
Detection of Tick-Borne Pathogens of the Genera Rickettsia, Anaplasma and Francisella in Ixodes ricinus Ticks in Pomerania (Poland)


Abstract
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
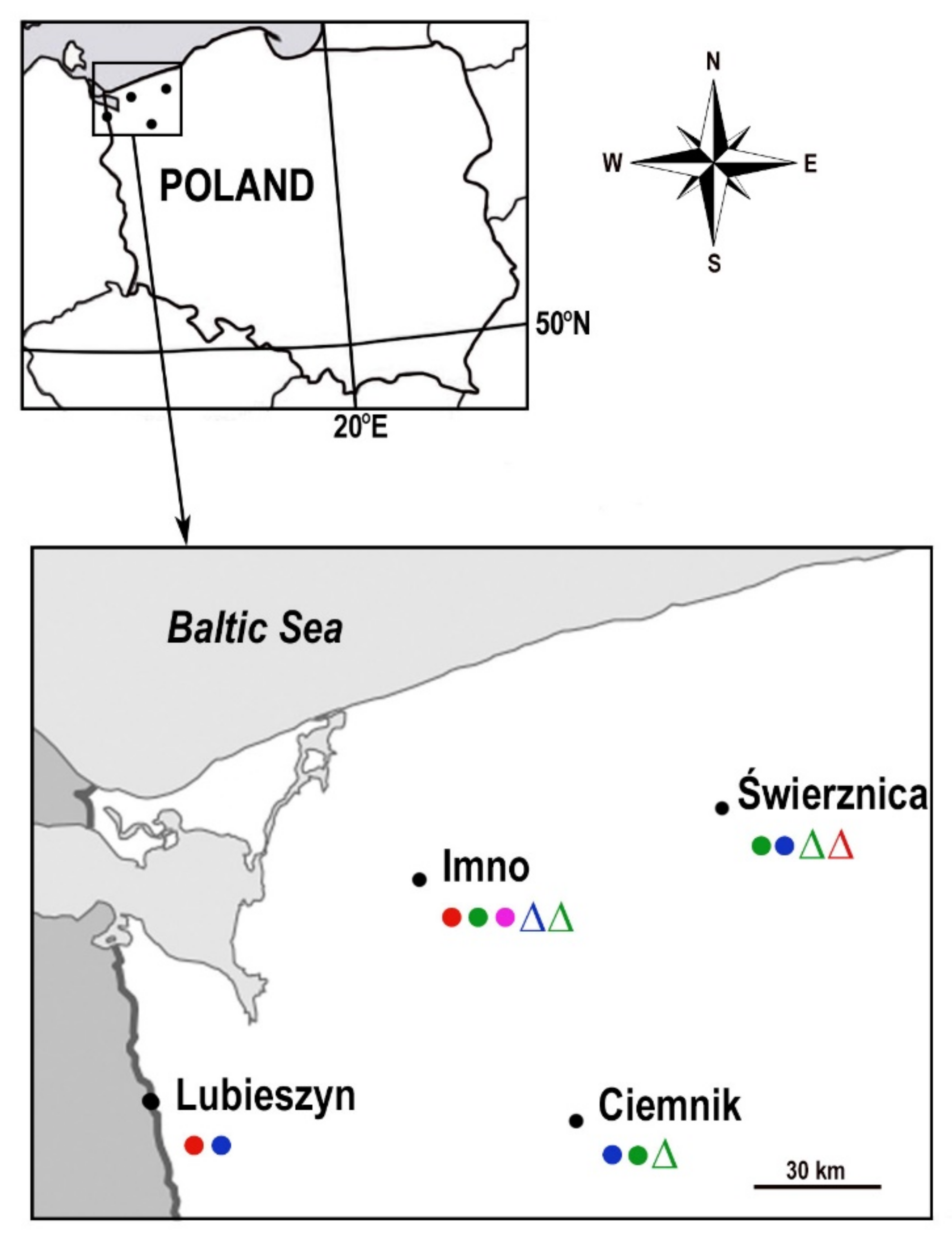
4.1. Characteristics of the Collection Sites
4.2. Sample Preparation and Molecular Analysis
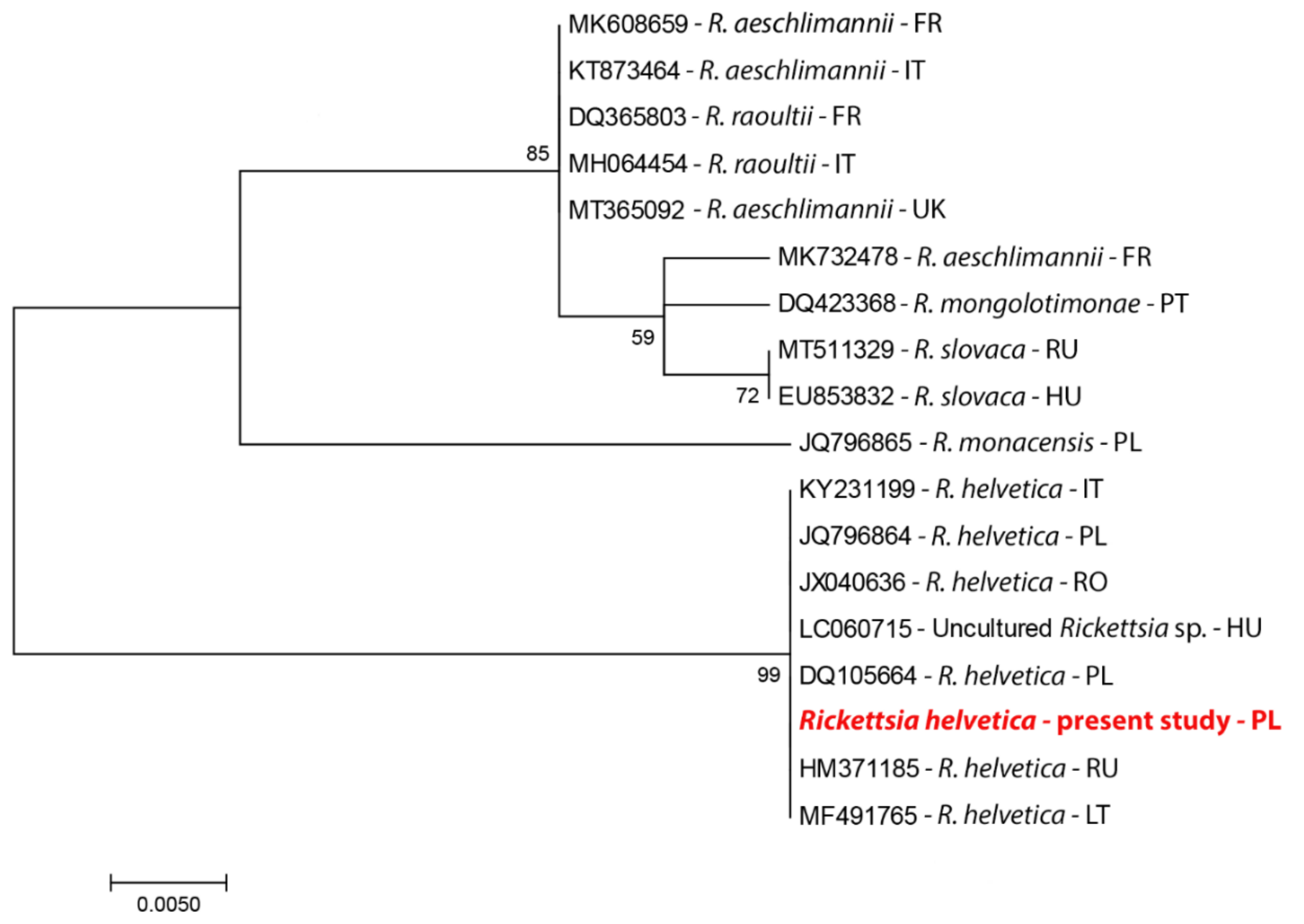
4.3. DNA Sequencing and Phylogenetic Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dumler, J.S.; Barbet, A.F.; Bekker, C.P.; Dasch, G.A.; Palmer, G.H.; Ray, S.C.; Rikihisa, Y.; Rurangirwa, F.R. Reorganization of genera in the families Rickettsiaceae and Anaplasmataceae in the order Rickettsiales: Unification of some species of Ehrlichia with Anaplasma, Cowdria with Ehrlichia and Ehrlichia with Neorickettsia, descriptions of six new species combinations and designation of Ehrlichia equi and ‘HGE agent’ as subjective synonyms of Ehrlichia phagocytophila. Int. J. Syst. Evol. Microbiol. 2001, 51, 2145–2165. [Google Scholar] [CrossRef] [Green Version]
- Rikihisa, Y. Mechanisms of obligatory intracellular infection with Anaplasma phagocytophilum. Clin. Microbiol Rev. 2011, 24, 469–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, W.; Bocklisch, H.; Schüler, G.; Hotzel, H.; Neubauer, H.; Otto, P. Detection of Francisella tularensis subsp. holarctica in a European brown hare (Lepus europaeus) in Thuringia, Germany. Vet. Microbiol. 2007, 123, 225–229. [Google Scholar] [CrossRef]
- Franke, J.; Fritzsch, J.; Tomaso, H.; Straube, E.; Dorn, W.; Hildebrandt, A. Coexistence of pathogens in host-seeking and feeding ticks within a single natural habitat in Central Germany. Appl. Environ. Microbiol. 2010, 76, 6829–6836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hestvik, G.; Warns-Petit, E.; Smith, L.A.; Fox, N.J.; Uhlhorn, H.; Artois, M.; Hannant, D.; Hutchings, M.R.; Mattsson, R.; Yon, L.; et al. The status of tularemia in Europe in a one-health context: A review. Epidemiol. Infect. 2015, 143, 2137–2160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamska, M. The role of different species of wild ungulates and Ixodes ricinus ticks in the circulation of genetic variants of Anaplasma phagocytophilum in a forest biotope in north-western Poland. Ticks Tick Borne Dis. 2020, 11, 101–465. [Google Scholar] [CrossRef] [PubMed]
- Piotrowski, M.; Rymaszewska, A. Expansion of tick-borne rickettsioses in the world. Microorganisms 2020, 8, 1906. [Google Scholar] [CrossRef] [PubMed]
- Kernif, T.; Leulmi, H.; Raoult, D.; Parola, P. Emerging tick-borne bacterial pathogens. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef]
- Londoño, A.F.; Acevedo-Gutiérrez, L.Y.; Marín, D.; Contreras, V.; Díaz, F.J.; Valbuena, G.; Labruna, M.B.; Hidalgo, M.; Arboleda, M.; Mattar, S.; et al. Human prevalence of the spotted fever group (SFG) rickettsiae in endemic zones of Northwestern Colombia. Ticks Tick Borne Dis. 2017, 8, 477–482. [Google Scholar] [CrossRef]
- Gillespie, J.J.; Williams, K.; Shukla, M.; Snyder, E.E.; Nordberg, E.K.; Ceraul, S.M.; Dharmanolla, C.; Rainey, D.; Soneja, J.; Shallom, J.M.; et al. Rickettsia phylogenomics: Unwinding the intricacies of obligate intracellular life. PLoS ONE 2008, 3, e2018. [Google Scholar] [CrossRef] [Green Version]
- Otto, P.; Kohlmann, R.; Müller, W.; Julich, S.; Geis, G.; Gatermann, S.G.; Peters, M.; Wolf, P.J.; Karlsson, E.; Forsman, M.; et al. Hare-to-human transmission of Francisella tularensis subsp. holarctica, Germany. Emerg. Infect. Dis. 2015, 21, 153–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seiwald, S.; Simeon, A.; Hofer, E.; Weiss, G.; Bellmann-Weiler, R. Tularemia Goes West: Epidemiology of an emerging infection in Austria. Microorganisms 2020, 8, 1597. [Google Scholar] [CrossRef]
- Yanushevych, M.; Komorowska-Piotrowska, A.; Feleszko, W. Tularaemia—A forgotten disease? Own experience. Med. Wieku Rozwoj 2013, 17, 355–359. [Google Scholar]
- Zasada, A.A.; Gierczyński, R.; Rzeczkowska, M.; Formińska, K.; Zacharczuk, K.; Rastawicki, W. Detection and identification of highly pathogenic bacteria within the framework of the EQADeBa project—Part I: Samples containing living pathogens. Przegl. Epidemiol. 2011, 65, 401–407. [Google Scholar] [PubMed]
- Matei, I.A.; Estrada-Peña, A.; Cutler, S.J.; Vayssier-Taussat, M.; Varela-Castro, L.; Potkonjak, A.; Zeller, H.; Mihalca, A.D. A review on the eco-epidemiology and clinical management of human granulocytic anaplasmosis and its agent in Europe. Parasites Vectors 2019, 12, 599. [Google Scholar] [CrossRef]
- Mączka, I.; Roguska, U.; Tylewska-Wierzbanowska, S. Occurrence of rickettsial infection in Poland in 2006–2012. Przegl. Epidemiol. 2013, 67, 721–723. [Google Scholar]
- Rymaszewska, A. Variability within the msp2 gene in populations of Anaplasma phagocythopilum. Folia Biol. 2010, 56, 269–275. [Google Scholar]
- Nilsson, K.; Lindquist, O.; Liu, A.J.; Jaenson, T.G.; Friman, G.; Påhlson, C. Rickettsia helvetica in Ixodes ricinus ticks in Sweden. J. Clin. Microbiol. 1999, 37, 400–403. [Google Scholar] [CrossRef] [Green Version]
- Wójcik-Fatla, A.; Zając, V.; Sawczyn, A.; Cisak, E.; Sroka, J.; Dutkiewicz, J. Occurrence of Francisella spp. in Dermacentor reticulatus and Ixodes ricinus ticks collected in eastern Poland. Ticks Tick Borne Dis. 2015, 6, 253–257. [Google Scholar] [CrossRef]
- Reye, A.L.; Hübschen, J.M.; Sausy, A.; Muller, C.P. Prevalence and seasonality of tick-borne pathogens in questing Ixodes ricinus ticks from Luxembourg. Appl. Environ. Microbiol. 2010, 76, 2923–2931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egyed, L.; Elő, P.; Sréter-Lancz, Z.; Széll, Z.; Balogh, Z.; Sréter, T. Seasonal activity and tick-borne pathogen infection rates of Ixodes ricinus ticks in Hungary. Ticks Tick Borne Dis. 2012, 3, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, S.; de la Fuente, J.; Nicollet, P.; Liu, X.; Madani, N.; Blanchard, B.; Maingourd, C.; Alongi, A.; Torina, A.; Fernández de Mera, I.G.; et al. Prevalence of tick-borne pathogens in adult Dermacentor spp. ticks from nine collection sites in France. Vector Borne Zoonot Dis. 2013, 13, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Kreizinger, Z.; Hornok, S.; Dán, A.; Hresko, S.; Makrai, L.; Magyar, T.; Bhide, M.; Erdélyi, K.; Hofmann-Lehmann, R.; Gyuranecz, M. Prevalence of Francisella tularensis and Francisella-like endosymbionts in the tick population of Hungary and the genetic variability of Francisella-like agents. Vector Borne Zoonot. Dis. 2013, 13, 160–163. [Google Scholar] [CrossRef] [Green Version]
- Milutinović, M.; Masuzawa, T.; Tomanović, S.; Radulović, Z.; Fukui, T.; Okamoto, Y. Borrelia burgdorferi sensu lato, Anaplasma phagocytophilum, Francisella tularensis and their co-infections in host-seeking Ixodes ricinus ticks collected in Serbia. Exp. Appl. Acarol. 2008, 45, 171–183. [Google Scholar] [CrossRef]
- Gehringer, H.; Schacht, E.; Maylaender, N.; Zeman, E.; Kaysser, P.; Oehme, R.; Pluta, S.; Splettstoesser, W.D. Presence of an emerging subclone of Francisella tularensis holarctica in Ixodes ricinus ticks from south-western Germany. Ticks Tick Borne Dis. 2013, 4, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Michelet, L.; Bonnet, S.; Madani, N.; Moutailler, S. Discriminating Francisella tularensis and Francisella-like endosymbionts in Dermacentor reticulatus ticks: Evaluation of current molecular techniques. Vet. Microbiol. 2013, 163, 399–403. [Google Scholar] [CrossRef] [PubMed]
- Laaksonen, M.; Klemola, T.; Feuth, E.; Sormunen, J.J.; Puisto, A.; Mäkelä, S.; Penttinen, R.; Ruohomäki, K.; Hänninen, J.; Sääksjärvi, I.E.; et al. Tick-borne pathogens in Finland: Comparison of Ixodes ricinus and I. persulcatus in sympatric and parapatric areas. Parasite Vector 2018, 11, 556. [Google Scholar] [CrossRef] [Green Version]
- Rymaszewska, A.; Piotrowski, M. Use of DNA sequences for Rickettsia identification in Ixodes ricinus ticks: The first detection of Rickettsia monacensis in Poland. Microbes Infect. 2013, 15, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Kowalec, M.; Szewczyk, T.; Welc-Falęciak, R.; Siński, E.; Karbowiak, G.; Bajer, A. Rickettsiales occurrence and co-occurrence in Ixodes ricinus ticks in natural and urban areas. Microb. Ecol. 2019, 77, 890–904. [Google Scholar] [CrossRef] [Green Version]
- Michalski, M.M.; Kubiak, K.; Szczotko, M.; Dmitryjuk, M. Tick-borne pathogens in ticks collected from wild ungulates in North-Eastern Poland. Pathogens 2021, 10, 587. [Google Scholar] [CrossRef]
- Žákovská, A.; Bártová, E.; Pittermannová, P.; Budíková, M. Antibodies related to Borrelia burgdorferi sensu lato, Coxiella burnetii, and Francisella tularensis detected in serum and heart rinses of wild small mammals in the Czech Republic. Pathogens 2021, 10, 419. [Google Scholar] [CrossRef]
- Wächter, M.; Wölfel, S.; Pfeffer, M.; Dobler, G.; Kohn, B.; Moritz, A.; Pachnicke, S.; Silaghi, C. Serological differentiation of antibodies against Rickettsia helvetica, R. raoultii, R. slovaca, R. monacensis and R. felis in dogs from Germany by a micro-immunofluorescent antibody test. Parasite Vectors 2015, 23, 126. [Google Scholar] [CrossRef] [Green Version]
- Jansen, A.; La Scola, B.; Raoult, D.; Lierz, M.; Wichmann, O.; Stark, K.; Schneider, T. Antibodies against Rickettsia spp. in hunters, Germany. Emerg. Infect. Dis. 2008, 14, 1961–1963. [Google Scholar] [CrossRef]
- Nowak-Chmura, M. The Fauna of Ticks (Ixodida) of Central Europe; Wydawnictwo Naukowe Uniwersytetu Pedagogicznego: Kraków, Poland, 2013. [Google Scholar]
- Wodecka, B.; Rymaszewska, A.; Skotarczak, B. Host and pathogen DNA identification in blood meals of nymphal Ixodes ricinus ticks from forest parks and rural forests of Poland. Exp. Appl. Acarol. 2014, 62, 543–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levin, M.L.; Nicholson, W.L.; Massung, R.F.; Sumner, J.W.; Fish, D. Comparison of the reservoir competence of medium-sized mammals and Peromyscus leucopus for Anaplasma phagocytophilum in Connecticut. Vector Borne Zoonotic Dis. 2002, 2, 125–136. [Google Scholar] [CrossRef]
- Sjöstedt, A.; Eriksson, U.; Berglund, L.; Tärnvik, A. Detection of Francisella tularensis in ulcers of patients with tularemia by PCR. J. Clin. Microbiol. 1997, 35, 1045–1048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labruna, M.B.; Pacheco, R.C.; Nava, S.; Brandão, P.E.; Richtzenhain, L.J.; Guglielmone, A.A. Infection by Rickettsia bellii and Candidatus “Rickettsia amblyommii” in Amblyomma neumanni ticks from Argentina. Microb. Ecol. 2007, 54, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Tamura, K. Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G+C-content biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Site | Number of Ticks | Female n/% | Male n/% | Nymph n/% | Larvae n/% |
|---|---|---|---|---|---|
| Świerznica 21/4.42 | 475 | 14/2.9 | 15/3.16 | 252/53.05 | 194/40.84 |
| Ciemnik 15/2.86 | 524 | 35/6.68 | 29/5.53 | 276/52.67 | 184/35.11 |
| Imno 24/5.71 | 420 | 55/13.09 | 10/2.38 | 252/60 | 103/24.52 |
| Lubieszyn 2/0.76 | 263 | 7/2.66 | 14/5.32 | 228/86.69 | 14/5.32 |
| Total | 1682 | 111/6.60 | 68/4.04 | 1008/59.93 | 495/29.43 |
| Number of Ticks | Site/Infection | Female (n/%)●∆ | Male (n/%)●∆ | Nymph (n/%)●∆ | Larvae (n/%)●∆ | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Ric n/%PCR+ | Aph n/%PCR+ | Ric n/%PCR+ | Aph n/%PCR+ | Ric n/%PCR+ | Aph n/%PCR+ | Ric n/%PCR+ | Aph n/%PCR+ | Ric n/%PCR+ | Aph n/%PCR+ | |
| 457 | Świerznica | 0 | 1/7.14 | 3/20.0 | 0 | 18/7.14 | 3/1.19 | 0 | nt | |
| 21/4.42 | 4/0.84 | |||||||||
| 524 | Ciemnik | 0 | 0 | 4/13.7 | 0 | 11/3.99 | 1/0.36 | 0 | nt | |
| 15/2.86 | 1/0.19 | |||||||||
| 420 | Imno | 1/1.82 | 0 | 0 | 2/20.0 | 22/8.73 | 2 /0.79 | 1/0.97 | nt | |
| 24/5.71 | 4/0.95 | |||||||||
| 263 | Lubieszyn | 1/14.29 | 0 | 1/7.14 | 0 | 0 | 0 | 0 | nt | |
| 2/0.76 | 0/0.84 | |||||||||
| 1682 | Total | 2/1.80 | 1/0.90 | 8/11.76 | 2/2.94 | 51/1.80 | 6/0.90 | 1/0.2 | nt | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kirczuk, L.; Piotrowski, M.; Rymaszewska, A. Detection of Tick-Borne Pathogens of the Genera Rickettsia, Anaplasma and Francisella in Ixodes ricinus Ticks in Pomerania (Poland). Pathogens 2021, 10, 901. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10070901
Kirczuk L, Piotrowski M, Rymaszewska A. Detection of Tick-Borne Pathogens of the Genera Rickettsia, Anaplasma and Francisella in Ixodes ricinus Ticks in Pomerania (Poland). Pathogens. 2021; 10(7):901. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10070901
Chicago/Turabian StyleKirczuk, Lucyna, Mariusz Piotrowski, and Anna Rymaszewska. 2021. "Detection of Tick-Borne Pathogens of the Genera Rickettsia, Anaplasma and Francisella in Ixodes ricinus Ticks in Pomerania (Poland)" Pathogens 10, no. 7: 901. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10070901