
Impact of Probiotics on the Salivary Microbiota and Salivary Levels of Inflammation-Related Proteins during Short-Term Sugar Stress: A Randomized Controlled Trial

Abstract
:1. Introduction
2. Results
2.1. Background Data
2.2. Sequencing Metadata
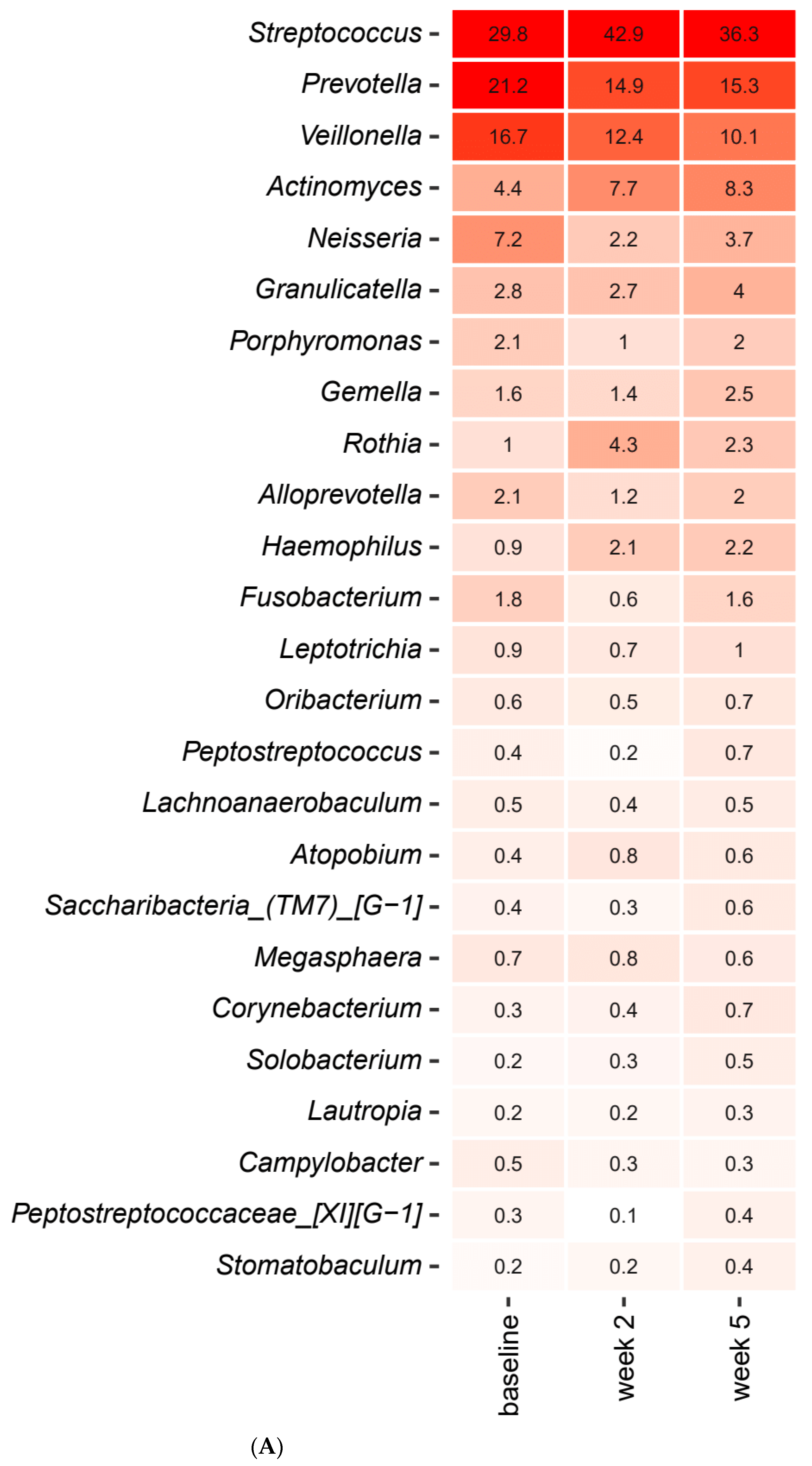
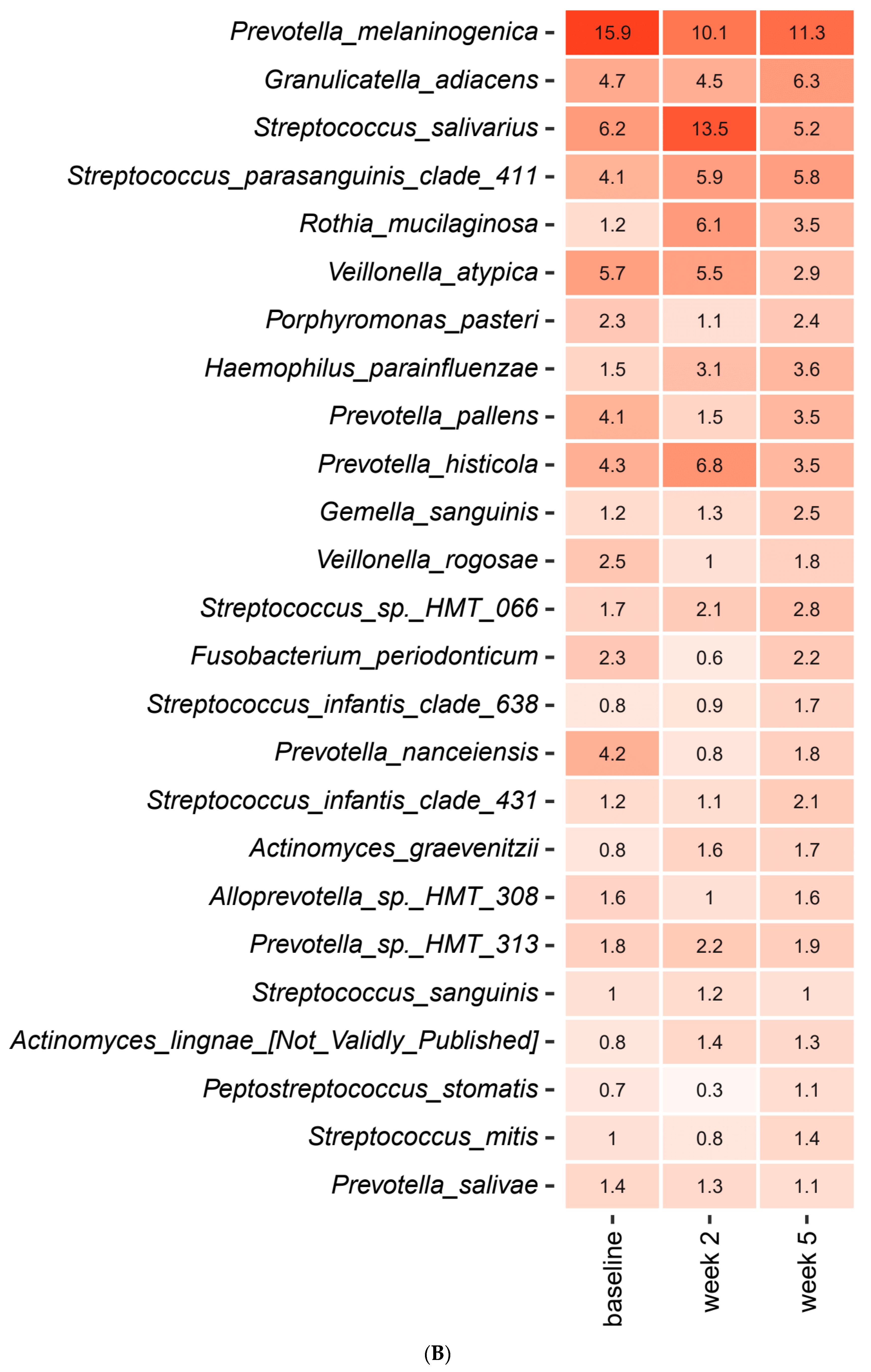
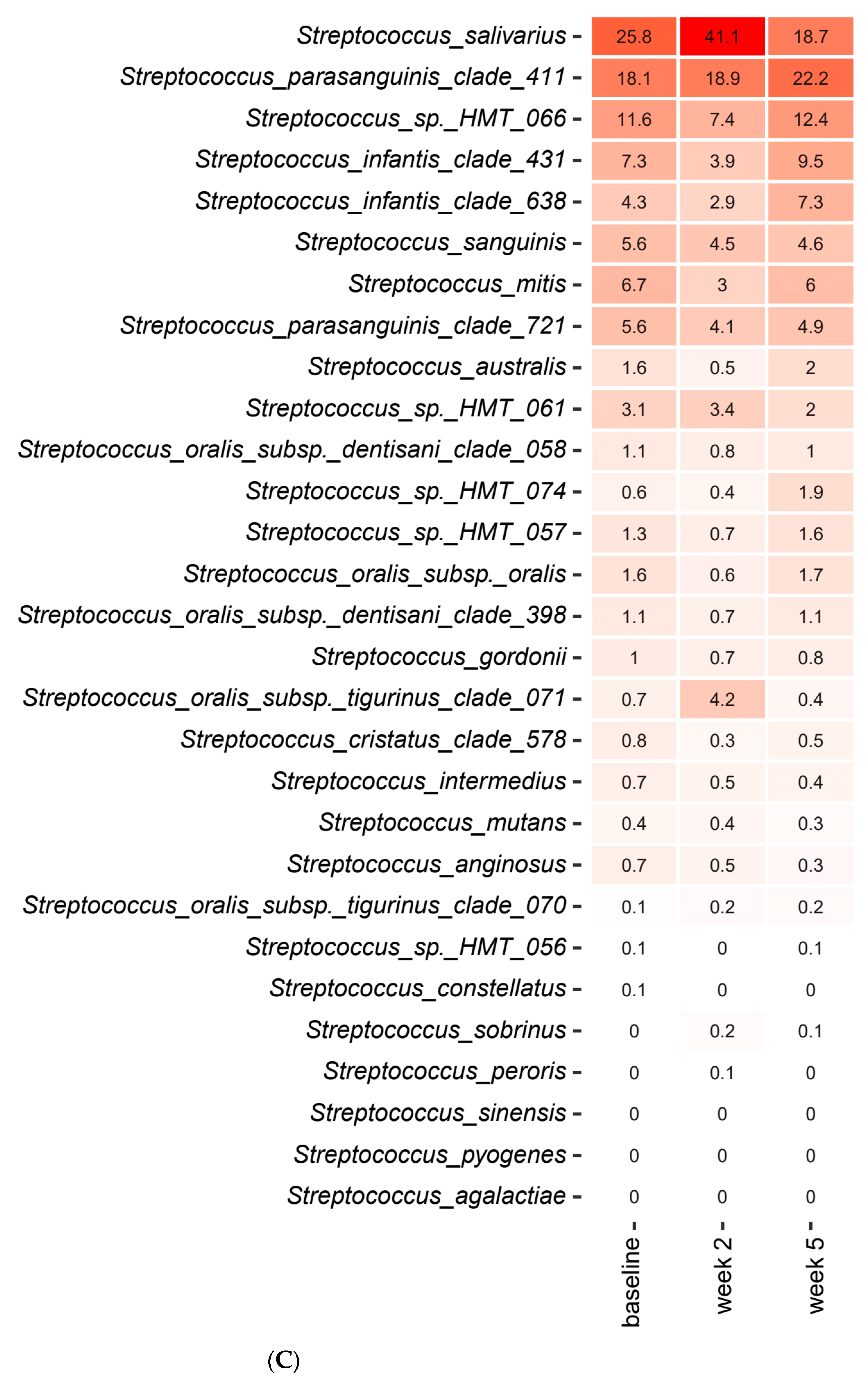
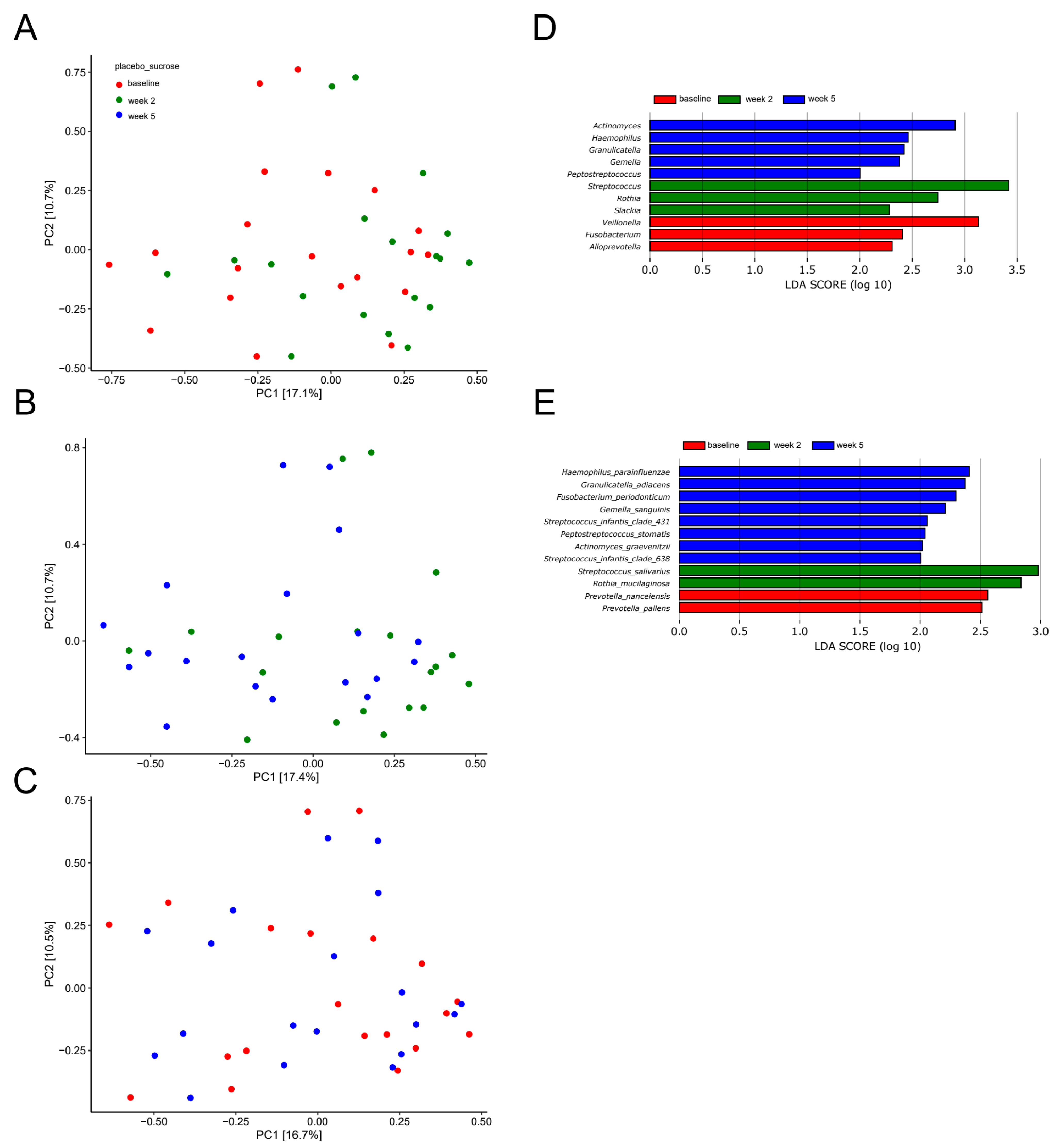
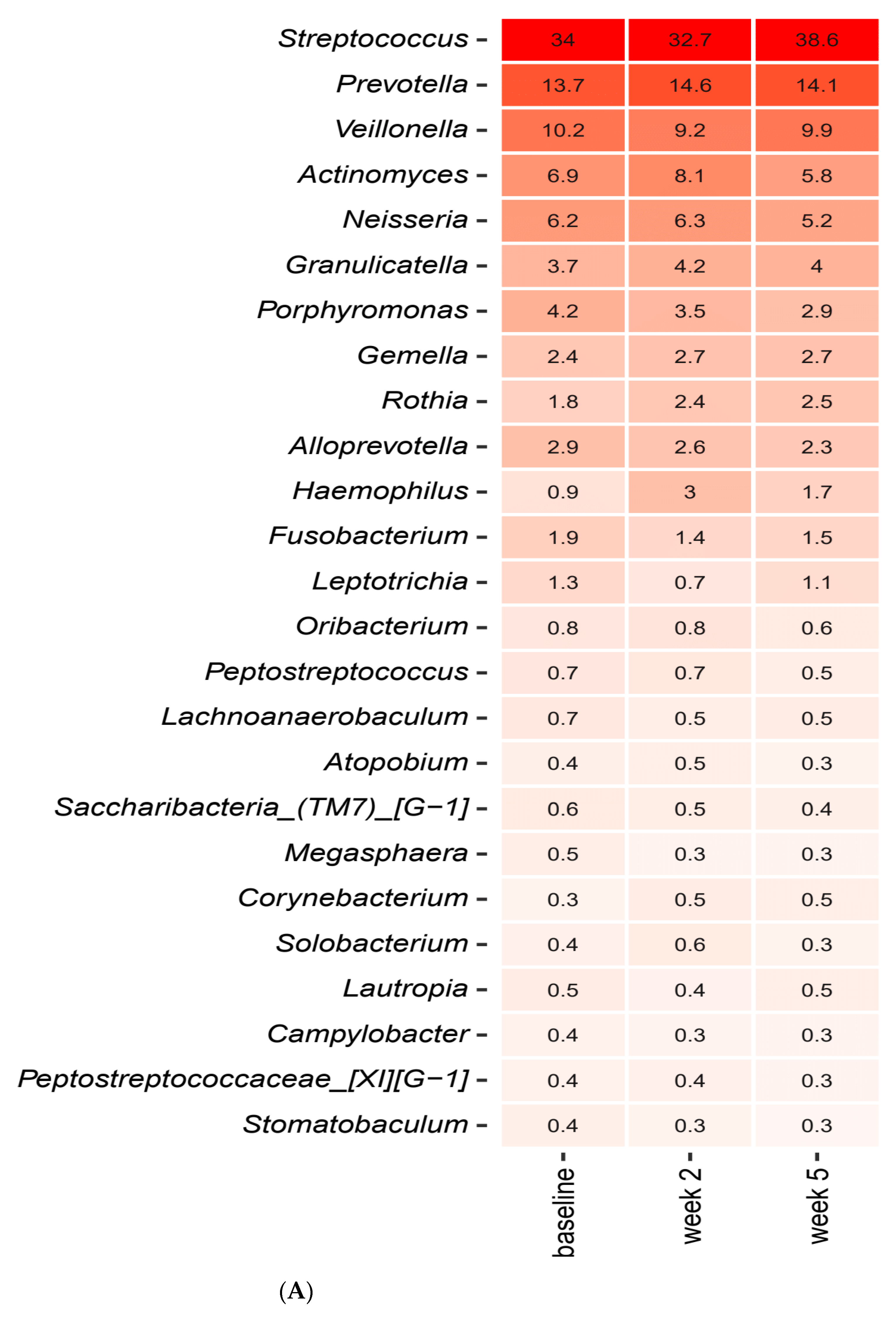
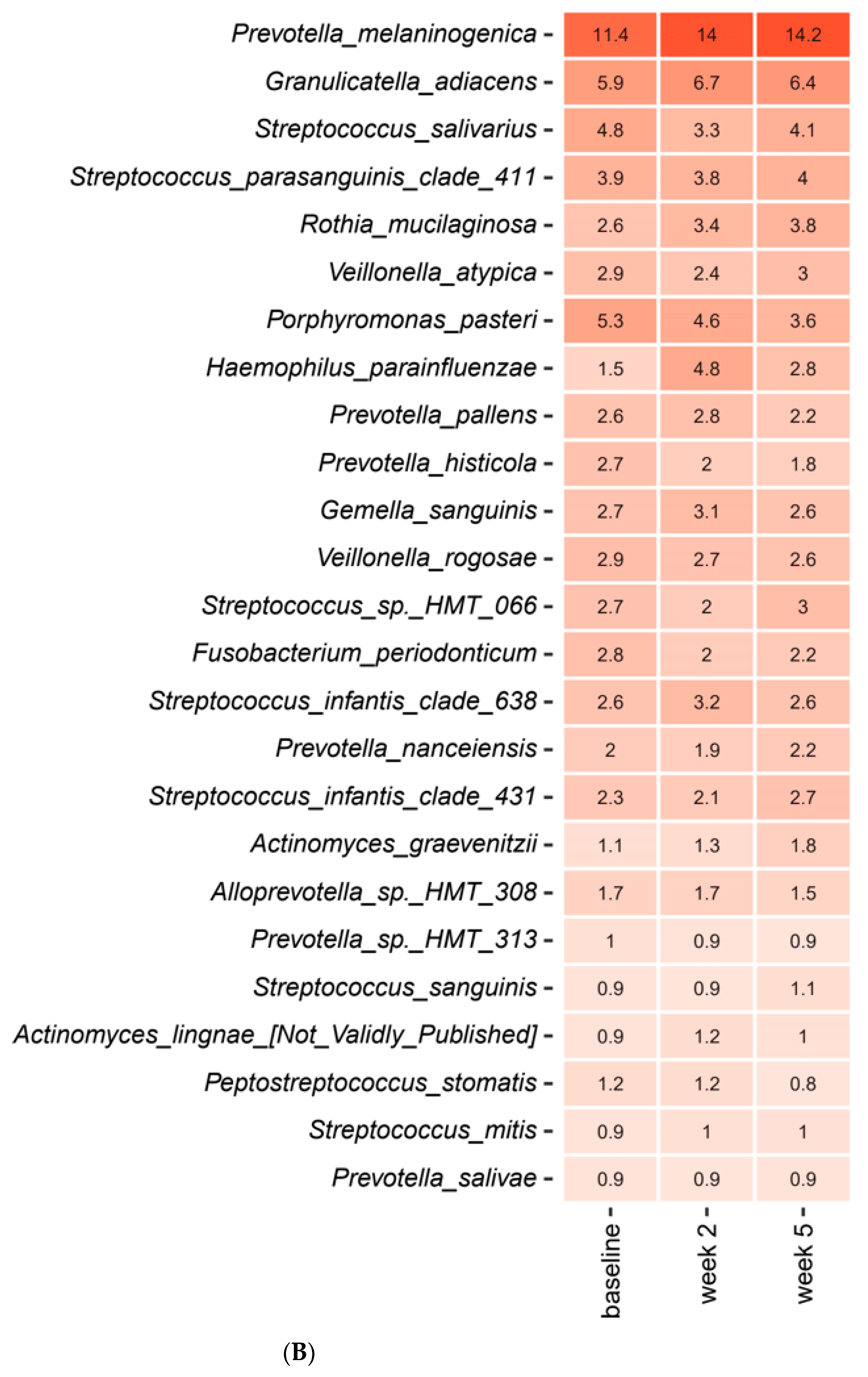
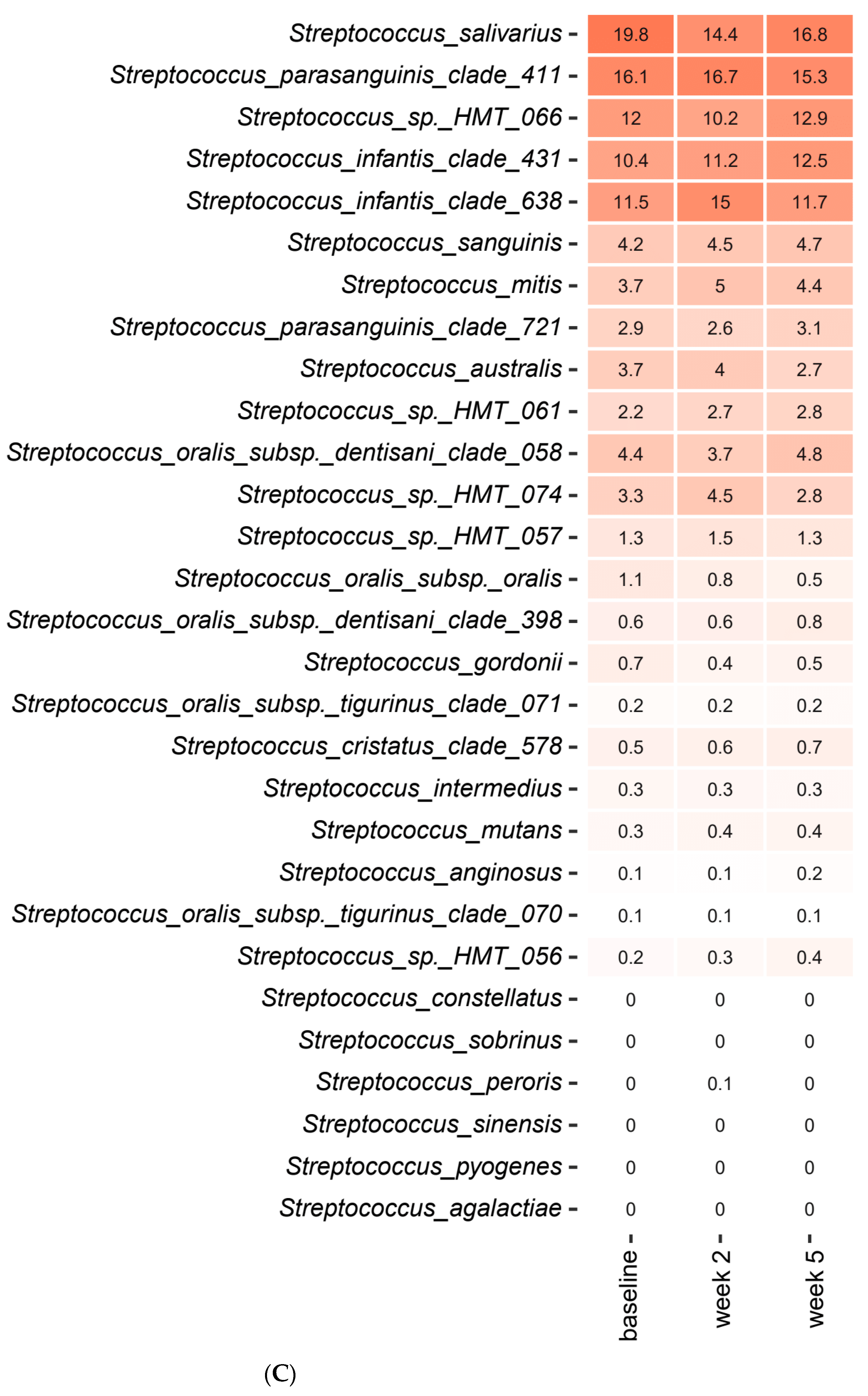
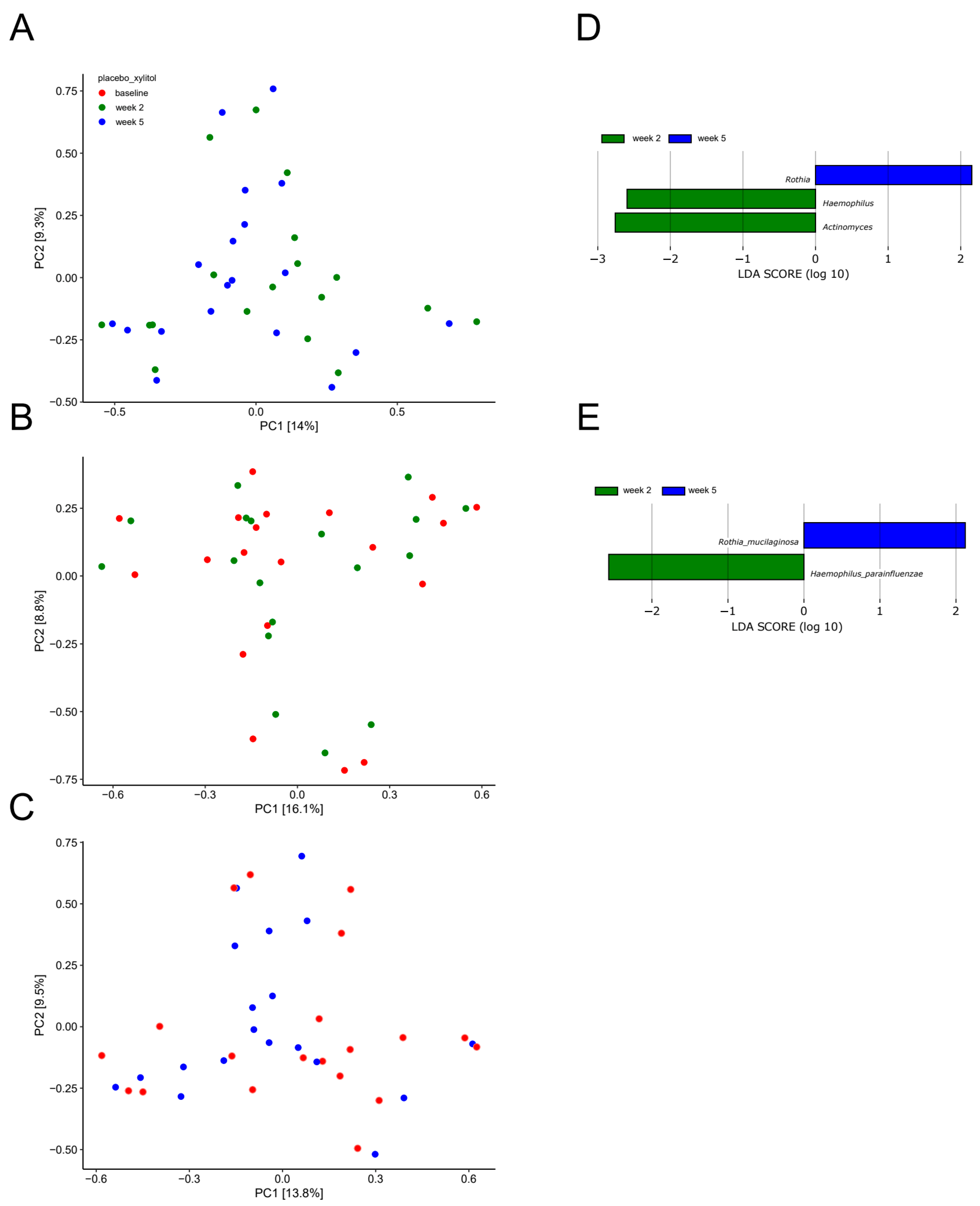
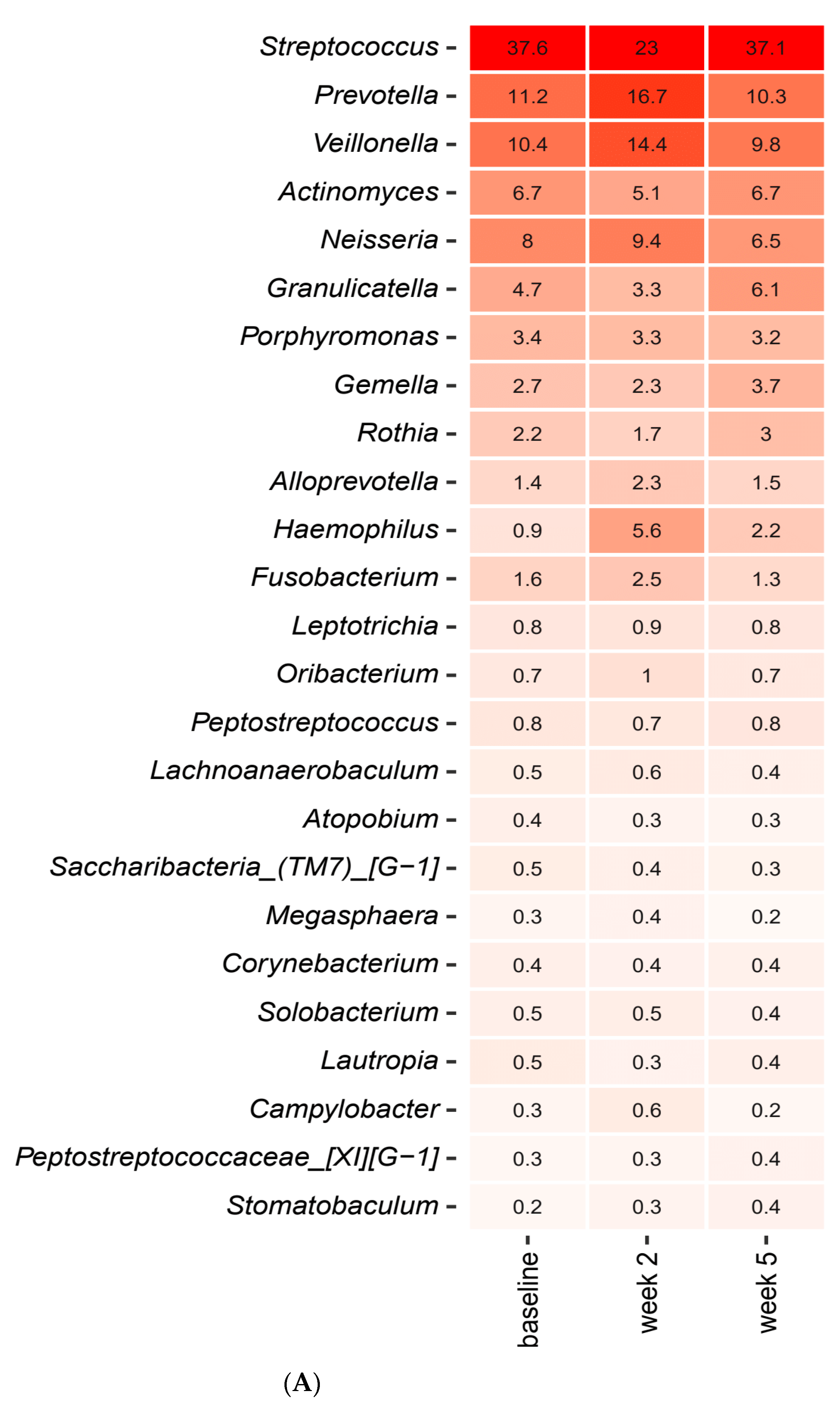
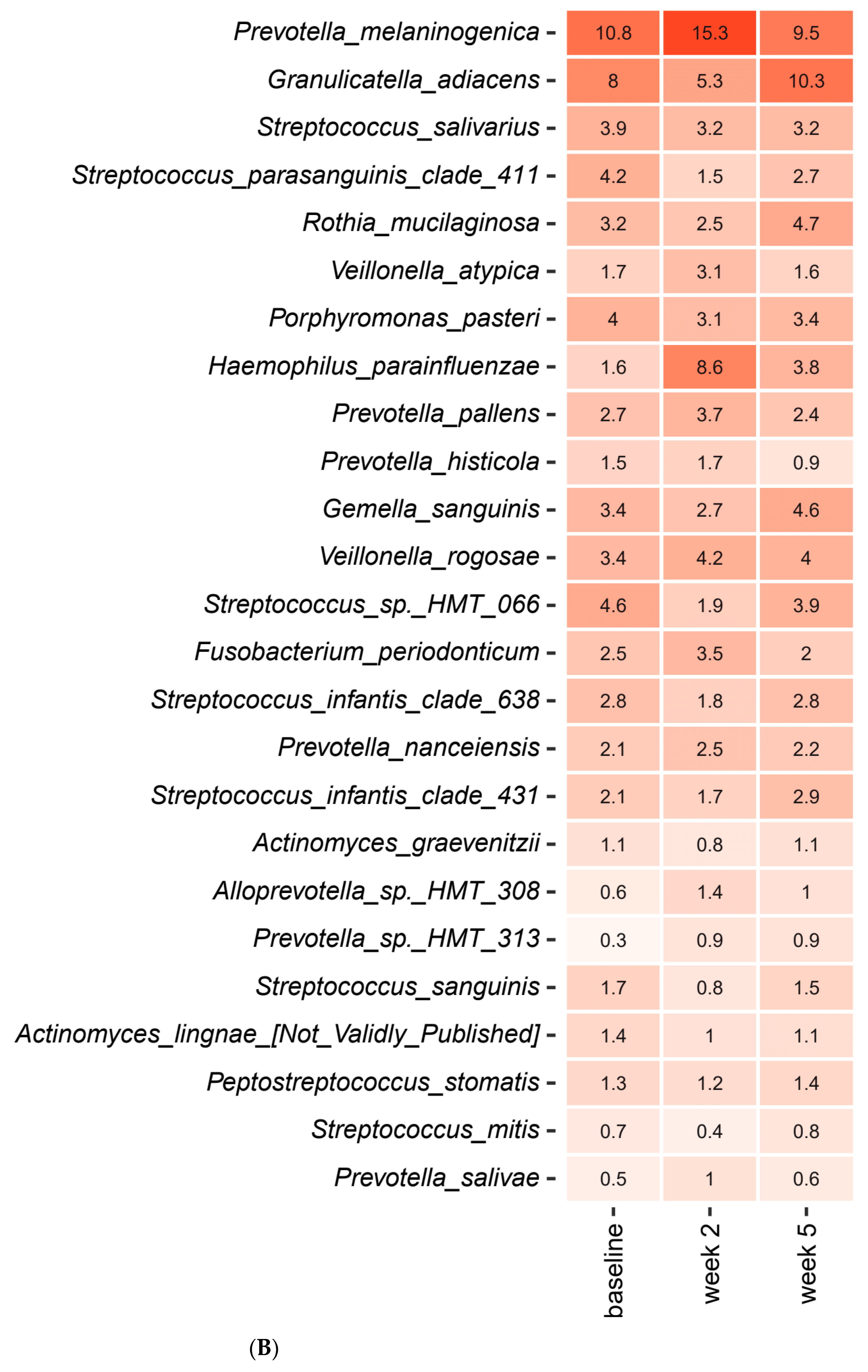
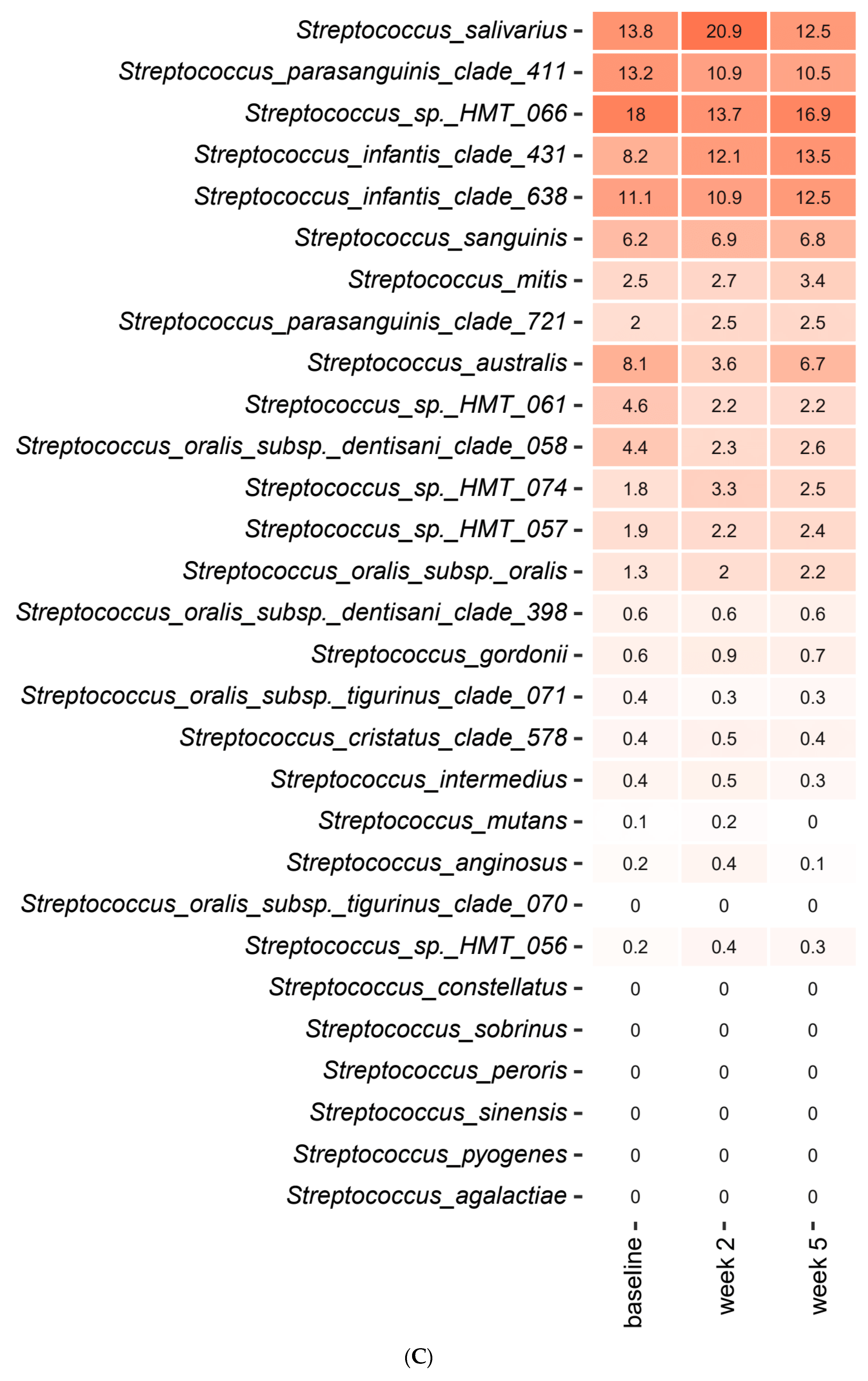
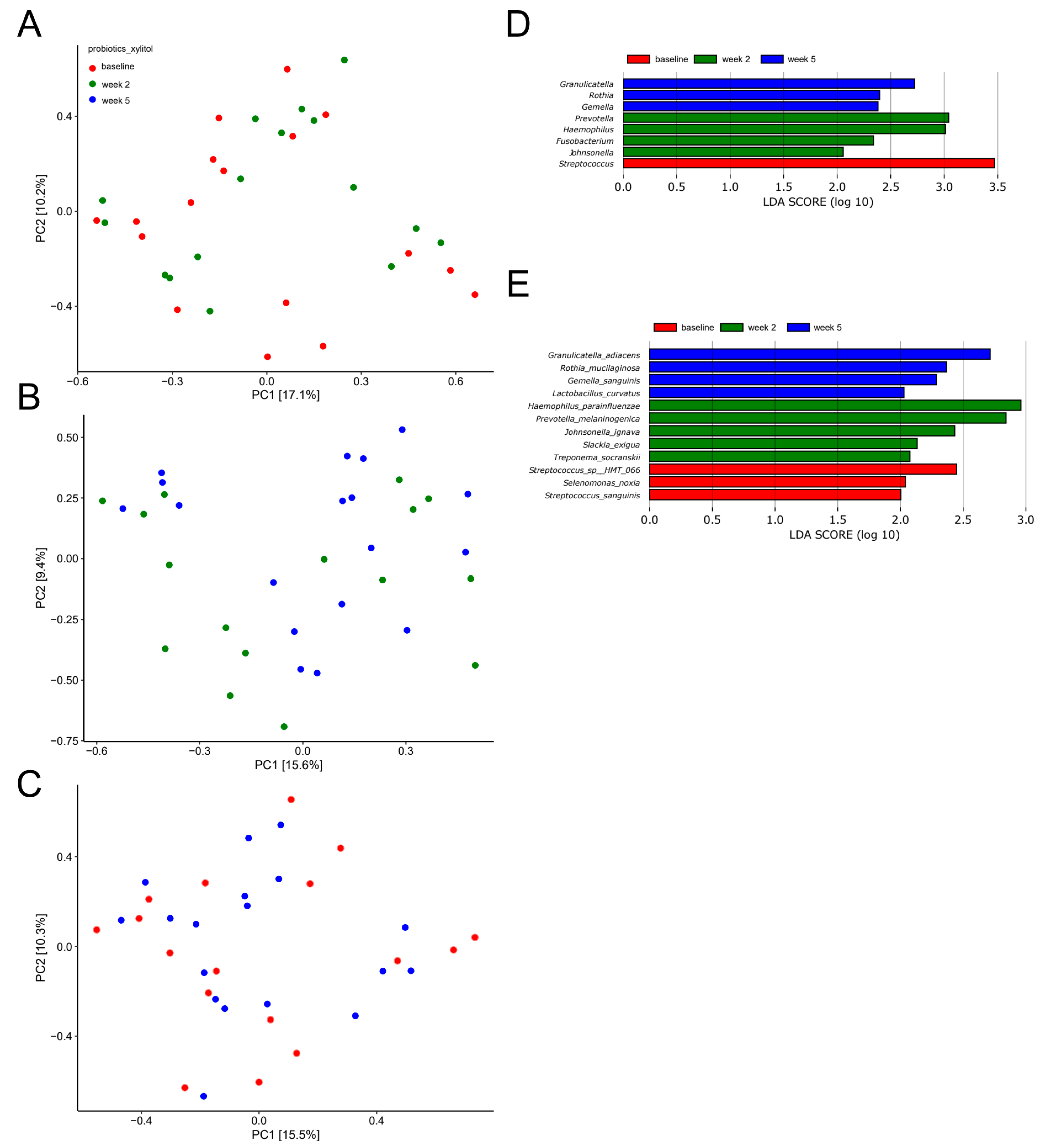
2.3. Short-Term Sugar Stress Changes the Composition of the Salivary Microbiota
2.4. Xylitol Has Minimum Effect on the Composition of the Salivary Microbiota
2.5. Probiotics in Combination with Xylitol Have a Major Impact on the Composition of the Salivary Microbiota
2.6. Impact of Sucrose, Xylitol and Probiotics on Salivary Levels of NGAL and Transferrin
3. Discussion
4. Materials and Methods
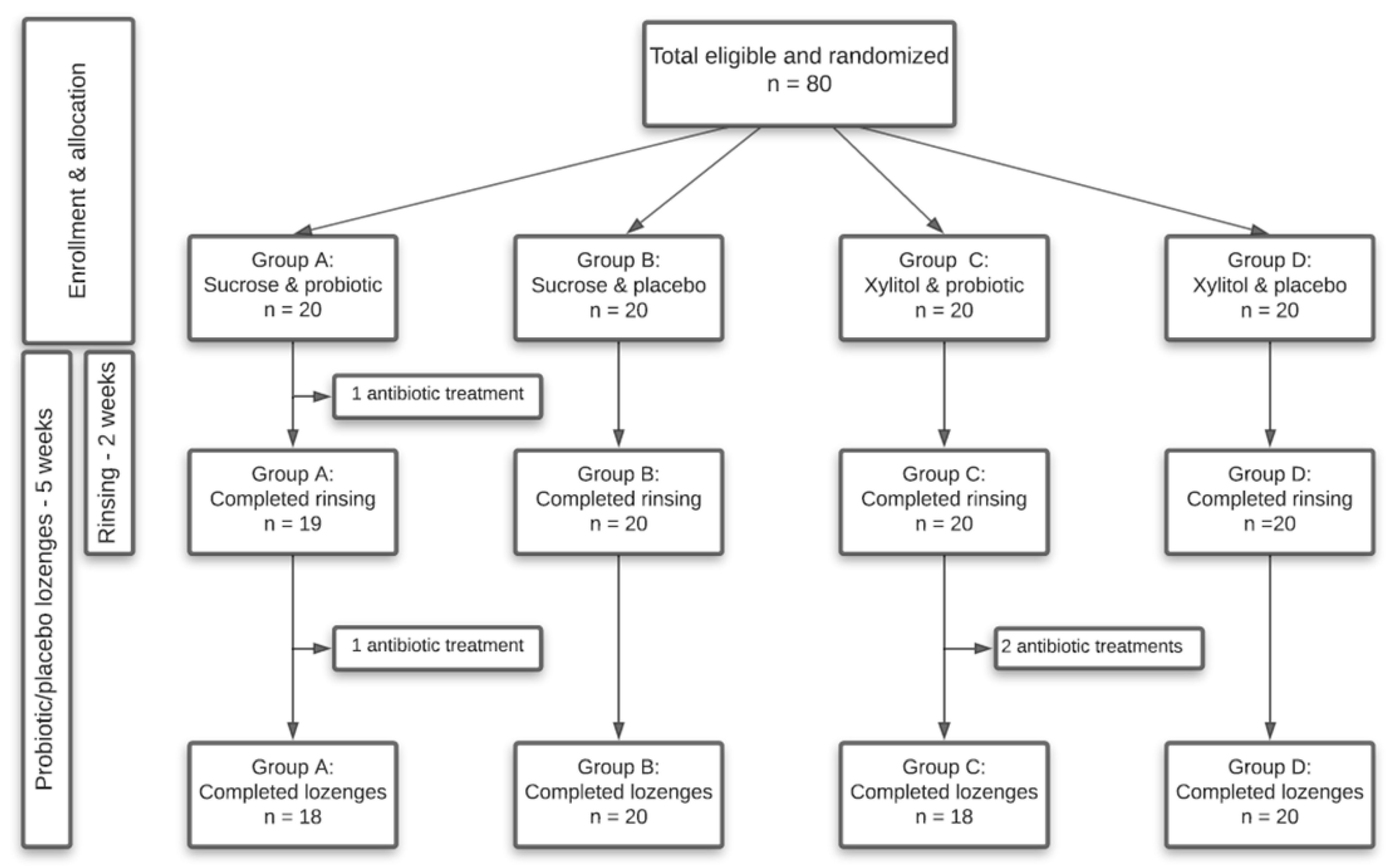
4.1. Study Design
4.2. Study Population
4.3. Collection of Samples
4.4. Sucrose and Xylitol Solution
4.5. Probiotics and Placebo
4.6. DNA Extraction
4.7. Library Preparation
4.8. DNA Sequencing
4.9. Bioinformatic Processing
4.10. Immunological Analysis and NGAL and Transferrin
4.11. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marsh, P. In Sickness and in Health—What Does the Oral Microbiome Mean to Us? An Ecological Perspective. Adv. Dent. Res. 2018, 29, 60–65. [Google Scholar] [CrossRef]
- Kilian, M.; Chapple, I.L.C.; Hannig, M.; Marsh, P.D.; Meuric, V.; Pedersen, A.M.L.; Tonetti, M.S.; Wade, W.G.; Zaura, E. The oral microbiome—An update for oral healthcare professionals. Br. Dent. J. 2016, 221, 657–666. [Google Scholar] [CrossRef]
- Twetman, S. Prevention of dental caries as a non-communicable disease. Eur. J. Oral Sci. 2018, 126, 19–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitts, N.B.; Zero, D.T.; Marsh, P.D.; Ekstrand, K.; Weintraub, J.A.; Ramos-Gomez, F.; Tagami, J.; Twetman, S.; Tsakos, G.; Ismail, A. Dental caries. Nat. Rev. Dis Primers 2017, 25, 17030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Divaris, K. Searching Deep and Wide: Advances in the Molecular Understanding of Dental Caries and Periodontal Disease. Adv. Dent. Res. 2019, 30, 40–44. [Google Scholar] [CrossRef]
- Mäkinen, K.K. Sugar Alcohols, Caries Incidence, and Remineralization of Caries Lesions: A Literature Review. Int. J. Dent. 2010, 2010, 1–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaura, E.; Twetman, S. Critical Appraisal of Oral Pre- and Probiotics for Caries Prevention and Care. Caries Res. 2019, 53, 514–526. [Google Scholar] [CrossRef] [PubMed]
- Gruner, D.; Paris, S.; Schwendicke, F. Probiotics for managing caries and periodontitis: Systematic review and meta-analysis. J. Dent. 2016, 48, 16–25. [Google Scholar] [CrossRef]
- Cagetti, M.G.; Mastroberardino, S.; Milia, E.; Cocco, F.; Lingström, P.; Campus, G. The Use of Probiotic Strains in Caries Prevention: A Systematic Review. Nutrients 2013, 5, 2530–2550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laleman, I.; Detailleur, V.; Slot, D.E.; Slomka, V.; Quirynen, M.; Teughels, W. Probiotics reduce mutans streptococci counts in humans: A systematic review and meta-analysis. Clin. Oral Investig. 2014, 18, 1539–1552. [Google Scholar] [CrossRef] [Green Version]
- Ferrer, M.D.; López-López, A.; Nicolescu, T.; Salavert, A.; Méndez, I.; Cuñé, J.; Llena, C.; Mira, A. A pilot study to assess oral colonization and pH buffering by the probiotic Streptococcus dentisani under different dosing regimes. Odontology 2020, 108, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Krasse, B. The Vipeholm Dental Caries Study: Recollections and Reflections 50 Years Later. J. Dent. Res. 2001, 80, 1785–1788. [Google Scholar] [CrossRef]
- Belstrøm, D. The salivary microbiota in health and disease. J. Oral Microbiol. 2020, 12, 1723975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynge Pedersen, A.M.; Belstrom, D. The role of natural salivary defences in maintaining a healthy oral microbiota. J. Dent. 2019, 80 (Suppl. 1), S3–S12. [Google Scholar] [CrossRef] [PubMed]
- Dzidic, M.; Collado, M.C.; Abrahamsson, T.; Artacho, A.; Stensson, M.; Jenmalm, M.C.; Mira, A. Oral microbiome development during childhood: An ecological succession influenced by postnatal factors and associated with tooth decay. ISME J. 2018, 12, 2292–2306. [Google Scholar] [CrossRef] [PubMed]
- Relvas, M.; Regueira-Iglesias, A.; Balsa-Castro, C.; Salazar, F.; Pacheco, J.J.; Cabral, C.; Henriques, V.; Tomás, I. Relationship between dental and perio-dontal health status and the salivary microbiome: Bacterial diversity, co-occurrence networks and predictive models. Sci. Rep. 2021, 11, 929. [Google Scholar] [CrossRef]
- Belstrøm, D.; Sembler-Møller, M.; Grande, M.; Kirkby, N.; Cotton, S.; Paster, B.; Twetman, S.; Holmstrup, P. Impact of Oral Hygiene Discontinuation on Supragingival and Salivary Microbiomes. JDR Clin. Transl. Res. 2017, 3, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Murugesan, S.; Al Ahmad, S.F.; Singh, P.; Saadaoui, M.; Kumar, M.; Al Khodor, S. Profiling the Salivary microbiome of the Qatari population. J. Transl. Med. 2020, 18, 1–16. [Google Scholar] [CrossRef]
- Hansen, T.H.; Kern, T.; Bak, E.G.; Kashani, A.; Allin, K.H.; Nielsen, T.; Hansen, T.; Pedersen, O. Impact of a vegan diet on the human salivary microbiota. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef]
- Moynihan, P.J.; Kelly, S.A. Effect on caries of restricting sugars intake: Systematic review to inform WHO guidelines. J. Dent. Res. 2014, 93, 8–18. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, N.; Nyvad, B. The role of bacteria in the caries process: Ecological perspectives. J. Dent. Res. 2011, 90, 294–303. [Google Scholar] [CrossRef]
- Keevil, C.W.; Marsh, P.D.; Ellwood, D.C. Regulation of glucose metabolism in oral streptococci through independent pathways of glucose 6-phosphate and glucose 1-phosphate formation. J. Bacteriol. 1984, 157, 560–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.; Qiao, Y.; Zhang, Y.; Leng, C.; Chen, H.; Sun, J.; Fan, X.; Li, A.; Feng, Z. Metabolic Profiles of Carbohydrates in Streptococcus thermophilus During pH-Controlled Batch Fermentation. Front. Microbiol. 2020, 11, 1131. [Google Scholar] [CrossRef]
- Lazarevic, V.; Manzano, S.; Gaïa, N.; Girard, M.; Whiteson, K.; Hibbs, J.; François, P.; Gervaix, A.; Schrenzel, J. Effects of amoxicillin treatment on the salivary microbiota in children with acute otitis media. Clin. Microbiol. Infect. 2013, 19, e335–e342. [Google Scholar] [CrossRef] [Green Version]
- Nyvad, B.; Takahashi, N. Integrated hypothesis of dental caries and periodontal diseases. J. Oral Microbiol. 2020, 12, 1710953. [Google Scholar] [CrossRef] [Green Version]
- Keller, M.K.; Brandsborg, E.; Holmstrøm, K.; Twetman, S. Effect of tablets containing probiotic candidate strains on gingival in-flammation and composition of the salivary microbiome: A randomised controlled trial. Benef. Microbes. 2018, 9, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Belstrøm, D.; Eiberg, J.M.; Enevold, C.; Grande, M.A.; Jensen, C.A.J.; Skov, L.; Hansen, P.R. Salivary microbiota and inflammation-related proteins in patients with psoriasis. Oral Dis. 2020, 26, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Baima, G.; Iaderosa, G.; Citterio, F.; Grossi, S.; Romano, F.; Berta, G.N.; Buduneli, N.; Aimetti, M. Salivary metabolomics for the diagnosis of periodontal diseases: A systematic review with methodological quality assessment. Metabolomics 2021, 17, 1–21. [Google Scholar] [CrossRef]
- Sharma, A.; Badea, M.; Tiwari, S.; Marty, J. Wearable Biosensors: An Alternative and Practical Approach in Healthcare and Disease Monitoring. Molecules 2021, 26, 748. [Google Scholar] [CrossRef]
- Belstrøm, D.; Grande, M.A.; Sembler-Møller, M.L.; Kirkby, N.; Cotton, S.L.; Paster, B.J.; Holmstrup, P. Influence of periodontal treatment on subgingival and salivary microbiotas. J. Periodontol. 2018, 89, 531–539. [Google Scholar] [CrossRef] [Green Version]
- Proctor, D.M.; Fukuyama, J.A.; Loomer, P.M.; Armitage, G.C.; Lee, S.A.; Davis, N.M.; Ryder, M.I.; Holmes, S.P.; Relman, D.A. A spatial gradient of bacterial diversity in the human oral cavity shaped by salivary flow. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belstrøm, D.; Paster, B.J.; Fiehn, N.-E.; Bardow, A.; Holmstrup, P. Salivary bacterial fingerprints of established oral disease revealed by the Human Oral Microbe Identification using Next Generation Sequencing (HOMINGS) technique. J. Oral Microbiol. 2016, 8, 30170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bardow, A.; Lykkeaa, J.; Qvist, V.; Ekstrand, K.; Twetman, S.; Fiehn, N.-E. Saliva composition in three selected groups with normal stimulated salivary flow rates, but yet major differences in caries experience and dental erosion. Acta Odontol. Scand. 2014, 72, 466–473. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escapa, I.F.; Chen, T.; Huang, Y.; Gajare, P.; Dewhirst, F.E.; Lemon, K.P. New Insights into Human Nostril Microbiome from the Expanded Human Oral Microbiome Database (eHOMD): A Resource for the Microbiome of the Human Aerodigestive Tract. mSystems 2018, 3, e00187-18. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C.; Flyvbjerg, H. Error filtering, pair assembly and error correction for next-generation sequencing reads. Bioinformatics 2015, 31, 3476–3482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. SINTAX: A simple non-Bayesian taxonomy classifier for 16S and ITS sequences. bioRxiv 2016, 074161. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [Green Version]
- Hochberg, Y.; Benjamini, Y. More powerful procedures for multiple significance testing. Stat. Med. 1990, 9, 811–818. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sucrose + Placebo (n = 20) | Sucrose + Probiotics (n = 20) | Xylitol + Probiotics (n = 20) | Xylitol + Placebo (n = 20) | |
|---|---|---|---|---|
| Gender, female/male | 16/4 | 16/4 | 14/6 | 13/7 |
| Age (mean, range) years | 24.2 (21–29) | 24.3 (20–32) | 24.6 (20–32) | 23.7 (20–29) |
| Dental professions * | 11/20 | 16/20 | 14/20 | 17/20 |
| NGAL | ||||
|---|---|---|---|---|
| Baseline | Week 2 | Week 5 | p-Value | |
| Sucrose + probiotics | 1422 (1189–2236) | 1780 (1271–2638) | 1249 (877–1986) | 0.13 |
| Sucrose + placebo | 1837 (1423–2677) | 2208 (775–2761) | 2462 (1211–4141) | 0.5 |
| Xylitol + probiotics | 1429 (1053–1757) | 1458 (815–2367) | 1061 (559–1908) | 0.31 |
| Xylitol + placebo | 1209 (825–2112) | 1709 (937–2553) | 1399 (914–3000) | 0.26 |
| Transferrin | ||||
|---|---|---|---|---|
| Baseline | Week 2 | Week 5 | p-Value | |
| Sucrose + probiotics | 4.4 (3.0–7.4) | 4.8 (3.5–6.1) | 3.8 (2.7–7.1) | 0.1 |
| Sucrose + placebo | 4.5 (3.8–7.5) | 5.5 (3.0–6.6) | 6.8 (3.9–12.7) | 0.04 |
| Xylitol + probiotics | 4.7 (3.4–7.2) | 3.8 (2.6–5.3) | 3.9 (2.5–5.0) | 0.05 |
| Xylitol + placebo | 3.7 (2.7–6.1) | 4.2 (2.9–6.0) | 4.2 (2.5–7.5) | 0.77 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lundtorp-Olsen, C.; Enevold, C.; Juel Jensen, C.A.; Stofberg, S.N.; Twetman, S.; Belstrøm, D. Impact of Probiotics on the Salivary Microbiota and Salivary Levels of Inflammation-Related Proteins during Short-Term Sugar Stress: A Randomized Controlled Trial. Pathogens 2021, 10, 392. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10040392
Lundtorp-Olsen C, Enevold C, Juel Jensen CA, Stofberg SN, Twetman S, Belstrøm D. Impact of Probiotics on the Salivary Microbiota and Salivary Levels of Inflammation-Related Proteins during Short-Term Sugar Stress: A Randomized Controlled Trial. Pathogens. 2021; 10(4):392. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10040392
Chicago/Turabian StyleLundtorp-Olsen, Christine, Christian Enevold, Claus Antonio Juel Jensen, Steen Nymann Stofberg, Svante Twetman, and Daniel Belstrøm. 2021. "Impact of Probiotics on the Salivary Microbiota and Salivary Levels of Inflammation-Related Proteins during Short-Term Sugar Stress: A Randomized Controlled Trial" Pathogens 10, no. 4: 392. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10040392