified as different species with distinct sarcocyst wall morphologies, they had similar 18S rDNA and ITS1 sequences [39]. Sarcocystis attenuati and S. clethrionomyelaphis share the same DH, Elaphe taeniura, and they were isolated from the same geographical region in China. However, they differ in morphology, and phylogenetically they are placed in distinct subgroups based on 18S rDNA.
Species such as S. clethrionomyelaphis and S. muriviperae are the exceptions to the DH specificity, as they use three and two different genera as hosts, respectively; S. muriviperae also use animals of distinct families of snakes as DHs. Nevertheless, when S. clethrionomyelaphis was first identified, all natural and experimental species belonged to the genus Elaphe [46], but its taxonomy later changed. Thus, nowadays, three genera are reported as DHs. Moreover, evidence indicates that this species may be widely spread, regarding the large range of DHs native to distinct regions, associated with the fact that it was detected in two continents [46,47].
Results of experimental studies with S. muriviperae demonstrated an interesting fact in terms of its IHs. Sporocysts obtained from naturally infected Palestine viper (Vipera palaestinaea) and black whipsnake (Coluber jugularis) demonstrated differences in the applied doses to induce the same clinical signs in the early stages of merogony [35]. Although they were assumed to be the same species, morphologically and biologically, a cross-infection experiment using both isolated species was not performed to elucidate whether they are in fact the same species. Molecular analysis would help to clarify the discrepant results following infection in the DH.
The genus Coluber sp. was the first snake taxon related to the snake–reptile cycle [48] and it is known to harbor three species of Sarcocystis, besides S. muriviperae: S. podarcicolubris [48], S. chalcidicolubris [49] and Sarcocystis stenodactycolubris [50]. Although they have the DH in common, they differ in morphological characteristics of the sarcocyst wall and in the parasite’s biology. Sarcocystis chalcidicolubris [49] and Sarcocystis stenodactycolubris [50] were observed in lizards of different genera and families, respectively; they differ from those Sarcocystis spp. previously reported [48,51]. The distinction between these species and S. podarcicolubris was based on sarcocyst morphology and the parasite’s biology. Biologically, the IHs of S. chalcidicolubris did not acquire infection after ingesting sporocysts from S. podarcicolubris; S. stenodactycolubris was only infective to a genus of the gekkonidae family, indicating that they are different species.
Sarcocystis lacertae has an IH in common with S. podarcicolubris. However, experimental infections involving the latter species evidenced that specimens of the genus Malpolon, Elaphe and Coronella were not infected under experimental infections with S. podarcicolubris [51]. Posteriorly, the smooth snake (Coronella austriaca) was identified as the natural DH for Sarcocystis lacertae [52]. S. lacertae DNA was detected in the feces of a black-headed cat snake (Boiga nigriceps) from Malaysia [53], but sporocysts were not seen in the stools. Regarding geographic barrier, as the genus Podarcis does not occur in Southeast Asia [54], this finding may be related to another species closely related to S. lacertae or the IH might be other Lacertidae species in the island.
Through experimental assessment, the diadem snake (Spalerosophis diadema) was found to be the DH for Sarcocystis gongyli; besides sharing the same IH genus of S. chalcidicolubris, they differ in DHs and sarcocysts’ morphology [55]. The diadem snake was also identified as the DH of Sarcocystis acanthocolubri through transmission experiments [56], and as in the experiment performed for S. gongyli [55], the other two tested snakes were not infected.
All the above-mentioned species that use other reptiles as IHs were found in Europe, Africa and Asia. Nowadays, there are a few species of Sarcocystis classified in the Americas [38,42,43], and just one reported to have a snake–lizard cycle, namely Sarcocystis ameivamastigodryasi [57]. This species, described in Brazil, was the first reported of reptiles as IHs of Sarcocystis sp. from snakes in the Americas. The Boddaert’s tropical racer (Mastigodryas boddaerti) and the swamp racer (M. bifossatus) act as DHs. In this experiment, one specimen of the common boa (Boa constrictor) was tested; however, it was not infected [57]. Sarcocystis spp. that use snakes as DHs are summarized in Table 1 and Table 2.
Published reports of unnamed Sarcocystis spp. from snakes are available in numerous databases; nonetheless, besides the detection of the parasite, a few have additional information such as morphologic assessment, molecular and phylogenetic analysis, or experimental transmission tests [27,29,70,71,72].
In South America, most studies about Sarcocystis spp. in snakes correspond to general parasitic surveillance in snakes without further tests for species identification [26,28]. The presence of sporocysts in the feces of two specimens of common boas (Boa constrictor constrictor) and one rainbow boa (Epicrates cenchria cenchria) were reported in the north of Brazil. No additional tests were performed [57]. Recently, sporocysts shed by a single specimen of B. constrictor were detected and molecularly characterized in the northeast of Brazil [72]. Based on 18S rDNA and Cox1 markers, the isolate obtained from the B. constrictor is similar to Sarcocystis sp. isolated from the rodent plains viscacha (Lagostomus maximus) in Argentina [73]. As this Argentinian rodent species does not occur in the northeast of Brazil, it is possible that a different species of rodent acts as the IH of this Sarcocystis sp. derived from B. constrictor.
Although molecular analysis is mandatory to discriminate Sarcocystis spp., this methodology alone is not sufficient for a definitive diagnosis of a snake as the DH of the parasite. The multiplication of the parasite in the intestinal tract of a snake should be confirmed in order to prove that the shed sporocysts were indeed generated in the snake; a snake may shed sporocysts that already existed in any preyed animals. However, molecular evidence of infection indicates which Sarcocystis species may occur in that area [22,23,53,74] and may help to direct further investigations.
4. Snakes as Intermediate Hosts of Sarcocystis spp.
Based on the prey–predator life cycle of Sarcocystis sp. [1], and the fact that some species of snakes are preyed on by other animals, snakes might serve as the IH for this coccidian. Some mammals, birds of prey such as Falconiformes, and reptiles (including other snakes) are known to prey on snakes [75,76,77]. Although reports on Sarcocystis sp. infecting snakes focus their role as DHs [1], a few studies show snakes as IHs of the parasite [78,79,80]; in all of these reports, the DHs remain unknown (Table 3).
The first report of the presence of sarcocysts in snakes was in an Australian carpet snake or Australian Python (Morelia argus). This species was named Sarcocystis pythonis. It was described as having a thin-walled cyst with a type I wall [79]. The other species, Sarcocystis atractaspidis, was identified later from two burrowing vipers (Atractaspis sp.). In this case, sarcocysts were observed in the mesentery and lungs and were macroscopic [80]. In both reports, these species were considered inquirendae, because no visual evidence (images) was reported in these papers, besides the lack of TEM analysis, differential morphological descriptions and molecular tests [1,82].
Through molecular analysis, Sarcocystis sp. was incidentally detected in tissues of the snakes Hemorrhois sp. and Malpolon sp. during an investigation of Hepatozoon sp. using the 18S rDNA gene. GenBank screening indicated that the closest match was S. lacertae, which uses snakes as the DH and lizards as the IH [78]. In the following year, sequences of 28S rDNA gene were reported from the Sarcocystis sp. of a Psammophis sp. snake in the Mediterranean region [83]. A phylogenetic analysis was performed, but the obtained results did not clarify the prediction of the DH; the obtained nucleotide sequences did not form a monophyletic group with other Sarcocystis infecting snakes, and the available sequences were placed in unrelated groups [83]. A number of studies indicate that natural Sarcocystis spp. groups are mainly influenced by their DH correlations, as opposed to their relationships with IHs [84,85]. Molecular evidence of Sarcocystis sp. in tissues of the crowned leafnose snake (Lytorhynchus diadema) was reported, although no phylogenetic analysis was conducted [86]. In these three studies, only molecular evidence was shown for Sarcocystis spp. infecting snakes’ tissues.
A South American rattlesnake (Crotalus durissus terrificus) was confiscated from an illegal owner in Tennessee. The animal died in custody and a necropsy was performed. Macroscopically, no lesions or cysts were observed in the animal’s tissues. Under light microscopy, cysts measuring up to 2000 µm in length were visualized in the skeletal muscles from three areas near the vertebrae, as well in the tongue and head. The sarcocysts’ wall was thin and classified as type I. At the time, there was no evidence of active merogony. The authors considered the cyst morphology distinct from the usual sarcocyst form in some aspects, leading them to hypothesize the possibility of the snake being an aberrant IH [82].
A molecular analysis of Sarcocystis sp. from the South American rattlesnake demonstrated a high similarity with Sarcocystis mucosa isolated from Australian marsupials; this finding led the authors to hypothesize that an opossum species from South America, which is also a marsupial and shares the same native habitat as this snake species, could be its DH. However, the nucleotide sequence of 18S rDNA obtained in this study was relatively short, and no phylogenetic test was performed. Other animals have been suggested as DHs based on their food habits, but this remains unclear [82]. In another study, 99% identity was found for L. diadema isolate and Sarcocystis sp. from the Virginia opossum (Didelphis virginiana) from Mexico and Sarcocystis neurona from fisher (Martes pennanti). Thus, it was suggested that the probable DH could be an omnivorous or carnivorous mammal [86].
Recently, the presence of Sarcocystis sp. in the head, trunk and tail muscles of the green tree python (Morelia cf. viridis) has been reported [81]. This parasite was classified as a new species and was named Sarcocystis moreliae, based on morphological and molecular characteristics. This isolate differs from the one found in C. d. terrificus in terms of size and cyst wall thickness. Molecular analyses successfully generated sequences of the 18S rDNA, 28S rDNA, ITS1, and cox1 loci. Despite obtaining robust sequences, the comparison with other species that infect reptiles as IHs was limited due to the lack of genetic information, such as for the ITS-1 region or very short sequences available. The absence of morphological information also hampered the comparison between species. Phylogenetic analysis could not predict the DH of the species. Nevertheless, the authors proposed that the DHs of this species might include monitors, birds, birds of prey, quolls, or even other snakes [81].
5. Intermediate Hosts of Sarcocystis spp. from Snakes
Serpents have a wide range of animals in their food spectrum, including mammals, birds, reptiles and amphibians, depending on the snake species, life habits and body size [87,88]. Nonetheless, only mammals and reptiles have been recorded as IHs of Sarcocystis spp. which use snakes as DHs [1,38,39] (Figure 1). We hypothesize that birds that commonly feed on the soil are more exposed to the ingestion of sporocysts and possibly serve as IHs for certain species of Sarcocystis that cycle in snakes. Sarcocystis spp. that have mammals as IHs are more numerous when compared to those that use reptiles, embracing 16 out of 23 species [1,38,39]. Initial descriptions of Sarcocystis sp. were mainly based on sarcocysts’ morphology [31,48,51]. However, the morphology of sarcocyst walls is not reliable because some walls may be almost identical among certain species and may be affected during sample processing or may be changed with age [89]. Molecular analysis is a valuable and necessary resource for the classification of Sarcocystis on the species level, besides morphological and biological aspects of the parasite and animal hosts [3,90,91].
Rodents comprise the largest order in the class Mammalia, encompassing more than 2000 animal species and almost 500 genera [92]. Sarcocystis spp. of snakes are reported to infect around 20 genera of rodents (Table 1). Sarcocystis spp. shed by snakes are less specific to their IHs than to the DHs. Sarcocystis singaporensis, for example, infects at least 12 IHs of the genus Rattus and Bandicota [16]. Sarcocystis dirunpens and S. hoarensis infect six different animal genera belonging to two families: Muridae and Cricetidae [31]. Exceptions are observed as well, and a single animal species acts as IH for S. muriviperae [37]. Other Sarcocystis species that demonstrate specificity to their IHs are S. idahoensis, S. zamani, S. zuoi, S. pantherophis, S crotali and S. attenuati; however, with exception of S. idahoensis, experimental infections were not performed for the remaining Sarcocystis spp., or were conducted with a limited range of potential IHs for these Sarcocystis spp. Moreover, attention must be given to S. pantherophis, since the only IH is an experimental immunosuppressed mouse (KO Mus musculus); thus, the putative IH could be a different murid. Although S. gerbilliechis has also been shown to have low IH specificity on the genus and species levels, only members of the subfamilie Gerbillinae were demonstrated to be IHs of the parasite under experimental infections [34].
The genus Rattus is reported to be an IH for S. zuoi, S. zamani, S. villivilosi, S. murinotechis and S. singaporensis. Out of these species, S. murinotechis and S. singaporensis stand out due to their pathogenicity to their IH. Sarcocystis singaporensis is one of the most studied Sarcocystis spp. of snakes [93] and this species is highly lethal to different species of Rattus sp. and to the short-tailed bandicoot rat (Nesokia indica) [14]. Due to its pathogenicity to IH, S. singaporensis was tested as an efficient biocontrol agent for non-native rats [14,15,94].
Voles serve as IHs for S. clethrionomyelaphis [46,47,60]. Initially, only the genera Microtus and Clethrionomys were described as IHs for S. clethrionomyelaphis, and, at that time, these animals were classified as IHs under experimental infections [46]. Years later, the large oriental vole (Eothenomys miletus) was identified as a natural IH of S. clethrionomyelaphis through morphology and molecular analysis [47]. This vole is endemic to Yunnan Province, China [95], and the frequency of S. clethrionomyelaphis infection in these animals was 75.8% [47], demonstrating that this Sarcocystis species is spread in the region. Recently, a new species was classified as S. attenuati from the Asian gray shrew (Crocidura attenuata) in the same region where S. clethrionomyelaphis was found; morphological and molecular features were employed to differentiate S. attenuati from S. clethrionomyelaphis. Still, different from the other Sarcocystis spp., sarcocysts from S. attenuati were also found in the heart of the IH [39].
Intermediate hosts of snakes’ Sarcocystis spp. are more susceptible to developing clinical signs than the DHs. In contrast to reptiles as IH, rodents have been demonstrated to be more susceptible to developing a pathology caused by Sarcocystis spp. The occurrence of lesions is induced by early stages of the parasite and is associated with the immune response of the host and the dose of parasites [1,33]. In most Sarcocystis spp. that cause acute sarcocystosis, the pathology is associated with damage in the liver. This may be observed in S. idahoensis, S. muriveperidae and S. atheridis, where meronts were observed in hepathocytes. Hepatosplenomegaly, anorexia, weakness and icterus are some of the clinical signs commonly observed [33,37,42]. The pathology is so severe that it can lead the animal to death. Specificity is also noted in the pathogenicity level. The same infective dose of S. atheridis was given to both IHs of this species; nevertheless, only the barbary striped grass mouse (Lemniscomys barbarus) was demonstrated to be susceptible to disease. Sarcocystis gerbilliechis also cause hepatopathy in their IHs; however, the lesions are focused in the liver parenchyma and endothelial cells of other organs and usually the animals survive to the acute signs [34].
Other pathologies reported in IHs of Sarcocystis spp. from snakes are neurological and respiratory alterations. Meronts of S. pantherophisi were seen in the cerebellum of gamma-interferon gene knockout mice (Mus musculus) and were related to clinical signs observed in these animals. Early stages of the parasites Sarcocystis murinotechis and S. singaporensis can be found in the endothelium of different tissues through the body; however, lungs are the most infected tissue, leading to severe respiratory alterations and culminating in death [10,12,17]. In both species, the induction of edematous lungs and effusion were observed, but hemorrhage was only reported for S. singaporensis. Dyspnea was observed in IHs susceptible to the disease caused by S. singaporensis [14].
Molecular diagnosis and phylogenetic analysis results may predict the DH for certain Sarocyctis spp. Some sarcocysts found in rodents were identified and classified even without knowledge of their DH. Through phylogenetic data it is possible to assume the suitable DH, as observed for S. eothenomysi and S. scandentiborneensis [96,97].
Reptiles from the Saurian families Lacertidae, Scincidae and Gekkonidae are known to be IHs for seven species that have snakes as DHs; these species are as follows: S. podarcicolubris [48], S. gongyli [68], S. stenodactylicolubris [50], S. chalcidicolubris [49], S. acanthocolubri [56], S. ameivamastigodryasi [57], S. lacertae [66] (Table 2).
Sarcocystis podarcicolubris was the first species found to exhibit a life cycle involving two reptiles, with a lizard being the IH [48]. Initially, two species from the genus Podarcis were described as IHs following experimental infections [48]. In the same paper, a reference was made to a report on a different species of Podarcis sp., sharing close characteristics with S. podarcicolubris; however, this species was not formally named. Subsequently, experimental transmission defined the genera Lacerta, Podarcis and Algyroides as additional IHs for S. podarcicolubris [51]. Regarding the extensive range of IHs for reptilian sarcosporidia, other lizard genera that could be infected by S. podarcicolubris were tested, and fortuitously a distinct sarcocyst was detected in the musculature of Chalcides spp. (eyed skink, Chalcides ocellatus; Gran Canaria skink, Chalcides sexlineatus; West canary skink, Chalcides viridanus). A new species was postulated and designated as Sarcocystis chalcidicolubris, based on observations made through light and electron microscopy, as well as on the biology of the parasite [49].
Trinci [68] described a cyst found in the muscles of an eyed skink (Chalcides ocellatus) and named it Sarcocystis gongyli. Years later, numerous sarcocysts in the muscles of the skink from a different genus were described, although the DH was not defined [98]. Posteriorly, a study revealed the presence of sarcocysts in skeletal muscles of a native skink (Chalcides ocellatus) from Egypt, and through light and transmission electron microscopy, these sarcocysts were identified according to Trinci’s classification (S. gongyli) [55]. Despite sharing the same IH, S. gongyli and S. chalcidicolubris were proved to be distinct species based on differences in the sarcocyst’s morphology and experimental hosts.
Babudieri [66] re-described a species initially found by Lühe [99] in a common wall lizard (Podarcis muralis) and designated it Sarcocystis lacertae; nonetheless, there was no evidence of its DH. Sporocysts were isolated from a colubrid snake, the smooth snake (Coronella austriaca); then, experiments were conducted to identify its possible IH, and Podarcis muralis was the only species that became infected [52]. As S. podarcicolubris shares the same IH as S. lacertae, it could potentially be the same species, but the life cycle trials demonstrated specificity related to the DH, and certain morphological differences kept them classified as separate species [48,52].
Chronologically, until the 1990s, only lizards from the families Lacertida and Scincidae had been identified as IHs for Sarcocystis species with snakes as DHs [44,48,55]. Sarcocystis stenodactycolubris was the first identified species that uses geckos as IHs [50]. Sarcocysts had been found in gekkonids skeletal muscles [100,101,102,103], and were classified as Sarcocystis platydactyli [104]. However, the DH of this species remains unknown to date. Modry et al. [50] isolated sporocysts from a Colubridae snake and performed experimental infections using six different lizard and two rodent species. Only geckos (Fan-fingered gecko, Ptyodactylus guttatus, and Jordan short-fingered gecko, Stenodactylus grandiceps) were found to be infected and sarcocysts were observed in tail, limb and tongue muscles.
The giant ameiva or Amazon racerruner (Ameiva ameiva) was identified as the IH of S. ameivamastrigodryasi through both natural and experimental infections. This Sarcocystis species is the only one with a snake–reptile cycle classified in the Americas until now [57]. Sarcocystis acanthocolubri is currently the most recent species known to have snakes as DHs and other reptiles as IHs [56]. The assessment revealed two differently sized cysts in all the studied species (Bosc’s fringe-toed lizard, Acanthodactylus boskianus; Nidua fringe-toed lizard, Acanthodactylus sculentus; and leopard fringe-fingered lizard, Acanthodactylus pardalis). However, it was demonstrated that they belong to the same species [56]. All Sarcocystis spp. detected in other reptiles as IHs were classified solely based on morphological and biological findings through natural or experimental infections. Molecular screening would elucidate whether these species are unique or not, and would help to identify other species.
6. Infection in Humans by Sarcocystis nesbitti
A few Sarcocystis species are known to infect humans and, in some cases, are capable of causing disease [19,20,105]. Humans serve as DHs or IHs in the life cycle of certain Sarcocystis spp. They act as DHs after consuming mature sarcocysts present in undercooked meat [89]. Usually, humans do not develop any symptoms and remain asymptomatic. But in certain cases, this intestinal sarcocystosis culminates in a gastrointestinal condition that is mostly self-limited and bland [105]. When humans ingest water or food contaminated with sporocysts, they might develop muscular sarcocystosis. In such cases, they may experience some symptoms such as fever, myalgia, headache, fatigue and arthralgia [19,20,89,106].
Early descriptions of Sarcocystis spp. in humans had been based on the sarcocyst wall morphology [89]. However, this method of classification and species distinction is questionable, as mentioned before. More than 100 studies in Europe, Latin America, Africa and Asia have reported sarcocysts in human tissues [19,20,89,107,108]. Most of the reported cases are concentrated in Southeast Asia. Some of these cases had been published a long time ago as isolated occurrences, and the diagnosis was based on symptoms, epidemiology and, in some cases, by biopsy findings. During that time, no species had been truly identified, but sarcocysts were morphologically described.
Southeast Asia has been known as an endemic region for acute muscular sarcocystosis (AMS) [89,109]. However, epidemiological surveillance of human AMS in this region is considered scant [19]. In a study in Malaysia, the prevalence of 21% of Sarcocystis spp. cysts was found in 100 humans’ tongues [18]. The samples were obtained from autopsies in a university hospital in Malaysia. A survey for sarcocysts in more than 1500 samples of limb muscles was undertaken in the same institute and no sarcocysts were observed (unpublished data) [20]. A seroprevalence survey was also carried out [110]; however, serology does not allow a distinction between muscular and intestinal sarcocystosis [111].
In the 1990s, an outbreak of human AMS was reported involving militaries from the U.S. that were in Malaysia for work. In this report, seven out of fifteen individuals exhibited similar symptoms in some degree, but only one case was confirmed through biopsy. The other patients were assumed to have AMS based on the common exposure, symptoms and laboratorial changes [106]. In the past decade, a series of outbreaks of AMS in Tioman Island and Pangkor in Malaysia were published, leading to an in-depth investigation with many affected people [19,20,23,109]. One of the biggest outbreaks to happen in Southeast Asia was a case involving 100 individuals in different periods between 2011 and 2012; most of them were travelers from Europe, and the initial diagnosis was based on clinical and laboratorial patterns of myositis. A biopsy was performed in eight patients, with six showing typical histological myositis, and sarcocysts were visualized in two cases [19]. In 2012, another outbreak was reported involving 89 symptomatic individuals who had visited Pangkor Island [20,109]. In this outbreak, sarcocysts were visualized through biopsy and the causative agent was identified by PCR; the 18S rDNA sequences were from Sarcocystis nesbitti.
Later, a previously reported AMS outbreak [19] was re-described [112], extending the study period to 2013 and considering 62 out of 99 cases as probable AMS with 6 confirmed cases. They performed a molecular analysis on a single sample of a patient accessing the 18S rDNA gene and identified S. nesbitti as the causative agent of the infection. In both cases [20,112], the authors presumed that all symptomatic patients were probably infected with the same species, given their shared location, similar travelling habits and symptoms. In recent years, a confirmed case of S. nesbitti was reported involving one Vietnamese person who traveled to Malaysia [108]. Humans are known to be IHs of S. nesbitti. Nevertheless, humans are reported as accidental hosts, because they do not have any natural predators to move the cycle forward [89,113].
Sarcocystis nesbitti was first mentioned in 1969 by Mandour [62], who found a different cyst in the muscles of two Rhesus monkeys (Macaca mulatta) from North India. However, the validity of S. nesbitti was questioned [63,114] due to the lack of TEM analysis, the absence of photomicrographs and the controversial description of cyst morphology. Afterward, sarcocysts were identified in the musculature of a long-tailed macaque (Macaca fascicularis) from a monkey farm in Yunnan Province, China. The sarcocysts were thoroughly described through LM and TEM, leading to their classification as a different species from Sarcocystis kortei, and then classified as S. nesbitti [63]. Based on ultrastructure, four other reports in monkeys [115,116,117,118] and two in humans [119,120] were mentioned as S. nesbitti by Yang et al. [63]. Consequently, it was assumed that S. nesbitti may infect other species of monkeys such as sooty mangabey (Cercocebus atys) and Guinea baboon (Papio papio) besides the genus Macaca.
A molecular analysis of S. nesbitti was performed years later by sequencing Sarcocystis sp. 18S rDNA derived from muscle samples of a long-tailed macaque [21]. Phylogeny demonstrated that these sequences were closely related to two Sarcocystis spp. that have snake–rodent life cycles, as well as one that was isolated from a racoon in Japan, but with origin in North America [121]. Therefore, it was hypothesized that snakes may be the DHs of S. nesbitti. The first indication of this assumption came from the molecular detection of this species in snakes’ stools, although no oocysts/sporocysts were observed [22]. Phylogenetic analysis using the 18S rDNA region demonstrated that two sequences, one from the monocled cobra (Naja kaouthia) and the other from the reticulated python (Phyton reticulatus), were closely related to S. nesbitti [22]. The following year, a similar approach was implemented, and once again no oocysts/sporocysts were observed, but a sequence from the Indian cobra (Naja naja) was obtained and the isolate was placed near other isolate of S. nesbitti [21], along with two human isolates that were sequenced in this research [23].
Despite molecular evidence, the identification of oocysts/sporocysts in fecal samples is important to diagnose intestinal sarcocystosis [89]. Sporocysts were observed in the stool shed by the amethystine python (Simalia amethistina) in Australia, and presented 18S rDNA sequences with high similarity to S. nesbitti; phylogenetic analysis grouped them closely to S. nesbitti clades, including isolates of humans, monkeys and snakes. This finding confirms the earlier hypothesis of a snake as the DH of S. nesbitti [13]. Previous findings were concentrated in Southeast Asia, and the reticulated python was assumed to be the potential DH for S. nesbitti. Nonetheless, in the same study in which S. nesbitti was probably found in Australia, only S. singaporensis was detected in Indonesia, Malaysia, Thailand, the Philippines and Singapore [13]. Later, a molecular screening using 18S rDNA indicated S. nesbitti in reticulated python in Tioman Island [53]. As the first identified IH of S. nesbitti (macaque) does not occur in Australia, the authors [13] have questioned the snake–monkey cycle based on the diet of its potential predator, classifying this IH as accidental. Thus, they suggest rodents as potential IHs for the parasite. Further experiments associated with molecular analysis using additional markers rather than 18S rDNA are needed to respond to all the gaps involving S. nesbitti.
Regarding the source of S. nesbitti infection in humans in the past outbreaks, food and water were considered potential sources of infection [19,20,106]. Surveillance studies were conducted for the molecular detection of Sarcocystis sp. in water samples; however, the results were negative [20,122]. Molecular evidence of S. nesbitti was found in tank and river water from two villages located in Tioman, Malaysia. Although no oocysts/sporocysts were found, this study suggested that water was the main source of infection [123]. For this reason, it is important to develop preventive policies for both natives and tourists in Southeast Asia, focusing on water safety; it is recommended to drink boiled or bottled water, and to use this same type of water for activities such as brushing teeth, and to avoid uncooked food [20,123].
7. Molecular Diagnosis of Sarcocystis spp. of Snakes
Studies on the identification of sarcocystid organisms have been counting on the valuable contribution of molecular methods to resolve phylogenetic and taxonomic issues. Molecular identification has long been recommended as a mandatory approach for the complete description of a species, not only for the Sarcocystidae family [124,125,126], but also for other taxonomic groups [127]. Among the approximately 20 studies on the description of species of the genus Sarcocystis in snakes, those that were published before the 1990s had no molecular data, which has become an obligatory requirement since this period. Nowadays, the most used markers for the molecular identification of Sarcocystidae of the genus Sarcocystis are nucleotide sequencing nuclear ribosomal loci and loci encoding mitochondrial proteins, located in the mitochondrial genome [38,128,129,130].
Among the Sarcocystis species that infect snakes already described so far, there is information from molecular data for only ten of them, and partial sequences of the gene coding for the small subunit ribosomal RNA (18S rDNA) were provided for all these ten cases. Gene coding for 18S rDNA is a universally used molecular marker that is formed by variable regions interspersed with conserved regions, which allows the design of probes capable of amplifying organisms of different taxonomic levels by PCR. About 1800 nucleotides spanning nine helices, numbered V1 to V9, typically form the 18S coding sequence of Toxoplasma gondii, the most known Sarcocystid. Within 18S, the V4 region is a hypervariable and distinctive region among eukaryotes, rendering V4 the most used segment in sarcocystids [131]. The hypervariable V4 segment is present in 100% of the 18S rDNA sequences from Sarcocystis sp. identified in snakes that are available in Genbank.
Although superior in terms of phylogenetic informativeness for evolutionary studies [126], genes encoding the large subunit ribosomal RNA (28S rDNA) are much less frequently explored in studies on sarcocystids and, as a consequence, as well in studies on sarcocystids of snakes. Larger than 18S rDNA (28S is about 4000 nucleotides long in sarcocystidae), eukaryotic 28S rDNA sequences are also composed of conserved segments interspersed with hypervariable segments [131]. The hypervariable domains of 28S rDNA are characterized by expansions of nucleotide sequences, numbered from D1 to D12, whose origin and function are not elucidated [131]. Typically, the segments usually identified in Sarcocystids are those with the greatest discriminatory power and include the D2 and D3 domains. From the three species of Sarcocystis of snakes that have been 28S rRNA gene sequenced, all of them encompassed the domains D2-D3.
A third marker located within the ribosomal locus used for the identification of species of the Sarcocystidae family are the transcribed internal spacers (ITS). These sequences do not encode known products and their function is unknown. The transcribed internal spacers are interregnum nucleotide sequences between the 18S rDNA and 5.8S ribosomal RNA coding genes, and between 5.8S and 28S rDNA, called ITS1 and ITS2, respectively [132]. Phylogenetic studies using ITS are less common, because it is not possible to use approaches based on structural alignment between homologous sequences, since they are not coding sequences and therefore the alignment of distant taxa is not very accurate [133,134]. However, ITS1 is a marker with the potential to discriminate between species with high similarity and for which differentiation with markers of lower evolutionary rate was not possible [130]. Sarcocystis moreliae, Sarcocystis zuoi, Sarcocystis pantherophisi, Sarcocystis attenuati, Sarcocystis singaporensis and Sarcocystis zamani were identified by both 18S rDNA and ITS1.
Information from non-nuclear genes such as those from the mitochondrial and apicoplast genomes has also been used to identify sarcocystids, but less frequently than the ribosomal locus genes. Among them, studies using cytochrome oxidase coding genes predominate. Cytochrome oxidase is an enzyme that participates in the electron transport chain in eukaryotes and is therefore involved in the cellular energy metabolism. This gene is present in all organisms that have mitochondria and hence is widely used as a marker in molecular phylogenies [127]. Although widely used in DNA barcoding studies, cox1 has insufficient discriminatory power to identify species that use birds as IHs. This group of organisms can only be discriminated using ITS1 analysis, whereas they are almost identical regarding both the 18S rRNA gene and the D2/D3 domain of the 28S rRNA gene [130]. Cox1 has been used to identify three species of snake-derived Sarcocystids: Sarcocystis moreliae, Sarcocystis pantherophisi and Sarcocystis attenuati.
8. Future Directions
For most reports of Sarcocystis spp. in snakes, no molecular data are available. Therefore, besides relevant biological and morphological aspects of the parasites and their animal hosts, it is crucial to provide genetic information for the observed Sarcocystis spp. This will favor the identification of potential hosts involved in their life cycles. When sporocysts or tissue cysts are detected in snakes, PCR and nucleotide sequencing of the most informative markers (ex. ITS1, cox1, 28S rDNA and 18S rDNA) are needed. Then, the sequencing matches may show putative hosts for the parasite.
If the molecular analysis of sporocysts detected in snakes does not provide any information on the potential IH involved in its life cycle, a bioassay may be an alternative approach. Since most reports of snakes acting as DHs use small mammals as IHs, initial attempts at bioassays should focus on frequently found small mammals inhabiting the same region as the snake. Small rodents such as rats and mice are good candidates in bioassays, as they are widely distributed in most geographic regions and laboratory rodents may be acquired for experimental purposes. Searching for lizards as IHs of Sarcocystis sp. from snakes is more challenging, as these animals are rarely raised for research or for human food.
For those snakes that act as IHs for certain species of Sarcocystis, their DHs are completely unknown to date. Future studies should be conducted to generate information about sarcocysts in snakes, including genetic characterization of the parasite. The search for potential DHs for these Sarcocystis spp. is also difficult, as these DHs may include several classes of predators, including mammals, birds and reptiles that may prey on snakes.
Author Contributions
Conceptualization, T.L.B. and L.F.P.G.; writing—original draft preparation, T.L.B.; section about molecular diagnosis of Sarcocystis spp. of snakes, R.M.S.; writing—review and editing, T.L.B., L.F.P.G. and R.M.S.; supervision, L.F.P.G.; funding acquisition, L.F.P.G. All authors have read and agreed to the published version of the manuscript.
Funding
Luís Gondim and Rodrigo Soares are recipients of productivity fellowships from the National Council for Scientific and Technological Development (CNPq). Taynar Bezerra received a scholarship from Fundação de Amparo à Pesquisa do Estado da Bahia (FAPESB).
Acknowledgments
The authors are grateful to the artist Dilermano Maciel for drawing the life cycle illustrated in Figure 1.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dubey, J.P.; Calero-Bernal, B.M.; Rosenthal, B.M.; Speer, C.A.; Fayer, R. Sarcocystis of Animals and Humans; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar] [CrossRef]
- Mugridge, N.B.; Morrison, D.A.; Jakel, T.; Heckeroth, A.R.; Tenter, A.M.; Johnson, A.M. Effects of sequence alignment and structural domains of ribosomal DNA on phylogeny reconstruction for the protozoan family sarcocystidae. Mol. Biol. Evol. 2000, 17, 1842–1853. [Google Scholar] [CrossRef] [PubMed]
- Votypka, J.; Hypsa, V.; Jirku, M.; Flegr, J.; Vavra, J.; Lukes, J. Molecular phylogenetic relatedness of Frenkelia spp. (Protozoa, Apicomplexa) to Sarcocystis falcatula Stiles 1893: Is the genus Sarcocystis paraphyletic? Eukaryot. Microbiol. 1998, 45, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Odening, K. The present state of species-systematics in Sarcocystis Lankester, 1882 (Protista, Sporozoa, Coccidia). Syst. Parasitol. 1998, 41, 209–233. [Google Scholar] [CrossRef]
- Mugridge, N.B.; Morrison, D.A.; Johnson, A.M.; Luton, K.; Dubey, J.P.; Votypka, J.; Tenter, A.M. Phylogenetic relationships of the genus Frenkelia: A review of its history and new knowledge gained from comparison of large subunit ribosomal ribonucleic acid gene sequences. Int. J. Parasitol. 1999, 29, 957–972. [Google Scholar] [CrossRef]
- Speer, C.A.; Dubey, J.P.; Blixt, J.A.; Blagburn, B. Development of Hammondia heydorni in cultured bovine and ovine cells. J. Protozool. 1988, 35, 352–356. [Google Scholar] [CrossRef] [PubMed]
- Riahi, H.; Darde, M.L.; Bouteille, B.; Leboutet, M.J.; Pestre-Alexandre, M. Hammondia hammondi cysts in cell cultures. J. Parasitol. 1995, 81, 821–824. [Google Scholar] [CrossRef]
- Frenkel, J.K.; Smith, D.D. Determination of the genera of cyst-forming coccidia. Parasitol. Res. 2003, 91, 384–389. [Google Scholar] [CrossRef]
- Rzepchzyk, C.M. Evidence of a rat-snake life cycle for Sarcocystis. Int. J. Parasitol. 1974, 4, 447–449. [Google Scholar] [CrossRef]
- Zaman, V.; Colley, F.C. Light and electron microscopic observations of the life cycle of Sarcocystis orientalis sp. n. in the rat (Rattus norvegicus) and the Malaysian reticulated python (Python reticulatus). Z. Parasitenkd. 1975, 47, 169–185. [Google Scholar] [CrossRef]
- Zaman, V.; Colley, F.C. Replacement of Sarcocystis orientalis Zaman and Colley, 1975 by Sarcocystis singaporensis sp.n. Z. Parasitenkd. 1976, 51, 137. [Google Scholar] [CrossRef]
- Beaver, P.C.; Maleckar, J.R. Sarcocystis singaporensis Zaman and Colley, (1975) 1976, Sarcocystis villivillisi sp. n., and Sarcocystis zamani sp. n.: Development, morphology, and persistence in the laboratory rat, Rattus norvegicus. J. Parasitol. 1981, 67, 241–256. [Google Scholar] [CrossRef] [PubMed]
- Wassermann, M.; Raisch, L.; Lyons, J.A.; Natusch, D.J.D.; Richter, S.; Wirth, M.; Preeprem, P.; Khoprasert, Y.; Ginting, S.; Mackenstedt, U.; et al. Examination of Sarcocystis spp. of giant snakes from Australia and Southeast Asia confirms presence of a known pathogen–Sarcocystis nesbitti. PLoS ONE 2017, 12, e0187984. [Google Scholar] [CrossRef]
- Jakel, T.; Burgstaller, H.; Frank, W. Sarcocystis singaporensis: Studies on host specificity, pathogenicity, and potential use as a biocontrol agent of wild rats. J. Parasitol. 1996, 82, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Jäkel, T.; Khoprasert, Y.; Endepols, S.; Archer-Baumann, C.; Suasa-ard, K.; Promkerd, P.; Kliemt, D.; Boonsong, P.; Hongnark, S. Biological control of rodents using Sarcocystis singaporensis. Int. J. Parasitol. 1999, 29, 1321–1330. [Google Scholar] [CrossRef] [PubMed]
- Häfner, U.; Frank, W. Host specificity and host range of the genus Sarcocystis in three snake-rodent life cycles. Zentralblatt Bakteriol. Mikrobiol. Hyg. A 1984, 256, 296–299. [Google Scholar] [CrossRef]
- Munday, B.L.; Mason, R.W. Sarcocystis and related organisms in Australian wildlife: III. Sarcocystis murinotechis sp.n. life cycle in rats (Rattus, Pseudomys and Mastocomys spp.) and tiger snakes (Notechis ater). J. Wildl. Dis. 1980, 16, 83–88. [Google Scholar] [CrossRef]
- Wong, K.T.; Pathmanathan, R. High prevalence of human skeletal muscle sarcocystosis in south-east Asia. Trans. R. Soc. Trop. Med. Hyg. 1992, 86, 631–632. [Google Scholar] [CrossRef]
- Esposito, D.H.; Freedman, D.O.; Neumayr, A.; Parola, P. Ongoing outbreak of an acute muscular Sarcocystis-like illness among travellers returning from Tioman Island, Malaysia, 2011–2012. Eurosurveillance 2012, 17, 20310. [Google Scholar] [CrossRef]
- Italiano, C.M.; Wong, K.T.; AbuBakar, S.; Lau, Y.L.; Ramli, N.; Syed Omar, S.F.; Kahar Bador, M.; Tan, C.T. Sarcocystis nesbitti causes acute, relapsing febrile myositis with a high attack rate: Description of a large outbreak of muscular sarcocystosis in Pangkor Island, Malaysia, 2012. PLoS Negl. Trop. Dis. 2014, 8, e2876. [Google Scholar] [CrossRef]
- Tian, M.; Chen, Y.; Wu, L.; Rosenthal, B.M.; Liu, X.; He, Y.; Dunams, D.B.; Cui, L.; Yang, Z. Phylogenetic analysis of Sarcocystis nesbitti (Coccidia: Sarcocystidae) suggests a snake as its probable definitive host. Vet. Parasitol. 2012, 183, 373–376. [Google Scholar] [CrossRef]
- Lau, Y.L.; Chang, P.Y.; Subramaniam, V.; Ng, Y.H.; Mahmud, R.; Ahmad, A.F.; Fong, M.Y. Genetic assemblage of Sarcocystis spp. in Malaysian snakes. Parasites Vectors 2013, 6, 257. [Google Scholar] [CrossRef] [PubMed]
- Lau, Y.L.; Chang, P.Y.; Tan, C.T.; Fong, M.Y.; Mahmud, R.; Wong, K.T. Sarcocystis nesbitti infection in human skeletal muscle: Possible transmission from snakes. Am. J. Trop. Med. Hyg. 2014, 90, 361–364. [Google Scholar] [CrossRef] [PubMed]
- White, J.; Persson, H. Snakes. In Human Toxicology; Descotes, J., Ed.; Elsevier: Amsterdam, The Netherlands, 1996; p. 839. [Google Scholar]
- Enzeroth, R.; Chobotar, B.; Scholtyseck, E. Sarcocystis crotali sp. n. with the Mojave Rattlesnake (Crotalus scutulatus scutulatus)—Mouse (Mus musculus) cycle. Arch. Protistenkd. 1985, 129, 19–23. [Google Scholar] [CrossRef]
- de Oliveira Toledo, F.A.; Alves, P.V.; Arthuso Vasconcelos, I.M.; Oliveira, A.R.; Dos Santos, D.O.; Gomes Cabral, J.A.; Righi de Toledo, R.A.; Pinto, H.A.; Henriques Cunha, P.H.; Oliveira Paes Leme, F.; et al. Parasitologic and pathologic study of free-ranging south american rattlesnakes. J. Zoo Wildl. Med. 2022, 53, 515–527. [Google Scholar] [CrossRef] [PubMed]
- McAllister, C.T.; Upton, S.J.; Trauth, S.E.; Dixon, J.R. Coccidian parasites (Apicomplexa) from snakes in the southcentral and southwestern United States: New host and geographic records. J. Parasitol. 1995, 81, 63–68. [Google Scholar] [CrossRef]
- Souza, J.L.; Barbosa, A.a.S.; Vazon, A.P.; Uchôa, C.M.; Nunes, B.C.; Cortez, M.B.; Silva, V.L.; Más, L.B.; Melgarejo, A.R.; Bastos, O.M. Parasitological and immunological diagnoses from feces of captive-bred snakes at Vital Brazil Institute. Rev. Bras. Parasitol. Vet. 2014, 23, 123–128. [Google Scholar] [CrossRef]
- McAllister, C.T.; Upton, S.J.; Barker, D.G.; Painter, C.W. Sarcocystis sp.(Apicomplexa) from the New Mexico ridgenose rattlesnake, Crotalus willardi obscurus (Serpentes: Viperidae) from Sonora, Mexico. J. Helminthol. Soc. Wash. 1996, 63, 128–130. [Google Scholar]
- Matuschka, F.R.; Mehlhorn, H.; Abd-Al-Aal, Z. Replacement of Besnoitia Matuschka and Häfner 1984 by Sarcocystis hoarensis. Parasitol. Res. 1987, 74, 94–96. [Google Scholar] [CrossRef]
- Häfner, U.; Matuschka, F.-R. Life cycle studies on Sarcocystis dirumpens sp. n. with regard to host specificity. Z. Parasitenknd. 1984, 70, 715–720. [Google Scholar] [CrossRef]
- Matuschka, F.-R.; Häfner, U. Cyclic transmission of an African Besnoitia species by snakes of the genus Bitis to several rodents. Z. Parasitenknd. 1984, 70, 471–476. [Google Scholar] [CrossRef]
- Slapeta, J.R.; Modry, D.; Koudela, B. Sarcocystis atheridis sp. nov., a new sarcosporidian coccidium from Nitsche’s bush viper, Atheris nitschei Tornier, 1902, from Uganda. Parasitol. Res. 1999, 85, 758–764. [Google Scholar] [CrossRef]
- Jäkel, T. Cyclic transmission of Sarcocystis gerbilliechis n. sp. by the Arabian saw-scaled viper, Echis coloratus, to rodents of the subfamily gerbillinae. J. Parasitol. 1995, 81, 626–631. [Google Scholar] [CrossRef] [PubMed]
- Paperna, I.; Finkelman, S. Early generation merogonies of Sarcocystis muriviperae in liver and muscles of white mice. Folia Parasitol. 1996, 43, 91–99. [Google Scholar]
- Paperna, I.; Finkelman, S. Ultrastructural study of Sarcocystis muriviperae development in the intestine of its snake hosts. Folia Parasitol. 1996, 43, 13–19. [Google Scholar]
- Matuschka, F.R.; Heydorn, A.O.; Mehlhorn, H.; Abd-Al-Aal, Z.; Diesing, L.; Biehler, A. Experimental transmission of Sarcocystis muriviperae n. sp. to laboratory mice by sporocysts from the Palestinian viper (Vipera palaestinae): A light and electron microscope study. Parasitol. Res. 1987, 73, 33–40. [Google Scholar] [CrossRef]
- Verma, S.K.; Lindsay, D.S.; Mowery, J.D.; Rosenthal, B.M.; Dubey, J.P. Sarcocystis pantherophisi n. sp., from Eastern Rat Snakes (Pantherophis alleghaniensis) as Definitive Hosts and Interferon Gamma Gene Knockout Mice as Experimental Intermediate Hosts. J. Parasitol. 2017, 103, 547–554. [Google Scholar] [CrossRef]
- Hu, J.; Sun, J.; Guo, Y.; Zeng, H.; Zhang, Y.; Tao, J. Infection of the Asian gray shrew Crocidura attenuata (Insectivora: Soricidae) with Sarcocystis attenuati n. sp. (Apicomplexa: Sarcocystidae) in China. Parasites Vectors 2022, 15, 13. [Google Scholar] [CrossRef]
- Thornton, S.L. Snakes. In Encyclopedia of Toxicology, 3rd ed.; Wexler, P., Ed.; Academic Press/Elsevier: London, UK, 2014; Volume 4, p. 5220. [Google Scholar]
- Hedley, J.; Eatwell, K. Nonvenomous colubrid snakes (Colubridae). In Companion Animal Care Welfare: The UFAW Companion Animal Handbook, 1st ed.; Yeates, J., Ed.; Universities Federation for Animal Welfare: Hertfordshire, UK, 2018; pp. 412–424. [Google Scholar]
- Bledsoe, B. Sarcocystis idahoensis sp. n. in deer mice Peromyscus maniculatus (Wagner) and gopher snakes Pituophis melanoleucus (Daudin). J. Protozool. 1980, 27, 93–102. [Google Scholar] [CrossRef]
- Bledsoe, B. Transmission studies with Sarcocystis idahoensis of deer mice (Peromyscus maniculatus) and gopher snakes (Pituophis melanoleucus). J. Wildl. Dis. 1980, 16, 195–200. [Google Scholar] [CrossRef]
- Matuschka, F.R. Reptiles as intermediate and/or final hosts of Sarcosporidia. Parasitol. Res. 1987, 73, 22–32. [Google Scholar] [CrossRef]
- Hu, J.J.; Meng, Y.; Guo, Y.M.; Liao, J.Y.; Song, J.L. Completion of the life cycle of Sarcocystis zuoi, a parasite from the Norway rat, Rattus norvegicus. J. Parasitol. 2012, 98, 550–553. [Google Scholar] [CrossRef]
- Matuschka, F.-R. Sarcocystis clethrionomyelaphis n. sp. from snakes of the genus Elaphe and different voles of the family Arvicolidae. J. Parasitol. 1986, 72, 226–231. [Google Scholar] [CrossRef]
- Hu, J.J.; Liu, T.T.; Liu, Q.; Esch, G.W.; Chen, J.Q. Sarcocystis clethrionomyelaphis Matuschka, 1986 (Apicomplexa: Sarcocystidae) infecting the large oriental vole Eothenomys miletus (Thomas) (Cricetidae: Microtinae) and its phylogenetic relationships with other species of Sarcocystis Lankester, 1882. Syst. Parasitol. 2015, 91, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Matuschka, F.-R. Life Cycle of Sarcocystis between Poikilothermic Hosts. Lizards are Intermediate Hosts for S. podarcicolubris sp. nov, Snakes Function as Definitive Hosts. Z. Naturforsch. C 1981, 36, 1093–1096. [Google Scholar] [CrossRef]
- Matuschka, F.-R. Sarcocystis chalcidicolubris n. sp.: Recognition of the life cycle in skinks of the genus Chalcides and snakes of the genus Coluber. J. Parasitol. 1987, 73, 1014–1018. [Google Scholar] [CrossRef]
- Modry, D.; Koudela, B.; Slapeta, J.R. Sarcocystis stenodactylicolubris n. sp., a new sarcosporidian coccidium with a snake-gecko heteroxenous life cycle. Parasite 2000, 7, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Matuschka, F.-R. Experimental investigations on the host range of Sarcocystis podarcicolubris. Int. J. Parasitol. 1985, 15, 77–80. [Google Scholar] [CrossRef]
- Volf, J.; Modrý, D.; Koudela, B.; Šlapeta, J.R. Discovery of the life cycle of Sarcocystis lacertae Babudieri, 1932 (Apicomplexa: Sarcocystidae), with a species redescription. Folia Parasitol. 2013, 46, 257–262. [Google Scholar]
- Mohd Fadil, N.F.; Tengku-Idris, T.I.N.; Shahari, S.; Fong, M.Y.; Lau, Y.L. Molecular Evidence of Sarcocystis Species Infecting Reptiles in Peninsular Malaysia. Iran. J. Parasitol. 2019, 14, 623–630. [Google Scholar] [CrossRef]
- Grismer, L.L.; Quah, E.S.H. An updated and annotated checklist of the lizards of Peninsular Malaysia, Singapore, and their adjacent archipelagos. Zootaxa 2019, 4545, 230–248. [Google Scholar] [CrossRef]
- Abdel-Ghaffar, F.; Rahman Bashtar, A.; Ashour, M.B.; Sakran, T. Life cycle of Sarcocystis gongyli Trinci 1911 in the skink Chalcides ocellatus ocellatus and the snake Spalerosophis diadema: A light and electron microscopic study. Parasitol. Res. 1990, 76, 444–450. [Google Scholar] [CrossRef]
- Morsy, K.; Bashtar, A.R.; Abdel-Ghaffar, F.; Mehlhorn, H.; Al Quraishy, S.; Al-Ghamdi, A.; Koura, E.; Maher, S. Sarcocystis acanthocolubri sp. n. infecting three lizard species of the genus Acanthodactylus and the problem of host specificity. Light and electron microscopic study. Parasitol. Res. 2012, 110, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Lainson, R.; Paperna, I. The life-cycle and ultrastructure of Sarcocystis ameivamastigodryasi n. sp., in the lizard Ameiva ameiva (Teiidae) and the snake Mastigodryas bifossatus (Colubridae). Parasite 2000, 7, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.J.; Ma, T.C.; Li, X.R. A new species of Sarcocystis (Sporozoea, Eucoccidiida) from Rattus norvegicus. Acta Zootaxon. Sin. 2005, 30, 287–290. [Google Scholar]
- Šlapeta, J.R.; Modrý, D.; Votýpka, J.; Jirků, M.; Lukeš, J.; Koudela, B. Evolutionary relationships among cyst-forming coccidia Sarcocystis spp.(Alveolata: Apicomplexa: Coccidea) in endemic African tree vipers and perspective for evolution of heteroxenous life cycle. Mol. Phylogenet. Evol. 2003, 27, 464–475. [Google Scholar] [CrossRef]
- Mehlhorn, H.; Matuschka, F.-R. Ultrastructural studies of the development of Sarcocystis clethrionomyelaphis within its final and intermediate hosts. Protistologica 1986, 22, 97–104. [Google Scholar]
- Brehm, H.; Frank, W. Der Entwicklungskreislauf von Sarcocystis singaporensis Zaman und Colley, 1976 im End-und Zwischenwirt. Z. Parasitenkd. 1980, 62, 15–30. [Google Scholar] [CrossRef]
- Mandour, A.M. Sarcocystis nesbitti n. sp. from the rhesus monkey. J. Protozool. 1969, 16, 353–354. [Google Scholar] [CrossRef]
- Yang, Z.Q.; Wei, C.G.; Zen, J.S.; Song, J.L.; Zuo, Y.X.; He, Y.S.; Zhang, H.F.; Attwood, S.W.; Chen, X.W.; Yang, G.C.; et al. A taxonomic re-appraisal of Sarcocystis nesbitti (Protozoa: Sarcocystidae) from the monkey Macaca fascicularis in Yunnan, PR China. Parasitol. Int. 2005, 54, 75–81. [Google Scholar] [CrossRef]
- Hoare, C.A. Studies on some new ophidian and avian coccidia from Uganda, with a revision of the classification of the Eimeriidea. Parasitology 1933, 25, 359–388. [Google Scholar] [CrossRef]
- Häfner, U.; Frank, W. Morphological studies on the muscle cysts of Sarcocystis dirumpens (Hoare 1933) Häfner and Matuschka 1984 in several host species revealing endopolygeny in metrocytes. Z. Parasitenknd. 1986, 72, 453–461. [Google Scholar] [CrossRef]
- Babudieri, B. I sarcosporidi e le sarcosporidiosi. Arch. Protistenknd. 1932, 76, 421–580. [Google Scholar]
- Slapeta, J.R.; Modry, D.; Votypka, J.; Jirku, M.; Koudela, B.; Lukes, J. Multiple origin of the dihomoxenous life cycle in sarcosporidia. Int. J. Parasitol. 2001, 31, 413–417. [Google Scholar] [CrossRef]
- Trinci, G. Nota sopra una Sarcocystis parassita di Gongylus ocellatus Wagl., con considerazioni critiche sulla morfologia e sulla biologia dei Sarcosporidi. Monit. Zool. Ital. 1911, 22, 309–326. [Google Scholar]
- Matuschka, F.-R.; Mehlhorn, H. Sarcosysts of Sarcocystis podarcicolubris from experimentally infected Tyrrhenian wall ligards (Podarcis tiliguerta), S. gallotiae from naturally infected Canarian lizards (Gallotia galloti) and S. dugesii from Madeirian lizards (Lacerta dugesii). Protistologica 1984, 20, 133–139. [Google Scholar]
- Grego, K.F.; Gardiner, C.H.; Catão-Dias, J.L. Comparative pathology of parasitic infections in free-ranging and captive pit vipers (Bothrops jararaca). Vet. Rec. 2004, 154, 559–562. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.K.; Lindsay, D.S.; Rosenthal, B.M.; Dubey, J.P. Ancient, globally distributed lineage of Sarcocystis from sporocysts of the Eastern rat snake (Pantherophis alleghaniensis) and its relation to neurological sequalae in intermediate hosts. Parasitol. Res. 2016, 115, 2697–2704. [Google Scholar] [CrossRef]
- Bezerra, T.L.; De Jesus, R.F.; Soares, R.M.; Borges-Silva; Gondim, L.F.P. Sarcocystis sp. shed by the common boa snake (Boa constrictor) in Brazil. Int. J. Parasitol. Parasites Wildl. 2023, 22, 84–91. [Google Scholar] [CrossRef]
- Canova, V.; Helman, E.; Robles, M.D.R.; Abba, A.M.; Moré, G. First report of Sarcocystis spp. (Apicomplexa, Sarcocystidae) in Lagostomus maximus (Desmarest, 1917) (Rodentia, Chinchillidae) in Argentina. Int. J. Parasitol. Parasites Wildl. 2023, 20, 180–186. [Google Scholar] [CrossRef]
- Abe, N.; Matsubara, K.; Tamukai, K.; Miwa, Y.; Takami, K. Molecular evidence of Sarcocystis species in captive snakes in Japan. Parasitol. Res. 2015, 114, 3175–3179. [Google Scholar] [CrossRef]
- Wuster, W.; Ferguson, J.E.; Quijada-Mascarenas, J.A.; Pook, C.E.; Salomao Mda, G.; Thorpe, R.S. Tracing an invasion: Landbridges, refugia, and the phylogeography of the Neotropical rattlesnake (Serpentes: Viperidae: Crotalus durissus). Mol. Ecol. 2005, 14, 1095–1108. [Google Scholar] [CrossRef] [PubMed]
- Wiseman, K.D.; Greene, H.W.; Koo, M.S.; Long, D.J. Feeding ecology of a generalist predator, the California kingsnake (Lampropeltis californiae): Why rare prey matter. Herpetol. Conserv. Biol. 2019, 14, 1–30. [Google Scholar]
- Motta-Junior, J.C.; Granzinolli, M.A.M.; Monteiro, A.R. Miscellaneous ecological notes on Brazilian birds of prey and owls. Biota Neotrop. 2010, 10, 255–259. [Google Scholar] [CrossRef]
- Tome, B.; Maia, J.P.; Harris, D.J. Hepatozoon infection prevalence in four snake genera: Influence of diet, prey parasitemia levels, or parasite type? J. Parasitol. 2012, 98, 913–917. [Google Scholar] [CrossRef]
- Tiegs, O.W. Note on the occurrence of Sarcocystis in muscle of python. Parasitology 1931, 23, 412–414. [Google Scholar] [CrossRef]
- Parenzan, P. Sarcosporidiosi (psorospermosi) da nuova specie (Prot.: Sarcocystis atractaspidis n. sp.) in reptile (Atractaspis). Boll. Soc. Nat. Napoli 1947, 55, 117–119. [Google Scholar]
- Maca, O.; Gonzalez-Solis, D. Sarcocystis moreliae sp. nov. in the imported green tree python Morelia cf. viridis (Reptilia, Pythonidae). Front. Vet. Sci. 2023, 10, 1211522. [Google Scholar] [CrossRef]
- Roberts, J.F.; Wellehan, J.F., Jr.; Weisman, J.L.; Rush, M.; Childress, A.L.; Lindsay, D.S. Massive Muscular Infection by a Sarcocystis Species in a South American Rattlesnake (Crotalus durissus terrificus). J. Parasitol. 2015, 101, 386–389. [Google Scholar] [CrossRef]
- Tome, B.; Maia, J.P.; Harris, D.J. Molecular assessment of apicomplexan parasites in the snake Psammophis from North Africa: Do multiple parasite lineages reflect the final vertebrate host diet? J. Parasitol. 2013, 99, 883–887. [Google Scholar] [CrossRef]
- Dolezel, D.; Koudela, B.; Jirku, M.; Hypsa, V.; Obornik, M.; Votypka, J.; Modry, D.; Slapeta, J.R.; Lukes, J. Phylogenetic analysis of Sarcocystis spp. of mammals and reptiles supports the coevolution of Sarcocystis spp. with their final hosts. Int. J. Parasitol. 1999, 29, 795–798. [Google Scholar] [CrossRef]
- Tenter, A.M.; Baverstock, P.R.; Johnson, A.M. Phylogenetic relationships of Sarcocystis species from sheep, goats, cattle and mice based on ribosomal RNA sequences. Int. J. Parasitol. 1992, 22, 503–513. [Google Scholar] [CrossRef]
- Maia, J.P.; Harris, D.J.; Carranza, S.; Gomez-Diaz, E. Assessing the diversity, host-specificity and infection patterns of apicomplexan parasites in reptiles from Oman, Arabia. Parasitology 2016, 143, 1730–1747. [Google Scholar] [CrossRef] [PubMed]
- Pizzatto, L.; Marques, O.; Facure, K. Food habits of Brazilian boid snakes: Overview and new data, with special reference to Corallus hortulanus. Amphib. -Reptil. 2009, 30, 533–544. [Google Scholar] [CrossRef]
- Pough, F.H.; Groves, J.D. Specializations of the body form and food habits of snakes. Am. Zool. 1983, 23, 443–454. [Google Scholar] [CrossRef]
- Fayer, R.; Esposito, D.H.; Dubey, J.P. Human infections with Sarcocystis species. Clin. Microbiol. Rev. 2015, 28, 295–311. [Google Scholar] [CrossRef]
- Fischer, S.; Odening, K. Characterization of bovine Sarcocystis species by analysis of their 18S ribosomal DNA sequences. J. Parasitol. 1998, 84, 50–54. [Google Scholar] [CrossRef]
- Jeffries, A.C.; Schnitizler, B.; Heydorn, A.O.; Johnson, A.M.; Tenter, A.M. Identification of synapomorphic characteris in the genus Sarcocystis based on 18S rDNA sequence comparison. J. Eukaryot. Microbiol. 1997, 44, 388–392. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.E.; Reeder, D.M. Mammal Species of the World: A Taxonomic and Geographic Reference, 3rd ed.; JHU Press: Baltimore, MD, USA, 2005; Volume 1, p. 2142. [Google Scholar]
- Slapeta, J.R.; Kyselová, I.; Richardson, A.O.; Modrý, D.; Lukes, J. Phylogeny and sequence variability of the Sarcocystis singaporensis Zaman and Colley, (1975) 1976 ssrDNA. Parasitol. Res. 2002, 88, 810–815. [Google Scholar] [CrossRef] [PubMed]
- Jäkel, T.; Promkerd, P.; Sitthirath, R.; Guedant, P.; Khoprasert, Y. Biocontrol of rats in an urban environment in Southeast Asia using Sarcocystis singaporensis. Pest. Manag. Sci. 2019, 75, 2148–2157. [Google Scholar] [CrossRef]
- Luo, Z.X.; Chen, W.; Gao, W. Fauna sinica mammalia. In Rodentia Part III: Cricetidae; Science Press: Beijing, China, 2000; Volume 6. [Google Scholar]
- Hu, J.J.; Liu, Q.; Yang, Y.F.; Esch, G.W.; Guo, Y.M.; Zou, F.C. Sarcocystis eothenomysi n. sp. (Apicomplexa: Sarcocystidae) from the large oriental vole Eothenomys miletus (Thomas) (Cricetidae: Microtinae) from Anning, China. Syst. Parasitol. 2014, 89, 73–81. [Google Scholar] [CrossRef]
- Ortega Perez, P.; Wibbelt, G.; Brinkmann, A.; Galindo Puentes, J.A.; Tuh, F.Y.Y.; Lakim, M.B.; Nitsche, A.; Wells, K.; Jakel, T. Description of Sarcocystis scandentiborneensis sp. nov. from treeshrews (Tupaia minor, T. tana) in northern Borneo with annotations on the utility of COI and 18S rDNA sequences for species delineation. Int. J. Parasitol. Parasites Wildl. 2020, 12, 220–231. [Google Scholar] [CrossRef]
- Munday, B.L.; Hartley, W.J.; Harrigan, K.E.; Presidente, P.J.; Obendorf, D.L. Sarcocystis and related organisms in Australian wildlife: II. Survey findings in birds, reptiles, amphibians and fish. J. Wildl. Dis. 1979, 15, 57–73. [Google Scholar] [CrossRef]
- Lühe, M. Ergebnisse der Neueren Sporozoenforschung: Zusammenfassende Darstellung mit Besonderer Berücksichtigung der Malariaparasiten und Ihrer Nächsten Verwandten; Verlag von Gustav Fischer: Jena, Germany, 1900; Volume 28. [Google Scholar]
- Weber, A. Recherches sur la sarcosporidie du Gecko (Sarcocystis platydactyli Bertram). Arch. d’Anat. Microsp. 1910, 11, 167–178. [Google Scholar]
- Weber, A. Sur la morphologie de la Sarcosporidie du Gecko (Sarcocystis platydactyli Bertram). CR Soc. Biol. 1909, 66, 1061–1062. [Google Scholar]
- Chatton, E.; Avel, M. Sur la sarcosporidie du Gecko et ses cytophanères: La paroi et le réticulum sarcosporidiens appartiennent en propre au parasite. C. R. Sci. Soc. Biol. 1923, 89, 181–185. [Google Scholar]
- Dupouy, J.; Kechemir, N. Les cestodes de reptiles en Algerie. Essai de revision du genre Oochoristica Lühe (Cestoda, Anoplocephalidae). Bull. Soc. Hist. Nat. Afr. Nord 1973, 64, 47–98. [Google Scholar]
- Bertram, A. Beiträge zur Kenntniss der Sarcosporidien, nebst einem Anhange über parasitische Schläuche in der Leibshöhle von Rotatorien. Zool. Jahrb. Abt. Anat. Ontog. Tiere 1892, 5, 581–604. [Google Scholar]
- Rosenthal, B.M. Zoonotic Sarcocystis. Res. Vet. Sci. 2021, 136, 151–157. [Google Scholar] [CrossRef]
- Arness, M.K.; Brown, J.D.; Dubey, J.P.; Neafie, R.C.; Granstrom, D.E. An outbreak of acute eosinophilic myositis attributed to human Sarcocystis parasitism. Am. J. Trop. Med. Hyg. 1999, 61, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.; Nathoo, N.; Lu, J.Q.; Kowalewska-Grochowska, K.T.; Power, C. Sarcocystis myopathy in a patient with HIV-AIDS. J. Neurovirol. 2018, 24, 376–378. [Google Scholar] [CrossRef]
- Kwok, C.Y.; Ting, Y. Atypical Presentation of Human Acute Muscular Sarcocystosis: Sarcocystis Nesbitti Confirmed on Molecular Testing. Am. J. Case Rep. 2019, 20, 499–502. [Google Scholar] [CrossRef] [PubMed]
- Abubakar, S.; Teoh, B.T.; Sam, S.S.; Chang, L.Y.; Johari, J.; Hooi, P.S.; Lakhbeer-Singh, H.K.; Italiano, C.M.; Omar, S.F.; Wong, K.T.; et al. Outbreak of human infection with Sarcocystis nesbitti, Malaysia, 2012. Emerg. Infect. Dis. 2013, 19, 1989–1991. [Google Scholar] [CrossRef]
- Thomas, V.; Dissanaike, A.S. Antibodies to Sarcocystis in Malaysians. Trans. R. Soc. Trop. Med. Hyg. 1978, 72, 303–306. [Google Scholar] [CrossRef] [PubMed]
- Markus, M.B. Antibodies to Sarcocystis in human sera. Trans. R. Soc. Trop. Med. Hyg. 1979, 73, 346–347. [Google Scholar] [CrossRef] [PubMed]
- Esposito, D.H.; Stich, A.; Epelboin, L.; Malvy, D.; Han, P.V.; Bottieau, E.; da Silva, A.; Zanger, P.; Slesak, G.; van Genderen, P.J.; et al. Acute muscular sarcocystosis: An international investigation among ill travelers returning from Tioman Island, Malaysia, 2011–2012. Clin. Infect. Dis. 2014, 59, 1401–1410. [Google Scholar] [CrossRef] [PubMed]
- Fayer, R. Sarcocystis spp. in human infections. Clin. Microbiol. Rev. 2004, 17, 894–902. [Google Scholar] [CrossRef]
- Levine, N.D. Protozoan Parasites of Domestic Animals and of Man; Andesite Press: Minneapolis, MN, USA, 1973; p. 406. [Google Scholar]
- Mehlhorn, H.; Heydorn, A.O.; Janitschke, K. Light and electron microscopical study on sarcocysts from muscles of the rhesus monkey (Macaca mulatta), Baboon (Papio cynocephalus) and Tamarin (Saguinus(=Oedipomidas) oedipus). Z. Parasitenkd. 1977, 51, 165–178. [Google Scholar] [CrossRef]
- Mehlhorn, H.; Heydorn, A.O. The sarcosporidia (Protozoa, Sporozoa): Life cycle and fine structure. Adv. Parasitol. 1978, 16, 43–91. [Google Scholar] [CrossRef]
- Karr, S.L., Jr.; Wong, M.M. A survey of Sarcocystis in nonhuman primates. Lab. Anim. Sci. 1975, 25, 641–645. [Google Scholar]
- Kan, S.P.; Prathap, K.; Dissanaike, A.S. Light and electron microstructure of a Sarcocystis sp. from the Malaysian long-tailed monkey, Macaca fascicularis. Am. J. Trop. Med. Hyg. 1979, 28, 634–642. [Google Scholar] [CrossRef]
- Wong, K.T.; Pathmanathan, R. Ultrastructure of the human skeletal muscle sarcocyst. J. Parasitol. 1994, 80, 327–330. [Google Scholar] [CrossRef] [PubMed]
- Pamphlett, R.; O’Donoghue, P. Sarcocystis infection of human muscle. Aust. N. Z. J. Med. 1990, 20, 705–707. [Google Scholar] [CrossRef]
- Jinnai, M.; Kawabuchi-Kurata, T.; Tsuji, M.; Nakajima, R.; Fujisawa, K.; Nagata, S.; Koide, H.; Matoba, Y.; Asakawa, M.; Takahashi, K.; et al. Molecular evidence for the presence of new Babesia species in feral raccoons (Procyon lotor) in Hokkaido, Japan. Vet. Parasitol. 2009, 162, 241–247. [Google Scholar] [CrossRef]
- Husna Maizura, A.M.; Khebir, V.; Chong, C.K.; Azman Shah, A.M.; Azri, A.; Lokman Hakim, S. Surveillance for sarcocystosis in Tioman Island, Malaysia. Malays. J. Public Health Med. 2012, 12, 39–44. [Google Scholar]
- Shahari, S.; Tengku-Idris, T.I.; Fong, M.Y.; Lau, Y.L. Molecular evidence of Sarcocystis nesbitti in water samples of Tioman Island, Malaysia. Parasites Vectors 2016, 9, 598. [Google Scholar] [CrossRef]
- Olsen, G.J. Phylogenetic analysis using ribosomal RNA. Methods Enzymol. 1988, 164, 793–812. [Google Scholar] [CrossRef]
- Tenter, A.M.; Johnson, A.M. Phylogeny of the tissue cyst-forming coccidia. Adv. Parasitol. 1997, 39, 69–139. [Google Scholar] [CrossRef]
- Mugridge, N.B.; Morrison, D.A.; Heckeroth, A.R.; Johnson, A.M.; Tenter, A.M. Phylogenetic analysis based on full-length large subunit ribosomal RNA gene sequence comparison reveals that Neospora caninum is more closely related to Hammondia heydorni than to Toxoplasma gondii. Int. J. Parasitol. 1999, 29, 1545–1556. [Google Scholar] [CrossRef]
- Hebert, P.D.; Cywinska, A.; Ball, S.L.; deWaard, J.R. Biological identifications through DNA barcodes. Proc. Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef]
- Prakas, P.; Kutkiene, L.; Butkauskas, D.; Sruoga, A.; Zalakevicius, M. Description of Sarcocystis lari sp. n. (Apicomplexa: Sarcocystidae) from the great black-backed gull, Larus marinus (Charadriiformes: Laridae), on the basis of cyst morphology and molecular data. Folia Parasitol. 2014, 61, 11–17. [Google Scholar] [CrossRef]
- Lindsay, D.S.; Verma, S.K.; Scott, D.; Dubey, J.P.; von Dohlen, A.R. Isolation, molecular characterization, and in vitro schizogonic development of Sarcocystis sp. ex Accipiter cooperii from a naturally infected Cooper’s hawk (Accipiter cooperii). Parasitol. Int. 2017, 66, 106–111. [Google Scholar] [CrossRef]
- Gjerde, B.; Vikoren, T.; Hamnes, I.S. Molecular identification of Sarcocystis halieti n. sp., Sarcocystis lari and Sarcocystis truncata in the intestine of a white-tailed sea eagle (Haliaeetus albicilla) in Norway. Int. J. Parasitol. Parasites Wildl. 2018, 7, 1–11. [Google Scholar] [CrossRef]
- Gagnon, S.; Bourbeau, D.; Levesque, R.C. Secondary structures and features of the 18S, 5.8S and 26S ribosomal RNAs from the Apicomplexan parasite Toxoplasma gondii. Gene 1996, 173, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Hillis, D.M.; Dixon, M.T. Ribosomal DNA: Molecular evolution and phylogenetic inference. Q. Rev. Biol. 1991, 66, 411–453. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J.T.; Morrison, D.A.; Liddell, S.; Jenkins, M.C.; Mohammed, O.B.; Ryce, C.; Dubey, J.P. The genus Hammondia is paraphyletic. Parasitology 1999, 118 Pt 4, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Prakas, P.; Kutkiene, L.; Butkauskas, D.; Sruoga, A.; Zalakevicius, M. Molecular and morphological investigations of Sarcocystis corvusi sp. nov. from the jackdaw (Corvus monedula). Parasitol. Res. 2013, 112, 1163–1167. [Google Scholar] [CrossRef]
Figure 1.
Life cycle of Sarcocystis spp. involving snakes as definitive hosts. Their intermediate hosts encompass distinct species of small mammals and lizards.
Figure 1.
Life cycle of Sarcocystis spp. involving snakes as definitive hosts. Their intermediate hosts encompass distinct species of small mammals and lizards.


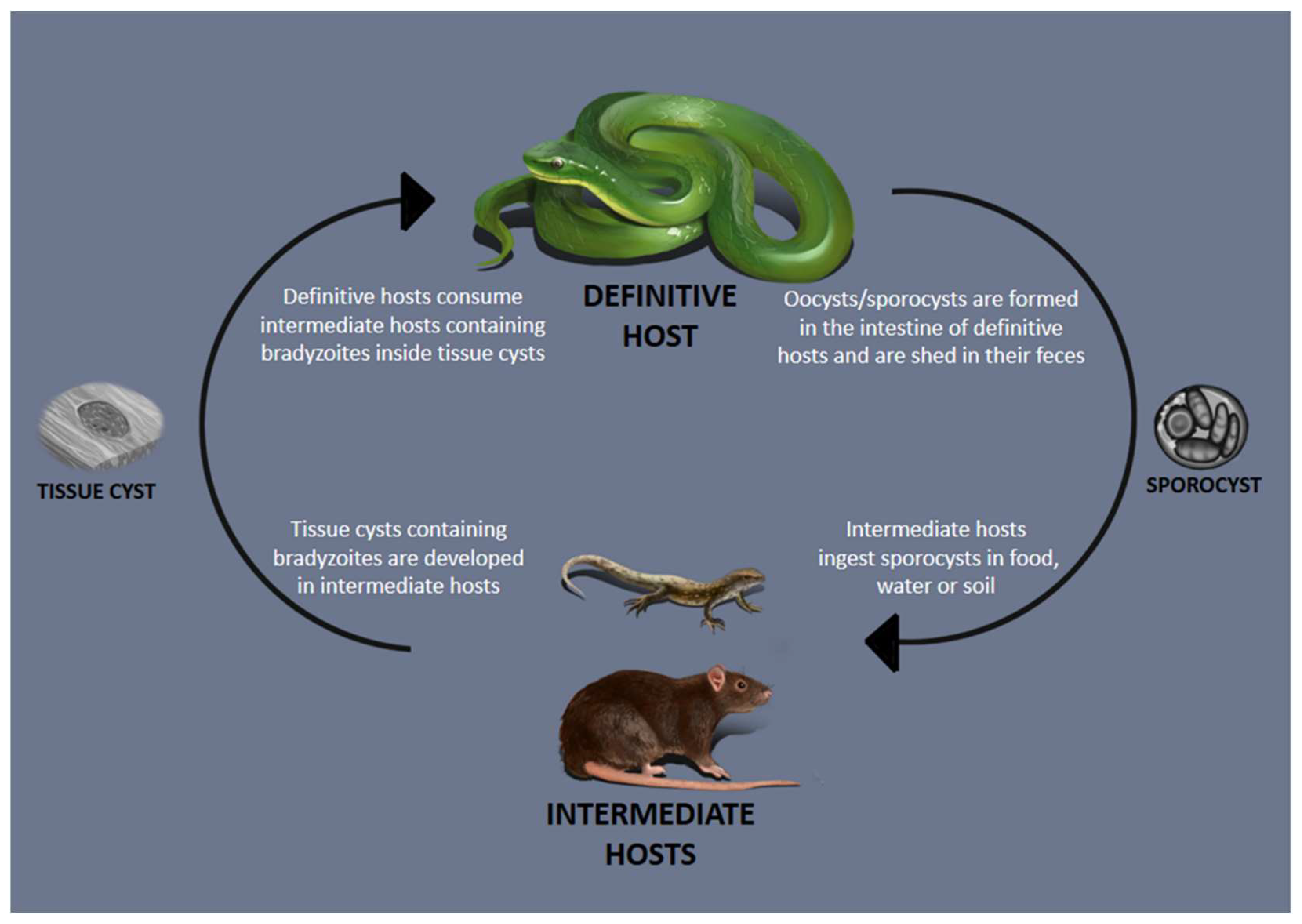{kind=link}
Table 1.
Snakes as definitive hosts of Sarcocystis spp. and mammals as their intermediate hosts.
| Sarcocystis Species | Snake Species (Family) | Intermediate Host | Sarcocyst Description/Animal Tissue | Place of Sampling | Molecular Marker | Reference |
|---|---|---|---|---|---|---|
| S. attenuati | Elaphe taeniura (Colubridae) | Crocidura attenuata a | 740–1355 × 117–250 μm, thick wall (3.3–4.5 μm), type 9h, protrusions 3.3–4.5 µm, GS 1.2–1.6 μm, bradyzoite 8.2–10.4 × 2.0–3.0 μm/esophagus, diaphragm, tongue, skeletal muscles (thigh, loin, rump, and ribs) and heart | Yunnan Province, China a | 18S rDNA, Cox1, Cox3, ITS1, cytb, rpoB, clpC | [39] |
| S. pantherophisi | Pantherophis alleghaniensis a (Colubridae) | KO Mus musculus | Up to 3500 × 70 μm, smooth thin wall (<1 μm), protrusions 100 × 100 nm, GS 500 nm, bradyzoite 7.1–8.4 × 1.5–2.9 μm/tongue, abdomen, limbs, and masseter muscles. | Maryland, U.S. a | 18S rDNA, 28S rDNA, Cox1, ITS1 | [38] |
| S. zuoi | Elaphe carinata (Colubridae) | Rattus norvegicus a | 2325–700 × 225–81 μm, striated wall, type 17, protrusion 10.2–12.5 × 2.0–2.5 μm, GS 0.4–0.6 μm, bradyzoite 7–10 × 2.6–3.0 μm/skeletal muscle through [54] the body, tongue, esophagus, and diaphragm | China a | 18S rDNA, ITS1, ITS2 | [45,58] |
| S. atheridis | Atheris nitschei a (Viperidae) | Mus musculus Lemniscomys barbarus | 3000 × 0.9 µm, smooth thin wall (0.6–0.8 µm), type 1b, bradyzoite 6–8 × 1–1.5 µm, protrusions up to 150 × 90 nm, GS 0.6 ± 0.8 µm/tongue, triceps brachii, m. quadriceps femoris, abdominal wall, labium superior and inferior, masseter, diaphragm (one specimen) | Uganda, Africa a | 18S rDNA | [33,59] |
| S. gerbilliechis | Echis coloratus a (Viperidae) | Gerbillus gerbillus Gerbillus perpallidus Psammomys obesus Pachyuromys duprasi natronensi | Up to 11.7 mm–190 µm, smooth, thin wall, type 1b, protrusions 80–180 × 50–120 nm, GS 300–1500 nm/esophagus, tongue, diaphragm, and thoracic and abdominal wall muscles | Galala mountain, Egypt a | No data | [34] |
| S. muriviperae | Vipera palaestinae a (Viperidae) Coluber jugularis (Colubridae) | Mus musculus | 5–8 mm × 150–400 µm, thick wall (3 µm), type 18b, bradyzoite 12–13 µm, protrusions ~3.5 µm/skeletal muscles | Tel Aviv, Israel a | No data | [35,36,37] |
| S. clethrionomyelaphis | Zamenis longíssimus a Z. scalaris Elaphe. quatuorlineata E. dione E. taeniura Pantherophis. obsoleta P. guttata (Colubridae) | Microtus arvalis M. oeconomus, M. guentheri Clethrionomys glareolus Eothenomys miletus b | 1200–4500 × 70–170 µm, smooth wall, type 9, protrusions ~3 µm, bradyzoites 10–12 × 2 µm/skeletal muscle (thigh, loin, rump, ribs), tongue, esophagus and diaphragm | Germany a Yunnan Province, China b | 18S rDNA | [46,47,60] |
| S. crotali | Crotalus scuiulatus scutulatus a (Viperidae) | Mus musculus | 4000 × 500 µm, smooth thin wall, type 1a, homogeneous GS (1 µm)/skeletal muscle of abdomen and flanks | California, U.S. a | No data | [25] |
| S. hoarensis | Bittis nasicornis a B. arietans B. caudalis B. gabonica (Viperidae) | Meriones unguiculatus Mesocricetus auratus Phodopus sungorus Gerbillus perpallidus Mastomys natalensis Mus musculus | 2500 µm, secondary cyst wall, type 42, bradyzoite 11–12 × 2 µm/connective tissue (snout, tongue), subcutis and connective tissue (ear, scrotum, anus, base of the tail and soles of the feet) | East Africa a | No data | [30,32] |
| S. zamani | Python reticulatus a (Pythonidae) | Rattus norvegicus | 1000–2000 × 300–500 µm, smooth thick wall (1–3 µm), type 18c, bradyzoite 10–12 × 1.5 µm (living cyst), 7–8 × 1–1.5 µm (formalin-fixed)/skeletal muscle throughout the body, including the tongue, esophagus, vagina, and diaphragm | Singapore a Malaysia a | No data | [12] |
| S. villivillosi | Python reticulatus a P. timorensis, Aspidites melanocephalus Python sebae (Pythonidae) | Rattus norvegicus and others Rattus spp. Bandicota spp. | 700–1100 × 70–90 µm, striated thick wall (<2 µm), type 22, bradyzoite 5.0 × 1.0 µm/skeletal muscles through the body | Singapore a Malaysia a | No data | [12,16] |
| S. idahoensis | Pituophis melanoleucus a (Colubridae) | Peromyscus aniculatus a | Up to 9400 × 800 µm, smooth thin wall (~1 µm), bradyzoite 6–7 × 2.0 µm/tongue, thorax, abdomen and hind limb | Idaho, U.S. a | No data | [42,43] |
| S. murinotechis | Notechis ater a (Elapidae) | Rattus norvegicus, R. lutreolus [57] R. rattus Pseudomys higginsi Mastacomys fuscus | 300 × 60–90 µm, thick wall, protrusion 6 × 4 µm/tongue, diaphragm and skeletal muscle | Southeastern Australia a | No data | [17] |
| S. singaporensis | Python reticulatus a P. timorensis, Aspidites melanocephalus Python sebae | Rattus norvegicus and others Rattus spp. Bandicota spp. Nesokia indicaArvicanthis niloticus | Up to 4400 µm, striated thick wall (3–5 µm), type 19, bradyzoite 4.5–6 × 1.0–1.5 µm/muscles throughout the body, including tongue, neck, and appendages, diaphragm, trunk and skin muscles, striated muscles of the eye, esophagus, scrotum and vagina | Malaysia a Singapore a [6] | 18S rDNA, ITS1, ITS2, 28S rDNA | [10,11,12,14,16,61] |
| S. nesbitti | Python reticulatus c (?) Simalia amethistina d (Pythonidae) Naja kaouthia c (?) (Elapidae) | Macaca mulatta a Macaca fascicularis b | Up to 2000 µm, smooth thin wall (0.46–0.60 µm), type 1, GS 0.5–0.76 µm, bradyzoite 7.5–10 × 1.25 µm/cervix, scapular region, abdomen, psoas, and diaphragm muscle | North India a Malaysia c Indonesia c Australia d China b | 18S rDNA | [13,20,22,62,63] |
| S. dirumpens | Bittis nasicornis a B. arietans B. caudalis B. gabonica (Viperidae) | Meriones unguiculatus Mesocricetus auratus Phodopus sungorus Gerbillus perpallidus Mastomys natalensis Mus musculus Grammomys caniceps Lemniscomys barbarous | Up to 25,000 × 50–100 µm, smooth thin wall, type 1b, GS 0.5–1 µm, bradyzoite 8–9 × 2 µm/abdominal wall | East Africa a | No data | [31,64,65] |
a, b, c, d: Equal letters mean that the naturally infected animals from which Sarcocystis sp. was isolated inhabit the same region where they were captured or sampled. Experimental hosts are not labeled with superscribed letters. Sarcocyst description: the word “type” refers to the 82 revised and/or proposed distinct types of sarcocyst walls [1].
Table 2.
Snakes as definitive hosts of Sarcocystis spp. and lizards as their intermediate hosts.
| Sarcocystis Species | Snake Species (Family) | Intermediate Host | Sarcocyst Description/Tissue | Place of Sampling | Molecular Marker | Reference |
|---|---|---|---|---|---|---|
| S. acanthocolubri | Spalerosophis diadema (Colubridae) | Acanthodactylus boskianus a A. sculentu a A. paradalis a | 10,120 × 950 µm, striated thick wall (3.9 μm), protrusions 2.55 × 1.41 μm, GS 0.37 μm/fore-limb, hind-limb subcutaneous tissue and tail | Giza, Cairo, Baltem, Rashid, Marsa-Matrouh, Beni-suef, Egypt a | No data | [56] |
| S. ameivamastigodryasi | M astigodryas bifossatus (Colubridae) | Ameiva ameiva a | 1000–2000 × 80 µm, smooth thin wall, bradyzoite 7.2 × 2.5 µm/tongue, tail and hind limb | Pará, Brazil a | No data | [57] |
| S. stenodactylicolubris | Coluber najadum a Coluber rogersi (Colubridae) | Ptyodactylus guttatus Stenodactylus grandiceps | 175–200 × 35–50 µm, thick wall (2 µm), type 4, protrusions up to 800 × 200–250 nm, bradyzoite 5–6 × 1.5 µm, GS 200–300 nm/tail, limbs, tongue | Syria a | No data | [50] |
| S. lacertae | Coronella austriaca a (Colubridae) | Podarcis muralis | Up to 640 × 230 µm, striated thick wall (1 µm), type 37a, bradyzoite 6.6 × 2.5 µm, GS 0.5–2.0 µm/tail | Čabraď, Slovak Republic a | 18S rDNA | [52,66,67] |
| S. gongyli | Spalerosophis diadema (Colubridae) | Chalcides ocellatus ocellatus a | 60–100 × 200–900 µm, striated wall (2–6 µm), type 9d, protrusions 1.9–3 µm, bradyzoite 4.1–5.3 × 0.8–2.0 µm/tail, hind limb and different parts of the body | Giza, Egypt a | No data | [55,68] |
| S. chalcidicolubris | Coluber ravergieri C. viridiflavus C. gemonensis C. najadum (Colubridae) | Chalcides spp. a | 124–275 × 27–53 µm, smooth thin wall, type 1e, protrusions 2 × 1 µm, bradyzoite 10–12 × 2 µm/tail | Israel a | No data | [49] |
| S. podarcicolubris | Coluber viridiflavus a C. gemonensis a Balkan whip C. najadum C. jugularis C. ravergieri Macroprotodon cucullatus (Colubridae) | Lacerta spp. Podarcis spp. Algyroides nigropunctatus | 430–1300 × 90–290 µm, striated wall, type 9 m, protrusions up to 5 µm, bradyzoite 7.7–10 × 2 µm/tail’s base and hind limb | Sardina a Sicilya a Jugoslavia a Turkey | No data | [44,48,51,69] |
a: Equal letters mean that the naturally infected animals from which Sarcocystis sp. was isolated inhabit the same region where they were captured or sampled. Experimental hosts are not labeled with superscribed letters. Sarcocyst description: the word “type” refers to the 82 revised and/or proposed distinct types of sarcocyst walls [1].
Table 3.
Snakes as intermediate hosts of Sarcocystis sp. and with unknown definitive hosts.
| Sarcocystis Species | Snake Species | Sarcocyst Description/Tissue | Natural Habitat/Place of Sampling | Molecular Marker | Reference |
|---|---|---|---|---|---|
| Sarcocystis moreliae | Morelia cf. viridis | 1.118 × 123 μm, smooth thin wall (0.55 μm), type I, bradyzoites 663,799 × 1.43–1.87 μm (LM)/head, trunk and tail | Indonesia/Czech Republic | 18S rDNA, 28S rDNA, ITS1, cox1 | [81] |
| Sarcocystis sp. | Crotalus durissus terrificus | up to 2000 × 50–150 µm, thin wall (<1 μm), type I wall, minimal GS, bradyzoite 1–2 μm, (LM; TEM)/Tongue, head, skeletal muscle adjacent to vertebra | South America/Tennessee, USA | 18S rDNA | [82] |
| Sarcocystis atractaspidis | Atractaspis leucomelas | up to 7 × 4 mm | Kenya/Kenya | No data | [80] |
| S. pythonis | Python spilota | up to 1100 µm, thin wall, bradyzoite 4 × 7 μm | No data | [79] |
Sarcocyst description: the word “type” refers to the 82 revised and/or proposed distinct types of sarcocyst walls [1].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bezerra, T.L.; Soares, R.M.; Gondim, L.F.P. Sarcocystis Species (Apicomplexa, Eucoccidiorida) Parasitizing Snakes. Parasitologia 2023, 3, 327-347. https://0-doi-org.brum.beds.ac.uk/10.3390/parasitologia3040032
AMA Style
Bezerra TL, Soares RM, Gondim LFP. Sarcocystis Species (Apicomplexa, Eucoccidiorida) Parasitizing Snakes. Parasitologia. 2023; 3(4):327-347. https://0-doi-org.brum.beds.ac.uk/10.3390/parasitologia3040032
Chicago/Turabian StyleBezerra, Taynar Lima, Rodrigo Martins Soares, and Luís Fernando Pita Gondim. 2023. "Sarcocystis Species (Apicomplexa, Eucoccidiorida) Parasitizing Snakes" Parasitologia 3, no. 4: 327-347. https://0-doi-org.brum.beds.ac.uk/10.3390/parasitologia3040032