
Challenges in Tick-Borne Pathogen Detection: The Case for Babesia spp. Identification in the Tick Vector

 ,
,
Abstract
:1. Introduction
2. Detection Methods for Babesia Identification
2.1. Bovine Babesiosis
2.1.1. Microscopy Tools
2.1.2. Molecular Tools
2.1.3. Other Babesia Species that Affect Cattle
2.2. Canine Babesiosis
Molecular Tools
2.3. Cervid Babesiosis
Molecular Tools
2.4. Equine Babesiosis
Molecular Tools
2.5. Human Babesiosis
2.5.1. Molecular Tools
2.5.2. Biological Tools
2.6. Ovine Babesiosis
2.6.1. Microscopy Tools
2.6.2. Molecular Tools
3. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gondard, M.; Cabezas-Cruz, A.; Charles, R.A.; Vayssier-Taussat, M.; Albina, E.; Moutailler, S. Ticks and tick-borne pathogens of the Caribbean: Current understanding and future directions for more comprehensive surveillance. Front. Cell. Infect. Microbiol. 2017, 7, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Dantas-Torres, F.; Chomel, B.B.; Otranto, D. Ticks and tick-borne diseases: A one health perspective. Trends Parasitol. 2012, 28, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Dantas-Torres, F. Species concepts: What about ticks? Trends Parasitol. 2018, 34, 1017–1026. [Google Scholar] [CrossRef]
- Gray, J.S.; Estrada-Peña, A.; Zintl, A. Vectors of babesiosis. Annu. Rev. Entomol. 2019, 64, 149–165. [Google Scholar] [CrossRef] [PubMed]
- Guglielmone, A.A.; Robbins, R.G.; Apanaskevich, D.A.; Petney, T.N.; Estrada-Peña, A.; Horak, I.G.; Shao, R.; Barker, S.C. The Argasidae, Ixodidae and Nuttalliellidae (Acari: Ixodida) of the world: A list of valid species names. Zootaxa 2010, 2528, 1–28. [Google Scholar] [CrossRef] [Green Version]
- Schnittger, L.; Rodriguez, A.E.; Florin-Christensen, M.; Morrison, D.A. Babesia: A world emerging. Infect. Genet. Evol. 2012, 12, 1788–1809. [Google Scholar] [CrossRef]
- Bock, R.; Jackson, L.; De Vos, A.; Jorgensen, W. Babesiosis of cattle. Parasitology 2004, 129, 247–269. [Google Scholar] [CrossRef]
- Uilenberg, G. Babesia a historical overview. Vet. Parasitol. 2006, 138, 3–10. [Google Scholar] [CrossRef]
- Vannier, E.G.; Diuk-Wasser, M.A.; Ben Mamoun, C.; Krause, P.J. Babesiosis. Infect. Dis. Clin. North. Am. 2015, 29, 357–370. [Google Scholar] [CrossRef] [Green Version]
- Levine, N.D. Taxonomy of the piroplasms. Trans. Am. Microsc. Soc. 1971, 90, 2–33. [Google Scholar] [CrossRef]
- Babes, V. Sur l’hemoglobinurie bacterienne du boeuf (on the bacterian hemoglobinuria of cattle). Compt. Rend. Hebd. Acad. Sci. 1888, 107, 692–694. (In French) [Google Scholar]
- Babes, V. L’etiologie d’une enzootie des moutons, dénommé Carceag en Roumanie. Compt. Rend. Hebd. Acad. Sci. 1892, 105, 359–361. [Google Scholar]
- Smith, T.; Kilborne, F.L. Investigation into the Nature Causation and Prevention of Texas or Southern Cattle Fever; US Department of Agriculture, Bureau of Animal Industry: Washington, DC, USA, 1893; Volume 1, pp. 1–301. [Google Scholar]
- Skrabalo, Z.; Deanovic, Z. Piroplasmosis in man. Report of a case. Doc. Med. Geogr. Trop. 1957, 9, 11–16. [Google Scholar] [PubMed]
- Western, K.A.; Benson, G.D.; Gleason, N.N.; Healy, G.R.; Schultz, M.G. Babesiosis in a Massachusetts resident. N. Engl. J. Med. 1970, 283, 854–856. [Google Scholar] [CrossRef] [PubMed]
- Vannier, E.; Gewurz, B.E.; Krause, P.J. Human babesiosis. Infect. Dis. Clin. North Am. 2008, 22, 469–488. [Google Scholar] [CrossRef] [Green Version]
- Gray, S.J.; Weiss, M.L. Babesia microti. In Emerging Protozoan Pathogens; Khan, N.A., Ed.; Taylor & Francis: New York, NY, USA, 2008; pp. 303–349. [Google Scholar]
- Lack, J.B.; Reichard, M.V.; Van Den Bussche, R.A. Phylogeny and evolution of the Piroplasmida as inferred from 18S rRNA sequences. Int. J. Parasitol. 2012, 42, 353–363. [Google Scholar] [CrossRef]
- Mehlhorn, H.; Schein, E. The piroplasms: Life cycle and sexual stages. Adv. Parasitol. 1985, 23, 37–103. [Google Scholar] [CrossRef]
- Chauvin, A.; Moreau, E.; Bonnet, S.; Plantard, O.; Malandrin, L. Babesia and its hosts: Adaptation to long-lasting interactions as a way to achieve efficient transmission. Vet. Res. 2009, 40, 37. [Google Scholar] [CrossRef] [Green Version]
- Jalovecka, M.; Hajdusek, O.; Sojka, D.; Kopacek, P.; Malandrin, L. The complexity of piroplasms life cycles. Front. Cell. Infect. Microbiol. 2018, 8, 248. [Google Scholar] [CrossRef] [Green Version]
- Jalovecka, M.; Sojka, D.; Ascencio, M.; Schnittger, L. Babesia life cycle—When phylogeny meets biology. Trends Parasitol. 2019, 35, 356–368. [Google Scholar] [CrossRef]
- Alvarez, J.A.; Rojas, C.; Figueroa, J.V. Diagnostic tools for the identification of Babesia sp. in persistently infected cattle. Pathoghens 2019, 8, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Homer, M.J.; Aguilar-Delfin, I.; Telford, S.R.; Krause, P.J.; Persing, D.H. Babesiosis. Clin. Microbiol. Rev. 2000, 13, 451–469. [Google Scholar] [CrossRef] [PubMed]
- Lei, R.; Wang, X.; Zhang, D.; Liu, Y.; Chen, Q.; Jiang, N. Rapid isothermal duplex real-time recombinase polymerase amplification (RPA) assay for the diagnosis of equine piroplasmosis. Sci. Rep. 2020, 10, 4096. [Google Scholar] [CrossRef] [Green Version]
- Lempereur, L.; Beck, R.; Fonseca, I.; Marques, C.; Duarte, A.; Santos, M.; Zúquete, S.; Gomes, J.; Walder, G.; Domingos, A.; et al. Guidelines for the detection of Babesia and Theileria parasites. Vector Borne Zoonotic. Dis. 2017, 17, 51–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, A.; Guan, G.; Du, P.; Gou, H.; Liu, Z.; Liu, J.; Ma, M.; Yang, J.; Li, Y.; Niu, Q.; et al. Loop-mediated isothermal amplification (LAMP) method based on two species-specific primer sets for the rapid identification of Chinese Babesia bovis and B. bigemina. Parasitol Int. 2012, 61, 658–663. [Google Scholar] [CrossRef] [PubMed]
- Mosqueda, J.; Olvera-Ramirez, A.; Aguilar-Tipacamu, G.; Canto, G.J. Current advances in detection and treatment of babesiosis. Curr. Med. Chem. 2012, 19, 1504–1518. [Google Scholar] [CrossRef] [Green Version]
- Sparagano, O.A.; Allsopp, M.T.; Mank, R.A.; Rijpkema, S.G.; Figueroa, J.V.; Jongejan, F. Molecular detection of pathogen DNA in ticks (Acari: Ixodidae): A review. Exp. Appl. Acarol. 1999, 23, 929–960. [Google Scholar] [CrossRef]
- Westblade, L.F.; Simon, M.S.; Mathison, B.A.; Kirkman, L.A. Babesia microti: From mice to ticks to an increasing number of highly susceptible humans. J. Clin. Microbiol. 2017, 55, 2903–2912. [Google Scholar] [CrossRef] [Green Version]
- Sivakumar, T.; Igarashi, I.; Yokoyama, N. Babesia ovata: Taxonomy, phylogeny and epidemiology. Vet. Parasitol. 2016, 229, 99–106. [Google Scholar] [CrossRef]
- Jacob, S.S.; Sengupta, P.P.; Paramanandham, K.; Suresh, K.P.; Chamuah, J.K.; Rudramurthy, G.R.; Roy, P. Bovine babesiosis: An insight into the global perspective on the disease distribution by systematic review and meta-analysis. Vet. Parasitol. 2020, 283, 109136. [Google Scholar] [CrossRef]
- Guglielmone, A.A.; Gaido, A.B.; Mangold, A.J. Light microscopy diagnosis of Babesia bovis and Babesia bigemina kinetes in the haemolymph of artificially infected Boophilus microplus engorged female ticks. Vet. Parasitol. 1996, 61, 15–20. [Google Scholar] [CrossRef]
- Oliveira, M.C.S.; Oliveira-Sequeira, T.C.G.; Araujo, J.P., Jr.; Amarante, A.F.T.; Oliveira, H.N. Babesia spp. infection in Boophilus microplus engorged females and eggs in Sao Paulo State, Brazil. Vet. Parasitol. 2005, 130, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Howell, J.M.; Ueti, M.W.; Palmer, G.H.; Scoles, G.A.; Knowles, D.P. Transovarial transmission efficiency of Babesia bovis tick stages acquired by Rhipicephalus (Boophilus) microplus during acute infection. J. Clin. Microbiol. 2007, 45, 426–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howell, J.M.; Ueti, M.W.; Palmer, G.H.; Scoles, G.A.; Knowles, D.P. Persistently infected calves as reservoirs for acquisition and transovarial transmission of Babesia bovis by Rhipicephalus (Boophilus) microplus. J. Clin. Microbiol. 2007, 45, 3155–3159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, M.C.S.; Oliveira-Sequeira, T.C.G.; Regitano, L.C.A.; Alencar, M.M.; Néo, T.A.; Silva, A.M.; Oliveira, H.N. Detection of Babesia bigemina in cattle of different genetic groups and in Rhipicephalus (Boophilus) microplus tick. Vet. Parasitol. 2008, 155, 281–286. [Google Scholar] [CrossRef]
- Figueroa, J.V.; Chieves, L.P.; Johnson, G.S.; Buening, G.M. Multiplex polymerase chain reaction based assay for the detection of Babesia bigemina, Babesia bovis and Anaplasma marginale DNA in bovine blood. Vet. Parasitol. 1993, 50, 69–81. [Google Scholar] [CrossRef]
- Figueroa, J.V.; Álvarez, J.A. Investigaciones sobre la aplicación de técnicas moleculares en el diagnóstico y control de la babesiosis bovina. Cienc. Vet. 2003, 9, 75–104. [Google Scholar]
- Smeenk, I.; Kelly, P.J.; Wray, K.; Musuka, G.; Trees, A.J.; Jongejan, F. Babesia bovis and B. bigemina DNA detected in cattle and ticks from Zimbabwe by polymerase chain reaction. J. South Afr. Vet. Assoc. 2000, 71, 21–24. [Google Scholar] [CrossRef] [Green Version]
- Oliveira-Sequeira, T.C.; Oliveira, M.C.; Araujo, J.P., Jr.; Amarante, A.F. PCR-based detection of Babesia bovis and Babesia bigemina in their natural host Boophilus microplus and cattle. Int. J. Parasitol. 2005, 35, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Bhat, S.A.; Singh, N.K.; Singh, H.; Rath, S.S. Molecular prevalence of Babesia bigemina in Rhipicephalus microplus ticks infesting cross-bred cattle of Punjab, India. Parasite Epidemiol. Control. 2017, 2, 85–90. [Google Scholar] [CrossRef]
- Gonzalez, J.; Echaide, I.; Pabón, A.; Gabriel Piñeros, J.J.; Blair, S.; Tobón-Castaño, A. Babesiosis prevalence in malaria-endemic regions of Colombia. J. Vector Borne Dis. 2018, 55, 222–229. [Google Scholar] [CrossRef]
- Guerrero, F.D.; Bendele, K.G.; Davey, R.B.; George, J.E. Detection of Babesia bigemina infection in strains of Rhipicephalus (Boophilus) microplus collected from outbreaks in South Texas. Vet. Parasitol. 2007, 145, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Adham, F.K.; Abd-El-Samie, E.M.; Gabre, R.M.; El-Hussein, H. Detection of tick blood parasites in Egypt using PCR assay I-Babesia bovis and Babesia bigemina. Parasitol. Res. 2009, 105, 721–730. [Google Scholar] [CrossRef] [PubMed]
- Carret, C.; Walas, F.; Carcy, B.; Grande, N.; Précigout, E.; Moubri, K.; Schetters, T.P.; Gorenflot, A. Babesia canis canis, Babesia canis vogeli, Babesia canis rossi: Differentiation of the three subspecies by a Restriction Fragment Length Polymorphism analysis on amplified small subunit ribosomal RNA genes. J. Eukaryot. Microbiol. 1999, 46, 298–303. [Google Scholar] [CrossRef] [PubMed]
- Figueroa-Millán, J.V.; Lira-Amaya, J.J.; Castañeda-Arriola, R.; Álvarez-Martínez, J.A.; Rojas-Martínez, C.; Bautista-Garfias, C.R. Optimización de una prueba de PCR-RFLP para detección y diferenciación de Babesia sp. en garrapatas Rhipicephalus microplus. Entomol. Mex. 2014, 1, 978–983. [Google Scholar]
- Giglioti, R.; Oliveira, H.N.; Santana, C.H.; Ibelli, A.M.G.; Néo, T.A.; Bilhassi, T.B.; Rabelo, M.D.; Machado, R.Z.; Brito, L.G.; Oliveira, M.C.S. Babesia bovis and Babesia bigemina infection levels estimated by qPCR in Angus cattle from an endemic area of São Paulo state, Brazil. Ticks Tick Borne Dis. 2016, 7, 657–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giglioti, R.; Nunes de Oliveira, H.; Okino, C.H.; de Sena Oliveira, M.C. qPCR estimates of Babesia bovis and Babesia bigemina infection levels in beef cattle and Rhipicephalus microplus larvae. Exp. Appl. Acarol. 2018, 75, 235–240. [Google Scholar] [CrossRef] [Green Version]
- Obregón, D.; Corona-González, B.; Díaz-Sánchez, A.A.; Armas, Y.; Roque, E.; de Sena Oliveira, M.C.; Cabezas-Cruz, A. Efficient transovarial transmission of Babesia spp. in Rhipicephalus microplus ticks fed on water buffalo (Bubalus bubalis). Pathoghens 2020, 9, 280. [Google Scholar] [CrossRef]
- Merino, O.; Almazán, C.; Canales, M.; Villar, M.; Moreno-Cid, J.A.; Galindo, R.C.; de la Fuente, J. Targeting the tick protective antigen subolesin reduces vector infestations and pathogen infection by Anaplasma marginale and Babesia bigemina. Vaccine 2011, 29, 8575–8579. [Google Scholar] [CrossRef]
- Antunes, S.; Galindo, R.C.; Almazán, C.; Rudenko, N.; Golovchenko, M.; Grubhoffer, L.; Shkap, V.; do Rosário, V.; de la Fuente, J.; Domingos, A. Functional genomics studies of Rhipicephalus (Boophilus) annulatus ticks in response to infection with the cattle protozoan parasite, Babesia bigemina. Int. J. Parasitol. 2012, 42, 187–195. [Google Scholar] [CrossRef]
- Hussein, H.E.; Johnson, W.C.; Taus, N.S.; Suarez, C.E.; Scoles, G.A.; Ueti, M.W. Silencing expression of the Rhipicephalus microplus vitellogenin receptor gene blocks Babesia bovis transmission and interferes with oocyte maturation. Parasit. Vectors 2019, 12, 7. [Google Scholar] [CrossRef] [PubMed]
- AL-Hosary, A.A.T.A. Loop-mediated isothermal amplification (LAMP) assay for diagnosis of bovine babesiosis (Babesia bovis infection) in Egypt. J. Adv.Vet. Res. 2017, 7, 71–74. [Google Scholar]
- Bonnet, S.; Jouglin, M.; Malandrin, L.; Becker, C.; Agoulon, A.; L’Hostis, M.; Chauvin, A. Transstadial and transovarial persistence of Babesia divergens DNA in Ixodes ricinus ticks fed on infected blood in a new skin-feeding technique. Parasitology 2007, 134, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Hilpertshauser, H.; Deplazes, P.; Schnyder, M.; Gern, L.; Mathis, A. Babesia spp. Identified by PCR in ticks collected from domestic and wild ruminants in southern Switzerland. Appl. Environ. Microbiol. 2006, 72, 6503–6507. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Zhou, Y.Q.; Zhou, D.N.; Liu, E.Y.; Du, K.; Chen, S.G.; Yao, B.A.; Zhao, J.L. Semi-nested PCR detection of Babesia orientalis in its natural hosts Rhipicephalus haemaphysaloides and buffalo. Vet. Parasitol. 2007, 143, 260–266. [Google Scholar] [CrossRef]
- He, L.; Zhou, Y.Q.; Oosthuizen, M.C.; Zhao, J.L. Loop-mediated isothermal amplification (LAMP) detection of Babesia orientalis in water buffalo (Bubalus babalis, Linnaeus, 1758) in China. Vet Parasitol. 2009, 165, 36–40. [Google Scholar] [CrossRef] [Green Version]
- Gray, S.; de Vos, A. Studies on a bovine Babesia transmitted by Hyalomma marginatum rufipes Koch, 1844. OnderstepoortJ. Vet. Res. 1981, 48, 215–223. [Google Scholar]
- Ros-García, A.; M’Ghirbi, Y.; Bouattour, A.; Hurtado, A. First detection of Babesia occultans in Hyalomma ticks from Tunisia. Parasitology 2011, 138, 578–582. [Google Scholar] [CrossRef]
- Sivakumar, T.; Tattiyapong, M.; Okubo, K.; Suganuma, K.; Hayashida, K.; Igarashi, I.; Zakimi, S.; Matsumoto, K.; Inokuma, H.; Yokoyama, N. PCR detection of Babesia ovata from questing ticks in Japan. Ticks Tick Borne Dis. 2014, 5, 305–310. [Google Scholar] [CrossRef] [Green Version]
- Uilenberg, G.; Franssen, F.F.J.; Perié, N.M.; Spanjer, A.A.M. Three groups of Babesia canis distinguished and a proposal for nomenclature. Vet. Q. 1989, 11, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Irwin, P.J. Canine babesiosis: From molecular taxonomy to control. Parasit. Vectors. 2009, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birkenheuer, A.J. Babesiosis. Canine Feline Infect. Dis. 2014, 727–738. [Google Scholar] [CrossRef]
- Petra, B.; Josipa, K.; Renata, B.R.; Vladimir, M. Canine babesiosis: Where do we stand? Acta Vet. 2018, 68, 127–160. [Google Scholar] [CrossRef] [Green Version]
- Matijatko, V.; Torti, M.; Schetters, T.P. Canine babesiosis in Europe: How many diseases? Trends Parasitol. 2012, 28, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Olmeda, A.S.; Armstrong, P.M.; Rosenthal, B.M.; Valladares, B.; del Castillo, A.; de Armas, F.; Miguelez, M.; González, A.; Rodríguez Rodríguez, J.A.; Spielman, A.; et al. A subtropical case of human babesiosis. Acta Trop. 1997, 67, 229–234. [Google Scholar] [CrossRef]
- Birkenheuer, A.J.; Levy, M.G.; Breitschwerdt, E.B. Development and evaluation of a seminested PCR for detection and differentiation of Babesia gibsoni (Asian genotype) and B. canis DNA in canine blood samples. J. Clin. Microbiol. 2003, 41, 4172–4177. [Google Scholar] [CrossRef] [Green Version]
- Trotta, M.; Nicetto, M.; Fogliazza, A.; Montarsi, F.; Caldin, M.; Furlanello, T.; Solano-Gallego, L. Detection of Leishmania infantum, Babesia canis, and rickettsiae in ticks removed from dogs living in Italy. Ticks Tick Borne Dis. 2012, 3, 294–297. [Google Scholar] [CrossRef]
- Lira-Amaya, J.J.; Rojas-Martinez, C.; Alvarez-Martinez, A.; Pelaez-Flores, A.; Martinez-Ibañez, F.; Perez-de la Rosa, D.; Figueroa-Millan, J.V. First molecular detection of Babesia canis vogeli in dogs and Rhipicephalus sanguineus from Mexico. Arch. Palliat. Care. 2017, 2, 1013. [Google Scholar]
- Azmi, K.; Ereqat, S.; Nasereddin, A.; Al-Jawabreh, A.; Baneth, G.; Abdeen, Z. Molecular detection of Theileria, Babesia, and Hepatozoon spp. in ixodid ticks from Palestine. Ticks Tick Borne Dis. 2016, 7, 734–741. [Google Scholar] [CrossRef]
- Casati, S.; Sager, H.; Gern, L.; Piffaretti, J.C. Presence of potentially pathogenic Babesia sp. for human in Ixodes ricinus in Switzerland. Ann. Agric. Environ. Med. 2006, 13, 65–70. [Google Scholar]
- Jain, J.K.; Lakshmanan, B.; Nagaraj, H.V.; Praveena, J.E.; Syamala, K.; Aravindakshan, T. Detection of Babesia canis vogeli, Babesia gibsoni and Ehrlichia canis by multiplex PCR in naturally infected dogs in South India. Vet. Arh. 2018, 88, 215–224. [Google Scholar] [CrossRef]
- Jain, J.; Lakshmanan, B.; Wahlang, L.; Syamala, K.; Aravindakshan, T.V. Molecular evidence of haemoparasites in ixodid ticks of dogs-first report in India. Vet. Parasitol. Reg. Stud. Rep. 2018, 13, 177–179. [Google Scholar] [CrossRef] [PubMed]
- Annoscia, G.; Latrofa, M.S.; Cantacessi, C.; Olivieri, E.; Manfredi, M.T.; Dantas-Torres, F.; Otranto, D. A new PCR assay for the detection and differentiation of Babesia canis and Babesia vogeli. Ticks Tick Borne Dis. 2017, 8, 862–865. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Peña, A.; Gray, J.S.; Kahl, O.; Lane, R.S.; Nijhof, A.M. Research on the ecology of ticks and tick-borne pathogens methodological principles and caveats. Front. Cell. Infect. Microbiol. 2013, 3, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, F.D.; Wall, L.E.R. Prevalence of Babesia and Anaplasma in ticks infesting dogs in Great Britain. Vet. Parasitol. 2013, 198, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Chao, L.L.; Liao, H.T.; Ho, T.Y.; Shih, C.M. First detection and molecular identification of Babesia gibsoni from Rhipicephalus sanguineus ticks. Acta Trop. 2017, 166, 356–362. [Google Scholar] [CrossRef]
- René, M.; Chêne, J.; Beaufils, J.P.; Valiente Moro, C.; Bourdoiseau, G.; Mavingui, P.; Chabanne, L. First evidence and molecular characterization of Babesia vogeli in naturally infected dogs and Rhipicephalus sanguineus ticks in southern France. Vet. Parasitol. 2012, 187, 399–407. [Google Scholar] [CrossRef]
- Yisaschar-Mekuzas, Y.; Jaffe, C.L.; Pastor, J.; Cardoso, L.; Baneth, G. Identification of Babesia species infecting dogs using reverse line blot hybridization for six canine piroplasms, and evaluation of co-infection by other vector-borne pathogens. Vet. Parasitol. 2013, 191, 367–373. [Google Scholar] [CrossRef]
- M’Ghirbi, Y.; Bouattour, A. Detection and molecular characterization of Babesia canis vogeli from naturally infected dogs and Rhipicephalus sanguineus ticks in Tunisia. Vet. Parasitol. 2008, 152, 1–7. [Google Scholar] [CrossRef]
- Harrus, S.; Perlman-Avrahami, A.; Mumcuoglu, K.Y.; Morick, D.; Eyal, O.; Baneth, G. Molecular detection of Ehrlichia canis, Anaplasma bovis, Anaplasma platys, Candidatus Midichloria mitochondrii and Babesia canis vogeli in ticks from Israel. Clin. Microbiol. Infect. 2011, 17, 459–463. [Google Scholar] [CrossRef] [Green Version]
- Manoj, R.R.S.; Iatta, R.; Latrofa, M.S.; Capozzi, L.; Raman, M.; Colella, V.; Otranto, D. Canine vector-borne pathogens from dogs and ticks from Tamil Nadu, India. Acta Trop. 2020, 203, 105308. [Google Scholar] [CrossRef] [PubMed]
- Jongejan, F.; Su, B.L.; Yang, H.J.; Berger, L.; Bevers, J.; Liu, P.C.; Fang, J.C.; Cheng, Y.W.; Kraakman, C.; Plaxton, N. Molecular evidence for the transovarial passage of Babesia gibsoni in Haemaphysalis hystricis (Acari: Ixodidae) ticks from Taiwan: A novel vector for canine babesiosis. Parasit. Vectors 2018, 11, 134. [Google Scholar] [CrossRef] [PubMed]
- Matjila, T.P.; Nijhof, A.M.; Taoufik, A.; Houwers, D.; Teske, E.; Penzhorn, B.L.; de Lange, T.; Jongejan, F. Autochthonous canine babesiosis in The Netherlands. Vet. Parasitol. 2005, 131, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Jongejan, F.; Fourie, J.J.; Chester, S.T.; Manavella, C.; Mallouk, Y.; Pollmeier, M.G.; Baggott, D. The prevention of transmission of Babesia canis canis by Dermacentor reticulatus ticks to dogs using a novel combination of fipronil, amitraz and (S)-methoprene. Vet. Parasitol. 2011, 179, 343–350. [Google Scholar] [CrossRef]
- Zygner, W.; Jaros, S.; Wedrychowicz, H. Prevalence of Babesia canis, Borrelia afzelii, and Anaplasma phagocytophilum infection in hard ticks removed from dogs in Warsaw (central Poland). Vet. Parasitol. 2008, 153, 139–142. [Google Scholar] [CrossRef]
- Karbowiak, G.; Vichová, B.; Slivinska, K.; Werszko, J.; Didyk, J.; Petko, B.; Stanko, M.; Akimov, I. The infection of questing Dermacentor reticulatus ticks with Babesia canis and Anaplasma phagocytophilum in the Chernobyl exclusion zone. Vet. Parasitol. 2014, 204, 372–375. [Google Scholar] [CrossRef]
- Hornok, S.; Kartali, K.; Takács, N.; Hofmann-Lehmann, R. Uneven seasonal distribution of Babesia canis and its two 18S rDNA genotypes in questing Dermacentor reticulatus ticks in urban habitats. Ticks Tick Borne Dis. 2016, 7, 694–697. [Google Scholar] [CrossRef]
- Geurden, T.; Becskei, C.; Six, R.H.; Maeder, S.; Latrofa, M.S.; Otranto, D.; Farkas, R. Detection of tick-borne pathogens in ticks from dogs and cats in different European countries. Ticks Tick Borne Dis. 2018, 9, 1431–1436. [Google Scholar] [CrossRef]
- Sprong, H.; Fonville, M.; Docters van Leeuwen, A.; Devillers, E.; Ibañez-Justicia, A.; Stroo, A.; Hansford, K.; Cull, B.; Medlock, J.; Heyman, P.; et al. Detection of pathogens in Dermacentor reticulatus in northwestern Europe: Evaluation of a high-throughput array. Heliyon 2019, 5, E01270. [Google Scholar] [CrossRef] [Green Version]
- Rar, V.A.; Fomenko, N.V.; Dobrotvorsky, A.K.; Livanova, N.N.; Rudakova, S.A.; Fedorov, E.G.; Astanin, V.B.; Morozova, O.V. Tickborne pathogen detection, Western Siberia, Russia. Emerg. Infect. Dis. 2005, 11, 1708–1715. [Google Scholar] [CrossRef]
- Radzijevskaja, J.; Mardosaitė-Busaitienė, D.; Aleksandravičienė, A.; Paulauskas, A. Investigation of Babesia spp. in sympatric populations of Dermacentor reticulatus and Ixodes ricinus ticks in Lithuania and Latvia. Ticks Tick Borne Dis. 2018, 9, 270–274. [Google Scholar] [CrossRef] [PubMed]
- Adaszek, Ł.; Jankowska, M.; Kalinowski, M.; Banach, T.; Wułupek, D.; Winiarczyk, S. The loop-mediated isothermal amplification assay for rapid diagnosis of Babesia canis canis infections in dogs. Pol. J. Vet. Sci. 2013, 16, 131–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Modarelli, J.J.; Ferro, P.J.; Perez de León, A.A.; Esteve-Gasent, M.D. TickPath Layerplex: Adaptation of a real-time PCR methodology for the simultaneous detection and molecular surveillance of tick-borne pathogens. Sci. Rep. 2019, 9, 6950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pastor, A.; Milnes, E. Babesiosis in cervidae. Fowler’s Zoo Wild Anim. Med. Curr. Ther. 2019, 9, 647–655. [Google Scholar] [CrossRef]
- Wiegmann, L.; Silaghi, C.; Obiegala, A.; Karnath, C.; Langer, S.; Ternes, K.; Kämmerling, J.; Osmann, C.; Pfeffer, M. Occurrence of Babesia species in captive reindeer (Rangifer tarandus) in Germany. Vet. Parasitol. 2015, 211, 16–22. [Google Scholar] [CrossRef]
- Karnath, C.; Obiegala, A.; Speck, S.; Essbauer, S.; Derschum, H.; Scholz, H.; Kiefer, D.; Tserennorov, D.; Dashdavaa, O.; Tsogbadrakh, N.; et al. Detection of Babesia venatorum, Anaplasma phagocytophilum and Candidatus Neoehrlichia mikurensis in Ixodes persulcatus ticks from Mongolia. Ticks Tick Borne Dis. 2016, 7, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Kazimírová, M.; Hamšíková, Z.; Špitalská, E.; Minichová, L.; Mahríková, L.; Caban, R.; Sprong, H.; Fonville, M.; Schnittger, L.; Kocianová, E. Diverse tick-borne microorganisms identified in free-living ungulates in Slovakia. Parasit. Vectors. 2018, 11, 495. [Google Scholar] [CrossRef]
- Milnes, E.L.; Thornton, G.; Léveillé, A.N.; Delnatte, P.; Barta, J.R.; Smith, D.A.; Nemeth, N. Babesia odocoilei and zoonotic pathogens identified from Ixodes scapularis ticks in southern Ontario, Canada. Ticks Tick Borne Dis. 2019, 10, 670–676. [Google Scholar] [CrossRef]
- Lejal, E.; Moutailler, S.; Šimo, L.; Vayssier-Taussat, M.; Pollet, T. Tick-borne pathogen detection in midgut and salivary glands of adult Ixodes ricinus. Parasit. Vectors. 2019, 12, 152. [Google Scholar] [CrossRef] [Green Version]
- Rothschild, C.M. Equine piroplasmosis. J. Equine Vet. Sci. 2013, 33, 497–508. [Google Scholar] [CrossRef]
- Onyiche, T.E.; Suganuma, K.; Igarashi, I.; Yokoyama, N.; Xuan, X.; Thekisoe, O. A review on equine piroplasmosis: Epidemiology, vector ecology, risk factors, host immunity, diagnosis and control. Int. J. Environ. Res. Public Health. 2019, 16, 1736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tirosh-Levy, S.; Gottlieb, Y.; Fry, L.M.; Knowles, D.P.; Steinman, A. Twenty years of equine piroplasmosis research: Global distribution, molecular diagnosis, and phylogeny. Pathogens 2020, 9, 926. [Google Scholar] [CrossRef]
- Battsetseg, B.; Xuan, X.; Ikadai, H.; Rodriguez, B.J.L.; Byambaa, B.; Boldbaatar, D.; Battur, B.; Battsetseg, G.; Batsukh, Z.; Igarashi, I.; et al. Detection of Babesia caballi and Babesia equi in Dermacentor nuttalli adult ticks. Int. J. Parasitol. 2001, 31, 384–386. [Google Scholar] [CrossRef]
- Medrano-Bugarini, R.A.; Figueroa-Millán, J.V.; Rivera-Chavira, B.E.; Lira-Amaya, J.J.; Rodríguez-Alarcón, C.A.; Beristain-Ruiz, D.M.; Adame-Gallegos, J.R. Detection of Theileria equi, Babesia caballi, and Anaplasma phagocytophilum DNA in soft ticks and horses at Ciudad Juarez, Mexico. Southwest. Entomol. 2019, 44, 647–658. [Google Scholar] [CrossRef]
- Abedi, V.; Razmi, G.; Seifi, H.; Naghibi, A. Molecular and serological detection of Theileria equi and Babesia caballi infection in horses and ixodid ticks in Iran. Ticks Tick Borne Dis. 2014, 5, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Nagore, D.; García-Sanmartín, J.; García-Pérez, A.L.; Juste, R.A.; Hurtado, A. Detection and identification of equine Theileria and Babesia species by reverse line blotting: Epidemiological survey and phylogenetic analysis. Vet. Parasitol. 2004, 123, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Ros-García, A.; M’Ghirbi, Y.; Hurtado, A.; Bouattour, A. Prevalence and genetic diversity of piroplasm species in horses and ticks from Tunisia. Infect. Genet. Evol. 2013, 17, 33–37. [Google Scholar] [CrossRef]
- Alhassan, A.; Govind, Y.; Tam, N.T.; Thekisoe, O.M.M.; Yokoyama, N.; Inoue, N.; Igarashi, I. Comparative evaluation of the sensitivity of LAMP, PCR and in vitro culture methods for the diagnosis of equine piroplasmosis. Parasitol. Res. 2007, 100, 1165–1168. [Google Scholar] [CrossRef]
- Karbowiak, G.; Biernat, B.; Stańczak, J.; Werszko, J.; Szewczyk, T.; Sytykiewicz, H. The role of particular ticks developmental stages in the circulation of tick-borne pathogens in Central Europe. 6. Babesia Ann. Parasitol. 2018, 64, 265–284. [Google Scholar] [CrossRef]
- Krause, P.J. Human babesiosis. Int. J. Parasitol. 2019, 49, 165–174. [Google Scholar] [CrossRef]
- Jahfari, S.; Hofhuis, A.; Fonville, M.; van der Giessen, J.; van Pelt, W.; Sprong, H. Molecular detection of tick-borne pathogens in humans with tick bites and erythema migrans, in the Netherlands. PLoS Negl. Trop. Dis. 2016, 10, E0005042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, S.H.; Kim, S.Y.; Song, B.G.; Roh, J.R.; Cho, C.R.; Kim, C.N.; Um, T.H.; Kwak, Y.G.; Cho, S.H.; Lee, S.E. Detection and characterization of an emerging type of Babesia sp. similar to Babesia motasi for the first case of human babesiosis and ticks in Korea. Emerg. Microbes Infect. 2019, 8, 869–878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diuk-Wasser, M.A.; Liu, Y.; Steeves, T.K.; Folsom-O’Keefe, C.; Dardick, K.R.; Lepore, T.; Bent, S.J.; Usmani-Brown, S.; Telford, S.R.; Fish, D.; et al. Monitoring human babesiosis emergence through vector surveillance New England, USA. Emerg. Infect. Dis. 2014, 20, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Swei, A.; O’Connor, K.E.; Couper, L.I.; Thekkiniath, J.; Conrad, P.A.; Padgett, K.A.; Burns, J.; Yoshimizu, M.H.; Gonzales, B.; Munk, B.; et al. Evidence for transmission of the zoonotic apicomplexan parasite Babesia duncani by the tick Dermacentor albipictus. Int. J. Parasitol. 2019, 49, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Stensvold, C.R.; Al Marai, D.; Andersen, L.O.; Krogfelt, K.A.; Jensen, J.S.; Larsen, K.S.; Nielsen, H.V. Babesia spp. and other pathogens in ticks recovered from domestic dogs in Denmark. Parasit. Vectors. 2015, 8, 262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Liu, G.; Yang, L.; Xu, R.; Cao, W. Babesia microti-like rodent parasites isolated from Ixodes persulcatus (Acari: Ixodidae) in Heilongjiang Province, China. Vet. Parasitol. 2008, 156, 333–339. [Google Scholar] [CrossRef]
- Li, L.H.; Zhu, D.; Zhang, C.C.; Zhang, Y.; Zhou, X.N. Experimental transmission of Babesia microti by Rhipicephalus haemaphysaloides. Parasit Vectors. 2016, 25, 231. [Google Scholar] [CrossRef] [Green Version]
- Bonnet, S.; Liu, X.Y. Laboratory artificial infection of hard ticks: A tool for the analysis of tick-borne pathogen transmission. Acarologia 2012, 52, 453–464. [Google Scholar] [CrossRef] [Green Version]
- Becker, C.A.; Bouju-Albert, A.; Jouglin, M.; Chauvin, A.; Malandrin, L. Natural transmission of zoonotic Babesia spp. by Ixodes ricinus ticks. Emerg Infect Dis. 2009, 15, 320–322. [Google Scholar] [CrossRef]
- Liu, X.Y.; Cote, M.; Paul, R.E.; Bonnet, S.I. Impact of feeding system and infection status of the blood meal on Ixodes ricinus feeding. Ticks Tick Borne Dis. 2014, 5, 323–328. [Google Scholar] [CrossRef]
- Yeruham, I.; Hadani, A.; Galker, F. Some epizootiological and clinical aspects of ovine babesiosis caused by Babesia ovis—A review. Vet. Parasitol. 1998, 74, 153–163. [Google Scholar] [CrossRef]
- Esmaeilnejad, B.; Tavassoli, M.; Asri-Rezaei, S.; Dalir-Naghadeh, B.; Mardani, K.; Jalilzadeh-Amin, G.; Golabi, M.; Arjmand, J. PCR-based detection of Babesia ovis in Rhipicephalus bursa and small ruminants. J. Parasitol. Res. 2014, 2014, 294704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erster, O.; Roth, A.; Wolkomirsky, R.; Leibovich, B.; Savitzky, I.; Shkap, V. Transmission of Babesia ovis by different Rhipicephalus bursa developmental stages and infected blood injection. Ticks Tick Borne Dis. 2016, 7, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Guan, G.; Ma, M.; Moreau, E.; Liu, J.; Lu, B.; Bai, Q.; Luo, J.; Jorgensen, W.; Chauvin, A.; Yin, H. A new ovine Babesia species transmitted by Hyalomma anatolicum anatolicum. Exp. Parasitol. 2009, 122, 261–267. [Google Scholar] [CrossRef]
- Ozubek, S.; Aktas, M. Molecular evidence of a new Babesia sp. in goats. Vet. Parasitol. 2017, 233, 1–8. [Google Scholar] [CrossRef]
- Yeruham, I.; Hadani, A.; Galker, F. The effect of the ovine host parasitaemia on the development of Babesia ovis (Babes, 1892) in the tick Rhipicephalus bursa (Canestrini and Fanzago, 1877). Vet. Parasitol. 2001, 96, 195–202. [Google Scholar] [CrossRef]
- Razmi, G.R.; Naghibi, A.; Aslani, M.R.; Fathivand, M.; Dastjerdi, K. An epidemiological study on ovine babesiosis in the Mashhad suburb area, province of Khorasan, Iran. Vet. Parasitol. 2002, 108, 109–115. [Google Scholar] [CrossRef]
- Aktaş, M.; Altay, K.; Dumanli, N. Development of a polymerase chain reaction method for diagnosis of Babesia ovis infection in sheep and goats. Vet. Parasitol. 2005, 133, 277–281. [Google Scholar] [CrossRef]
- Aouadi, A.; Leulmi, H.; Boucheikhchoukh, M.; Benakhla, A.; Raoult, D.; Parola, P. Molecular evidence of tick-borne hemoprotozoan-parasites (Theileria ovis and Babesia ovis) and bacteria in ticks and blood from small ruminants in Northern Algeria. Comp. Immunol. Microbiol. Infect. Dis. 2017, 50, 34–39. [Google Scholar] [CrossRef]
- Herwaldt, B.L.; Cacciò, S.; Gherlinzoni, F.; Aspöck, H.; Slemenda, S.B.; Piccaluga, P.; Martinelli, G.; Edelhofer, R.; Hollenstein, U.; Poletti, G.; et al. Molecular characterization of a non-Babesia divergens organism causing zoonotic babesiosis in Europe. Emerg. Infect. Dis. 2003, 9, 942–948. [Google Scholar] [CrossRef]
- Erster, O.; Roth, A.; Wolkomirsky, R.; Leibovich, B.; Savitzky, I.; Zamir, S.; Molad, T.; Shkap, V. Molecular detection of Babesia ovis in sheep and ticks using the gene encoding B. ovis surface protein D (BoSPD). Vet. Parasitol. 2015, 214, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Erster, O.; Roth, A.; Wollkomirsky, R.; Leibovich, B.; Savitzky, I.; Zamir, S.; Molad, T.; Shkap, V. Quantitative analysis of Babesia ovis infection in sheep and ticks. Vet. Parasitol. 2016, 221, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Guan, G.; Chauvin, A.; Luo, J.; Inoue, N.; Moreau, E.; Liu, Z.; Gao, J.; Thekisoe, O.M.M.; Ma, M.; Liu, A.; et al. The development and evaluation of a loop-mediated isothermal amplification (LAMP) method for detection of Babesia spp. infective to sheep and goats in China. Exp. Parasitol. 2008, 120, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Rooney, A.P. Mechanisms underlying the evolution and maintenance of functionally heterogeneous 18S rRNA genes in Apicomplexans. Mol. Biol. Evol. 2004, 21, 1704–1711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunfeld, K.P.; Hildebrandt, A.; Gray, J.S. Babesiosis: Recent insights into an ancient disease. Int. J. Parasitol. 2008, 38, 1219–1237. [Google Scholar] [CrossRef] [PubMed]
- Hanron, A.E.; Billman, Z.P.; Seilie, A.M.; Chang, M.; Murphy, S.C. Detection of Babesia microti parasites by highly sensitive 18S rRNA reverse transcription PCR. Diagn. Microbiol. Infect. Dis. 2017, 87, 226–228. [Google Scholar] [CrossRef]
- Cacciò, S.; Cammà, C.; Onuma, M.; Severini, C. The β-tubulin gene of Babesia and Theileria parasites is an informative marker for species discrimination. Int. J. Parasitol. 2000, 30, 1181–1185. [Google Scholar] [CrossRef]
- Uilenberg, G.; Gray, J.; Kahl, O. Research on Piroplasmorida and other tick-borne agents: Are we going the right way? Ticks Tick Borne Dis. 2018, 9, 860–863. [Google Scholar] [CrossRef]
- Dantas-Torres, F. Climate change, biodiversity, ticks and tick-borne diseases: The butterfly effect. Int. J. Parasitol. Parasites Wildl. 2015, 4, 452–461. [Google Scholar] [CrossRef] [Green Version]
- Michelet, L.; Delannoy, S.; Devillers, E.; Umhang, G.; Aspan, A.; Juremalm, M.; Chirico, J.; van der Wal, F.J.; Sprong, H.; Boye-Pihl, T.P.; et al. High-throughput screening of tick-borne pathogens in Europe. Front. Cell. Infect. Microbiol. 2014, 4, 103. [Google Scholar] [CrossRef]
- Moutailler, S.; Valiente-Moro, C.; Vaumourin, E.; Michelet, L.; Tran, F.H.; Devillers, E.; Cosson, J.F.; Gasqui, P.; Van, T.V.; Mavingui, P.; et al. Co-infection of ticks: The rule rather than the exception. PLoS Negl. Trop. Dis. 2016, 10, e0004539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabezas-Cruz, A.; Allain, E.; Ahmad, A.S.; Saeed, M.A.; Rashid, I.; Ashraf, K.; Yousfi, L.; Shehzad, W.; Indjein, L.; Rodriguez-Valle, M.; et al. Low genetic diversity of Ehrlichia canis associated with high co-infection rates in Rhipicephalus sanguineus (s.l.). Parasit Vectors 2019, 12, 12. [Google Scholar] [CrossRef]
- Grech-Angelini, S.; Stachurski, F.; Vayssier-Taussat, M.; Devillers, E.; Casabianca, F.; Lancelot, R.; Uilenberg, G.; Moutailler, S. Tick-borne pathogens in ticks (Acari: Ixodidae) collected from various domestic and wild hosts in Corsica (France), a Mediterranean island environment. Transbound. Emerg. Dis. 2020, 67, 745–757. [Google Scholar] [CrossRef] [PubMed]
- Ghafar, A.; Khan, A.; Cabezas-Cruz, A.; Gauci, C.G.; Niaz, S.; Ayaz, S.; Mateos-Hernández, L.; Galon, C.; Nasreen, N.; Moutailler, S.; et al. An assessment of the molecular diversity of ticks and tick-borne microorganisms of small ruminants in Pakistan. Microorganisms 2020, 8, 1428. [Google Scholar] [CrossRef] [PubMed]
- Ghafar, A.; Cabezas-Cruz, A.; Galon, C.; Obregon, D.; Gasser, R.B.; Moutailler, S.; Jabbar, A. Bovine ticks harbour a diverse array of microorganisms in Pakistan. Parasit. Vectors. 2020, 13, 1. [Google Scholar] [CrossRef] [Green Version]
- Gondard, M.; Delannoy, S.; Pinarello, V.; Aprelon, R.; Devillers, E.; Galon, C.; Pradel, J.; Vayssier-Taussat, M.; Albina, E.; Moutailler, S. Upscaling the surveillance of tick-borne pathogens in the French caribbean islands. Pathogens 2020, 9, 176. [Google Scholar] [CrossRef] [Green Version]
- Ghafar, A.; Koehler, A.V.; Hall, R.S.; Gauci, C.G.; Gasser, R.B.; Jabbar, A. Targeted next-generation sequencing and informatics as an effective tool to establish the composition of bovine piroplasm populations in endemic regions. Microorganisms 2021, 9, 21. [Google Scholar] [CrossRef]



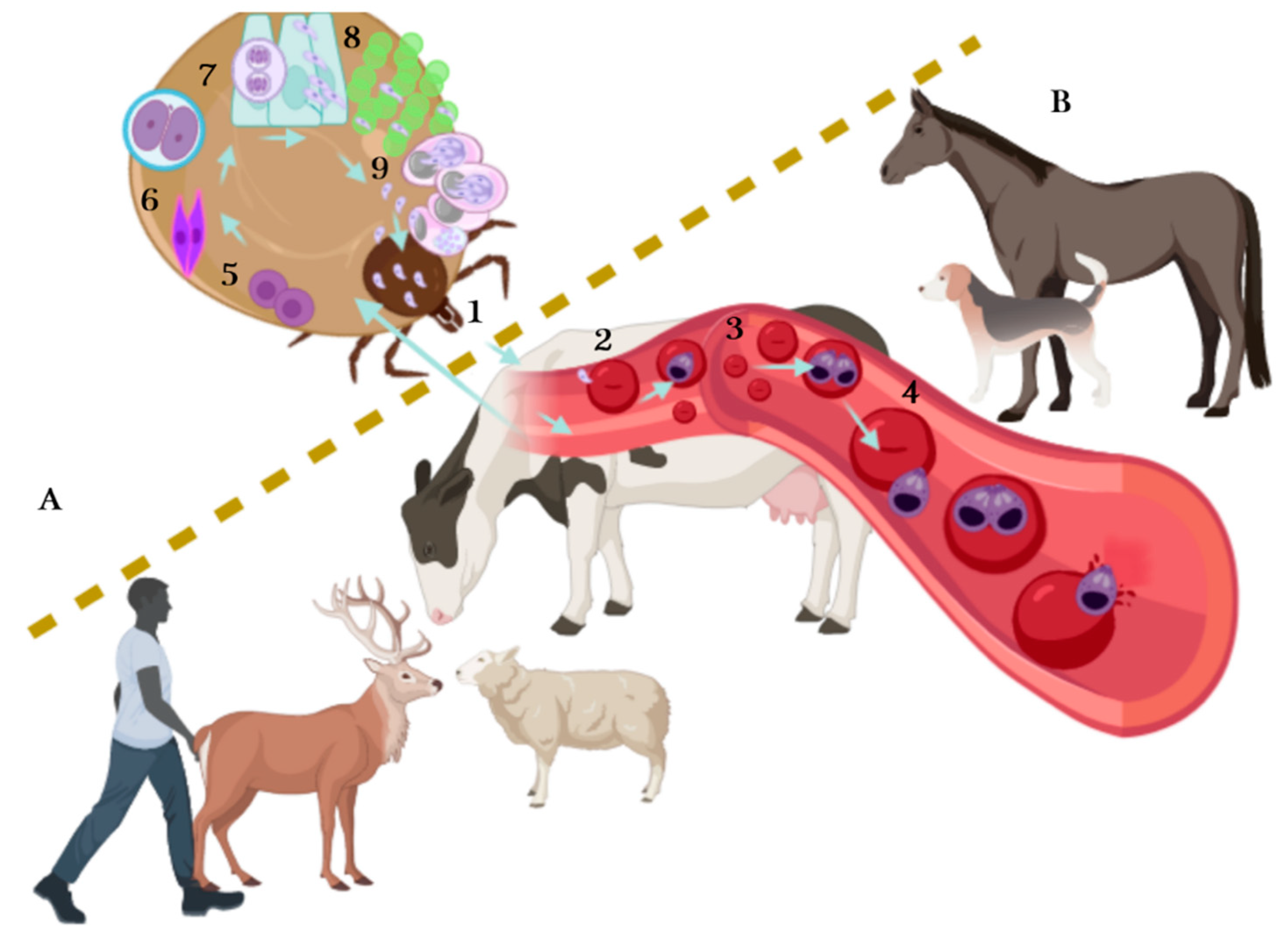{kind=link}
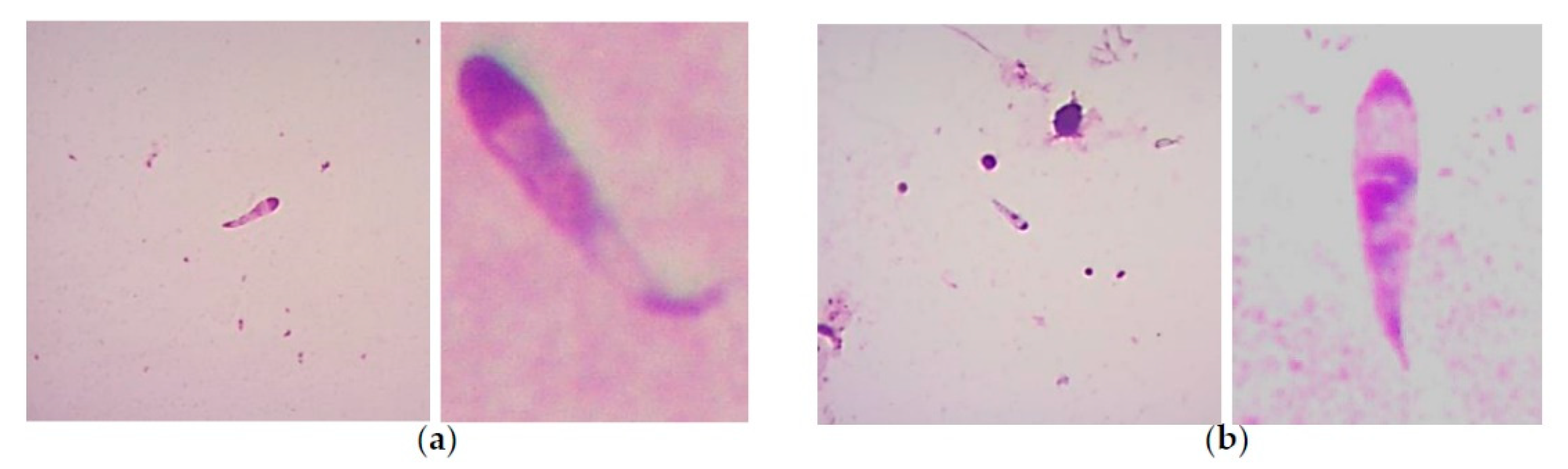{kind=link}
| Babesia Species | Main Tick Vectors | Mammal Host | Geographical Distribution | Detection Methods | Tick Stage Evaluated | References |
|---|---|---|---|---|---|---|
| B. bigemina | Rhipicephalus microplus, Rh. decoloratus, Rh. annulatus, Rh. geigyi, Rh. evertsi | Cattle, Water buffalo | Africa, America, Australia | Microscopy, nPCR, PCR, PCR-RFLP, qPCR | Adult, nymphs, larvae, eggs. Hemolymph and eggs | [34,37,40,41,42,43,45,47,48,49,50,51,52] |
| B. bovis | Rhipicephalus microplus, Rh. decoloratus, Rh. annulatus, Rh. geigyi | Cattle, Water buffalo | Africa, America, Australia | Microscopy, nPCR, PCR, PCR-RFLP, qPCR | Adult, nymphs, larvae, eggs. Hemolymph and eggs | [34,35,36,37,40,41,43,45,47,48,49,50,53] |
| B. divergens | Ixodes ricinus, I. persulcatus | Cattle | North West Europe, Great Britain, Ireland, and Spain | PCR, PCR-RLB hybridization | Adult, nymphs, (Salivary glands) larvae, eggs. | [55,113] |
| B. occultans | Hyalomma marginatum | Cattle | Africa | RLB hybridization B. occultans-specific probe | Adult | [60] |
| B. ovata | Haemaphysalis longicornis, Ixodes ovatus | Cattle | China, Japan, Korea, Mongolia, Thailand | PCR | ND | [61] |
| B. canis | Dermacentor reticulatus | Dogs | Asia and Europe | PCR, real-time PCR-based assay, qPCR, PCR-RLB hybridization, B. canis-specific probe | Adult | [69,75,85,86,87,88,89,90,91,93] |
| B. vogeli | Haemaphysalis spp., Rhipicephalus sanguineus, Rh. turanicus, Rh. haemaphysaloides | Dogs | Africa, Asia, Australia, North and South America, Europe | PCR-RFLP, semi-nested PCR, PCR, nPCR, PCR-RLB hybridization, real-time PCR-based assay, Multiplex PCR | Adult, nymphs, larvae, males, and unfed females | [70,71,73,74,78,79,81,82,83,84,91] |
| B. gibsoni | Haemaphysalis spp., Rh. sanguineus, Rh. turanicus, Ixodes ricinus | Dogs | Africa, America, Asia, Australia, Europe | PCR, nPCR, PCR-RLB hybridization, Multiplex PCR | Adult, nymphs, larvae | [73,74,77,78,84] |
| B. venatorum | Ixodes ricinus, I. persulcatus | Dogs, Roe deer, Red deer, Fallow deer, Moose, White-tailed deer, European reindeer | Canada, China, Europe, Mongolia and USA | PCR, microfluidic real-time PCR, PCR-RLB hybridization | Adult, larvae | [77,93,98,99,101,113] |
| B. odocoilei | Ixodes scapularis | White-tailed deer | Canada, USA | semi-nested PCR | Adult, Larvae | [100] |
| B. caballi | Dermacentor spp., D. nuttalli, D. nitens, Hyalomma marginatus, H. truncatum, Otobius megnini | Horses, Donkeys, Mules and Zebras | Africa, America, Asia, Europe | nPCR | Adult | [105,106] |
| B. microti | Ixodes scapularis, I. ricinus | White-footed mouse, Humans | United States, Europe | PCR-RLB hybridization, nPCR, RT-PCR, in vitro culture-PCR | Adult females, Nymphs | [113,114,115,118] |
| B. motasi | Rhipicephalus bursa, Haemaphysalis longicornis, H. punctata, | Goats, Sheep | Korea | Microscopy, PCR | ND | [114,129] |
| B. duncani | Dermacentor albipictus | Mule deer | United States | nPCR, PCR | Adult, larvae | [116] |
| B. ovis | Rhipicephalus bursa | Goats, Sheep | Iran | Microscopy, PCR-RLB, qPCR, PCR | Adult, eggs, larvae | [124,125,128,129,131,133,134] |
| Babesia Species Detected | PCR Format | Target Gene or Region | Primers Name | Product Size | Primer Sequence | References |
|---|---|---|---|---|---|---|
| B. bigemina and B. bovis | nPCR | SpeI-AvaI rap-1 | BiIA/BiIB BiIAN/BiIBN BoF/BoR BoFN/BoRN | 278 bp 170 bp 356 bp 291 bp | 5′-CATCTAATTTCTCTCCATACCCCTCC-3′ 5′-CCTCGGCTTCAACTCTGATGCCAAAG-3′ 5′-CGCAAGCCCAGCACGCCCCGGTGC-3′ 5′-CCGACCTGGATAGGCTGTGTGATG-3′ 5′-CACGAGGAAGGAACTACCGATGTTGA-3′ 5′ CCAAGGAGCTTCAACGTACGAGGTCA 3′ 5′-TCAACAAGGTACTCTATATGGCTACC-3′ 5′-CTACCGAGCAGAACCTTCTTCACCAT-3′ | [38,40,41,42,43] |
| B. bovis | nPCR | msa-1 | external forward external reverse internal forward internal reverse | ND 212 bp | 5′-TTCGACCAGACCAAATTGT-3′ 5′-CGCATCAAAAGA CTCAACA-3′ 5′-GCCCTGATCTATTTAATGCA-3′ 5′-CCCCGTATAAACATGCTTC-3′ | [35,36] |
| B. bovis | qRT-PCR | msa-1 | Forward/reverse Probe | 150 bp | 5′-GATGCGTTTGCACATGCTAAG-3′ 5′-TGAGAGCACCGAAGTACCCG-3′ 5′-CACGCTCAAGTAGGAAATTTTGTTAAACCTGGA-3′ | [35,36] |
| B. bovis | nPCR | rap-1 | BoF/BoR BoFN/BoRN | 354 bp 291 bp | 5′-CACGAGGAAGGAACTACCGATGTTGA-3′ 5′ CCAAGGAGCTTCAACGTACGAGGTCA 3′ 5′-TCAACAAGGTACTCTATATGGCTACC-3′ 5′-CTACCGAGCAGAACCTTCTTCACCAT-3′ | [53] |
| B. bigemina and B. bovis | nPCR | 18S rRNA | KB-16/KB-17 KB-18/KB-19 KB-24/KB-25 | ND 262 bp 217 bp | 5′-CATCAGCTTGACGGTAGGG-3′ 5′-GTCCTTGGCAAATGCTTTC-3′ 5′-GATGTACAACCTCACCAGAGTACC-3′ 5′-CAACAAAATAGAACCAAGGTCCTAC-3′ 5′-GGGGGCGACCTTCAC-3′ 5′-CTCAATTATACAGGCGAAAC-3′ | [44] |
| B. bigemina and B. bovis | PCR | ssrDNA | A/B C/B | 118 bp 225 bp | 5′-TGTCCTCGTTTGCTTCTTAGAGGGACTCCT-3′ 5′-CCGACACGATGCACACTAAACATTACCCAA-3′ 5′-TTGGCATGGGGGCGACCTTCACCCTCGCCC-3′ 5′-CCAAAGTCAACCAACGGTACGACAGGGTCA-3′ | [45] |
| B. bigemina | qPCR | 18S rDNA | RTBbF/RTBbR | ND | 5′- AGCTTGCTTTCACAACTCGCC -3′ 5′- TTGGTGCTTTGACCGACGACAT -3′ | [51] |
| B. bigemina | qPCR | 18S rDNA | Forward/reverse | ND | 5′- AATAACAATACAGGGCTTTCGTCT -3′ 5′- AACGCGAGGCTGAAATACAACT -3′ | [52] |
| B. bovis, and B. bigemina | PCR-RFLP | 18S rDNA | PiroA/Piro B RE: MspI BoxI | 400 bp 250, 150 bp 290, 110 bp | 5′-AATACCCAATCCTGACACAGGG-3′ 5′-TTAAATACGAATGCCCCCAAC-3′ | [47] |
| B. bigemina and B. bovis | qPCR | mitochondrialcytochrome b | Cbisg 1 and 2 Cbosg 1 and 2 | 88 bp 88 bp | 5′-TGTTCCAGGAGATGTTGATTC-3′ 5′-AGCATGGAAATAACGAAGTGC-3 5′-TGTTCCTGGAAGCGTTGATTC-3′ 5′-AGCGTGAAAATAACGCATTGC-3′ | [49,50] |
| B. divergens | PCR | 18S rRNA | BAB GF2/GR2 | 559 bp | 5′-GYYTTGTAATTGGAATGATGG-3′ 5′-CCAAAGACTTTGATTTCTCTC-3′ | [55] |
| Bovine Babesia spp. | PCR | 18S rRNA | ND | 422–440 bp | 5′-GTTTCTGMCCCATCAGCTTGAC-3′ 5′-CAAGACAAAAGTCTGCTTGAAAC-3′ | [56] |
| B. divergens | PCR | 18S rRNA | ND | 353 bp | 5′-GTTTCTGMCCCATCAGCTTGAC-3′ 5′-CAATATTAACACCACGCAAAAATTC-3′ | [56] |
| Babesia sp. genotype EU1 | PCR | 18S rRNA | ND | 362 bp | 5′-GTTTCTGMCCCATCAGCTTGAC-3′ 5′-AGACAAGAGTCAATAACTCGATAAC-3′ | [56] |
| B. orientalis | Semi-nested PCR | 18S rRNA | P1/B-R2 B-P2/B-R2 | ND 257 bp | 5′-AACCTGGTTGATCCTGCCAGTAGT-3′ 5′-CACACGCACAACGCTGAA-3′ 5′-TGAGAAACGGCTACCACA-3′ 5′-CACACGCACAACGCTGAA-3′ | [57] |
| B. occultans | PCR/RLB hybridization | 18S rRNA (V4 variable region) | RLB-F2/RLB-R2 Probe | 460 bp | 5′-GACACAGGGAGGTAGTGACAAG-3′ 5′-biotin-CTAAGAATTTCACCTCTGACAGT-3′ 5′-GTGTGCCTCTTTTGGCCCATC-3′ Species-specific containing a C12 amino linker in 5′ | [60] |
| B. ovata | PCR | β-tubulin | ND | ND | 5′-ACACTGTGCATCCTCACCGTCATAT-3′ 5′-CTCGCGGATCTTGCTGATCAGCAGA-3′ | [61] |
| B. vogeli | PCR-RFLP | 18S rRNA | PiroA/PiroB RE: TaqI | 400 bp 203, 171, 26 bp | 5′-AATACCCAATCCTGACACAGGG-3′ 5′-TTAAATACGAATGCCCCCACC-3′ | [46,70,83] |
| B. vogeli | Semi-nested PCR | 18S rRNA | 455-459/793-772 BCV/793-772 | 339 bp 192 bp | 5′-GTCTTGTAATTGGAATGATGGTGAC-3′ 5′-ATGCCCCCAACCGTTCCTATTA-3 5′-GTTCGAGTTTGCCATTCGTT-3′ 5′- ATGCCCCCAACCGTTCCTATTA-3′ | [68,70] |
| B. vogeli | PCR | 18S rRNA | BCV-F/Ba721R | 422 bp | 5′- GTGTTCGAGTTTGCCATTCG-3′ 5′-CCCAGAACCCAAAGACTTTGATTTCTCTCAAG-3′ | [79] |
| B. vogeli | PCR-RFLP | 18S rRNA | BJ1/BN2 RE: ApoI | 489 bp 367, 122 bp | 5′-GTCTTGTAATTGGAATGATGG-3′ 5′-TAGTTTATGGTTAGGACTACG-3′ | [71] |
| B. vogeli and B. canis | PCR | cox1 | BFor/BvRev/BcRev | 450 bp 750 bp | 5′-GCATCTGGAATAGCTAGTGC-3′ 5′-CTGCTTCTAAACCAGAAGTG-3′ 5′-TGGAAATGACCTACAACATAC-3′ | [75] |
| B. gibsoni | PCR | 18S rRNA | PiroA/PiroB | 408 bp | 5′-AATACCCAATCCTGACACAGGG-3′ 5′-TTAAATACGAATGCCCCCACC-3′ | [77] |
| B. gibsoni and B. vogeli | nPCR (non-specific) | 18S ssrRNA | 5-22F/1661R 455-479F/793-722R (generic primers) | 293–338 bp | 5′-GTTGATCCTGCCAGTAGT-3′ 5′-AACCTTGTTACGACTTCTC-3′ 5′-GTCTTGTAATTGGAATGATGGTGAC-3′ 5′-ATGCCCCCAACCGTTCCTATTA-3′ | [78] |
| B. gibsoni, B. vogeli, B. canis | PCR/RLB hybridization | 18S rRNA (V4 variable region) | RLB-F2/RLB-R2 Probes | 460 bp | 5′-GACACAGGGAGGTAGTGACAAG-3′ 5′-biotin-CTAAGAATTTCACCTCTGACAGT-3′ Oligonucleotides probes (species-specific) linked to N-terminal N-(trifluoracetamidohexyl-cyanoethyl, N, N-diisopropyl phosphoramidite [TFA])-C6 amino | [81,82,84,86] |
| B. gibsoni and B. vogeli | Multiplex PCR | 18S rRNA | BAGI F/BAGI R BAB1 F/BAB4 R | 590 bp 488 bp | 5′- TTGGCGGCGTTTATTAGTTC-3′ 5′- AAAGGGGAAAACCCCAAAAG-3′ 5′- GTGAACCTTATCACTTAAAGG-3′ 5′- CAACTCCTCCAC GCAATCG-3′ | [73,74] |
| B. canis | PCR | 18S rRNA | BcW-A/BcW-B | 500 bp | 5′-CATCTAAGGAAGGCAGCAGG-3′ 5′- TTAATGGAAACGTCCTTGGC-3′ | [87] |
| B. canis | PCR | 18S rRNA | PiroA/PiroB | 408 bp | 5′-AATACCCAATCCTGACACAGGG-3′ 5′-TTAAATACGAATGCCCCCACC-3′ | [69] |
| B. canis | PCR (non-specific) | 18S rRNA | BJ1/BN2 (generic primers) | ND | 5′-GTCTTGTAATTGGAATGATGG-3′ 5′-TAGTTTATGGTTAGGACTACG-3′ | [72,88,89,90] |
| B. canis | PCR nPCR | 18S rRNA | BS1/PiroC PiroA/PiroC | ND | 5′-GACGGTAGGGTATTGGCCT-3′ 5′-CCAACAAAATAGAACCAAAGTCCTAC-3′ 5′-ATTACCCAATCCTGACACAGGG- 3′ 5′-CCAACAAAATAGAACCAAAGTCCTAC-3′ | [92,93] |
| B. venatorum | PCR (non-specific) | 18S rRNA | BJ1/BN2 (generic primers) | 411–452 bp | 5′-GTCTTGTAATTGGAATGATGG-3′ 5′-TAGTTTATGGTTAGGACTACG-3′ | [72,98,99] |
| B. odocoilei | Semi-nested PCR | 18S rDNA | Piro_18S_300F/Piro_18S_1688R Cocci_18S_595F/Piro_18S_1688R | 1393 bp 1147 bp | 5′-GACGGTAGGGTATTGGCCTA-3′ 5′-CGACTTCTCCTTCCTTTAAGTGATAAG-3′ 5′-CCGCGGTAATTCCAGCTCCAAT-3′ 5′-CGACTTCTCCTTCCTTTAAGTGATAAG-3′ | [100] |
| B. caballi | nPCR | BC48 | BC48F1/BC48R3 BC48F11/BC48R31 | 530 bp 430 bp | 5′-ACGAATTCCCACAACAGCCGTGTT-3′ 5′-ACGATTTCGTAAAGCGTGGCCATG-3′ 5′-GGGCGACGTGACTAAGACATG-3′ 5′-GTTCTCAATGTCAGTGACATCCGC-3′ | [105,106] |
| B. caballi | PCR/RLB hybridization | 18S rRNA (hypervariable V4 region) | RLB-F2/RLB-R2 Genotypes A Genotypes B | ND | 5′-GACACAGGGAGGTAGTGACAAG-3′ biotin-5′-CTAAGAATTTCACCTCTGACAGT-3′ 5′-GTTGCGTTGTTCTTGCTTTTTGCTT-3′ 5′-CGGGTTATTGACTTCGCTTTTTCTT-3′ | [109] |
| B. divergens/B. venatorum/B. microti | PCR/RLB hybridization | 18S rRNA | Bath-F/Bath-R (generic primers) | ND | 5′-TAAGAATTTCACCTCTGACAGTTA-3′ 5′-ACACAGGGAGGTAGTGACAAG-3′ | [113] |
| B. motasi | PCR (non-specific) | 18S rRNA | BTH 1F/BTH 1R GF2F/GR2R (generic primers) | 561 bp | 5′-CCTGAGAAACGGCTACCACATCT-3′ 5′-TTGCGACCATACTCCCCCCA-3′ 5′-GTCTTGTAATTGGAATGATG-3′ 5′-CCAAAGACTTTGATTTCTCTC-3′ | [114] |
| B. motasi | PCR (non-specific) | Cytochrome b (COB) | COB F/COB R (generic primers) | 550 bp | 5′-CCATAGCAATTAATCCAGCTA-3′ 5′-ACCTTGGTCATGGTATTCTGG-3′ | [114] |
| B. motasi | PCR (non-specific) | Cytochrome c (COX-3) | COX3 F/COX3 R (generic primers) | 552 bp | 5′-TCAACAAAATGCCAATATGT-3′ 5′-AAGTGCATCTTTGGGAGAAG-3′ | [114] |
| B. microti | nPCR | β-tubulin | Tubu93 F/Tubu897 R Tubu192 F/Tubu782 R | 551 bp | 5′-GAYAGYCCCTTRCAACTAGAAAGAGC-3′ 5′-CGRTCGAACGAACATTTGTTGHGTCARTTC-3′ 5′-ACHATGGATTCTGTTAGATCYGGC-3′ 5′-GGGAADGGDATRAGATTCACAGC-3′ | [114] |
| B. microti | RT-PCR | 18S rRNA | ND Probe | ND | 5′-AACAGGCATTCGCCTTGAAT-3′ 5′-CCAACTGCTCCTATTAACCATTACTCT-3′ 6FAM-CTACAGCATGGAATAATGA-MGBNFQ | [115] |
| B. duncani | nPCR | β-tubulin | F34/R323 BtubFn/BtubRn | ND 175–181 bp | 5′-TGTGGTAACCAGATYGGWGCCAA-3′ 5′-TCNGTRTARTGNCCYTTRGCCCA-3′ 5′-TCWGACGAGCACGGCATYGA-3′ 5′-CCAGGCTCCAARTCCATYAA-3′ | [116] |
| B. ovis | PCR | ssrRNA | Bbo-F/Bbo-R | 549 bp | 5′-TGGGCAGGACCTTGGTTGTTCT-3′ 5′-CCGCGTAGCGCCGGCTAAATA-3′ | [124,130] |
| B. ovis | qPCR (non-specific) | 18S | Bab_18s_F/Bab_18s_R Bab_18s_P | ND | 5′-TTGGGGGCATTCGTANTNRAC-3′ 5′-TTCTTGATTAATGAAAACGTCTTG-3′ 6FAM-AAGACGAACTACTGCGAAAGCATTTGC-TAMRA | [131] |
| B. ovis | PCR (non-specific) | 18S rRNA | CRYPTOF/CRYPTOR | ND | 5′-AACCTGGTTGATCCTGCCAGT-3′ 5′-GCTTGATCCTTCTGCAGGTTCACCTAC-3′ | [131,132] |
| B. ovis | PCR qPCR | BoSPD BoSPD | SDP forward/SDP reverse SDP forward/SDP reverse | 486 bp 141 bp | 5′-ATGTTGGCCAAGTATCTTGCC-3′ 5′-CTACGTCAATTTGGCCTTGAACTC-3′ 5′-TAATGACGCAGACCTGATGG-3′ 5′-GTTTGATCACCCTCGGAAAC-3′ | [125,133,134] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-García, G.; Santamaría-Espinosa, R.M.; Lira-Amaya, J.J.; Figueroa, J.V. Challenges in Tick-Borne Pathogen Detection: The Case for Babesia spp. Identification in the Tick Vector. Pathogens 2021, 10, 92. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10020092
Martínez-García G, Santamaría-Espinosa RM, Lira-Amaya JJ, Figueroa JV. Challenges in Tick-Borne Pathogen Detection: The Case for Babesia spp. Identification in the Tick Vector. Pathogens. 2021; 10(2):92. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10020092
Chicago/Turabian StyleMartínez-García, Grecia, R. Montserrat Santamaría-Espinosa, José J. Lira-Amaya, and Julio V. Figueroa. 2021. "Challenges in Tick-Borne Pathogen Detection: The Case for Babesia spp. Identification in the Tick Vector" Pathogens 10, no. 2: 92. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10020092