
Antibiotics as a Stressing Factor Triggering the Harboring of Helicobacter pylori J99 within Candida albicans ATCC10231

 ,
, 
 , ,
, ,
Abstract
:1. Introduction
2. Results
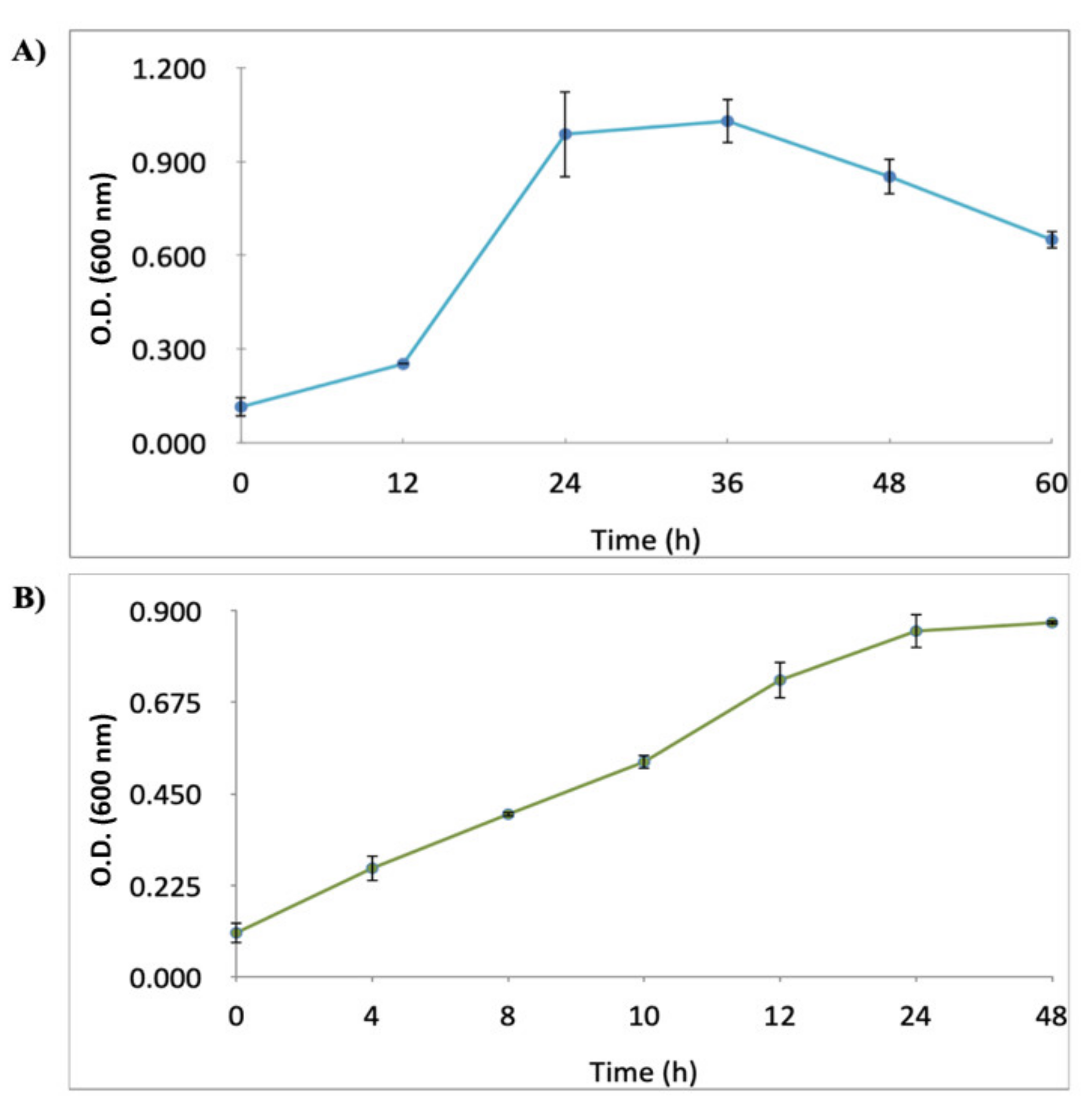
2.1. Growth Curves of H. pylori J99 (ATCC 700824) Strain and C. albicans ATCC10231 Strain
2.2. H. pylori J99-C. albicans ATCC 10231 Co-Culture Assay in the Presence of a Subinhibitory Concentration (¼ Minimal Inhibitory Concentration MIC = 0.0075 μg mL−1) of AMX or CLT
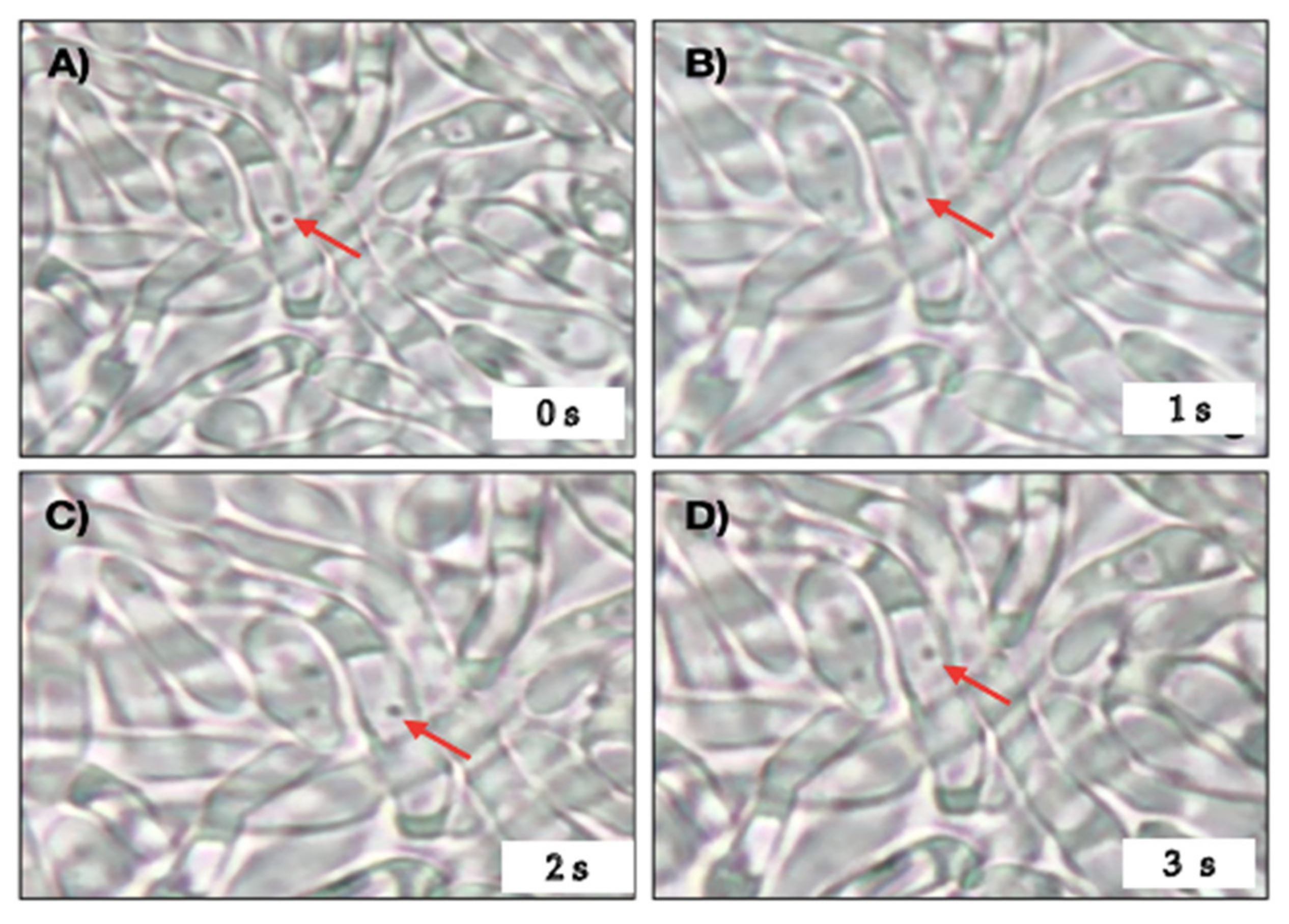
2.3. Cell Viability Assay
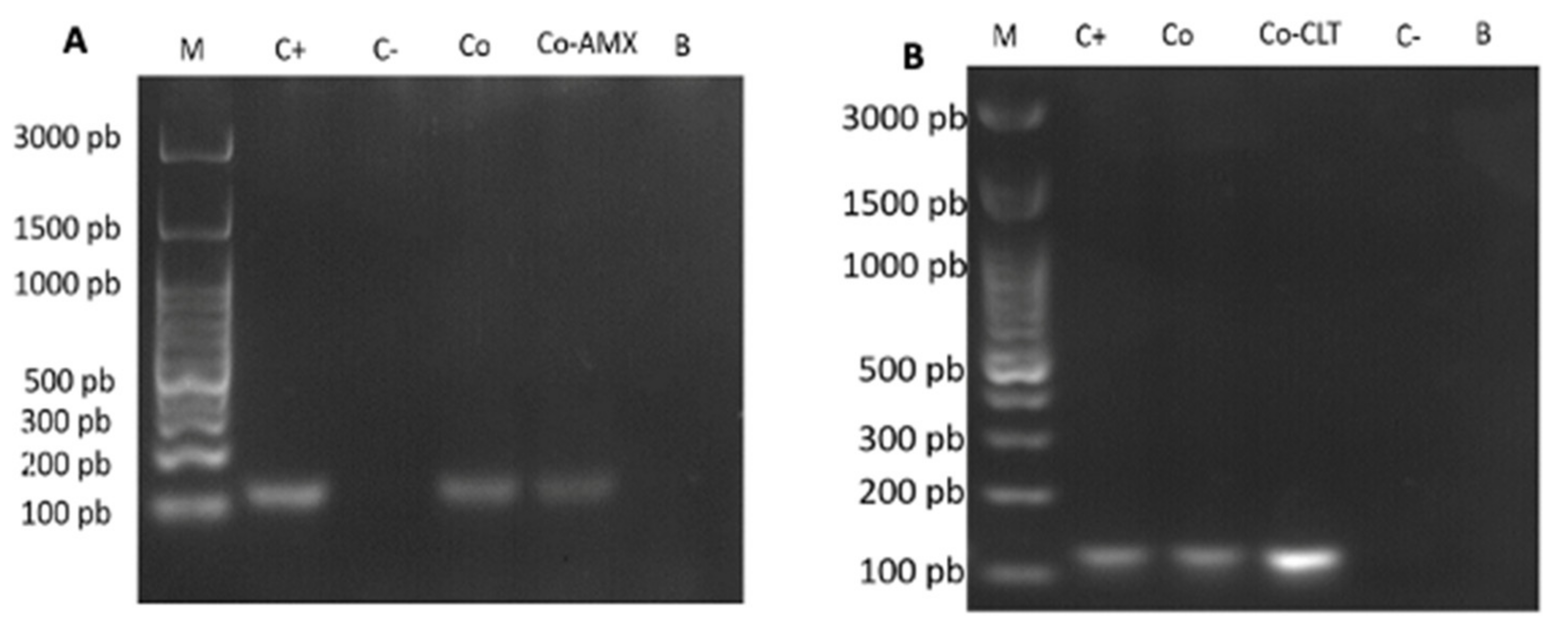
2.4. Detection of H. pylori Genes in Total DNA Extracted from C. albicans Cells
3. Discussion
4. Materials and Methods
4.1. Strains and Culture Conditions
4.2. Growth Curves of H. pylori J99 and C. albicans ATCC 10231 Strains
4.3. Preparation of Antibiotics Stock Solutions
4.4. Determination of the MIC of Amoxicillin and Clarithromycin for H. pylori J99
4.5. Determination of H. pylori J99 Internalization into C. albicans ATCC 10231 Cells in the Presence of Amoxicillin or Clarithromycin
4.6. Cell Viability Assay
4.7. Detection of H. pylori Genes in Total DNA Extracted from C. albicans Cells
4.8. Extraction and Quantification of DNA from Yeasts
4.9. Amplification of the 16S rDNA Gene of H. pylori J99
4.10. Agarose gel Electrophoresis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abadi, A.T.B.; Ierardi, E. Vonoprazan and Helicobacter pylori Treatment: A Lesson from Japan or a Limited Geographic Phenomenon? Front. Pharm. 2019, 10, 316. [Google Scholar] [CrossRef] [Green Version]
- IARC Working Group on the Evaluation of Carcinogenic Risks to Humans. Schistosomes, Liver Flukes and Helicobacter pylori. IARC Working Group on the Evaluation of Carcinogenic Risks to Humans; International Agency for Research on Cancer: Lyon, France, 1994. [Google Scholar]
- Hooi, J.K.Y.; Lai, W.Y.; Ng, W.K.; Suen, M.M.Y.; Underwood, F.E.; Tanyingoh, D.; Malfertheiner, P.; Graham, D.Y.; Wong, V.W.S.; Wu, J.C.Y.; et al. Global Prevalence of Helicobacter pylori Infection: Systematic Review and Meta-Analysis. Gastroenterology 2017, 153, 420–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amieva, M.; Peek, R.M. Pathobiology of Helicobacter pylori-Induced Gastric Cancer. Gastroenterology 2016, 150, 64–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burucoa, C.; Axon, A. Epidemiology of Helicobacter pylori Infection. Helicobacter 2017, 22 (Suppl. 1). [Google Scholar] [CrossRef]
- Fischbach, W.; Malfertheiner, P. Helicobacter pylori Infection. Dtsch. Arztebl. Int. 2018, 115, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-H.; Lv, Z.-F.; Zhong, Y.; Liu, D.-S.; Chen, S.-P.; Xie, Y. The Internalization of Helicobacter pylori Plays a Role in the Failure of H. pylori Eradication. Helicobacter 2017, 22. [Google Scholar] [CrossRef]
- Calvet, X. Dealing with Uncertainty in the Treatment of Helicobacter pylori. Adv. Chronic Dis. 2018, 9, 93–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fallone, C.A.; Moss, S.F.; Malfertheiner, P. Reconciliation of Recent Helicobacter pylori Treatment Guidelines in a Time of Increasing Resistance to Antibiotics. Gastroenterology 2019, 157, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Goderska, K.; Agudo Pena, S.; Alarcon, T. Helicobacter pylori Treatment: Antibiotics or Probiotics. Appl. Microbiol. Biotechnol. 2018, 102, 1–7. [Google Scholar] [CrossRef]
- Hu, Y.; Zhu, Y.; Lu, N.-H. Recent Progress in Helicobacter pylori Treatment. Chin. Med. J. Engl. 2020, 133, 335–343. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, A.; Liou, J.-M.; Gisbert, J.P.; O’Morain, C. Review: Treatment of Helicobacter pylori Infection 2019. Helicobacter 2019, 24 (Suppl. 1), e12640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savoldi, A.; Carrara, E.; Graham, D.Y.; Conti, M.; Tacconelli, E. Prevalence of Antibiotic Resistance in Helicobacter pylori: A Systematic Review and Meta-Analysis in World Health Organization Regions. Gastroenterology 2018, 155, 1372–1382.e17. [Google Scholar] [CrossRef] [Green Version]
- Parra-Sepúlveda, C.; Merino, J.S.; Sáez-Carrillo, K.; González, C.; García-Cancino, A. Antibiotic Resistance Surveillance of Helicobacter pylori at the Biobío Region (Chile) in a Decade. Arq. Gastroenterol. 2019, 56, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, Research, and Development of New Antibiotics: The WHO Priority List of Antibiotic-Resistant Bacteria and Tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Kadkhodaei, S.; Siavoshi, F.; Noghabi, K. Mucoid and Coccoid Helicobacter pylori with Fast Growth and Antibiotic Resistance. Helicobacter 2019, 25. [Google Scholar] [CrossRef] [PubMed]
- Reshetnyak, V.I.; Reshetnyak, T.M. Significance of Dormant Forms of Helicobacter pylori in Ulcerogenesis. World J. Gastroenterol. 2017, 23, 4867–4878. [Google Scholar] [CrossRef]
- Krzyżek, P.; Grande, R. Transformation of Helicobacter pylori into Coccoid Forms as a Challenge for Research Determining Activity of Antimicrobial Substances. Pathogens 2020, 9, 184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matamala-Valdés, L.; Sánchez-Alonzo, K.; Parra, C.; Sáez, K.; Aguayo-Reyes, A.; García, A. Detection of Intracellular Helicobacter pylori in Candida. SPP from Neonate Oral Swabs. Rev. Assoc. Med. Bras. 2018, 64, 928–935. [Google Scholar] [CrossRef]
- Saniee, P.; Siavoshi, F.; Nikbakht Broujeni, G.; Khormali, M.; Sarrafnejad, A.; Malekzadeh, R. Localization of H. pylori within the Vacuole of Candida Yeast by Direct Immunofluorescence Technique. Arch. Iran Med. 2013, 16, 705–710. [Google Scholar]
- Saniee, P.; Siavoshi, F.; Nikbakht Broujeni, G.; Khormali, M.; Sarrafnejad, A.; Malekzadeh, R. Immunodetection of Helicobacter pylori-Specific Proteins in Oral and Gastric Candida Yeasts. Arch. Iran Med. 2013, 16, 624–630. [Google Scholar]
- Siavoshi, F.; Taghikhani, A.; Malekzadeh, R.; Sarrafnejad, A.; Kashanian, M.; Jamal, A.S.; Saniee, P.; Sadeghi, S.; Sharifi, A.H. The Role of Mother’s Oral and Vaginal Yeasts in Transmission of Helicobacter pylori to Neonates. Arch. Iran Med. 2013, 16, 288–294. [Google Scholar] [PubMed]
- Lai, C.-H.; Kuo, C.-H.; Chen, P.-Y.; Poon, S.-K.; Chang, C.-S.; Wang, W.-C. Association of Antibiotic Resistance and Higher Internalization Activity in Resistant Helicobacter pylori Isolates. J. Antimicrob. Chemother. 2006, 57, 466–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Alonzo, K.; Parra-Sepúlveda, C.; Vega, S.; Bernasconi, H.; Campos, V.L.; Smith, C.T.; Sáez, K.; García-Cancino, A. In Vitro Incorporation of Helicobacter pylori into Candida albicans Caused by Acidic PH Stress. Pathogens 2020, 9, 489. [Google Scholar] [CrossRef]
- Angebault, C.; Djossou, F.; Abélanet, S.; Permal, E.; Ben Soltana, M.; Diancourt, L.; Bouchier, C.; Woerther, P.-L.; Catzeflis, F.; Andremont, A.; et al. Candida albicans Is Not Always the Preferential Yeast Colonizing Humans: A Study in Wayampi Amerindians. J. Infect. Dis. 2013, 208, 1705–1716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moris, D.V.; Melhem, M.S.C.; Martins, M.A.; Mendes, R.P. Oral Candida Spp. Colonization in Human Immunodeficiency Virus-Infected Individuals. J. Venom. Anim. Toxins Incl. Trop. Dis. 2008, 14, 224–257. [Google Scholar] [CrossRef]
- Baltrus, D.A.; Guillemin, K. Multiple Phases of Competence Occur during the Helicobacter pylori Growth Cycle. Fems Microbiol. Lett. 2006, 255, 148–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bury-Moné, S.; Kaakoush, N.O.; Asencio, C.; Mégraud, F.; Thibonnier, M.; De Reuse, H.; Mendz, G.L. Is Helicobacter pylori a True Microaerophile? Helicobacter 2006, 11, 296–303. [Google Scholar] [CrossRef]
- Parreira, P.; Magalhães, A.; Gonçalves, I.C.; Gomes, J.; Vidal, R.; Reis, C.A.; Leckband, D.E.; Martins, M.C.L. Effect of Surface Chemistry on Bacterial Adhesion, Viability, and Morphology. J. Biomed. Mater. Res. Part A 2011, 99, 344–353. [Google Scholar] [CrossRef]
- Estibariz, I.; Overmann, A.; Ailloud, F.; Krebes, J.; Josenhans, C.; Suerbaum, S. The Core Genome M5C Methyltransferase JHP1050 (M.Hpy99III) Plays an Important Role in Orchestrating Gene Expression in Helicobacter pylori. Nucleic Acids Res. 2019, 47, 2336–2348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maidan, M.M.; De Rop, L.; Relloso, M.; Diez-Orejas, R.; Thevelein, J.M.; Van Dijck, P. Combined Inactivation of the Candida albicans GPR1 and TPS2 Genes Results in Avirulence in a Mouse Model for Systemic Infection. Infect. Immun. 2008, 76, 1686–1694. [Google Scholar] [CrossRef] [Green Version]
- Thewes, S.; Moran, G.P.; Magee, B.B.; Schaller, M.; Sullivan, D.J.; Hube, B. Phenotypic Screening, Transcriptional Profiling, and Comparative Genomic Analysis of an Invasive and Non-Invasive Strain of Candida albicans. BMC Microbiol. 2008, 8, 187. [Google Scholar] [CrossRef] [Green Version]
- Weerasekera, M.M.; Wijesinghe, G.K.; Jayarathna, T.A.; Gunasekara, C.P.; Fernando, N.; Kottegoda, N.; Samaranayake, L.P. Culture Media Profoundly Affect Candida albicans and Candida tropicalis Growth, Adhesion and Biofilm Development. Mem. Inst. Oswaldo Cruz. 2016, 111, 697–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EUCAST: Clinical Breakpoints and Dosing of Antibiotics. Available online: https://www.eucast.org/clinical_breakpoints/ (accessed on 24 December 2020).
- Donofrio, F.C.; Miranda, E.T.; Maia, D.C.G.; Carlos, I.Z.; Raddi, M.S.G. Subinhibitory Concentrations of Amoxicillin on Helicobacter pylori Increase Apoptosis in RAW 264.7 Cells. J. Chem. Pharm. Res. 2015, 7, 178–181. [Google Scholar]
- Momynaliev, K.; Chelysheva, V.; Selezneva, O.; Larin, A.; Akopian, T.; Alexeev, D.; Le Berre, V.; Sokol, S.; Francois, J.-M.; Govorun, V. Modulation of Helicobacter pylori Transcriptional Profile by Subinhibitory Concentrations of Rifampicin. Biotechnol. Theory Pract. 2013, 23–29. [Google Scholar] [CrossRef]
- Siavoshi, F.; Sahraee, M.; Ebrahimi, H.; Sarrafnejad, A.; Saniee, P. Natural Fruits, Flowers, Honey, and Honeybees Harbor Helicobacter pylori-Positive Yeasts. Helicobacter 2018, 23, e12471. [Google Scholar] [CrossRef] [PubMed]
- Siavoshi, F.; Saniee, P. Vacuoles of Candida Yeast as a Specialized Niche for Helicobacter pylori. World J. Gastroenterol. 2014, 20, 5263–5273. [Google Scholar] [CrossRef] [PubMed]
- Berry, V.; Jennings, K.; Woodnutt, G. Bactericidal and Morphological Effects of Amoxicillin on Helicobacter pylori. Antimicrob. Agents Chemother. 1995, 39, 1859–1861. [Google Scholar] [CrossRef] [Green Version]
- Shahamat, M.; Mai, U.E.; Paszko-Kolva, C.; Yamamoto, H.; Colwell, R.R. Evaluation of Liquid Media for Growth of Helicobacter pylori. J. Clin. Microbiol. 1991, 29, 2835–2837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Attaran, B.; Falsafi, T.; Ghorbanmehr, N. Effect of Biofilm Formation by Clinical Isolates of Helicobacter pylori on the Efflux-Mediated Resistance to Commonly Used Antibiotics. World J. Gastroenterol. 2017, 23, 1163–1170. [Google Scholar] [CrossRef]
- Yonezawa, H.; Osaki, T.; Hanawa, T.; Kurata, S.; Ochiai, K.; Kamiya, S. Impact of Helicobacter pylori Biofilm Formation on Clarithromycin Susceptibility and Generation of Resistance Mutations. PLoS ONE 2013, 8, e0073301. [Google Scholar] [CrossRef] [PubMed]
- Francesco, V.D.; Zullo, A.; Hassan, C.; Giorgio, F.; Rosania, R.; Ierardi, E. Mechanisms of Helicobacter pylori Antibiotic Resistance: An Updated Appraisal. World J. Gastrointest. Pathophysiol. 2011, 2, 35–41. [Google Scholar] [CrossRef]
- Sauvage, E.; Kerff, F.; Terrak, M.; Ayala, J.A.; Charlier, P. The Penicillin-Binding Proteins: Structure and Role in Peptidoglycan Biosynthesis. FEMS Microbiol. Rev. 2008, 32, 234–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bush, K. Antimicrobial Agents Targeting Bacterial Cell Walls and Cell Membranes. Rev. Sci. Tech. 2012, 31, 43–56. [Google Scholar] [CrossRef]
- Chaput, C.; Ecobichon, C.; Cayet, N.; Girardin, S.E.; Werts, C.; Guadagnini, S.; Prévost, M.-C.; Mengin-Lecreulx, D.; Labigne, A.; Boneca, I.G. Role of AmiA in the Morphological Transition of Helicobacter pylori and in Immune Escape. PLoS Pathog. 2006, 2, e97. [Google Scholar] [CrossRef] [Green Version]
- Lambert, T. Antibiotics That Affect the Ribosome. Rev. Sci. Tech. 2012, 31, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Braga, P.C. Effects of Subinhibitory Concentrations of Seven Macrolides and Four Fluoroquinolones on Adhesion of Staphylococcus aureus to Human Mucosal Cells. Chemotherapy 1994, 40, 304–310. [Google Scholar] [CrossRef] [PubMed]
- Shryock, T.R.; Mortensen, J.E.; Baumholtz, M. The Effects of Macrolides on the Expression of Bacterial Virulence Mechanisms. J. Antimicrob. Chemother. 1998, 41, 505–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wozniak, D.J.; Keyser, R. Effects of Subinhibitory Concentrations of Macrolide Antibiotics on Pseudomonas Aeruginosa. Chest 2004, 125, 62S–69S. [Google Scholar] [CrossRef]
- Garey, K.; Vo, Q.; Lewis, R.; Saengcharoen, W.; LaRocco, M.; Tam, V. Increased Bacterial Adherence and Biomass in Pseudomonas aeruginosa Bacteria Exposed to Clarithromycin. Diagn. Microbiol. Infect. Dis. 2008, 63, 81–86. [Google Scholar] [CrossRef]
- Faghri, J.; Poursina, F.; Moghim, S.; Zarkesh Esfahani, H.; Nasr Esfahani, B.; Fazeli, H.; Mirzaei, N.; Jamshidian, A.; Ghasemian Safaei, H. Morphological and Bactericidal Effects of Different Antibiotics on Helicobacter Pylori. Jundishapur J. Microbiol. 2014, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bessa, L.J.; Grande, R.; Iorio, D.D.; Giulio, M.D.; Campli, E.D.; Cellini, L. Helicobacter Pylori Free-Living and Biofilm Modes of Growth: Behavior in Response to Different Culture Media. APMIS 2013, 121, 549–560. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Mesonero, L.; Moreno, Y.; Alonso, J.L.; Ferrús, M.A. DVC-FISH and PMA-QPCR Techniques to Assess the Survival of Helicobacter pylori inside Acanthamoeba castellanii. Res. Microbiol. 2016, 167, 29–34. [Google Scholar] [CrossRef]
- Salmanian, A.-H.; Siavoshi, F.; Beyrami, Z.; Latifi-Navid, S.; Tavakolian, A.; Sadjadi, A. Foodborne Yeasts Serve as Reservoirs of Helicobacter pylori. J. Food Saf. 2012, 32, 152–160. [Google Scholar] [CrossRef]
- Siavoshi, F.; Salmanian, A.H.; Akbari, F.; Kbari, F.A.; Malekzadeh, R.; Massarrat, S. Detection of Helicobacter pylori-Specific Genes in the Oral Yeast. Helicobacter 2005, 10, 318–322. [Google Scholar] [CrossRef]
- Paredes-Osses, E.; Sáez, K.; Sanhueza, E.; Hebel, S.; González, C.; Briceño, C.; García Cancino, A. Association between CagA, VacAi, and DupA Genes of Helicobacter pylori and Gastroduodenal Pathologies in Chilean Patients. Folia Microbiol. Praha 2017, 62, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, N.; Kuwayama, H.; Ito, Y.; Iwasaki, A.; Arakawa, Y. Gastric Epithelial Cells Stimulate Helicobacter pylori Growth. J. Clin. Gastroenterol. 1998, 27 (Suppl. 1), S138–S140. [Google Scholar] [CrossRef] [PubMed]
- Premoli, G.; González, A.; Millán-Mendoza, B.; Percoco, T.; Vielma, A. Diagnóstico de Helicobacter pylori Mediante La Reacción En Cadena de La Polimerasa. Rev. Cuba. Med. Trop. 2004, 56, 85–90. [Google Scholar]
- Sánchez-Alonzo, K.; Parra-Sepúlveda, C.; Vergara, L.; Bernasconi, H.; García-Cancino, A.; Sánchez-Alonzo, K.; Parra-Sepúlveda, C.; Vergara, L.; Bernasconi, H.; García-Cancino, A. Detection of Helicobacter pylori in Oral Yeasts from Students of a Chilean University. Rev. Assoc. Med. Bras. 2020, 66, 1509–1514. [Google Scholar] [CrossRef] [PubMed]


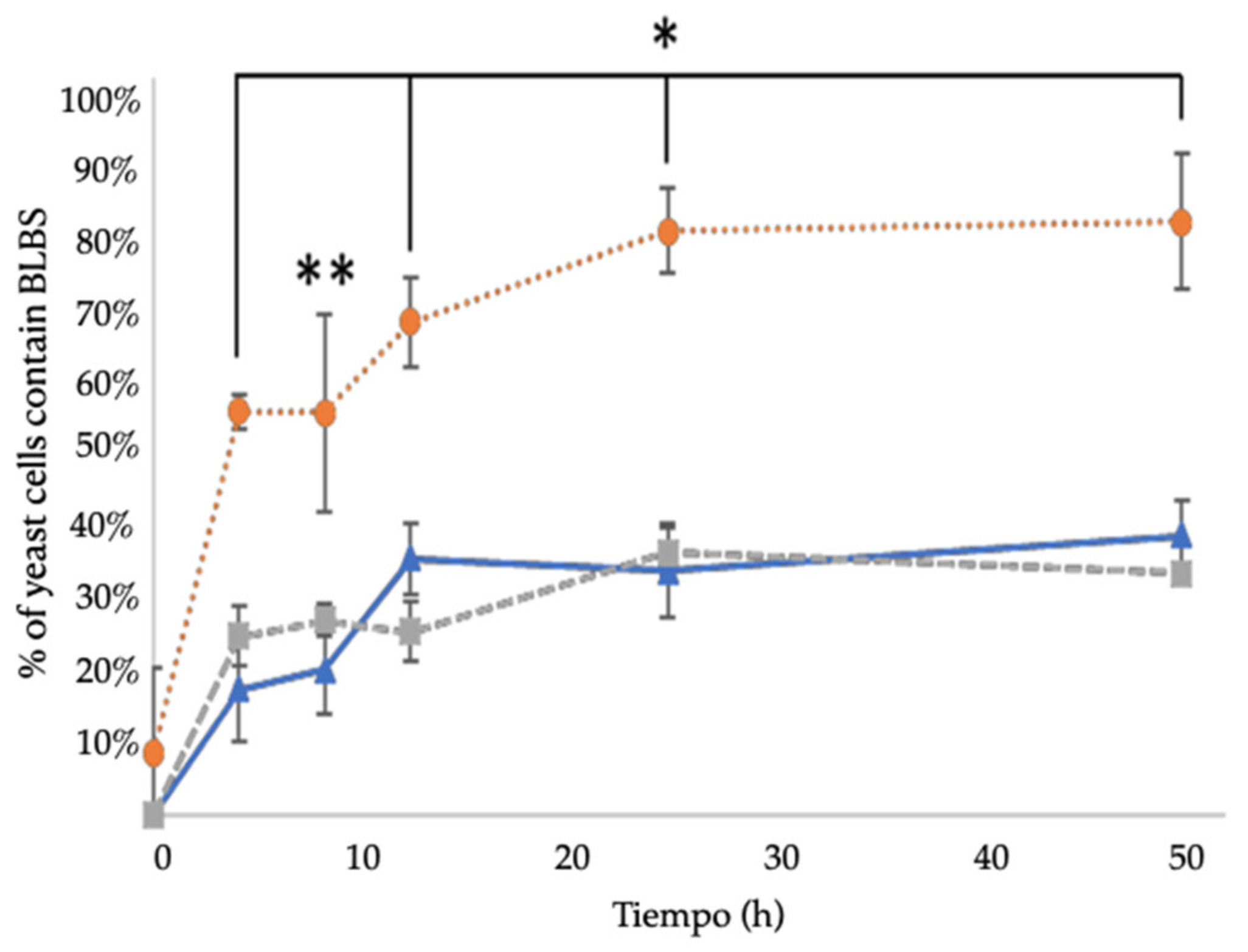
 ), co-culture in the presence of ¼ MIC CLT (
), co-culture in the presence of ¼ MIC CLT (  ), co-culture in the presence of ¼ MIC AMX (
), co-culture in the presence of ¼ MIC AMX (  ). Mean of duplicates. * p < 0.05, ** p < 0.1 with respect to control. BLBs: bacteria-like body MIC: minimal inhibitory concentration, CLT: clarithromycin, AMX: amoxicillin.
), co-culture in the presence of ¼ MIC CLT ( ), co-culture in the presence of ¼ MIC AMX ( ). Mean of duplicates. * p < 0.05, ** p < 0.1 with respect to control. BLBs: bacteria-like body MIC: minimal inhibitory concentration, CLT: clarithromycin, AMX: amoxicillin.
). Mean of duplicates. * p < 0.05, ** p < 0.1 with respect to control. BLBs: bacteria-like body MIC: minimal inhibitory concentration, CLT: clarithromycin, AMX: amoxicillin.
), co-culture in the presence of ¼ MIC CLT ( ), co-culture in the presence of ¼ MIC AMX ( ). Mean of duplicates. * p < 0.05, ** p < 0.1 with respect to control. BLBs: bacteria-like body MIC: minimal inhibitory concentration, CLT: clarithromycin, AMX: amoxicillin.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Media | DE | |
|---|---|---|---|
| Control | 35.4 | 3.39 | B |
| AMX | 46.85 | 2.19 | A |
| CLT | 31.15 | 1.59 | B |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Alonzo, K.; Belmar, L.; Parra-Sepúlveda, C.; Bernasconi, H.; Campos, V.L.; Smith, C.T.; Sáez, K.; García-Cancino, A. Antibiotics as a Stressing Factor Triggering the Harboring of Helicobacter pylori J99 within Candida albicans ATCC10231. Pathogens 2021, 10, 382. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10030382
Sánchez-Alonzo K, Belmar L, Parra-Sepúlveda C, Bernasconi H, Campos VL, Smith CT, Sáez K, García-Cancino A. Antibiotics as a Stressing Factor Triggering the Harboring of Helicobacter pylori J99 within Candida albicans ATCC10231. Pathogens. 2021; 10(3):382. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10030382
Chicago/Turabian StyleSánchez-Alonzo, Kimberly, Libnny Belmar, Cristian Parra-Sepúlveda, Humberto Bernasconi, Víctor L. Campos, Carlos T. Smith, Katia Sáez, and Apolinaria García-Cancino. 2021. "Antibiotics as a Stressing Factor Triggering the Harboring of Helicobacter pylori J99 within Candida albicans ATCC10231" Pathogens 10, no. 3: 382. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10030382