
Toxoplasma gondii in Marine Life of Italian Coasts, What Do We Know So Far?


1
Department of Infectious Diseases, Microbiology and Public Health, La Sapienza, University of Rome, 00185 Rome, Italy
2
Department of Translational Research, University of Pisa, 56126 Pisa, Italy
*
Author to whom correspondence should be addressed.
Parasitologia 2023, 3(4), 364-373; https://0-doi-org.brum.beds.ac.uk/10.3390/parasitologia3040035
Submission received: 14 September 2023
/
Revised: 27 November 2023
/
Accepted: 3 December 2023
/
Published: 5 December 2023
{kind=link}
Abstract
:Coastal areas of Italy experience high anthropogenic pressure, with a population density estimated to be 360 people per km2. This is correlated with the production of sewage or surface runoff of water contaminated with Toxoplasma gondii oocysts and other pathogens that can in turn enter the food chain and become a public health concern. Therefore, in the present study, we want to review the current knowledge on the presence of T. gondii in Italian coastal areas. The majority of research in Italy has been carried out on marine mammals, while fishes and mollusks have less often been screened. So far, T. gondii has been found in all sea animals tested, with genotypes II and III as well as atypical genotypes being the most commonly found when screened. However, more information is needed. We encourage routine screening of mollusks and fish that are sold in different food markets throughout the country in order to add information on the distribution of T. gondii and other parasites, preventing possible future outbreaks.
1. Introduction
Toxoplasma gondii is an obligate intracellular protozoan parasite with the ability to infect a broad range of hosts, being one of the most widespread parasites among warm-blooded animals. The seroprevalence ranges from 10 to 30% in North America, Southeast Asia, and Northern Europe; 30 to 50% in central and southern Europe; and up to 90% in South America and tropical Africa [1]. During its life cycle, the parasite goes through three different stages: tachyzoite, the free parasite form infects host cells in the acute stage; bradyzoite, tissue cysts are formed; and oocysts form that are shed in the feces of the definitive host (felids). T. gondii oocysts become infective only after sporulation. This occurs after excretion to the environment in feline feces, often within 2–5 days and depending on temperature, humidity, and oxygen [2,3]. Each oocyst contains two sporocysts with four sporozoites. Under favorable conditions, mature oocysts are very resistant and can remain infective in soil for over a year, while in water they can remain active for up to 2 years [4]. Humans are infected by consuming sporulated oocysts present in unwashed fruits and vegetables, or in water, as well as in raw or undercooked fish and mollusks; they are also infected by eating undercooked meat containing tissue cysts [5,6].
In recent years, special interest has been paid to the spillover of pathogens from terrestrial animals into aquatic ecosystems, resulting in different emerging infectious diseases (EIDs) [7]. Host–parasite ecology is linked to climate change, environmental degradation, and changes in human demography [7]. Therefore, coastal areas are of great importance for EIDs, since they are the end point of streams and rivers, storm drains, and overland freshwater runoff that can carry parasites, viruses, and bacteria from terrestrial sources such as feces from humans, pets, and farmed animals [8,9]. Indeed, the emergence of different diseases in marine plants and animals has already been demonstrated in cases of morbillivirus [10], herpesvirus [11], brucellosis [12], canine distemper virus (CDV) [13], and influenza B virus [14]. The dispersion of encysted protozoan parasites that can be filtered and concentrated by shellfish and potentially ingested by humans and marine mammals has also been documented [15]. These EIDs may have epizootic potential and zoonotic consequences [16].
It is therefore clear that the health of coastal ecosystems is strongly impacted by terrestrially derived pathogen pollution [17]. As an example, a long-spined sea urchin disease likely induced by boat traffic eradicated urchins from the Caribbean, shifting the algal communities. Macroalgae became the dominant benthic group, which inhibited coral recruitment; as a result, coral communities were diminished [18]. On the other hand, marine mammals such as otters, seals, and sea lions have been strongly impacted by different infectious diseases. It has been reported that mortality among southern sea otters on the coasts of California is mainly caused by protozoan infections, with T. gondii and Sarcocystis neurona being the cause of death in about 25% of sea otters examined between 1998 and 2001 [19]. These changes cause serious economic losses in terms of declining fishery revenue and ecosystem damage [20]. Some examples of infectious agents that lead to economic loss include coral diseases in the Caribbean, abalone disease in California, herpes and leptospirosis in California sea lions, lobster disease and salmon virus in Maine, and the presence of protozoans in oysters in Maryland and Texas [21].
Unlike other protozoans such as Cryptosporidium spp. or Giardia duodenalis, Toxoplasma was not considered to be a significant waterborne pathogen. However, different toxoplasmosis outbreaks linked to water consumption increased the concern in the scientific community about the presence of oocysts in the environment. One of the first outbreaks occurred in 1979 among soldiers in Panama, and it was linked to water sources used for drinking [22]. In British Columbia, in 1995, more than 100 cases of human acute T. gondii infection were reported. It was associated with the use of unfiltered and chloraminated surface water provided by the municipal water system, which was contaminated with oocysts from either cougar and/or domestic cat feces [23]. Indeed, oocysts can survive inactivation procedures from water treatment plants. They remain viable for up to 18 months at 4 °C even after exposure to aqueous 2% sulfuric acid, and they are resistant to detergents and disinfectants. According to this, drinking-water treatment plants that use chlorination as the only treatment method could potentially disseminate viable oocysts and/or discharge contaminated water into the ocean [24].
Conducting routine marine monitoring has logistical and financial issues, which often leads to unobserved marine mortality events. In many cases, the specific infectious agent of a disease outbreak has not been identified. This makes it even more important to provide an overview to identify possible trends in ecosystem decline in different parts of the world to try to mitigate possible outbreaks. From a One Health perspective, caring about our ecosystems and what inhabits them is closely connected to our own health. Very little has been done so far in Italy regarding the detection of T. gondii in the marine environment. However, it has been gaining more importance in other parts of the world, especially since devastating consequences of toxoplasmosis for some species such as sea otters became apparent [11]. In the present review, our aim is to gather all existing information on this topic in Italy in order to fill the gaps and enable future research that could shed light on the wellbeing of water, sea ecosystems, and human health.
2. A Closer Look into Italy
During the last 100 years, the European population residing in coastal areas has doubled. The same trend has been seen in Italy, which can offer a good model for the study of the effects of this development on coastal pathogen pollution. According to ISTAT (National Institute of Statistics), the population of Italy in 2022 consisted of almost 60 million people [25], representing the 3rd most populated country in the European Union and the 25th most populated country in the world. Coastal areas occupy 18% of the country’s area and host as much as 34.5% of its inhabitants, leading to a population density estimated to be 360 people per km2 compared to 198 inhabitants per km2 at the national level [26].
Due to its geographic position, Italy has become an important tourist destination. Even though coastal areas correspond to 18% of the Italian surface area, they host as much as 53% of the total tourist attendance in the country. This leads to tourism pressure of 11 tourists per inhabitant in these areas (double that of the rest of the country), representing an important socioeconomic force. However, mass tourism has exerted great pressure on the infrastructure of coastal areas and has led to an increase in waste production with a big impact on the environment. This results in territorial dynamics that have modified the structure of about 20% of coastal areas through land modification. Almost half of Italian coasts are made out of sand, and it is estimated that almost 43% of them are occupied by bathing establishments [26].
It is thus evident that there is significant anthropogenic pressure on the coasts of Italy. In a study of 6477 km of the Italian coast, approximately 51% was urbanized (Legambiente). Specifically, 696 km was colonized by industries, ports, and infrastructure while 918 km was occupied by urban centers. Another 1659 km (25% of the total coastline) was made up of less dense urban landscapes. The remaining 3206 km of the Italian coasts was divided into 764 km (12% of total coastal line) of agricultural land and 2442 km (37% of the total coastline) of natural landscapes. The amount of waste produced in this country is approximately 30 tons per year. It has already been demonstrated that the increase in human population density in coastal areas leads to biodiversity loss, which in turn increases the possibility of zoonotic transmission of pathogens [27,28].
3. Distribution of T. gondii in Italy
In Europe, animal husbandry biosecurity measures and good manufacturing practices in the meat industry have been considerably improved, shifting attention to the role of oocysts in the epidemiology of this parasite. In recent years, long-term observations of pregnant women showed that the infection cannot always be linked to known risk factors such as the consumption of cyst-contaminated meat. Environmental exposure to T. gondii oocysts can indeed be an important route for human infection [29]. More sensitive analytical methods have been developed for the detection of oocysts in soil and water samples, adding evidence on the spread of oocysts via shedding in the environment. The role of oocysts is no longer seen as secondary to tissue-cyst-based transmission.
Half of the game produced in Europe is estimated to be seropositive for T. gondii, and meat from game is identified as an important source of T. gondii infection for consumers, including venison and wild boar meat [30,31]. Moreover, atypical and recombinant genotypes have been detected in wild animals in northern and central Italy [32,33].
Different studies report the seroprevalence of T. gondii in livestock in different Italian regions. A recent study conducted in the north of Italy reports seropositivity in different animals: 42.9% in cats, 29.9% in sheep, 25% in roe deer, 21.8% in dogs, 18.7% in goats, 15.5% in wild boar, and 9.7% in pigs. The same study reports a seroprevalence of 20.4% on a screened population of 36,814 individuals, while, among pregnant women, toxoplasmosis was active in 0.39% of cases [34]. Overall, the seroprevalence among small ruminants in farms of central and northern Italy is 90.7% [35] and 68.4% in cattle farms [36]. In southern and insular regions of Italy, there is a similar epidemiological scenario, with a prevalence of 87% among small ruminants farmed in Sicily [37]. In Campania, a southern region, the prevalence among livestock is reported at 77.8% in sheep [38] and 13.7% in water buffalo [39]. A seroprevalence of 39.6% was found among wild boar in the south [40].
Most cats are infected by ingesting infected prey [2]. Up to one billion oocysts can be shed by a single cat over a 1–2-week period, and they can repeatedly shed oocysts during their lifetime [41]. According to a survey from FEDIAF [42], between 2020 and 2021, the total number of cats owned by Italian households increased by almost 27%, reaching a population of 10.2 billion. Official feline population data and cat censuses are difficult to gather since it is not always recognized as a public health concern. In Italy, cat registration and identification are carried out on a voluntary basis. A multi-center study carried out in 2021 including 987 cats distributed across the country estimated that 27.9% of cats were living in colonies, 69.2% had a private owner, and 2.9% were living in shelters [43]. Among owned cats, it has been shown that the majority are living in rural areas (67.8%), being kept outdoors and often in households with other pets [44]. The same survey estimated that 32% of cats in Italy do not attend a veterinary clinic [44]. However, few studies have been conducted at a national level to determine the T. gondii seroprevalence among domestic cats. In Europe, the prevalence among feral and stray cats ranges between 18.27% and 40.7%, varying with the age and lifestyle of the cat [45]. In Italy, a seroprevalence of 40.7% was estimated among stray cats, being higher in urban (45%) than suburban areas (35.7%) [46].
ISTAT reported a yearly average temperature of 15.6 °C for 2021, with an increase of +0.6 °C compared to the period 1981–2010 and +1 °C compared to the period 1971–2000. In total, the yearly precipitation accounted for 746 mm in 2021, but 2022 presented a very dry year, with 455 mm [47]. In the current year, precipitation has already reached 500 mm, and it is projected to be a rainy fall and winter. Climatic changes with periods of drought followed by intense periods of rain could facilitate the dispersion of oocysts, as previously stated.
4. Toxoplasma gondii: From Land to Sea
Anthropogenic pressure on the coastline increases pollution with sewage or surface runoff of water possibly contaminated with T. gondii oocysts. It has been demonstrated that the distribution of pet cats and feral cat colonies is linked to human settlements that can provide food and shelter, even when feral cats are not directly fed or taken care of [48]. This is probably the key factor in T. gondii epidemiology in marine environments, with feral and domestic cats as the sole source of oocysts [49]. After infection, a single cat can shed millions of oocysts within 1 week, and, as previously reported, they can remain viable in soil for up to one year [3].
Reports of infection among marine mammals are becoming more frequent worldwide [8,50,51,52,53]. Clinical symptoms and seroprevalence have been described in sea otters, seals, sea lions, manatees, walruses, and dolphins [8,50,51,52,53]. It was only in the early 2000s that it was possible to isolate T. gondii from brain and heart tissues of sea otters (Enhydra lutris) in the National Wildlife Health Center (NWHC) of Madison, Wisconsin (USA) [54]. In the waters of the European continent, T. gondii has been found in the common dolphin (Delphinus delphis), striped dolphin (Stenella coeruleoalba), common bottlenose dolphin (Tursiops truncatus), long-finned pilot whale (Globicephala melas), Risso’s dolphin (Grampus griseus), harbor porpoise (Phocoena phocoena), humpback whale (Megaloptera novaengliae), bearded seal (Erignathus barbatus), harbor seal (Phocavitulina), ringed seal (Pusa hispida) and grey seal (Halichoerus grypus) [54].
Novel routes of oocyst transmission involving suspended microplastics have been found [55]. Parasites can associate with microplastics in contaminated seawater, suggesting that they may facilitate pathogen entry into marine food webs. Parasites could therefore be incorporated into aggregates of nanoparticles and ingested by filter-feeding marine invertebrates that may not be able to capture freely moving parasites [55]. Moreover, microplastics have the capacity to float or sink. In the first case, by floating on the sea surface, they can travel large distances, which may facilitate the dispersion of pathogens far from the areas from where they came. In the second case, sinking particles will accumulate in the benthos, where pathogens will concentrate and filter-feeding invertebrates are more likely to ingest them [56].
5. Toxoplasma gondii in the Italian Sea Environment
5.1. Marine Mammals
Toxoplasma gondii constitutes a major zoonotic agent and a significant cause of clinical disease in wildlife such as abortion, pneumonia, or encephalitis. Specifically, this parasite has been associated with neurological disease and encephalitis in cetaceans, becoming a primary neurotropic pathogen for striped dolphins [57,58,59,60]. A recent meta-analysis on marine species affected with T. gondii in Europe shows a total prevalence of 13%. The most affected are marine fissipeds (53.1%), followed by mollusks (26.4%), fishes (21.8%), cetaceans (14.8%), and pinnipeds (2.8%) [61].
The majority of data on the presence of T. gondii in Italian waters is related to stranded marine mammals. The International Pelagos Sanctuary, which occupies circa 90,000 km2 of international waters between France, Italy, and the Principality of Monaco, is a valuable source of information on marine mammals. It was established in 1999 to protect cetaceans from the combined pressures of natural environmental fluctuations and human impacts. It constitutes the first transboundary marine protected area, being a region with a high biomass of diversified plankton that attracts all eight cetaceans present in the Mediterranean Sea. Stranded cetaceans have been registered and analyzed in recent decades in this area, providing information on the presence of T. gondii and other parasites affecting the health of marine mammals.
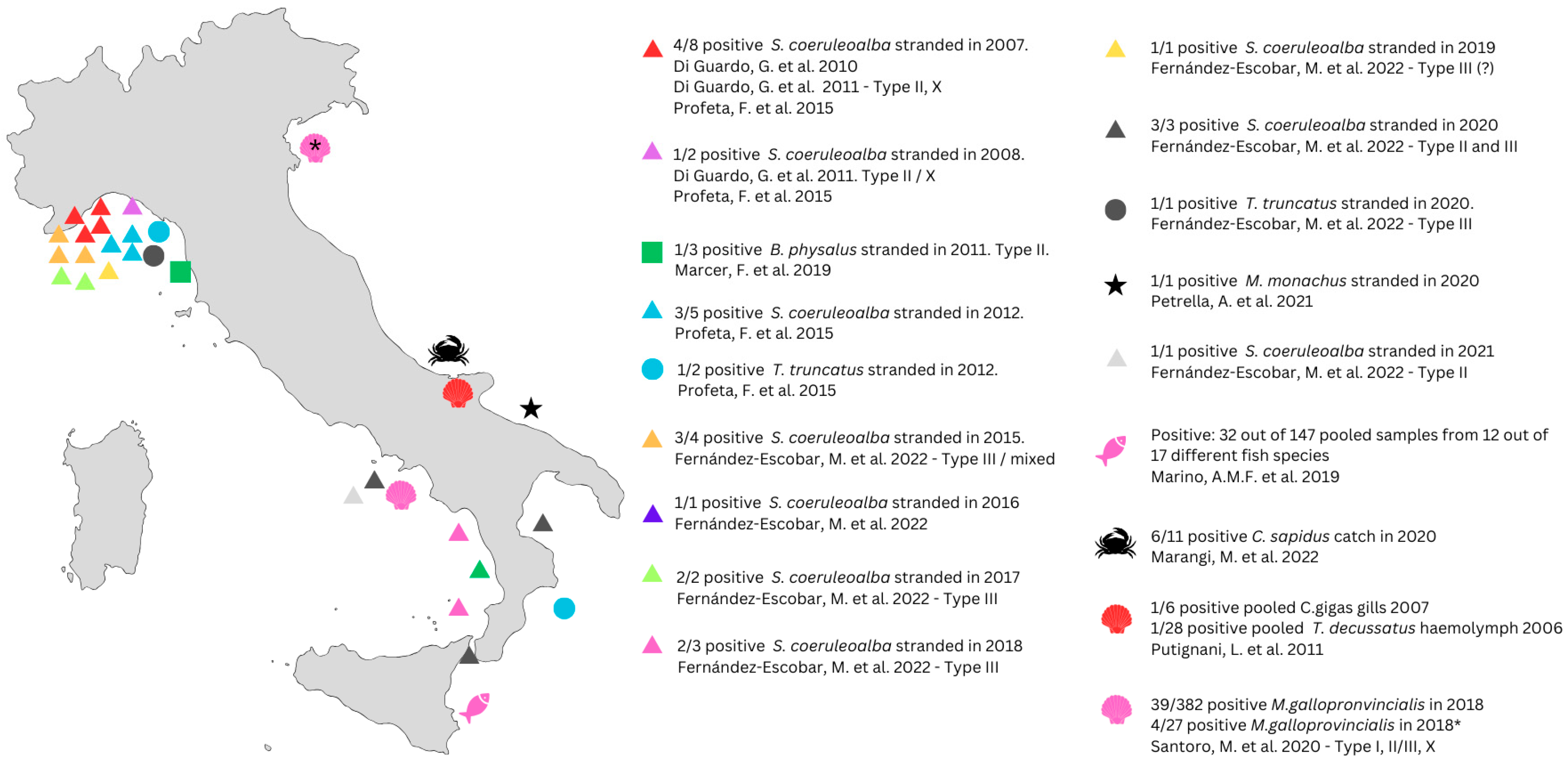
In the present review, we only included reported cases where marine mammals tested positive for T. gondii and organized them according to the year they were found (Figure 1, Supplementary Material S1). Cases considered in this review span from 2007 to 2021, with different tests for detecting the presence of T. gondii being used (more information in Supplementary Material S1). Striped dolphins (S. coeruleoalba) are the most common stranded marine mammals, accounting for 30 total cases, of which 21 (70%) resulted positive for T. gondii. Out of three cases of common bottlenose dolphins (T. truncatus), two tested positive for T. gondii. Out of three Fin whales (B. physalus), only one resulted positive. One rare case of Mediterranean monk seal (M. monachus) was reported in 2020 and resulted positive for T. gondii.
5.2. Fish
In Italy, some fish species, such as anchovies (E. encrasicolus), are consumed raw or following marination in lemon juice for a few hours, which is not enough to inactivate oocysts [62]. Moreover, most of the species consumed in Italy are demersal or benthopelagic, living on different types of seabeds (sand, mud, rocks, or seagrass beds). In this environment, T. gondii oocysts are more likely to settle, sometimes aided by aquatic invertebrates that facilitate settling and help benthos concentration.
Only one study has investigated the presence of T. gondii in fish from local markets, analyzing a total of 1293 individuals from 17 different species, pooled into 147 groups [49]. Samples were obtained by pooling intestines, gills, and skin/muscles. T. gondii DNA was found in 12 of 17 fish species tested with 32 positive samples out of 147 overall [49]. Of these samples, 16 were of the skin/muscle and 11 of intestines and gills [49]. Fish was purchased at MAAS (Sicily Agro-Food Markets), which is the biggest market in Sicily, and in other small-size fisheries that sell fish from the area FAO 37.2.2. According to this study, the species that tested positive for T. gondii accounted for 23.1% of the total caught in Sicily in 2015.
5.3. Shellfish
There have been four studies analyzing shellfish in Italy in the last ten years, one regarding crustaceans and three regarding bivalves. The study on crustaceans collected Atlantic blue crab (Callinectes sapidus) from Lesina Lagoon and analyzed hemolymph, gills, stomach, hepatopancreas, and gonads using PCR. T. gondii was mostly found in the gills (n = 4), hemolymph (n = 2), and stomach (n = 1) [63].
Of the three studies analyzing bivalves in Italy, two focused on Mediterranean mussels (Mytilus galloprovincialis). In the study by Marangi et al., 53 samples from Turkey and 60 from Foggia food markets (Italy) were analyzed using qPCR targeting the B1 gene [64]. While all the samples from Italy were negative, 7 out of 53 (13.2%) mussel DNA samples from Turkey tested positive for T. gondii, and the type I genotype was confirmed [64]. In the study by Santoro et al., they analyzed 382 samples of M. galloprovincialis sampled in seven production sites in the Gulf of Naples and 27 farmed Mediterranean mussels obtained from a mollusk depuration plant in Corigliano Calabro (Calabria region) [65]. Digestive glands were used for the detection of the T. gondii B1 gene using qPCR. T. gondii DNA was detected in 39 out of 382 (10.2%) Mediterranean mussels from 6 out of 7 sampling sites in the Gulf of Naples and 4 out of 27 individuals from the mollusk depuration plant in Corigliano Calabro [65].
Only one study analyzed edible farmed shellfish. The study collected a total of 1734 individuals divided into 62 pooled samples: 109 Crassostrea gigas (6 pools of gills), 660 Mytilus galloprovincialis (22 pools), 804 Tapes decussatus (28 pools), and 161 Tapes philippinarum (6 pools) [66]. T. gondii DNA was detected by both nested PCR and real-time FLAG assay in 2 pooled samples out of 62 (3.2%) [66].
Figure 1.
Summary of all available data on Toxoplasma gondii in Italian marine life from 2007 to 2021. Color represents the year of sampling: red = 2007; purple = 2008; dark green = 2011; light blue = 2012; orange = 2015; dark blue = 2016; light green = 2017; pink = 2018; yellow = 2019; black = 2020; grey = 2021. Different symbols represent the species analyzed: triangle = Stenella coeruleoalba; square = Balaenoptera physalus; circle = Tursiops truncatus; star = Monachus monachus [67,68,69,70,71,72,73]; fish = different species reported in Marino, et al., 2019 [49]; bivalve = Mytilus galloprovincialis [65,66], Crassostrea gigas [66], Thylacodes decussatus [66]; crab = Callinectes sapidus [63]. * The batch of mollusk analyzed from the depuration plant in Corigliano Calabro came from Porto Tolle (Veneto region) and arrived at the depuration plant one month before the analysis.
Figure 1.
Summary of all available data on Toxoplasma gondii in Italian marine life from 2007 to 2021. Color represents the year of sampling: red = 2007; purple = 2008; dark green = 2011; light blue = 2012; orange = 2015; dark blue = 2016; light green = 2017; pink = 2018; yellow = 2019; black = 2020; grey = 2021. Different symbols represent the species analyzed: triangle = Stenella coeruleoalba; square = Balaenoptera physalus; circle = Tursiops truncatus; star = Monachus monachus [67,68,69,70,71,72,73]; fish = different species reported in Marino, et al., 2019 [49]; bivalve = Mytilus galloprovincialis [65,66], Crassostrea gigas [66], Thylacodes decussatus [66]; crab = Callinectes sapidus [63]. * The batch of mollusk analyzed from the depuration plant in Corigliano Calabro came from Porto Tolle (Veneto region) and arrived at the depuration plant one month before the analysis.

6. Concluding Remarks
Toxoplasmosis is still a disconcerting problem for public health, both in Italy and worldwide. Human population growth and cat density associated with it increase the presence of oocysts in the environment. Moreover, changes in weather patterns that include intense rainfall events followed by periods of drought enhance the mobilization of oocysts by rainwater that accumulates during dry periods [74]. Therefore, the risk of oocyst-borne infections among humans and animals could increase in the following decades. An increasing number of studies are raising concern about parasite dispersion in the marine environment worldwide. However, Italy still needs more information in this regard. Very few studies have been conducted regarding the sea environment, particularly among species that are consumed by the population.
A recent report linked the consumption of raw shellfish to an increased risk of T. gondii infection in humans [75], which is relevant for Italy since people consume different mollusk and fish species raw. Therefore, routine screening of mollusks that are sold in different food markets throughout the country could add important information on the distribution of T. gondii and other parasites, providing the basis for a prevention strategy, which could preclude possible future outbreaks. Moreover, a collaborative and multidisciplinary effort among researchers in the areas of the biology, epidemiology, and ecology of T. gondii in the environment could shed more light on the distribution of this parasite. Furthermore, T. gondii oocyst detection methods are not standardized, leading to inadequate monitoring of water, soil, and foods. A lack of standardization and habitual use of different approaches also complicates study comparison. Consensus among laboratories would facilitate accurate detection of the parasite in the environment, preventing possible future toxoplasmosis outbreaks.
We therefore give a call to pursue new research activities in this sector.
Supplementary Materials
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Morozińska-Gogol, J. The presence of Toxoplasma gondii in the terrestrial and marine environments and its importance for public health. Ann. Parasitol. 2021, 67, 137–149. [Google Scholar] [PubMed]
- Dubey, J.P. Toxoplasmosis of Animals and Humans, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2009. [Google Scholar]
- Lélu, M.; Villena, I.; Dardé, M.-L.; Aubert, D.; Geers, R.; Dupuis, E.; Marnef, F.; Poulle, M.-L.; Gotteland, C.; Dumètre, A.; et al. Quantitative Estimation of the Viability of Toxoplasma gondii Oocysts in Soil. Appl. Environ. Microbiol. 2012, 78, 5127–5132. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P. Toxoplasma gondii oocyst survival under defined temperatures. J. Parasitol. 1998, 84, 862–865. [Google Scholar] [CrossRef] [PubMed]
- Pereira, K.S.; Franco, R.M.B.; Leal, D.A.G. Chapter 1—Transmission of Toxoplasmosis (Toxoplasma gondii) by Foods. In Advances in Food and Nutrition Research; Taylor, S.L., Ed.; Academic Press: Cambridge, MA, USA, 2010; pp. 1–19. Available online: https://0-www-sciencedirect-com.brum.beds.ac.uk/science/article/pii/S1043452610600010 (accessed on 8 August 2023).
- Robert-Gangneux, F.; Dardé, M.-L. Epidemiology of and Diagnostic Strategies for Toxoplasmosis. Clin. Microbiol. Rev. 2012, 25, 264–296. [Google Scholar] [CrossRef]
- Daszak, P.; Cunningham, A.A.; Hyatt, A.D. Anthropogenic environmental change and the emergence of infectious diseases in wildlife. Acta Trop. 2001, 78, 103–116. [Google Scholar] [CrossRef]
- Baker, R.E.; Mahmud, A.S.; Miller, I.F.; Rajeev, M.; Rasambainarivo, F.; Rice, B.L.; Takahashi, S.; Tatem, A.J.; Wagner, C.E.; Wang, L.-F.; et al. Infectious disease in an era of global change. Nat. Rev. Microbiol. 2022, 20, 193–205. [Google Scholar] [CrossRef]
- Shapiro, K. Climate and coastal habitat change: A recipe for a dirtier ocean. Mar. Pollut. Bull. 2012, 64, 1079–1080. [Google Scholar] [CrossRef]
- Di Guardo, G.; Mazzariol, S. Cetacean morbillivirus: A Land-to-Sea Journey and Back? Virol. Sin. 2019, 34, 240–242. [Google Scholar] [CrossRef]
- Gulland, F.M.D.; Hall, A.J.; Ylitalo, G.M.; Colegrove, K.M.; Norris, T.; Duignan, P.J.; Halaska, B.; Whitehouse, K.A.; Lowenstine, L.J.; Deming, A.C.; et al. Persistent Contaminants and Herpesvirus OtHV1 Are Positively Associated With Cancer in Wild California Sea Lions (Zalophus californianus). Front. Mar. Sci. 2020, 7, 1093. Available online: https://www.frontiersin.org/articles/10.3389/fmars.2020.602565 (accessed on 4 October 2023). [CrossRef]
- Nymo, I.H.; Tryland, M.; Godfroid, J. A review of Brucella infection in marine mammals, with special emphasis on Brucella pinnipedialis in the hooded seal (Cystophora cristata). Vet. Res. 2011, 42, 93. [Google Scholar] [CrossRef]
- Thomas, N.; White, C.L.; Saliki, J.; Schuler, K.; Lynch, D.; Nielsen, O.; Dubey, J.P.; Knowles, S. Canine distemper virus in the sea otter (Ehydra lutris) population in Washington Estate, USA. J. Wildl. Dis. 2020, 56, 873–883. [Google Scholar] [CrossRef] [PubMed]
- Osterhaus, A.D.; Rimmelzwaan, G.F.; Martina, B.E.; Bestebroer, T.M.; Fouchier, R.A. Influenza B virus in seals. Science 2000, 288, 1051–1053. [Google Scholar] [CrossRef]
- Fayer, R.; Dubey, J.P.; Lindsay, D.S. Zoonotic protozoa: From land to sea. Trends Parasitol. 2004, 20, 531–536. [Google Scholar] [CrossRef]
- Bossart, G.D. Marine Mammals as Sentinel Species for Oceans and Human Health. Vet. Pathol. 2011, 48, 676–690. [Google Scholar] [CrossRef] [PubMed]
- Doney, S.C. The growing human footprint on coastal and open-ocean biogeochemistry. Science 2010, 328, 1512–1516. [Google Scholar] [CrossRef] [PubMed]
- Hylkema, A.; Kitson-Walters, K.; Kramer, P.R.; Patterson, J.T.; Roth, L.; Sevier, M.L.B.; Vega-Rodriguez, M.; Warham, M.M.; Williams, S.M.; Lang, J.C. The 2022 Diadema antillarum die-off event: Comparisons with the 1983–1984 mass mortality. Front. Mar. Sci. 2023, 9, 2654. Available online: https://www.frontiersin.org/articles/10.3389/fmars.2022.1067449 (accessed on 8 August 2023). [CrossRef]
- Kreuder, C.; Miller, M.A.; Jessup, D.A.; Lowenstine, L.J.; Harris, M.D.; Ames, J.A.; Carpenter, T.E.; Conrad, P.A.; Mazet, J.A.K. Patterns of mortality in southern sea otters (Enhydra lutris nereis) from 1998–2001. J. Wildl. Dis. 2003, 39, 495–509. [Google Scholar] [CrossRef]
- Halpern, B.S.; Walbridge, S.; Selkoe, K.A.; Kappel, C.V.; Micheli, F.; D′Agrosa, C.; Bruno, J.F.; Casey, K.S.; Ebert, C.; Fox, H.E.; et al. A global map of human impact on marine ecosystems. Science 2008, 319, 948–952. [Google Scholar] [CrossRef]
- Harvell, D.; Aronson, R.; Baron, N.; Connell, J.; Dobson, A.; Ellner, S.; Gerber, L.; Kim, K.; Kuris, A.; McCallum, H.; et al. The rising tide of ocean diseases: Unsolved problems and research priorities. Front. Ecol. Environ. 2004, 2, 375–382. [Google Scholar] [CrossRef]
- Benenson, M.W.; Takafuji, E.T.; Lemon, S.M.; Greenup, R.L.; Sulzer, A.J. Oocyst-transmitted toxoplasmosis associated with ingestion of contaminated water. N. Engl. J. Med. 1982, 307, 666–669. [Google Scholar] [CrossRef]
- Aramini, J.J.; Stephen, C.; Dubey, J.P.; Engelstoft, C.; Schwantje, H.; Ribble, C.S. Potential contamination of drinking water with Toxoplasma gondii oocysts. Epidemiol. Infect. 1999, 122, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, K.; Bahia-Oliveira, L.; Dixon, B.; Dumètre, A.; de Wit, L.A.; VanWormer, E.; Villena, I. Environmental transmission of Toxoplasma gondii: Oocysts in water, soil and food. Food Waterborne Parasitol. 2019, 15, e00049. [Google Scholar] [CrossRef] [PubMed]
- Simone, M. La Dinamica Demografica del 2022; Istituto Nazionale di Statistica (ISTAT): Rome, Italy, Comunicato Stampa. Available online: https://www.istat.it/it/archivio/282316 (accessed on 20 October 2023).
- Mirto, A.P.M. Attività Antropiche e Salute delle Coste. Indicatori Territoriali di Rischio e Sostenibilità per Aree Costiere e Insulari in Italia e nell’Unione Europea; Istituto Nazionale di Statistica (ISTAT): Rome, Italy, 2022; ISBN 978-88-458-2084-7. Available online: https://www.istat.it/it/archivio/274891 (accessed on 20 October 2023).
- Keesing, F.; Ostfeld, R.S. Impacts of biodiversity and biodiversity loss on zoonotic diseases. Proc. Natl. Acad. Sci. USA 2021, 118, e2023540118. [Google Scholar] [CrossRef] [PubMed]
- Wilson, A.G.; Wilson, S.; Alavi, N.; Lapen, D.R. Human density is associated with the increased prevalence of a generalist zoonotic parasite in mammalian wildlife. Proc. R. Soc. B Biol. Sci. 2021, 288, 20211724. [Google Scholar] [CrossRef] [PubMed]
- Conrad, P.; Miller, M.; Kreuder, C.; James, E.; Mazet, J.; Dabritz, H.; Jessup, D.; Gulland, F.; Grigg, M. Transmission of Toxoplasma: Clues from the study of sea otters as sentinels of Toxoplasma gondii flow into the marine environment. Int. J. Parasitol. 2005, 35, 1155–1168. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Biological Hazards (BIOHAZ). Scientific Opinion on the public health hazards to be covered by inspection of meat from farmed game. EFSA J. 2013, 11, 3264. [Google Scholar] [CrossRef]
- World Health Organization. WHO Estimates of the Global Burden of Foodborne Diseases: Foodborne Disease Burden Epidemiology Reference Group 2007–2015. World Health Organization: Geneva, Switzerland, 2015. Available online: https://apps.who.int/iris/handle/10665/199350 (accessed on 20 October 2023).
- Battisti, E.; Zanet, S.; Trisciuoglio, A.; Bruno, S.; Ferroglio, E. Circulating genotypes of Toxoplasma gondii in Northwestern Italy. Vet. Parasitol. 2018, 253, 43–47. [Google Scholar] [CrossRef]
- Rocchigiani, G.; Nardoni, S.; D′Ascenzi, C.; Nicoloso, S.; Picciolli, F.; Papini, R.A.; Mancianti, F. Seroprevalence of Toxoplasma gondii and Neospora caninum in red deer from Central Italy. Ann. Agric. Environ. Med. 2016, 23, 699–701. [Google Scholar] [CrossRef]
- Dini, F.; Morselli, S.; Marangoni, A.; Taddei, R.; Maioli, G.; Roncarati, G.; Balboni, A.; Dondi, F.; Lunetta, F.; Galuppi, R. Spread of Toxoplasma gondii among animals and humans in Northern Italy: A retrospective analysis in a One-Health framework. Food Waterborne Parasitol. 2023, 32, e00197. [Google Scholar] [CrossRef]
- Gazzonis, A.L.; Zanzani, S.A.; Villa, L.; Manfredi, M.T. Toxoplasma gondii infection in meat-producing small ruminants: Meat juice serology and genotyping. Parasitol. Int. 2020, 76, 102060. [Google Scholar] [CrossRef]
- Gazzonis, A.L.; Marino, A.M.F.; Garippa, G.; Rossi, L.; Mignone, W.; Dini, V.; Giunta, R.P.; Luini, M.; Villa, L.; Zanzani, S.A.; et al. Toxoplasma gondii seroprevalence in beef cattle raised in Italy: A multicenter study. Parasitol. Res. 2020, 119, 3893–3898. [Google Scholar] [CrossRef] [PubMed]
- Vesco, G.; Buffolano, W.; La Chiusa, S.; Mancuso, G.; Caracappa, S.; Chianca, A.; Villari, S.; Currò, V.; Liga, F.; Petersen, E. Toxoplasma gondii infections in sheep in Sicily, southern Italy. Vet. Parasitol. 2007, 146, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Fusco, G.; Rinaldi, L.; Guarino, A.; Proroga, Y.T.R.; Pesce, A.; Giuseppina, D.M.; Cringoli, G. Toxoplasma gondii in sheep from the Campania region (Italy). Vet. Parasitol. 2007, 149, 271–274. [Google Scholar] [CrossRef] [PubMed]
- Ciuca, L.; Borriello, G.; Bosco, A.; D’Andrea, L.; Cringoli, G.; Ciaramella, P.; Maurelli, M.P.; Di Loria, A.; Rinaldi, L.; Guccione, J. Seroprevalence and Clinical Outcomes of Neospora caninum, Toxoplasma gondii and Besnoitia besnoiti Infections in Water Buffaloes (Bubalus bubalis). Animals 2020, 10, 532. [Google Scholar] [CrossRef] [PubMed]
- Sgroi, G.; Viscardi, M.; Santoro, M.; Borriello, G.; D′Alessio, N.; Boccia, F.; Pacifico, L.; Fioretti, A.; Veneziano, V.; Fusco, G. Genotyping of Toxoplasma gondii in wild boar (Sus scrofa) in southern Italy: Epidemiological survey and associated risk for consumers. Zoonoses Public Health 2020, 67, 805–813. [Google Scholar] [CrossRef]
- Dubey, J.P.; Miller, N.L.; Frenkel, J.K. The Toxoplasma gondii oocysts from cat feces. J. Exp. Med. 1970, 132, 636–662. [Google Scholar] [CrossRef] [PubMed]
- FEDIAF. European Pet Food, Annual Report 2023. Available online: www.europeanpetfood.org (accessed on 15 November 2023).
- Genchi, M.; Vismarra, A.; Zanet, S.; Morelli, S.; Galuppi, R.; Cringoli, G.; Lia, R.; Diaferia, M.; di Regalbono, A.F.; Venegoni, G.; et al. Prevalence and risk factors associated with cat parasites in Italy: A multicenter study. Parasites Vectors 2021, 14, 475. [Google Scholar] [CrossRef]
- Carvelli, A.; Iacoponi, F.; Scaramozzino, P. A Cross-Sectional Survey to Estimate the Cat Population and Ownership Profiles in a Semirural Area of Central Italy. BioMed Res. Int. 2016, 2016, e3796872. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P. Feline toxoplasmosis and coccidiosis: A survey of domiciled and stray cats. J. Am. Vet. Med. Assoc. 1973, 162, 873–877. [Google Scholar]
- Papini, R.; Sbrana, C.; Rosa, B.; Saturni, A.M.; Sorrentino, A.M.; Cerretani, M.; Raffaelli, G.; Guidi, G. Serological survey of Toxoplasma gondii infections in stray cats from Italy. Rev. Med. Vet. 2006, 157, 193–196. [Google Scholar]
- Vignani, D.; Budano, F. Istat, Istituto Nazionale di Statistica. Temperatura e Precipitazione delle Città Capoluogo Negli Anni 1971–2021. Report, 17 May 2023. Available online: www.istat.it (accessed on 20 October 2023).
- Horn, J.A.; Mateus-Pinilla, N.; Warner, R.E.; Heske, E.J. Home range, habitat use, and activity patterns of free-roaming domestic cats. J. Wildl. Manag. 2011, 75, 1177–1185. [Google Scholar] [CrossRef]
- Marino, A.M.F.; Giunta, R.P.; Salvaggio, A.; Castello, A.; Alfonzetti, T.; Barbagallo, A.; Aparo, A.; Scalzo, F.; Reale, S.; Buffolano, W.; et al. Toxoplasma gondii in edible fishes captured in the Mediterranean basin. Zoonoses Public Health 2019, 66, 826–834. [Google Scholar] [CrossRef] [PubMed]
- Cabezón, O.; Hall, A.; Vincent, C.; Pabón, M.; García-Bocanegra, I.; Dubey, J.; Almería, S. Seroprevalence of Toxoplasma gondii in North-eastern Atlantic harbor seal (Phoca vitulina vitulina) and grey seal (Halichoerus grypus). Vet. Parasitol. 2011, 179, 253–256. [Google Scholar] [CrossRef] [PubMed]
- Michael, S.A.; Howe, L.; Chilvers, B.L.; Morel, P.; Roe, W.D. Seroprevalence of Toxoplasma gondii in mainland and sub-Antarctic New Zealand sea lion (Phocarctos hookeri) populations. N. Z. Vet. J. 2016, 64, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Terracciano, G.; Fichi, G.; Comentale, A.; Ricci, E.; Mancusi, C.; Perrucci, S. Dolphins Stranded along the Tuscan Coastline (Central Italy) of the “Pelagos Sanctuary”: A Parasitological Investigation. Pathogens 2020, 9, 612. [Google Scholar] [CrossRef]
- VanWormer, E.; Carpenter, T.E.; Singh, P.; Shapiro, K.; Wallender, W.W.; Conrad, P.A.; Largier, J.L.; Maneta, M.P.; Mazet, J.A.K. Coastal development and precipitation drive pathogen flow from land to sea: Evidence from a Toxoplasma gondii and felid host system. Sci. Rep. 2016, 6, 29252. [Google Scholar] [CrossRef]
- Cole, R.A.; Lindsay, D.S.; Howe, D.K.; Roderick, C.L.; Dubey, J.P.; Thomas, N.J.; Baeten, L.A. Biological and Molecular Characterizations of Toxoplasma gondii Strains Obtained from Southern Sea Otters (Enhydra lutris nereis). J. Parasitol. 2000, 86, 526–530. [Google Scholar] [CrossRef]
- Zhang, E.; Kim, M.; Rueda, L.; Rochman, C.; VanWormer, E.; Moore, J.; Shapiro, K. Association of zoonotic protozoan parasites with microplastics in seawater and implications for human and wildlife health. Sci. Rep. 2022, 12, 6532. [Google Scholar] [CrossRef]
- De-la-Torre, G.E. Microplastics: An emerging threat to food security and human health. J. Food Sci. Technol. 2020, 57, 1601–1608. [Google Scholar] [CrossRef]
- Di Guardo, G.; Mazzariol, S. Toxoplasma gondii: Clues from stranded dolphins. Vet. Pathol. 2013, 50, 737. [Google Scholar] [CrossRef]
- Dubey, J.P.; Murata, F.H.A.; Cerqueira-Cézar, C.K.; Kwok, O.C.H.; Grigg, M.E. Recent epidemiologic and clinical importance of Toxoplasma gondii infections in marine mammals: 2009–2020. Vet. Parasitol. 2020, 288, 109296. [Google Scholar] [CrossRef] [PubMed]
- Giorda, F.; Crociara, P.; Iulini, B.; Gazzuola, P.; Favole, A.; Goria, M.; Serracca, L.; Dondo, A.; Crescio, M.I.; Audino, T.; et al. Neuropathological Characterization of Dolphin Morbillivirus Infection in Cetaceans Stranded in Italy. Animals 2022, 12, 452. [Google Scholar] [CrossRef] [PubMed]
- Pintore, M.D.; Mignone, W.; Di Guardo, G.; Mazzariol, S.; Ballardini, M.; Florio, C.L.; Goria, M.; Romano, A.; Caracappa, S.; Giorda, F.; et al. Neuropathologic findings in cetaceans stranded in Italy (2002–2014). J. Wildl. Dis. 2018, 54, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Ahmadpour, E.; Rahimi, M.T.; Ghojoghi, A.; Rezaei, F.; Hatam-Nahavandi, K.; Oliveira, S.M.R.; Pereira, M.d.L.; Majidiani, H.; Siyadatpanah, A.; Elhamirad, S.; et al. Toxoplasma gondii Infection in Marine Animal Species, as a Potential Source of Food Contamination: A Systematic Review and Meta-Analysis. Acta Parasit. 2022, 67, 592–605. [Google Scholar] [CrossRef]
- Dumètre, A.; Dardé, M.L. How to detect Toxoplasma gondii oocysts in environmental samples? FEMS Microbiol. Rev. 2003, 27, 651–661. [Google Scholar] [CrossRef]
- Marangi, M.; Lago, N.; Mancinelli, G.; Antonio, O.L.; Scirocco, T.; Sinigaglia, M.; Specchiulli, A.; Cilenti, L. Occurrence of the protozoan parasites Toxoplasma gondii and Cyclospora cayetanensis in the invasive Atlantic blue crab Callinectes sapidus from the Lesina Lagoon (SE Italy). Mar. Pollut. Bull. 2022, 176, 113428. [Google Scholar] [CrossRef]
- Marangi, M.; Giangaspero, A.; Lacasella, V.; Lonigro, A.; Gasser, R.B. Multiplex PCR for the detection and quantification of zoonotic taxa of Giardia, Cryptosporidium and Toxoplasma in wastewater and mussels. Mol. Cell. Probes 2015, 29, 122–125. [Google Scholar] [CrossRef]
- Santoro, M.; Viscardi, M.; Boccia, F.; Borriello, G.; Lucibelli, M.G.; Auriemma, C.; Anastasio, A.; Veneziano, V.; Galiero, G.; Baldi, L.; et al. Parasite Load and STRs Genotyping of Toxoplasma gondii Isolates from Mediterranean Mussels (Mytilus galloprovincialis) in Southern Italy. Front. Microbiol. 2020, 11, 355. Available online: https://www.frontiersin.org/articles/10.3389/fmicb.2020.00355 (accessed on 8 August 2023). [CrossRef]
- Putignani, L.; Mancinelli, L.; Del Chierico, F.; Menichella, D.; Adlerstein, D.; Angelici, M.; Marangi, M.; Berrilli, F.; Caffara, M.; di Regalbono, D.F.; et al. Investigation of Toxoplasma gondii presence in farmed shellfish by nested-PCR and real-time PCR fluorescent amplicon generation assay (FLAG). Exp. Parasitol. 2011, 127, 409–417. [Google Scholar] [CrossRef]
- Di Guardo, G.; Proietto, U.; Di Francesco, C.E.; Marsilio, F.; Zaccaroni, A.; Scaravelli, D.; Mignone, W.; Garibaldi, F.; Kennedy, S.; Forster, F.; et al. Cerebral Toxoplasmosis in Striped Dolphins (Stenella coeruleoalba) Stranded Along the Ligurian Sea Coast of Italy. Vet. Pathol. 2010, 47, 245–253. [Google Scholar] [CrossRef]
- Di Guardo, G.; Di Cesare, A.; Otranto, D.; Casalone, C.; Iulini, B.; Mignone, W.; Tittarelli, C.; Meloni, S.; Castagna, G.; Forster, F.; et al. Genotyping of Toxoplasma gondii isolates in meningo-encephalitis affected striped dolphins (Stenella coeruleoalba) from Italy. Vet. Parasitol. 2011, 183, 31–36. [Google Scholar] [CrossRef]
- Profeta, F.; Di Francesco, C.E.; Marsilio, F.; Mignone, W.; Di Nocera, F.; De Carlo, E.; Lucifora, G.; Pietroluongo, G.; Baffoni, M.; Cocumelli, C.; et al. Retrospective seroepidemiological investigations against Morbillivirus, Toxoplasma gondii and Brucella spp. in cetaceans stranded along the Italian coastline (1998–2014). Res. Vet. Sci. 2015, 101, 89–92. [Google Scholar] [CrossRef]
- Mazzariol, S.; Marcer, F.; Mignone, W.; Serracca, L.; Goria, M.; Marsili, L.; Di Guardo, G.; Casalone, C. Dolphin Morbillivirus and Toxoplasma gondii coinfection in a Mediterranean fin whale (Balaenoptera physalus). BMC Vet. Res. 2012, 8, 20. [Google Scholar] [CrossRef] [PubMed]
- Marcer, F.; Marchiori, E.; Centelleghe, C.; Ajzenberg, D.; Gustinelli, A.; Meroni, V.; Mazzariol, S. Parasitological and pathological findings in fin whales Balaenoptera physalus stranded along Italian coastlines. Dis. Aquat. Organ. 2019, 133, 25–37. [Google Scholar] [CrossRef]
- Fernández-Escobar, M.; Giorda, F.; Mattioda, V.; Audino, T.; Di Nocera, F.; Lucifora, G.; Varello, K.; Grattarola, C.; Ortega-Mora, L.M.; Casalone, C.; et al. Toxoplasma gondii Genetic Diversity in Mediterranean Dolphins. Pathogens 2022, 11, 909. [Google Scholar] [CrossRef] [PubMed]
- Petrella, A.; Mazzariol, S.; Padalino, I.; Di Francesco, G.; Casalone, C.; Grattarola, C.; Di Guardo, G.; Smoglica, C.; Centelleghe, C.; Gili, C. Cetacean Morbillivirus and Toxoplasma gondii Co-infection in Mediterranean Monk Seal Pup, Italy. Emerg. Infect. Dis. 2021, 27, 1237–1239. [Google Scholar] [CrossRef] [PubMed]
- Heim, R.R. An overview of weather and climate extremes—Products and trends. Weather. Clim. Extrem. 2015, 10, 1–9. [Google Scholar] [CrossRef]
- Jones, J.L.; Dargelas, V.; Roberts, J.; Press, C.; Remington, J.S.; Montoya, J.G. Risk factors for Toxoplasma gondii infection in the United States. Clin. Infect. Dis. 2009, 49, 878–884. [Google Scholar] [CrossRef]
- Grattarola, C.; Giorda, F.; Iulini, B.; Pintore, M.D.; Pautasso, A.; Zoppi, S.; Goria, M.; Romano, A.; Peletto, S.; Varello, K.; et al. Meningoencephalitis and Listeria monocytogenes, Toxoplasma gondii and Brucella spp. coinfection in a dolphin in Italy. Dis. Aquat. Organ. 2016, 118, 169–174. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Rodriguez-Fernandez, V.; Bruschi, F. Toxoplasma gondii in Marine Life of Italian Coasts, What Do We Know So Far? Parasitologia 2023, 3, 364-373. https://0-doi-org.brum.beds.ac.uk/10.3390/parasitologia3040035
AMA Style
Rodriguez-Fernandez V, Bruschi F. Toxoplasma gondii in Marine Life of Italian Coasts, What Do We Know So Far? Parasitologia. 2023; 3(4):364-373. https://0-doi-org.brum.beds.ac.uk/10.3390/parasitologia3040035
Chicago/Turabian StyleRodriguez-Fernandez, Veronica, and Fabrizio Bruschi. 2023. "Toxoplasma gondii in Marine Life of Italian Coasts, What Do We Know So Far?" Parasitologia 3, no. 4: 364-373. https://0-doi-org.brum.beds.ac.uk/10.3390/parasitologia3040035